Response of Pseudokirchneriella subcapitata in Free and Alginate Immobilized Cells to Heavy Metals Toxicity

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Alga
2.2. Metals
2.3. Alginate Source
2.4. Immobilization Procedure
2.5. Toxicity Testing
2.5.1. Microplate Procedure with Free Cells
2.5.2. Microplate Procedure with Algae-Alginate Beads
2.5.3. Toxicity-Response Parameter
2.5.4. Calculation of EC50 and EC100
2.6. Statistical Analysis of Data
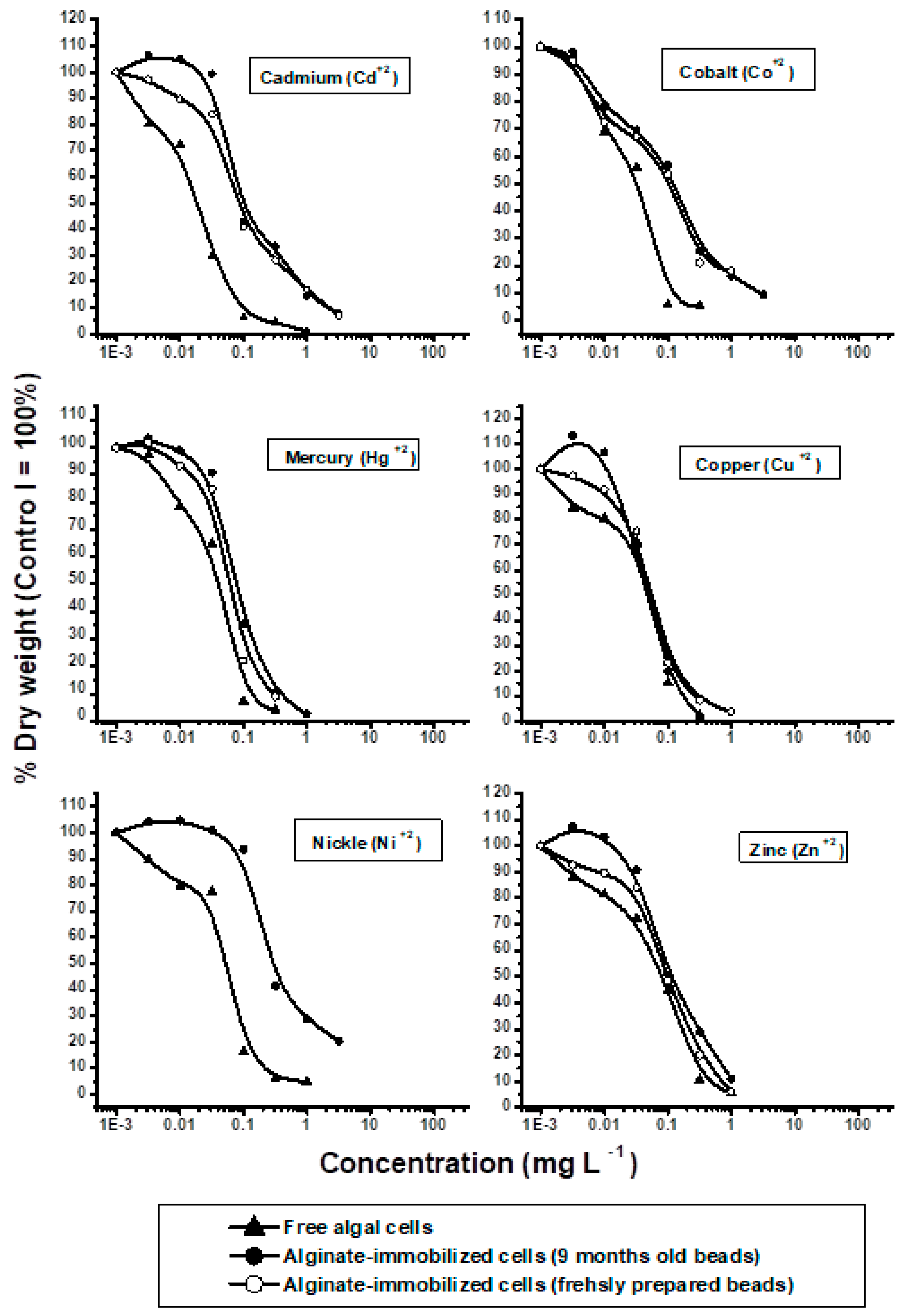
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Föistner, V.; Wittmann, G.T.W. Metal Pollution in Aquatic Environment; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1979. [Google Scholar]
- Saleem, M.H.; Ali, S.; Rehman, M.; Rana, M.S.; Rizwan, M.; Kamran, M.; Imran, M.; Riaz, M.; Soliman, M.H.; Elkelish, A.; et al. Influence of phosphorus on copper phytoextraction via modulating cellular organelles in two jute (Corchorus capsularis L.) varieties grown in a copper mining soil of Hubei Province, China. Chemosphere 2020, 248, 126032. [Google Scholar] [CrossRef] [PubMed]
- El-Esawi, M.A.; Elkelish, A.; Soliman, M.; Elansary, H.O.; Zaid, A.; Wani, S.H. Serratia marcescens BM1 Enhances Cadmium Stress Tolerance and Phytoremediation Potential of Soybean Through Modulation of Osmolytes, Leaf Gas Exchange, Antioxidant Machinery, and Stress-Responsive Genes Expression. Antioxidants 2020, 9, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forsberg, C.; Ryding, S.-O.; Claesson, A.; Forsberg, A. Water chemical analysis and/or algal assays? Sewage effluent and polluted lake water studies. Mitt. Int. Ver. Theor. Angew. Limnol. 1978, 21, 352–363. [Google Scholar]
- Miller, W.E.; Green, J.C.; Sheroyama, T. The Selenastrumcapricornutum Printz Algal Assay Bottle Test; Environmental Research Laboratory, Office of Research and Development, Environmental Protection Agency: Corvallis, OR, USA, 1978. [Google Scholar]
- Greene, J.C.; Bartles, C.I.; Warren-Hicks, W.J.; Parkhurst, B.R.; Linder, G.L.; Peterson, S.A.; Miller, W.E. Protocols for Short Term Toxicity Screening of Hazardous Waste Site; EPA/600/3-88/029; U.S. Environmental Protection Agency: Corvallis, OR, USA, 1989. [Google Scholar]
- ISO. Water Quality-Algal Growth Inhibition Test; Draft International Standard: Geneva, Switzerland, 1987. [Google Scholar]
- OECD. Guidelines for the Testing of Chemicals: Draft Revised Guideline and 201 (Freshwater Algae and Cyanobacteria, Growth Inhibition Test; Organization for Economic Cooperation and Development: Paris, France, 2002; p. 21. [Google Scholar]
