
Crystal Structure of a Tetrameric Type II β-Carbonic Anhydrase from the Pathogenic Bacterium Burkholderia pseudomallei

 , ,
, ,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
3. Materials and Methods
3.1. Enzyme Preparation
3.2. Crystallization and Data Collection
3.3. Structure Determination and Refinement
3.4. Kinetic and Inhibition Assay
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Limmathurotsakul, D.; Peacock, S.J. Melioidosis: A clinical overview. Br. Med. Bull. 2011, 99, 125–139. [Google Scholar] [CrossRef]
- Limmathurotsakul, D.; Golding, N.; Dance, D.; Messina, J.P.; Pigott, D.M.; Moyes, C.; Rolim, D.B.; Bertherat, E.; Day, N.P.; Peacock, S.J.; et al. Predicted global distribution of Burkholderia pseudomallei and burden of melioidosis. Nat. Microbiol. 2016, 1, 15008. [Google Scholar] [CrossRef] [Green Version]
- Wiersinga, W.J.; van der Poll, T.; White, N.J.; Day, N.P.; Peacock, S.J. Melioidosis: Insights into the pathogenicity of Burkholderia pseudomallei. Nat. Rev. Genet. 2006, 4, 272–282. [Google Scholar] [CrossRef]
- Cheng, A.; Currie, B.J. Melioidosis: Epidemiology, pathophysiology, and management. Clin. Microbiol. Rev. 2005, 18, 383–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotz, L.D.; Khan, A.S.; Lillibridge, S.R.; Ostroff, S.M.; Hughes, J.M. Public health assessment of potential biological terrorism agents. Emerg. Infect. Dis. 2002, 8, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Balaban, N.Q.; Gerdes, K.; Lewis, K.; McKinney, J.D. A problem of persistence: Still more questions than answers? Nat. Rev. Genet. 2013, 11, 587–591. [Google Scholar] [CrossRef] [PubMed]
- Monroe, D. Looking for chinks in the armor of bacterial biofilms. PLoS Biol. 2007, 5, e307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Supuran, C.T.; Capasso, C. An overview of the bacterial carbonic anhydrases. Metabolites 2017, 7, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Supuran, C.T.; Capasso, C. Biomedical applications of prokaryotic carbonic anhydrases. Expert Opin. Ther. Pat. 2018, 28, 745–754. [Google Scholar] [CrossRef]
