Green Tea Seed Isolated Theasaponin E1 Ameliorates AD Promoting Neurotoxic Pathogenesis by Attenuating Aβ Peptide Levels in SweAPP N2a Cells

Abstract
:
1. Introduction
2. Results
2.1. Saponin Extraction and Analysis
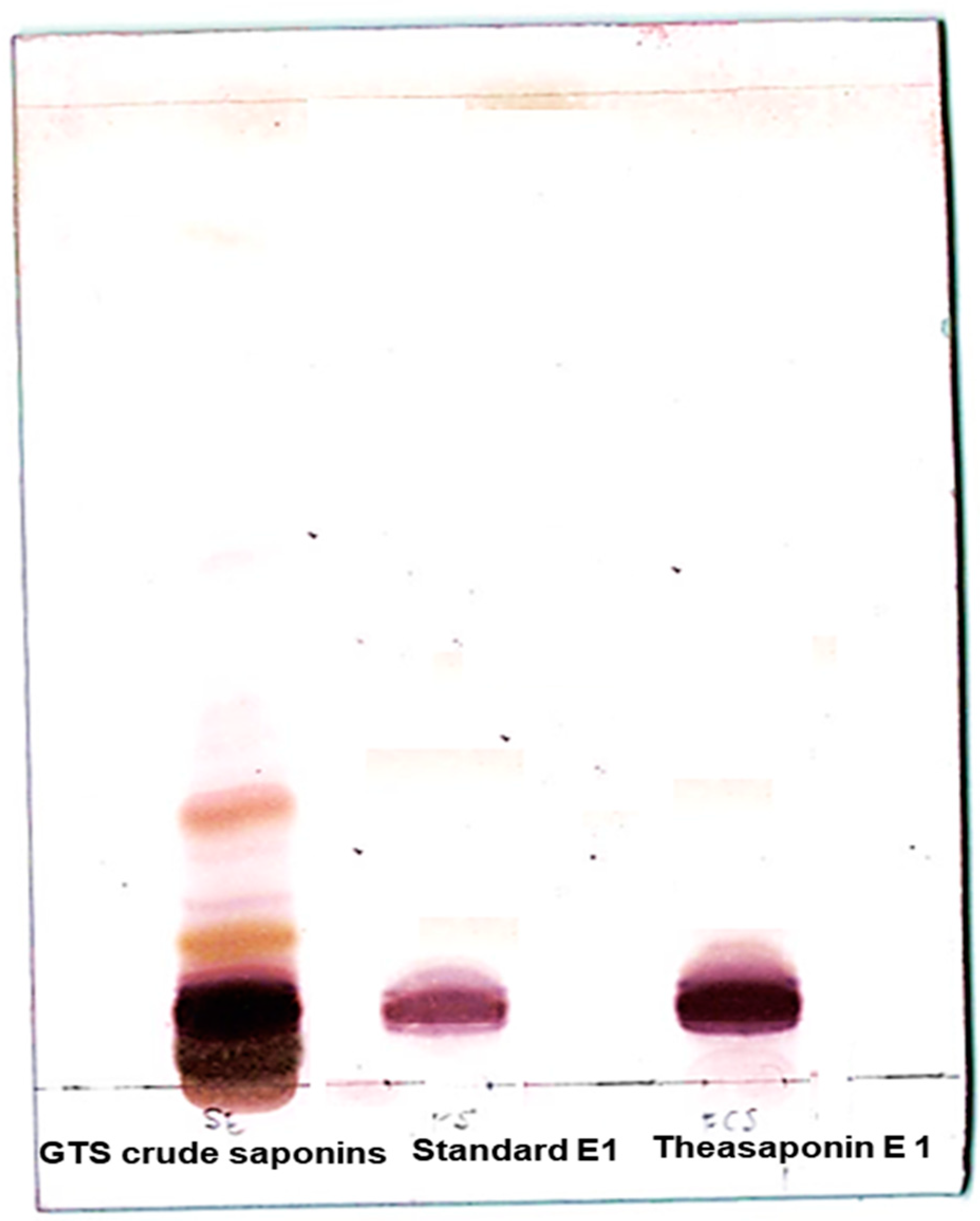
2.2. TLC Analysis
2.3. MTT Assay
2.4. Acetylcholinesterase Inhibitory Assay
3.5. Effects of Saponins on the Activities of α-Secretase (ADAM10), β-Secretase (BACE1), γ-Secretases (PS1 and NCT), IDE and Neprilysin
3.6. RT-PCR Analysis
3.7. Aß Peptides and APP Analysis by ELISA
3.8. Western Blotting
3. Discussion
4. Materials and Methods
4.1. Extraction of Saponins
4.2. Thin-Layer Chromatography
4.3. MTT Assay
4.4. Acetylcholinesterase Activity Inhibitory Assay
4.5. Enzyme Activity Analysis by Fluorometric Methods
4.6. RT- PCR Analysis
4.7. ELISA
4.8. Western Blotting
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Aβ | amyloid beta |
| ACh | acetylcholine |
| AChE | acetylcholinesterase |
| AD | Alzheimer’s disease |
| apoE | apolipoprotein E |
| APP | amyloid precursor protein |
| ADAM10 | A disintegrin and metalloproteinase domain-containing protein 10 |
| BACE1 | beta-secretase 1 |
| IDE | insulin-degrading enzymes |
| LC/TOF/MS | liquid chromatography time-of-flight mass spectrometry |
| NCT | nicastrin |
| NFT | neurofibrillary tangles |
| NMR | nuclear magnetic resonance |
| PS1 | presenilin |
| SweAPP N2a cells | Swedish amyloid precursor protein variant in neuro2a cells |
| TLC | thin layer chromatography |
References
- Hebert, L.E.; Scherr, P.A.; Bienias, J.L.; Bennett, D.A.; Evans, D.A. Alzheimer disease in the US population. Prevalence estimates using the 2000 census. Arch. Neurol. 2003, 60, 1119–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertram, L.; Tanzi, R.E. Genome-wide association studies in Alzheimer’s disease. Hum. Mol. Genet. 2009, 18, R137–R145. [Google Scholar] [CrossRef] [PubMed]
- Moya-Alvarado, G.; Gershoni-Emek, N.; Perlson, E.; Bronfman, F.C. Neurodegeneration and Alzheimer’s disease (AD). What Can Proteomics Tell Us About the Alzheimer’s Brain? Mol. Cell Proteom. 2016, 15, 409–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashyap, G.; Bapat, D.; Das, D.; Gowaikar, R.; Amritkar, R.E.; Rangarajan, G.; Ravindranath, V.; Ambika, G. Synapse loss and progress of Alzheimer’s disease -A network model. Sci. Rep. 2019, 9, 6555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beach, T.; Walker, R.; McGeer, E. Patterns of gliosis in alzheimer’s disease and aging cerebrum. Glia 1989, 2, 420–436. [Google Scholar] [CrossRef] [PubMed]
- Trojanowski, J.Q.; Lee, V.M. “Fatal attractions” of proteins. A comprehensive hypothetical mechanism underlying Alzheimer’s disease and other neurodegenerative disorders. Ann. New York Acad. Sci. 2000, 924, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Hou, H.; Rezai-Zadeh, K.; Giunta, B.; Ruscin, A. CD45 deficiency drives amyloid-β peptide oligomers and neuronal loss in Alzheimer’s disease mice. J. Neurosci. 2011, 26, 1355–1365. [Google Scholar] [CrossRef] [Green Version]