- Franklin, N.M.; Stauber, J.L.; Apte, S.C.; Lim, R.P. Effect of initial cell density on the bioavailability and toxicity of copper in microalgal bioassays. Enviro. Toxicol. Chem. 2002, 21, 742–751. [Google Scholar] [CrossRef]
- Lukavský, J.; Komárek, J.; Lukavská, A.; Ludvík, J.; Pokorný, J. Metabolic activity and cell structure of immobilized algal cells (Chlorella, Scenedesmus). Arch. Hydrobiol. 1986, 43, 261–297. [Google Scholar]
- Kaparapu, J.; Geddada, M.N.R. Applications of immobilized algae. J. Algal Biomass Util. 2016, 7, 122–128. [Google Scholar]
- Lau, P.S.; Tam, N.F.Y.; Wang, Y.S. Wastewater Nutrients (N and P) Removal by Carrageenan and Alginate Immobilized Chlorella Vulgaris. Environ. Technol. 1997, 18, 945–951. [Google Scholar] [CrossRef]
- Barkley, N.P. Extraction of mercury from groundwater using immobilized algae. J. Air Waste Manag. Assoc. 1991, 41, 1387–1393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joo, D.S.; Cho, M.G.; Lee, J.S.; Park, J.H.; Kwak, J.K.; Han, Y.H.; Bucholz, R. New strategy for the cultivation of microalgae using microencapsulation. J. Microencapsul. 2001, 18, 567–576. [Google Scholar]
- Dos Santos, M.M.; Moreno-Garrido, I.; Goncalves, F.; Soares, A.M.; Ribeiro, R. An in situ bioassay for estuarine environments using the microalga Phaeodactylumtricornutum. Environ. Toxicol Chem. 2002, 21, 567–574. [Google Scholar] [CrossRef]
- Podola, B.; Nowack, E.C.; Melkonian, M. The use of multiple strain algal sensor chips for the detection and identification of volatile organic compounds. Biosens. Bioelectron. 2004, 19, 1253–1260. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Hamid, M.I. Development and application of a simple procedure for toxicity testing using immobilized algae. Water Sci. Technol. 1996, 23, 129–138. [Google Scholar] [CrossRef]
- Davis, T.A.; Lpanes, F.; Volesky, B.; Mucci, A. Metal selectivity of Sargassum spp. and their alginates in relation to their α- L- guluronic acid content and conformation. Environ. Sci. Technol. 2003, 37, 261–267. [Google Scholar] [CrossRef]
- Davis, T.A.; Volesky, B.; Mucci, A. A review of the biochemistry of heavy metal biosorption by brown algae. Water Res. 2003, 37, 4311–4330. [Google Scholar] [CrossRef]
- Awasthi, M.; Rai, L.C. Adsorption of nickel, zinc and cadmium by immobilized green algae and cyanobacteria: A comparative study. Ann. Microbiol. 2004, 54, 257–267. [Google Scholar]
- Abdel-Hamid, M.I. Immobilization of algal cells: A first Biotechnological Step for the Handling of Cell Material for Toxicity Screening and Water Quality Studies. In Proceedings of the First Egyptian-Italian Symposium on Biotechnology, Cairo, Morocco, 21–23 November 1992; pp. 295–302. [Google Scholar]