- Capasso, C.; Supuran, C.T. Inhibition of bacterial carbonic anhydrases as a novel approach to escape drug resistance. Curr. Top Med. Chem. 2017, 17, 1237–1248. [Google Scholar] [CrossRef] [Green Version]
- Capasso, C.; Supuran, C.T. Bacterial, fungal and protozoan carbonic anhydrases as drug targets. Expert Opin. Ther. Targets 2015, 19, 1689–1704. [Google Scholar] [CrossRef] [PubMed]
- Capasso, C.; Supuran, C.T. An overview of the alpha-, beta- and gamma-carbonic anhydrases from Bacteria: Can bacterial carbonic anhydrases shed new light on evolution of bacteria? J. Enzyme Inhib. Med. Chem. 2015, 30, 325–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Supuran, C.T. Structure-based drug discovery of carbonic anhydrase inhibitors. J. Enzyme Inhib. Med. Chem. 2012, 27, 759–772. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T. Carbonic anhydrase inhibitors and activators for novel therapeutic applications. Future Med. Chem. 2011, 3, 1165–1180. [Google Scholar] [CrossRef]
- Bozdag, M.; Altamimi, A.S.A.; Vullo, D.; Supuran, C.T.; Carta, F. State of the art on carbonic anhydrase modulators for biomedical purposes. Curr. Med. Chem. 2019, 26, 2558–2573. [Google Scholar] [CrossRef]
- Supuran, C.T. Structure and function of carbonic anhydrases. Biochem. J. 2016, 473, 2023–2032. [Google Scholar] [CrossRef]
- Akocak, S.; Supuran, C.T. Activation of α-, β-, γ- δ-, ζ- and η- class of carbonic anhydrases with amines and amino acids: A review. J. Enzyme Inhib. Med. Chem. 2019, 34, 1652–1659. [Google Scholar] [CrossRef] [Green Version]
- Del Prete, S.; Nocentini, A.; Supuran, C.T.; Capasso, C. Bacterial ι-carbonic anhydrase: A new active class of carbonic anhydrase identified in the genome of the Gram-negative bacterium Burkholderia territorii. J. Enzyme Inhib. Med. Chem. 2020, 35, 1060–1068. [Google Scholar] [CrossRef] [Green Version]
- Capasso, C.; Supuran, C.T. Anti-infective carbonic anhydrase inhibitors: A patent and literature review. Expert Opin. Ther. Pat. 2013, 23, 693–704. [Google Scholar] [CrossRef]
- Supuran, C.T. Inhibition of bacterial carbonic anhydrases and zinc proteases: From orphan targets to innovative new antibiotic drugs. Curr. Med. Chem. 2012, 19, 831–844. [Google Scholar] [CrossRef]
