1. Introduction
Bladder cancer (BC) is a significant problem worldwide and one of the leading cause of death. American Cancer Society estimated 81,400 new cases of BC and approximately 17,980 deaths in 2020 [
1]. From the economic point of view, BC is the most expensive cancer to treat as it frequently requires diagnostic procedures such as cystourethroscopy, urine cytology, and radiological imaging [
2]. Numerous risk factors may be involved in the development of BC, including smoking, work-related exposure to aromatic amines and polyurethane products, hereditary genetic background, nutritional factors, certain medical conditions, long-term treatment with chemotherapy agents, such as cyclophosphamide or immunosuppressive drugs like glucocorticoids, age, sex, ethnicity, race, and socio-economic status. BC mortality varies in different countries. The highest rates are reported in European countries such as Denmark, UK, Belgium, and Italy, whereas the lowest rates are observed in Asian countries, including Japan, China, and Singapore [
3,
4]. Some epidemiological data show that populations in South-East Asia have a 4- to 10-fold lower incidence of, and death from BC, when compared with those in the USA. These geographic discrepancies related to the incidence of many tumor types may be associated with differences in diet and intake of nutrients, of which some may have chemopreventive properties [
4,
5].
Treatment of BC with the conventional chemotherapeutic regimens is often associated with serious side effects (a strong systemic toxicity and local irritation) [
6], while the efficacy of available intravesical immunotherapy agents such as bacillus Calmette–Guerin is ~30–70% [
7]. Accordingly, a novel and more effective therapeutic approach is urgently needed for the treatment of BC. Administration of intravesical or intraperitoneal hyperthermal chemotherapy (HIVEC or HIPEC) results in high locoregional drug concentration. With this approach, systemic toxicity is limited by peritoneal–plasma barrier and its duration is relatively short (from 30 min to 2 h) [
8]. This form of therapy leads to an increased exposure of tumor cells to the drug and long-term survival of patients and mice-bearing tumor [
9,
10,
11]. Hyperthermia increases chemotherapy penetration at the peritoneal surface and may increase the sensitivity of the cancer cells to chemotherapy by impairing DNA repair machinery [
9]. Hyperthermia may also induce apoptosis, activate heat-shock proteins that activate immune-modulating natural killer cells, and inhibit angiogenesis [
9].
It is known that cancers originating from organs in the peritoneal cavity (e.g., ovarian, pancreatic, colorectal, gastric, and liver) and extra-abdominal tumors originating from lymphoma, lung, and breast as well as a small number of unknown primary tumors account for approximately 250,000 new cancer cases annually in the USA [
10]. Despite advanced techniques and surgical removal of a macroscopically visible tumor in the peritoneal cavity, the problem of microscopic tumor residues remains, as well as the escape of tumor cells into the peritoneal cavity during surgery of advanced tumors that form an iatrogenic tumor [
11]. Malignant ascites is a sign of peritoneal carcinomatosis, the presence of malignant cells in the peritoneal cavity. In many types of cancer, ascites is a prognostic sign of advanced stage, with a survival rate of 11% for patients suffering from ascites for more than 6 months [
11]. Targeting cancer cells with nontoxic therapeutic agents differ in the mechanisms of action, enhances the treatment efficacy and is reported to have an additive or supra-additive effect [
9,
11]. Experimental tumors are known to be of great importance for the study of various forms of therapy. Ehrlich ascites tumor (EAT), a consequence of spontaneous breast cancer developed in female mice, is one of the most commonly used and adequate models of research in this area. EAT is referred as an undifferentiated carcinoma and is originally hyperdiploid, has high transplantable capability, no-regression, rapid proliferation, 100% malignancy, and also does not express tumor-specific transplantation antigen (TSTA) [
12]. In an effort to improve local tumor control, a multimodality treatment strategy including locoregional hyperthermia, chemotherapy, and immunomodulation with quercetin, seems to be very attractive for treatment of various tumors.