- Van Donk, E.; Abdel-Hamid, M.I.; Faafeng, B.A.; Källqvist, T. Effects of Dursban® 4E and its carrier on three algal species during exponential and P-limited growth. Aquat. Toxicol. 1992, 23, 181–192. [Google Scholar] [CrossRef]
- Steinman, A.D.; Lamberti, G.A. Biomass and pigments of benthic algae. In Methods in Stream Ecology; Hauer, F.R., Lamberti, G.A., Eds.; Academic Press: San Diego, CA, USA, 1996; p. 297. [Google Scholar]
- Walsh, G.E.; Deans, C.H.; Mclaughlin, L.L. Comparison of four methods for calculating the EC50 from algal population growth. Environ. Toxicol. Chem. 1987, 6, 767–770. [Google Scholar]
- Abdel-Hamid, M.I.; Shaaban-Dessouoki, S.A.; Skulberg, O.M. Biological Indexing of Water Quality of the River Nile. In Proceedings International Conference, Cairo, Morocco, 3–5 February 1992; Ministry of Public Works and Water Resources: Cairo, Morocco, 1992. [Google Scholar]
- Peterson, H.G.; Healey, F.P.; Wagemann, R. Metal toxicity to algae: A highly PH dependent phenomenon. Can. J. Fish. Aquat. Sci. 1984, 41, 974–979. [Google Scholar] [CrossRef]
- Petrovic, A.; Simonic, M. Removal of heavy metal ions from drinking water by alginate-immobilised Chlorella sorokiniana. Int. J. Environ. Sci. Technol. 2016, 13, 1761–1780. [Google Scholar] [CrossRef] [Green Version]
- Górak, M.; Żymańczyk-Duda, E. Reductive activity of free and immobilized cells of cyanobacteria toward oxophosphonates—Comparative study. J. Appl. Phycol. 2017, 29, 245–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartlet, L.; Rabe, F.W.; Funk, W.H. Effects of copper, zinc and cadmium on Selenastrum capricornatum. Water Res. 1974, 8, 179–185. [Google Scholar] [CrossRef]
- Blaise, C.; Legautt, R.; Bermingham, N.; van Coillie, R.; Vasseur, P. A simple microplate algal assay technique for aquatic toxicity. Toxic. Assess. 1986, 1, 261–281. [Google Scholar] [CrossRef]
- Collard, J.M.; Matagne, R.F. Isolation and genetic analysis of Chlamydomonas reinhardtii strains resistant to cadmium. Appl. Environ. Microbiol. 1990, 56, 2051–2055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cain, J.R.; Paschal, D.C.; Hayden, M.C. Toxicity and bioaccumulation of cadmium in the colonial green algae Scenedesmus obliquus. Arch. Environ. Contam. Toxicol. 1980, 9, 9–16. [Google Scholar] [CrossRef]
- Einicker-Lamas, M.; Soares, M.J.; Soares, M.S.; Oliveira, M.M. Effects of cadmium on Euglena gracilis membrane lipids. Braz. J. Med. Biol. Res. 1996, 29, 941–948. [Google Scholar]