- Du, X.; Wang, X.; Geng, M.-Y. Alzheimer’s disease hypothesis and related therapies. Transl. Neurodegener. 2018, 7, 2. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.C.; Jiang, Z.F. Accumulated amyloid-beta peptide and hyperphosphorylated tau protein: Relationship and links in Alzheimer’s disease. J. Alzheimers Dis. 2009, 16, 15–27. [Google Scholar] [CrossRef]
- O’Brien, R.J.; Wong, P.C. Amyloid precursor protein processing and Alzheimer’s disease. Ann. Rev. Neurosci. 2011, 34, 185–204. [Google Scholar] [CrossRef] [Green Version]
- Xu, X. Gamma-secretase catalyzes sequential cleavages of the AbetaPP transmembrane domain. J. Alzheimers Dis. 2009, 16, 211–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vassar, R.; Bennett, B.D.; Babu-Khan, S.; Kahn, S.; Mendiaz, E.A.; Denis, P. β-Secretase cleavage of Alzheimer’s amyloid precursor protein by the transmembrane aspartic protease BACE. Science 1999, 286, 735–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trevisan, K.; Cristina-Pereira, R.; Silva-Amaral, D.; Aversi-Ferreira, T.A. Theories of Aging and the Prevalence of Alzheimer’s Disease. Biomed. Res. Int. 2019, 2019, 9171424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cline, E.N.; Bicca, M.A.; Viola, K.L.; Klein, W.L. The Amyloid-β Oligomer Hypothesis: Beginning of the Third Decade. J. Alzheimer Dis. 2018, 64, S567–S610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, R.; Yang, G.; Guo, X.; Zhou, Q.; Lei, J.; Shi, Y. Recognition of the amyloid precursor protein by human γ-secretase. Science 2019, 363, 930. [Google Scholar] [CrossRef]
- Annaert, W.; Cupers, P.; Saftig, P.; De Strooper, B. Presenilin Function in APP Processing. Ann. New York Acad. Sci. 2006, 920, 158–164. [Google Scholar] [CrossRef]
- Peron, R.; Vatanabe, I.; Manzine, P.; Camins, A.; Cominetti, M.R. Alpha-Secretase ADAM10 Regulation: Insights into Alzheimer’s Disease Treatment. Pharmaceuticals 2018, 11, 12. [Google Scholar] [CrossRef] [Green Version]
- Yan, R.; Vassar, R. Targeting the β secretase BACE1 for Alzheimer’s disease therapy. Lancet Neurol. 2014, 13, 319–329. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Ma, G.; Cai, H.; Price, N.L.; Wong, P.C. Nicastrin Is Required for Assembly of Presenilin/γ-Secretase Complexes to Mediate Notch Signaling and for Processing and Trafficking of β-Amyloid Precursor Protein in Mammals. J. Neurosci. 2003, 23, 3272–3277. [Google Scholar] [CrossRef] [Green Version]
- Iwata, N.; Tsubuki, S.; Takaki, Y.; Shirotani, K.; Lu, B.; Gerard, N.P.; Hama, E.; Lee, H.-J.; Saido, T.C. Metabolic Regulation of Brain Abeta by Neprilysin. Science 2001, 292, 1550–1552. [Google Scholar] [CrossRef]