There are various hypotheses on reduced cancer risk in subjects who consume fruits and vegetables rich in polyphenols/flavonoids. The health-beneficial effects of flavonoids, such as quercetin (QU), could be attributed to anti-cancer, anti-oxidative, anti-inflammatory, and anti-proliferative activities that are associated with the inhibition of bioactive enzymes and induction of detoxification enzymes [
9,
13]. However, cellular effects of these polyphenolic compounds are poorly understood. Along with protecting cells from oxidative stress, flavonoids can promote apoptosis and genotoxicity of tumor cells through pro-oxidant activities [
14,
15]. In addition to the pro-oxidative action and the initiation of apoptosis, other anti-tumorigenic mechanisms of QU action include inhibition of glycolysis and enzyme synthesis, interaction with estrogen type II binding sites, and modulation of numerous molecular targets [
16,
17,
18]. The antitumor activity of QU has been investigated in different types of cancers including breast, colon, gastric, leukemia, melanoma, head and neck, and bladder cells [
13,
16,
17,
18,
19].
In addition to anti-tumor effects, QU acts as a hyperthermic sensitizer in HeLa cells, melanoma cells, and lymphoid-leukemia and ovarian cell lines, by inducing intracellular lactate accumulation and acidification. QU also inhibits tumor growth and amplifies the effects of hypothermia in PC-3 and DU-145 prostate tumor cells [
20]. Furthermore, QU reduces temperature-dependent expression of heat-shock protein-70 (HSP-70) by downregulating heat-shock-transcription factor 1 (HSF-1) in vitro [
21,
22,
23], and in vivo, as shown in colon carcinoma and Ehrlich ascites tumor (EAT) in mice [
9,
24]. Of note, several studies have demonstrated that hyperthermia may be useful in overcoming resistance to chemotherapeutic drugs such as CP by increased drug uptake, changing in the tumor microcirculation, blood flow, cell membrane permeability, and cellular metabolism [
9,
11,
19]. CP is the most widely used chemotherapeutic with numerous side effects, including hepatorenal toxicity, cardiotoxicity, neurotoxicity, ototoxicity, and changes of hematological parameters [
9]. It is applied intraperitoneally for bladder, cervical, ovarian cancer, gastric cancer, non–small cell lung cancer, mesothelioma, and head-and-neck squamous cell carcinoma. Cisplatin enters into cell by binding to DNA to form a DNA adduct that interferes with DNA transcription and replication. As a consequence, it abolishes the cell cycle, leading to activation of DNA repair mechanism and induction of apoptosis. In addition, CP can cause immunogenic cell death which is further enhanced by hyperthermia [
9,
11].
Therefore, QU has received much attention as a promising anticancer compound with little toxicity, but also as an adjuvant to the therapy capable to reduce CP-induced toxicity and sensitize cells to hyperthermia by antagonizing the induction of the heat shock response and ability to increase immunomodulatory activity [
9,
20]. However, it was also reported that low concentrations of QU may attenuate effects of CP and other anti-neoplastic drugs in ovarian cancer cells by reducing reactive oxygen species (ROS)-induced damage [
25].
Moreover, some of the mechanisms of QU action are very poorly defined. In particular, the differences between its in vitro and in vivo activities, the efficacy of multimodal approach in the therapy of tumors in combination with chemotherapy and hyperthermia, mode of QU interactions with chemotherapeutics and hyperthermia, as well as advantages and disadvantages of QU application in different models.
Hence, to better understand the efficacy of the combined treatment with QU and CP and effects of QU/CP interactions on tumor growth, we performed a multimodal study in in vitro and in vivo systems under physiological and hyperthermic conditions, which mimic intraoperative hyperthermic intraperitoneal chemotherapy (HIPEC) condition when the tumor is in an advanced stage of growth.