- Shanker, K.; Mishra, S.; Srivastava, S.; Srivastava, R.; Daas, S.; Prakash, S.; Srivastava, M.M. Effect of selenite and selenate on plant uptake of cadmium by maize (Zea mays). Bull. Environ. Contam. Toxicol. 1996, 56, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Price, N.M.; Morel, F.M.M. Cadmium and cobalt substitution for zinc in a marine diatom. Nature 1990, 344, 658–660. [Google Scholar] [CrossRef]
- Lee, J.G.; Roberts, S.B.; Morel, F.M. Cadmium: A nutrient for the marine diatom Thalassiosiraweissjlogii. Limnol. Oceanogr. 1995, 40, 1056–1063. [Google Scholar] [CrossRef]
- Lane, T.W.; Morel, F.M. Regulation of Carbonic Anhydrase Expression by Zinc, Cobalt, and Carbon Dioxide in the Marine Diatom Thalassiosiraweissflogii. Plant Physiol. 2001, 123, 345–352. [Google Scholar] [CrossRef] [Green Version]
- Butler, A. Acquisition and utilization of transition metal ions by marine organisms. Science 1998, 281, 207–210. [Google Scholar] [CrossRef] [Green Version]
- Rosko, J.J.; Rachlin, J.W. The Effect of Copper, Zinc, Cobalt and Manganese on the Growth of the Marine Diatom Nitzschia closterium; Torrey Botanical Society: New York, NY, USA, 1975. [Google Scholar]
- Plekhanov, S.E.; Chemeris, U.K. Early toxic effect of zinc, cobalt, and cadmium on photosynthetic activity of green alga Chlorella pyrenoidosa Chick S-39. Biol. Bull. 2003, 5, 610–616. [Google Scholar]
- Abdel Hameed, M.S. Continuous removal and recovery of lead by alginate beads, free and alginate-immobilized Chlorella vulgaris. Afr. J. Biotechnol. 2006, 5, 1819–1823. [Google Scholar]
- El-Sheekh, M.M.; El-Nagger, A.H.; Osman, M.E.H.; El-Mazaly, E. Effect of cobalt on growth, pigments and the photosynthetic electron transport in Monoraphidiumminutum and Nitzschiaperminta. Baraz. J. Plant Physiol. 2003, 15, 59–166. [Google Scholar]
- Gajdosova, J.; Reichrtova, E. Different growth response of Euglena gracilis to Hg, Cd, Cr, and Ni compounds: Fresenius. J. Anal. Chem. 1996, 354, 641–642. [Google Scholar] [CrossRef] [PubMed]
- De Filippis, L.F.; Ziegler, H. Effect of sub-lethal concentrations of zinc, cadmium and mercury on the photosynthetic carbon reduction cycle of Euglena. J. Plant Physiol. 1993, 142, 167–172. [Google Scholar] [CrossRef]
- Startton, G.W.; Hurker, A.L.; Corke, C.T. Effect of mercuric ion on the growth, photosynthesis, and nitrogenase activity of Anabaena inaequalis. Appl. Environ. Microbiol. 1979, 38, 537–543. [Google Scholar] [CrossRef] [Green Version]
- Rai, L.C.; Dey, R. Environmental effects on the toxicity of methyl mercuric chloride to Chlorella vulgaris. Acta Hydrochim. Hydrobiol. 1980, 8, 319–327. [Google Scholar]
- Sorentino, C. The effects of heavy metals on phytoplankton. A review phykos 18, 149–161. Stains resistant to cadmium. App. Environ. Microbiol. 1979, 56, 2051–2055. [Google Scholar]