- Kim, J.; Basak, J.M.; Holtzman, D.M. The Role of Apolipoprotein E in Alzheimer’s Disease. Neuron 2009, 63, 287–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bendiske, J.; Bahr, B.A. Lysosomal activation is a compensatory response against protein accumulation and associated synaptopathogenesis--an approach for slowing Alzheimer disease? J. Neuropathol. Exp. Neurol. 2003, 62, 451–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marambaud, P.; Zhao, H.; Davies, P. Resveratrol promotes clearance of Alzheimer’s disease amyloid-beta peptides. J. Biol. Chem. 2005, 280, 37377–37382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, W.L.; A Krafft, G.; E Finch, C. Targeting small Abeta oligomers: The solution to an Alzheimer’s disease conundrum? Trends Neurosci. 2001, 24, 219–224. [Google Scholar] [CrossRef]
- Selkoe, D.J. Translating cell biology into therapeutic advances in Alzheimer’s disease. Nature 1999, 399, A23–A31. [Google Scholar] [CrossRef]
- Walsh, D.M.; Selkoe, D.J. Oligomers on the brain: The emerging role of soluble protein aggregates in neurodegeneration. Protein Pept. Lett. 2004, 11, 213–228. [Google Scholar] [CrossRef]
- Tang, Y.P.; Gershon, E.S. Genetic studies in Alzheimer’s disease. Dialogues Clin. Neurosci. 2003, 5, 17–26. [Google Scholar]
- Kumar, A.; Nisha, C.M.; Silakari, C.; Sharma, I.; Anusha, K.; Gupta, N.; Nair, P.; Tripathi, T.; Kumar, A. Current and novel therapeutic molecules and targets in Alzheimer’s disease. J. Formos. Med. Assoc. 2016, 115, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Kihara, T.; Shimohama, S.; Sawada, H.; Honda, K.; Nakamizo, T.; Shibasaki, H.; Kume, T.; Akaike, A. Akaike alpha 7 nicotinic receptor transduces signals to phosphatidylinositol 3-kinase to block A beta-amyloid-induced neurotoxicity. J. Biol. Chem. 2001, 276, 13541–13546. [Google Scholar] [CrossRef] [Green Version]
- Söderman, A.; Mikkelsen, J.; West, M.J.; Christensen, D.Z.; Jensen, M.S. Activation of nicotinic α7 acetylcholine receptor enhances long term potentation in wild type mice but not in APPswe/PS1ΔE9 mice. Neurosci. Lett. 2011, 487, 325–329. [Google Scholar] [CrossRef]
- Woldemichael, G.M.; Wink, M. Identification and biological activities of triterpenoid saponins from Chenopodium quinoa. J. Agric. Food Chem. 2001, 49, 2327–2332. [Google Scholar] [CrossRef] [PubMed]
- Osbourn, A.E. Molecules of interest: Saponins in cereals. Phytochemistry 2003, 62, 1–4. [Google Scholar] [CrossRef]
- Serrano-Pozo, A.; Frosch, M.P.; Masliah, E.; Hyman, B.T. Neuropathological Alterations in Alzheimer Disease. Cold Spring Harb. Perspect. Med. 2011, 1, a006189. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, A. A review on Alzheimer’s disease pathophysiology and its management: An update. Pharmacol. Rep. 2015, 67, 195–203. [Google Scholar] [CrossRef]
- Qiu, W.Q.; Folstein, M. Insulin, insulin-degrading enzyme and amyloid-β peptide in Alzheimer’s disease: Review and hypothesis. Neurobiol. Aging 2006, 27, 190–198. [Google Scholar] [CrossRef]
- Schenk, D.; Basi, G.S.; Pangalos, M.N. Treatment strategies targeting amyloid beta-protein. Cold Spring Harb. Perspect Med. 2020, 2, a006387. [Google Scholar]