- Del Prete, S.; Vullo, D.; Di Fonzo, P.; Osman, S.M.; Alothman, Z.; Donald, W.A.; Supuran, C.T.; Capasso, C. Sulfonamide inhibition profile of the γ-carbonic anhydrase identified in the genome of the pathogenic bacterium Burkholderia pseudomallei the etiological agent responsible of melioidosis. Bioorg. Med. Chem. Lett. 2017, 27, 490–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vullo, D.; Del Prete, S.; Di Fonzo, P.; Carginale, V.; Donald, W.A.; Supuran, C.T.; Capasso, C. Comparison of the sulfonamide inhibition profiles of the β- and γ-carbonic anhydrases from the pathogenic bacterium Burkholderia pseudomallei. Molecules 2017, 22, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefanucci, A.; Angeli, A.; Dimmito, M.P.; Luisi, G.; Del Prete, S.; Capasso, C.; Donald, W.A.; Mollica, A.; Supuran, C.T. Activation of β- and γ-carbonic anhydrases from pathogenic bacteria with tripeptides. J. Enzyme Inhib. Med. Chem. 2018, 33, 945–950. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, S.; Vullo, D.; De Luca, V.; Carginale, V.; Osman, S.M.; Alothman, Z.; Supuran, C.T.; Capasso, C. Comparison of the sulfonamide inhibition profiles of the α-, β- and γ-carbonic anhydrases from the pathogenic bacterium Vibrio cholerae. Bioorg. Med. Chem. Lett. 2016, 26, 1941–1946. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, S.; Vullo, D.; Osman, S.M.; Alothman, Z.; Supuran, C.T.; Capasso, C. Sulfonamide inhibition profiles of the β-carbonic anhydrase from the pathogenic bacterium Francisella tularensis responsible of the febrile illness tularemia. Bioorg. Med. Chem. 2017, 25, 3555–3561. [Google Scholar] [CrossRef] [PubMed]
- Dedeoglu, N.; De Luca, V.; Isik, S.; Yildirim, H.; Köçkar, F.; Capasso, C.; Supuran, C.T. Cloning, characterization and anion inhibition study of a β-class carbonic anhydrase from the caries producing pathogen Streptococcus mutans. Bioorg. Med. Chem. 2015, 23, 2995–3001. [Google Scholar] [CrossRef] [PubMed]
- Shahidzadeh, R.; Opekun, A.; Shiotani, A.; Graham, D.Y. Effect of the carbonic anhydrase inhibitor, acetazolamide, on Helicobacter pylori infection in vivo: A pilot study. Helicobacter 2005, 10, 136–138. [Google Scholar] [CrossRef]