- O’kelly, J.C. Inorganic Nutrients. In Algal Physiology and Biochemistry; Stewart, W.D.P., Ed.; Blackwell Scientific publications: Oxford, UK, 1974; pp. 610–635. [Google Scholar]
- Wong, P.K.; Chang, L. Effects of copper, chromium and nickel on growth, photosynthesis and chlorophyll a synthesis of Chlorella pyrenoidosa. Environ. Pollut. 1991, 72, 127–139. [Google Scholar] [CrossRef]
- De Filippis, L.F. The effect of sub-lethal concentrations of mercury and zinc on Chlorella. v. The counteraction of metal toxicity by selenium and sulphhydryl compounds. Z. Pflanzenphysiol. 1979, 93, 63–68. [Google Scholar] [CrossRef]
- De Filippis. The effect of heavy metal compounds on the permeability of Chlorella celli. Z. Pflanzenphysiol. 1979, 922, 39–49. [Google Scholar]
- Quartacci, M.F.; Cosi, E.; Navari-Izzo, F. Lipids and NADPH-dependent superoxide production in plasma membrane vesicles from roots of wheat grown under copper deficiency or excess. J. Exp. Bot. 2001, 52, 77–84. [Google Scholar] [PubMed]
- Mostowska, A. Environmental Factors Affecting Chloroplasts. In Handbook of Photosynthesis; Pessarakli, M., Ed.; Marcel Dekker: New York, NY, USA, 1997; pp. 407–426. [Google Scholar]
- Reiriz, S.; Cid, A.; Torres, E.; Abalde, J.; Herrero, C. Different responses of the marine diatom Phaeodactylumtricornutum to copper toxicity. Microbiologia 1994, 10, 263–272. [Google Scholar] [PubMed]
- Trollope, D.R.; Evans, B. Concentrations of copper, iron, lead, nickel and zinc in freshwater algal blooms. Environ. Pollut. 1970, 11, 109–116. [Google Scholar] [CrossRef]
- Soliman, M.; Alhaithloul, H.A.; Hakeem, K.R.; Alharbi, B.M.; El-Esawi, M.; Elkelish, A. Elkelish Exogenous Nitric Oxide Mitigates Nickel-Induced Oxidative Damage in Eggplant by Upregulating Antioxidants, Osmolyte Metabolism, and Glyoxalase Systems. Plants 2019, 8, 562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitton, B.A.; Say, P.J.; Jupp, B.P. Accumulation of zinc, cadmium and lead by the aquatic liverwort Scapania. Environ. Pollut. 1982, 3, 299–316. [Google Scholar] [CrossRef]
- Loez, C.R.; Topalian, M.L.; Salibian, A. Effects of zinc on the structure and growth dynamics of a natural freshwater phyto- plankton assemblages reared in the laboratory. Environ. Pollut. 1995, 88, 275–281. [Google Scholar] [CrossRef]
- Meylan, S.; Odzak, N.; Behra, R.; Sigg, L. Speciation of copper and zinc in natural freshwater: Comparison of volumetric measurements, diffusive gradients in the thin film (DGT) and chemical equilibrium models. Anal. Chim. Acta 2004, 510, 91–100. [Google Scholar] [CrossRef]
- Falchuk, K.H.; Fawcett, D.W.; Vallee, B.L. Role of zinc in cell division of Euglena gracilis. J. Cell Sci. 1975, 17, 57–78. [Google Scholar]