- Chow, V.W.; Mattson, M.P.; Wong, P.C.; Gleichmann, M. An overview of APP processing enzymes and products. Neuromolecular Med. 2010, 12, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Sinha, S.; Anderson, J.P.; Barbour, R.; Basi, G.S.; Caccavello, R.; Davis, D.; Doan, M.; Dovey, H.F.; Frigon, N.; Hong, J.; et al. Purification and cloning of amyloid precursor protein β-secretase from human brain. Nature 1999, 402, 537–540. [Google Scholar] [CrossRef]
- Hemming, M.L.; Elias, J.E.; Gygi, S.P.; Selkoe, D.J. Identification of beta-secretase (BACE1) substrates using quantitative proteomics. PLoS ONE 2009, 4, e8477. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Prior, M.; He, W.; Hu, X.; Tang, X.; Shen, W.; Yadav, S.; Kiryu-Seo, S.; Miller, R.; Trapp, B.D.; et al. Cleavage of Neuregulin-1 by BACE1 or ADAM10 Protein Produces Differential Effects on Myelination. J. Boil. Chem. 2011, 286, 23967–23974. [Google Scholar] [CrossRef] [Green Version]
- De Strooper, B.; Iwatsubo, T.; Wolfe, M.S. Presenilins and gamma-secretase: Structure, function, and role in Alzheimer disease. Cold Spring Harb. Perspect. Med. 2012, 2, a006304. [Google Scholar] [CrossRef] [PubMed]
- Kaether, C.; Haass, C.; Steiner, H. Assembly, trafficking and function of g-secretase. Neurodegener. Dis. 2006, 3, 275–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.D.; Khan, M.I.; Shin, J.H.; Lee, M.G.; Seo, H.J.; Shin, T.S.; Kim, M.Y. HPLC fractionation and pharmacological assessment of green tea seed saponins for antimicrobial, anti-angiogenic and hemolytic activities. Biotechnol. Bioprocess. Eng. 2015, 20, 1035–1043. [Google Scholar] [CrossRef]
- Khan, M.I.; Ahhmed, A.; Shin, J.H.; Baek, J.S.; Kim, M.Y.; Kim, J.D. Green Tea Seed Isolated Saponins Exerts Antibacterial Effects against Various Strains of Gram Positive and Gram Negative Bacteria, a Comprehensive Study In vitro and In vivo. Evid. Based Complement. Altern. Med. 2018, 2018, 3486106–3486112. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.J.; Bailey, H.; Mukhtar, H. Green tea polyphenols for prostate cancer chemoprevention: A translational perspective. Phytomedicine 2010, 17, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Sundaram, R.; Naresh, R.; Shanthi, P.; Sachdanandam, P. Modulatory effect of green tea extract on hepatic key enzymes of glucose metabolism in streptozotocin and high fat diet induced diabetic rats. Phytomedicine 2013, 20, 577–584. [Google Scholar] [CrossRef]
- Kobayashi, H.; Tanaka, Y.; Asagiri, K.; Asakawa, T.; Tanikawa, K.; Kage, M.; Yagi, M. The antioxidant effect of green tea catechin ameliorates experimental liver injury. Phytomedicine 2010, 17, 197–202. [Google Scholar] [CrossRef]
- Mancini, E.; Beglinger, C.; Drewe, J.; Zanchi, D.; Lang, U.E.; Borgwardt, S. Green tea effects on cognition, mood and human brain function: A systematic review. Phytomedicine 2017, 34, 26–37. [Google Scholar] [CrossRef] [Green Version]