2. Results
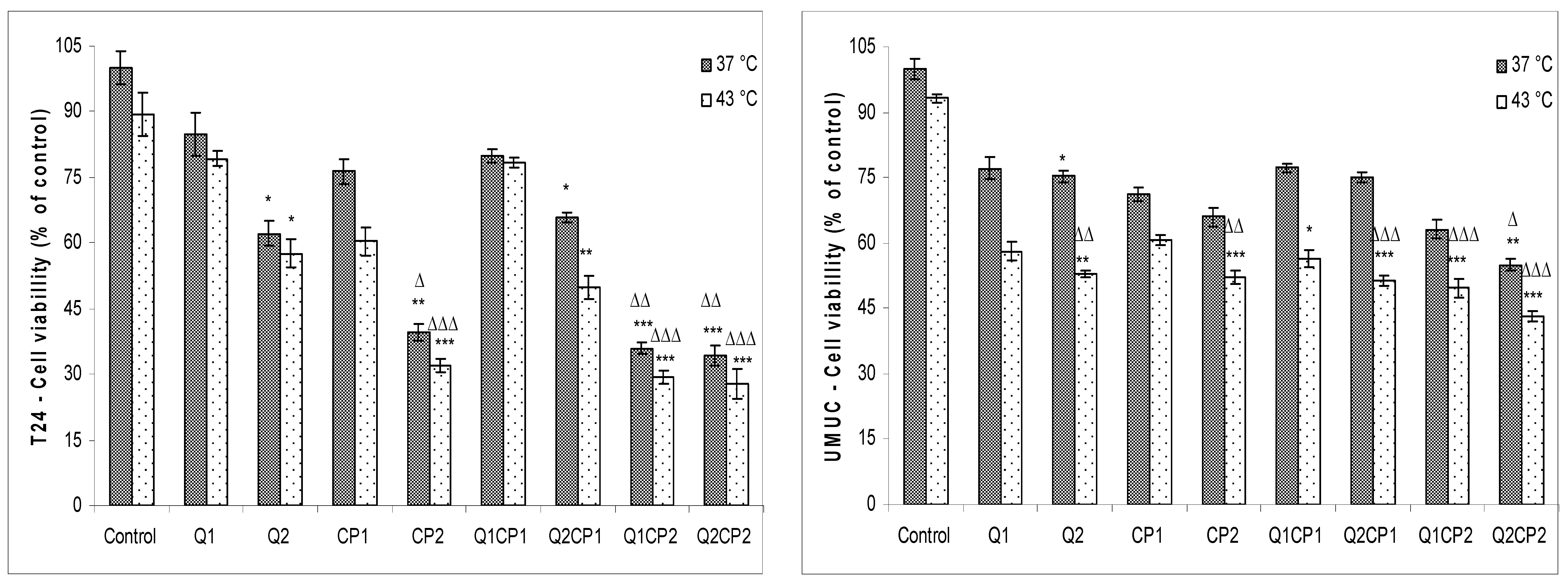
To examine the effects of QU and CP on T24 and UMUC human bladder cancer cells under physiological and hyperthermic conditions, exponentially growing cells were treated with 1 or 50 µM QU (QU1 or QU2) and incubated for 2 h, washed and treated with or without 1 or 50 µM CP (CP1 or CP2) for 1 h under physiological (37 °C) and hyperthermic conditions (43 °C), respectively. As determined by 3-(4,5-dimethylthiazol2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay, treatment with QU and/or CP reduced viability in both cell lines, although higher sensitivity was observed for T24 cells (
Figure 1).