- Paulsson, M.; Nyström, B.; Blanck, H. Long term toxicity of zinc to bacteria and algae in periphyton communities from the River GötaÄlv, based on a microcosm study. Aquat. Toxicol. 2000, 47, 243–257. [Google Scholar] [CrossRef]
- Bànerjee, M.; Mishra, S.; Chatterjee, J. Scavenging of nickel and chromium toxicity in Alucosirafertilissimaby immobilization. Effect on nitrogen assimilating enzymes. Electron. J. Biotechnol. 2004, 7, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Hörcsik, Z.T.; Balogh, Á. Intracellular distribution of chromium and toxicity on growth in Chlorella pyrenoidosa. Acta Biol. Szeged. 2002, 46, 57–58. [Google Scholar]
- Rachlin, J.W.; Grosso, A. The growth response of the green alga Chlorella vulgaris to combined cation exposure. Arch. Environ. Contam. Toxicol. 1993, 24, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Greger, M.; Tillberg, J.; Johansson, M. Aluminium effects on Scenedesmusobtusiusculus with different phosphorus status. I. Mineral uptake. Physiol. Plant. 1992, 84, 193–201. [Google Scholar]
- Abdel-Hamid, M.I.; Skulberg, O.M. Effect of selenium on the growth of some selected green and blue-green algae. Lakes Reserv.: Res. Manag. 1995, 1, 205–211. [Google Scholar] [CrossRef]
- Lindström, K. Selenium as a growth factor for plankton algae in laboratory experiments and in some Swedish lakes. Dev. Hydrobiol. 1983, 13, 35–47. [Google Scholar] [CrossRef]
- Lindström, K. Selenium requirement of the dinoflagellate Pendinopsisborgei (Lemm). Int. Revueges. Hydrobiol. 1985, 70, 77–78. [Google Scholar] [CrossRef]
- Sielicki, M.; Burnham, J.C. The effect of selenite on the physiological and morphological properties of the blue-green alga phormidiumluridum var. olivacea. J. Phycol. 1973, 9, 509–514. [Google Scholar]
- Agency for Toxic Substances, Disease Registry (ARSDR). Toxologic Profile of Arsenic; ATSDR: Atlanta, GA, USA, 2000. [Google Scholar]
- Blank, H. Species dependent variation among aquatic organisms in their sensitivity to chemicals. Ecol. Bull. 1984, 36, 107–119. [Google Scholar]
- Fuhua, C.; Weiqi, C.; Shugui, D. Toxicities of four arsenic species to Scenedesmusobliguus and influence of phosphate on inorganic arsenic toxicities. Toxicol. Environ. Chem. 1994, 41, 1–7. [Google Scholar] [CrossRef]
- Blanck, H.; Wangberg, S.A. Validity of an ecotoxicological test system Short-term and long-term effects of arsenate on marine periphyton communities in laboratory systems. Can. J. Fish. Aquat. Sci. 1988, 45, 1807–1815. [Google Scholar] [CrossRef]