- Eldeen, I.; Elgorashi, E.; Van Staden, J. Antibacterial, anti-inflammatory, anti-cholinesterase and mutagenic effects of extracts obtained from some trees used in South African traditional medicine. J. Ethnopharmacol. 2005, 102, 457–464. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors on proper request. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Accession NO. | Product Size (bp) | Forward | Reverse |
|---|---|---|---|---|
| α-secretase/ADAM10 | NM_007399 | 881 | 5′-GCCAGCCTATCTGTGGAAACGGG-3′ | 5′-TTAGCGTCGCATGTGTCCCATTTG-3′ |
| β-secretase/BACE1 | NM_011792 | 216 | 5′-GGATTATGGTGGCCTGAGCA-3′ | 5′- CACGAGAGGAGACGACGATG -3′ |
| γ-secretase/PSP1 | NM_001362271 | 195 | 5’-CTGTCCAGCTGCTCCAATGA-3’′ | 5’-CTTGAGTTACTCTGGGGCCG-3’ |
| γ-secretase/NCT | NM_021607 | 186 | 5′- CTAGAGAGGCCGCTACGAGT-3′ | 5′- CTCCACTGAGTTTCCCCCAC -3′ |
| Insulin-degrading enzyme/IDE | NM_031156 | 576 | 5′-ACCTGTGAACTGAGCTGCAGAA-3′ | 5′-TAGAAACGTCTGGTCATCCAACA-3′ |
| Neprilysin | NM_008604 | 250 | 5’-GCAGCCTCAGCCGAAACTAC-3’ | 5’-TCAGCATCCATCCAAGTAAG-3 |
| β-actin/Actb | NM_007393 | 172 | 5’-AGGGAAATCGTGCGTGACAT-3’ | 5’-GGAAAAGAGCCTCAGGGCAT-3’ |
| Antibody | Host | Species Reactivity | Target | Source |
|---|---|---|---|---|
| Anti-ADAM10 antibody | Rabbit | Mouse, Human | ADAM10 | Abcam ab1997 |
| Anti-human/mouse BACE1 | Monoclonal Mouse | Human, Mouse | BACE1 | R&D Systems MAB931 |
| Anti-Presenilin [APS 11] antibody | Rabbit | Mouse, Human | PS1 | Abcam ab15456 |
| Anti-NCSTN Antibody | Rabbit | Mouse, Human | NCT | US Biological 364323 |
| Anti-insulin degrading enzyme/IDE antibody | Rabbit | Mouse, Human | IDE | Abcam, ab32216 |
| Human/Mouse Neprilysin/CD10 Antibody | Monoclonal Rat | Mouse, Human | NEP | R&D Systems MAB1126 |
| Anti-amyloid precursor protein antibody | Rabbit | Mouse, Human | APP | Abcam ab2072 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, M.I.; Shin, J.H.; Kim, M.Y.; Shin, T.S.; Kim, J.D. Green Tea Seed Isolated Theasaponin E1 Ameliorates AD Promoting Neurotoxic Pathogenesis by Attenuating Aβ Peptide Levels in SweAPP N2a Cells. Molecules 2020, 25, 2334. https://doi.org/10.3390/molecules25102334
Khan MI, Shin JH, Kim MY, Shin TS, Kim JD. Green Tea Seed Isolated Theasaponin E1 Ameliorates AD Promoting Neurotoxic Pathogenesis by Attenuating Aβ Peptide Levels in SweAPP N2a Cells. Molecules. 2020; 25(10):2334. https://doi.org/10.3390/molecules25102334
Chicago/Turabian StyleKhan, Muhammad Imran, Jin Hyuk Shin, Min Yong Kim, Tai Sun Shin, and Jong Deog Kim. 2020. "Green Tea Seed Isolated Theasaponin E1 Ameliorates AD Promoting Neurotoxic Pathogenesis by Attenuating Aβ Peptide Levels in SweAPP N2a Cells" Molecules 25, no. 10: 2334. https://doi.org/10.3390/molecules25102334
APA StyleKhan, M. I., Shin, J. H., Kim, M. Y., Shin, T. S., & Kim, J. D. (2020). Green Tea Seed Isolated Theasaponin E1 Ameliorates AD Promoting Neurotoxic Pathogenesis by Attenuating Aβ Peptide Levels in SweAPP N2a Cells. Molecules, 25(10), 2334. https://doi.org/10.3390/molecules25102334