- Covarrubias, A.S.; Bergfors, T.; Jones, T.A.; Högbom, M. Structural mechanics of the pH-dependent activity of beta-carbonic anhydrase from Mycobacterium tuberculosis. J. Biol. Chem. 2006, 281, 4993–4999. [Google Scholar] [CrossRef] [Green Version]
- Kimber, M.S.; Pai, E.F. The active site architecture of Pisum sativum beta-carbonic anhydrase is a mirror image of that of alpha-carbonic anhydrases. EMBO J. 2000, 19, 1407–1418. [Google Scholar] [CrossRef] [Green Version]
- Strop, P.; Smith, K.S.; Iverson, T.M.; Ferry, J.G.; Rees, D.C. Crystal structure of the “cab”-type β class carbonic anhydrase from the archaeon Methanobacterium thermoautotrophicum. J. Biol. Chem. 2000, 276, 10299–10305. [Google Scholar] [CrossRef] [Green Version]
- Cronk, J.D.; Rowlett, R.; Zhang, K.Y.; Tu, C.; Endrizzi, J.A.; Lee, J.; Gareiss, P.C.; Preiss, J.R. Identification of a novel noncatalytic bicarbonate binding site in eubacterial β-carbonic anhydrase. Biochemistry 2006, 45, 4351–4361. [Google Scholar] [CrossRef]
- Hoffmann, K.M.; Million-Perez, H.R.; Merkhofer, R.; Nicholson, H.E.; Rowlett, R. Allosteric reversion of Haemophilus influenza β-Carbonic anhydrase via a proline shift. Biochemistry 2014, 54, 598–611. [Google Scholar] [CrossRef] [PubMed]
- Cronk, J.D.; Endrizzi, J.A.; Cronk, M.R.; O’Neill, J.W.; Zhang, K.Y. Crystal structure of E. coli β-carbonic anhydrase, an enzyme with an unusual pH-dependent activity. Protein Sci. 2001, 10, 911–922. [Google Scholar] [CrossRef] [PubMed]
- Mitsuhashi, S.; Mizushima, T.; Yamashita, E.; Yamamoto, M.; Kumasaka, T.; Moriyama, H.; Ueki, T.; Miyachi, S.; Tsukihara, T. X-ray structure of beta-carbonic anhydrase from the red alga, Porphyridium purpureum, reveals a novel catalytic site for CO(2) hydration. J. Biol. Chem. 2000, 275, 5521–5526. [Google Scholar] [CrossRef] [Green Version]
- Vagin, A.; Teplyakov, A. Molecular replacement with MOLREP. Acta Crystallogr Sect. D Biol Crystallogr. 2010, 66, 22–25. [Google Scholar] [CrossRef] [PubMed]