- Knauer, K.; Behra, R.; Hemond, H. Toxicity of inorganic and methylated arsenic to algal communities from lakes along an arsenic contamination gradient. Aquat. Toxicol. 1999, 46, 221–230. [Google Scholar] [CrossRef]
- El-Naggar, N.E.; Hamouda, R.A.; Mousa, I.E.; Abdel-Hamid, M.S.; Rabei, N.H. Biosorption optimization, characterization, immobilization and application of Gelidiumamansii biomass for complete Pb2+ removal from aqueous solutions. Sci. Rep. 2018, 8, 13456. [Google Scholar] [CrossRef]
- Rivkin, R.B. Effects of lead on growth of marine diatom Skeletonemacostatum. Mar. Biol. 1979, 50, 239–247. [Google Scholar] [CrossRef]
- Seiler, H.G.; Sigel, H.; Sigel, A. Handbook on the Toxicity of Tnorganic Compound; Marcel Dekker. Inc.: New York, NY, USA, 1988; p. 633. [Google Scholar]
- Clayton, G.D.; Clayton, F.E. Patty’s Industrial Hygiene and Toxicology, 3rd ed.; John Wiley Sons: New York NY, USA, 1981–1982; Volume 2A, 2B, 2C: Toxicology; p. 2131. [Google Scholar]
- Neal, C.; Smith, C.J.; Jeffery, H.A.; Jarvie, H.P.; Robson, A.J. Trace element concentrations in the major rivers entering the Humber estuary, NE England. J. Hydrol. 1996, 182, 37–64. [Google Scholar] [CrossRef]
- Baxter, M.; Jensen, T. Uptake of magnesium, strontium, barium, and manganese by Plectonemaboryanum (cyanophyceae) with special reference to polyphosphate bodies. Protoplasma 1980, 104, 81–89. [Google Scholar] [CrossRef]
- Wilkinson, S.C.; Goulding, K.H.; Robinson, P.K. Mercury removal by immobilized algae in batch culture systems. J. Appl. Phycol. 1990, 2, 223–230. [Google Scholar] [CrossRef]
- Tam, N.F.Y.; Wang, Y.S.; Simpson, C.G. Repeated removal of copper by alginate beads and the enhancement by microalgae. Biotechnol. Tech. 1998, 12, 187–190. [Google Scholar] [CrossRef]
- Myklestad, S. Ion-exchange properties of brown algae I. Determination of rate mechanism for calcium hydrogen ion exchange for particles from Laminariahyperborea and Laminariadigitata. J. Appl. Chem. 1968, 18, 30–36. [Google Scholar] [CrossRef]
- Prask, I.A.; Plocke, D.J. A Role for Zinc in the Structural Integrity of the Cytoplasmic Ribosomes of Euglena gracilis. Plant Physiol. 1971, 48, 150–155. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: Samples of the compounds and the algal species are available from the authors. |



{kind=link}
{kind=link}
| Name | Formula | Tested as Metal Ions | Obtained from |
|---|---|---|---|
| Cadmium chloride | CdCl2·2½ H2O | Cd2+ | BDH, Dubai, UAE |
| Cobalt chloride | CoCl2·6H2O | Co2+ | MERCK, Darmstadt, Germany |
| Mercuric chloride | HgCl2 | Hg2+ | MERCK, Darmstadt, Germany |
| Cupric chloride | CuCl2·2H2O | Cu2+ | MERCK, Darmstadt, Germany |
| Nickel sulfate | NiSO4·6H2O | Ni2+ | MERCK, Darmstadt, Germany |
| Zinc chloride | ZnCl2 | Zn2+ | MERCK, Darmstadt, Germany |
| Chromium nitrate | Cr (NO3)3·9H2O | Cr3+ | MERCK, Darmstadt, Germany |
| Aluminum chloride | AlCl3·6HO | Al3+ | MERCK, Darmstadt, Germany |
| Selenium dioxide | SeO2 | Se4+ | MERCK, Darmstadt, Germany |
| Sodium selenite a | Na2SeO3·5H2O | Se4+ | MERCK, Darmstadt, Germany |
| Sodium arsenate | Na2HAsO4·7HO | As5+ | MERCK, Darmstadt, Germany |
| Lead nitrate | Pb (NO3)2 | Pb2+ | BDH, Dubai, UAE |
| Strontium chloride | SrCl2·6H2O | Sr2+ | MERCK, Darmstadt, Germany |
| EC50 (mg L−1) a | EC100 (mg L−1) b | ||||||
|---|---|---|---|---|---|---|---|
| Free Cells | Immobilized Cells | Free Cells | Immobilized Cells | ||||
| Chemical | Tested as | Old c | New d | Old | New | ||
| CdCl2·2½ H2O | Cd2+ | 0.018 | 0.09 ***e | 0.09 | 0.1 | 1.0 | 1.0 |
| CoCl2·6H2O | Co2+ | 0.03 | 0.06 *** | 0.061 | 0.32 | 1.0 | 1.0 |
| HgCl2 | Hg2+ | 0.039 | 0.06 *** | 0.063 | 0.032 | 1.0 | 1.0 |
| CuCl2·2H2O | Cu2+ | 0.048 | 0.05 | 0.05 | 0.1 | 0.1 | 0.1 |
| NiSO4·6H2O | Ni2+ | 0.055 | 0.03 *** | NT f | 0.1 | 3.2 | NT |
| ZnCl2 | Zn2+ | 0.08 | 0.1 * | 0.1 | 0.32 | 0.32 | 0.32 |
| Cr(NO3)3·9H2O | Cr3+ | 0.2 | 0.3 *** | 0.3 | 1.0 | 3.2 | 3.2 |
| AlC3·6HO | Al3+ | 0.75 | 1.8 *** | NT | 1.0 | 3.2 | NT |
| SeO2 | Se4+ | 1.2 | 1.4 | NT | 3.2 | 3.2 | NT |
| Na2SeO3·5H2O | Se4+ | 3.0 | 3.0 | NT | 32 | 32 | NT |
| Na2HAsO4·7HO | As5+ | 3.0 | 4.0 * | 4.0 | 32 | 32 | 32 |
| Pb(NO3)2 | Pb2+ | 3.3 | 5.0 * | 5.0 | 32 | 32 | 32 |
| SrCl2·6H2O | Sr2+ | 160 | 180 *** | NT | >320 | >320 | NT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Hasawi, Z.M.; Abdel-Hamid, M.I.; Almutairi, A.W.; Touliabah, H.E. Response of Pseudokirchneriella subcapitata in Free and Alginate Immobilized Cells to Heavy Metals Toxicity. Molecules 2020, 25, 2847. https://doi.org/10.3390/molecules25122847
Al-Hasawi ZM, Abdel-Hamid MI, Almutairi AW, Touliabah HE. Response of Pseudokirchneriella subcapitata in Free and Alginate Immobilized Cells to Heavy Metals Toxicity. Molecules. 2020; 25(12):2847. https://doi.org/10.3390/molecules25122847
Chicago/Turabian StyleAl-Hasawi, Zaki M., Mohammad I. Abdel-Hamid, Adel W. Almutairi, and Hussein E. Touliabah. 2020. "Response of Pseudokirchneriella subcapitata in Free and Alginate Immobilized Cells to Heavy Metals Toxicity" Molecules 25, no. 12: 2847. https://doi.org/10.3390/molecules25122847
APA StyleAl-Hasawi, Z. M., Abdel-Hamid, M. I., Almutairi, A. W., & Touliabah, H. E. (2020). Response of Pseudokirchneriella subcapitata in Free and Alginate Immobilized Cells to Heavy Metals Toxicity. Molecules, 25(12), 2847. https://doi.org/10.3390/molecules25122847