- Pinard, M.A.; Lotlikar, S.R.; Boone, C.D.; Vullo, D.; Supuran, C.T.; Patrauchan, M.A.; McKenna, R. Structure and inhibition studies of a type II beta-carbonic anhydrase psCA3 from Pseudomonas aeruginosa. Bioorg. Med. Chem. 2015, 23, 4831–4838. [Google Scholar] [CrossRef]
- Kabsch, W. XDS. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Murshudov, G.N.; Skubak, P.; Lebedev, A.A.; Pannu, N.S.; Steiner, R.A.; Nicholls, R.; Winn, M.D.; Long, F.; Vagin, A.A. REFMAC5 for the refinement of macromolecular crystal structures. Acta Crystallogr. Sect. D Biol. Crystallogr. 2011, 67, 355–367. [Google Scholar] [CrossRef] [Green Version]
- Hough, M.A.; Wilson, K. From crystal to structure with CCP4. Acta Crystallogr. Sect. D Struct. Biol. 2018, 74, 67. [Google Scholar] [CrossRef] [Green Version]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [Green Version]
- Lamzin, V.S.; Perrakis, A.; Wilson, K.S. The ARP/wARP suite for automated construction and refinement of protein models. In International Tables for Crystallography: Volume F; Rossmann, M.G., Arnold, E., Eds.; Crystallography of Biological Macromolecules, Kluwer Academic Publishers: Dordrecht, The Netherlands, 2001; pp. 720–722. [Google Scholar]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalifah, R.G. The carbon dioxide hydration activity of carbonic anhydrase. I. Stop-flow kinetic studies on the native human isoenzymes B and C. J. Biol. Chem. 1971, 246, 2561. [Google Scholar] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |




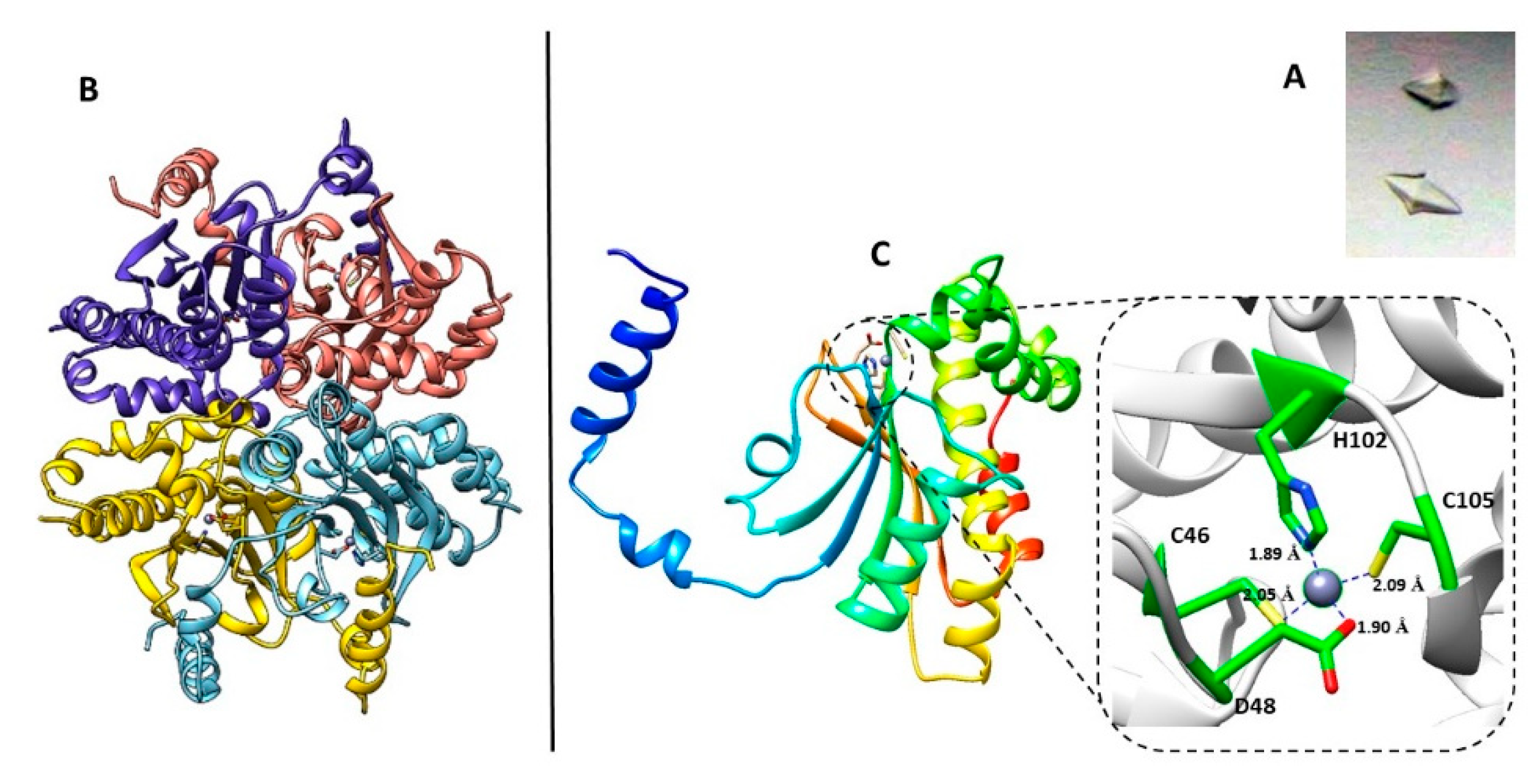{kind=link}
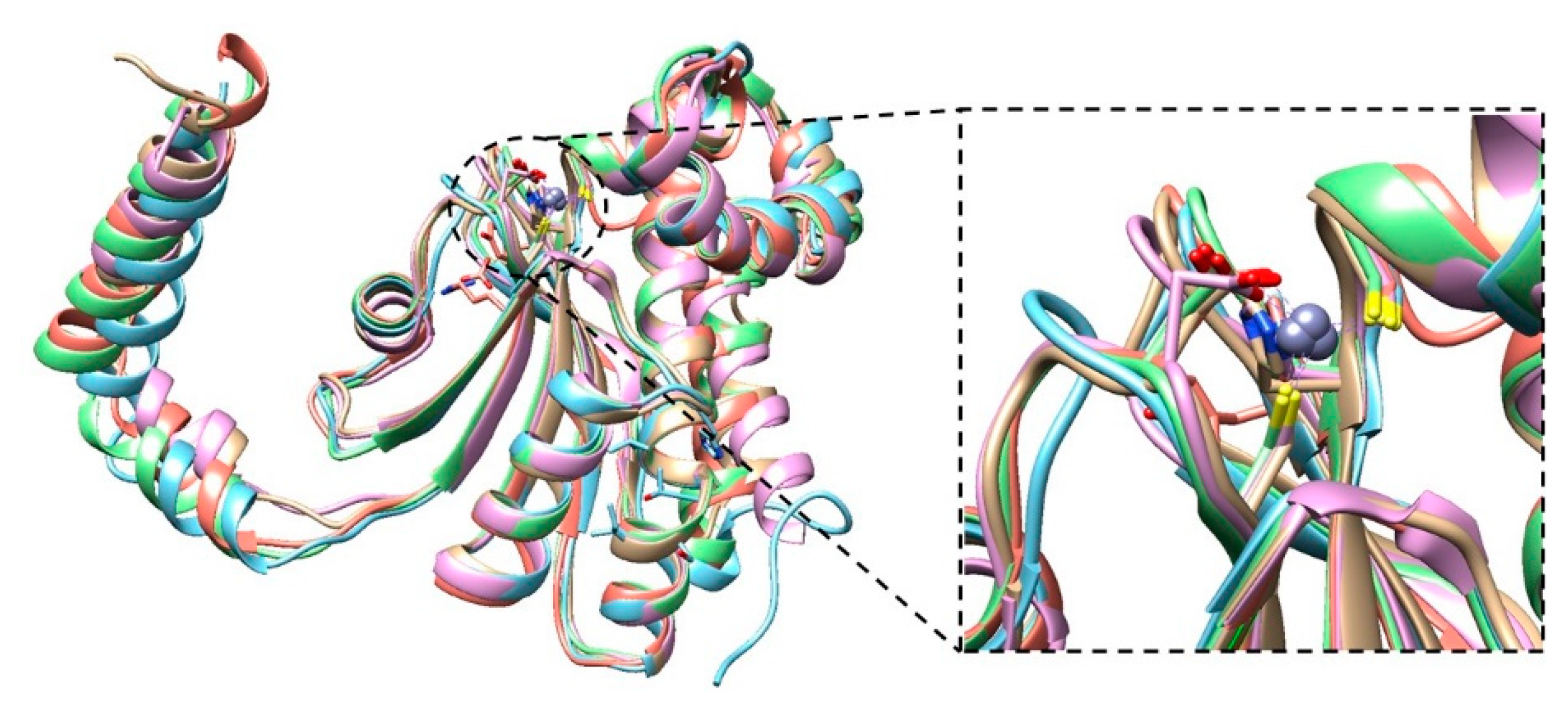{kind=link}
{kind=link}
| Enzyme | Activity Level | Class | kcat (s−1) | kcat/Km (M−1 s−1) | Ki (Acetazolamide) (nM) |
|---|---|---|---|---|---|
| BpsβCA | Moderate | β | 1.6 × 105 | 3.4 × 107 | 745 |
| BpsγCA | Moderate | γ | 5.3 × 105 | 2.5 × 107 | 149 |
| BteCAι | Moderate | ι | 3.0 × 105 | 9.7 × 107 | 64.9 |
| BpsβCA pH 7.5 | BpsβCA pH 8.5 | |
|---|---|---|
| PDB ID | 6YL7 | 6YJN |
| Wavelength (Å) | 1.0399 | 1.0000 |
| Space Group | P6422 | P6422 |
| Unit cell (a, b, c, α, β, γ) (Å,°) | 88.74;88.74;112.43; | 88.03;88.03;111.64; |
| 90.0;90.0;120.0 | 90.00;90.00;120.00 | |
| Limiting resolution (Å) | 45.37–3.16 (3.38–3.16) | 45.04–2.70 (2.83–2.70) |
| Unique reflections | 4858 (743) | 13330 (2141) |
| Rmerge (%) | 27.4 (265.4) | 26.0 (789.6) |
| Rmeas (%) | 28.08 (271.5) | 26.7 (811.5) |
| Redundancy | 24.7 (23.4) | 18.4 (18.6) |
| Completeness overall (%) | 99.7 (98.7) | 99.9 (99.6) |
| <I/σ(I)> | 10.42 (1.15) | 10.93 (0.34) |
| CC (1/2) | 99.8 (59.4) | 99.9 (32.1) |
| Refinement statistics | ||
| Resolution range (Å) | 45.412–3.166 | 45.080–2.701 |
| Unique reflections, working\free | 4605\3440 | 7508\7093 |
| Rfactor (%) | 19.26 | 21.4 |
| Rfree(%) | 29.62 | 32.4 |
| r.m.s.d. bonds(Å) | 0.0060 | 0.0052 |
| r.m.s.d. angles (°) | 1.5874 | 1.4987 |
| Ramachandran statistics (%) | ||
| Most favored | 79.3 | 82.9 |
| Additionally allowed | 17.8 | 11.4 |
| Outlier regions | 2.9 | 5.7 |
| Average B factor (Å2) | ||
| Solvent | 78.811 | 83.801 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angeli, A.; Ferraroni, M.; Pinteala, M.; Maier, S.S.; Simionescu, B.C.; Carta, F.; Del Prete, S.; Capasso, C.; Supuran, C.T. Crystal Structure of a Tetrameric Type II β-Carbonic Anhydrase from the Pathogenic Bacterium Burkholderia pseudomallei. Molecules 2020, 25, 2269. https://doi.org/10.3390/molecules25102269
Angeli A, Ferraroni M, Pinteala M, Maier SS, Simionescu BC, Carta F, Del Prete S, Capasso C, Supuran CT. Crystal Structure of a Tetrameric Type II β-Carbonic Anhydrase from the Pathogenic Bacterium Burkholderia pseudomallei. Molecules. 2020; 25(10):2269. https://doi.org/10.3390/molecules25102269
Chicago/Turabian StyleAngeli, Andrea, Marta Ferraroni, Mariana Pinteala, Stelian S. Maier, Bogdan C. Simionescu, Fabrizio Carta, Sonia Del Prete, Clemente Capasso, and Claudiu T. Supuran. 2020. "Crystal Structure of a Tetrameric Type II β-Carbonic Anhydrase from the Pathogenic Bacterium Burkholderia pseudomallei" Molecules 25, no. 10: 2269. https://doi.org/10.3390/molecules25102269
APA StyleAngeli, A., Ferraroni, M., Pinteala, M., Maier, S. S., Simionescu, B. C., Carta, F., Del Prete, S., Capasso, C., & Supuran, C. T. (2020). Crystal Structure of a Tetrameric Type II β-Carbonic Anhydrase from the Pathogenic Bacterium Burkholderia pseudomallei. Molecules, 25(10), 2269. https://doi.org/10.3390/molecules25102269