Hyperthermia in both cell lines additionally reduced the survival rate up to 10% and caused very low sensitization to CP. Again, the effect was more pronounced in T24 than UMUC cell line (
Figure 1). There were no significant differences in the percentage of cell viability (MTT test) under physiological and hyperthermic conditions for T24 cells treated with QU: percentage of cell viability for Q1 was 84.9 ± 4.97% at 37 °C vs. 79.3 ± 1.55% at 43 °C (
p ˃ 0.05) and for Q2 was 62.2 ± 2.87% at 37 °C compared to 57.67 ± 3.14% at 43 °C (
p ˃ 0.05). Treatment with CP reduced survival of T24 cells to 76.3 ± 2.89% (CP1) or 39.5 ± 1.98% (CP2) at 37 °C, in comparison to 60.4 ± 3.22% (CP1) or 32.1 ± 1.55% (CP2) at 43 °C. The combined treatment (QU1CP2 and QU2CP2) showed a significantly higher effect in relation to control under both condition (37 °C and 43 °C;
p < 0.001), Q2 (
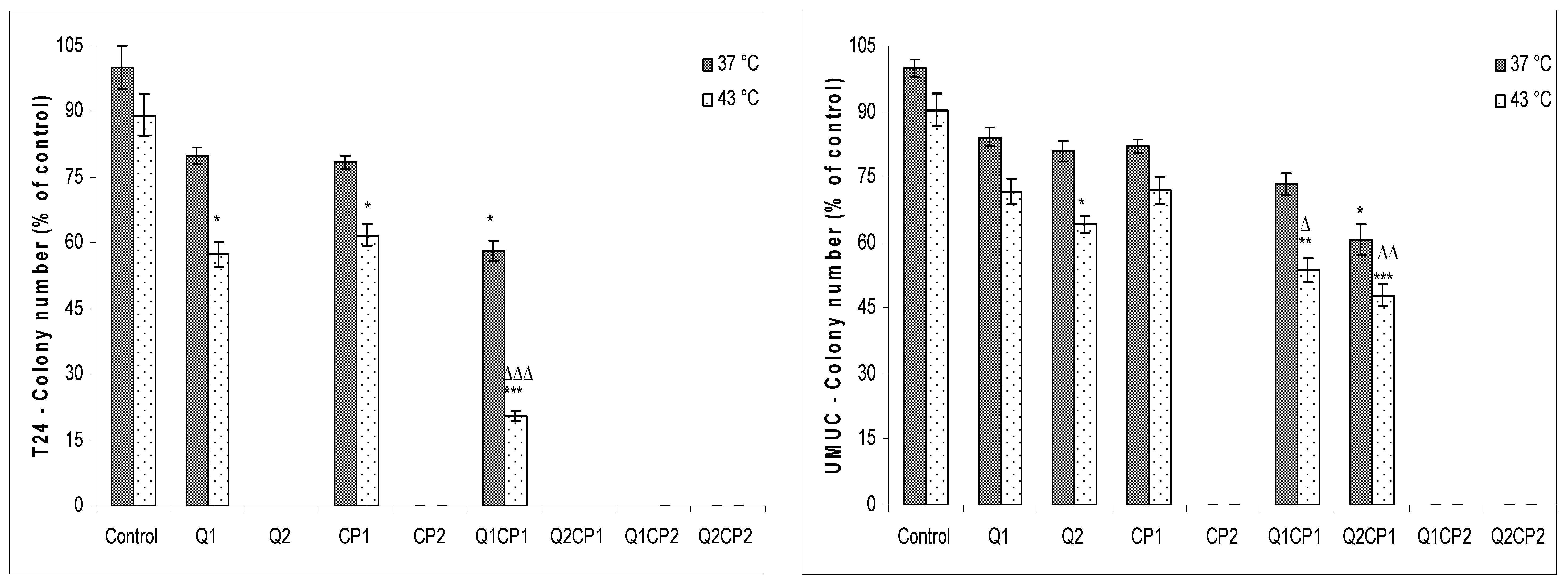
p < 0.05), but not in comparison to CP2. There was no significant difference between the different thermal conditions (37 or 43 °C) in combined treatment. Similar data were obtained for the UMUC human bladder cell line but with lower sensitivity on combined treatment and the different thermal conditions and without differences between applied concentration of QU and CP (1 or 50 µM). Apart from the results obtained with MTT assay, QU and CP showed even higher ability to reduce cell clonogenesis (
Figure 2).
Cell clonogenesity was significantly inhibited by hyperthermal treatment in both cell lines (
Figure 2). These data indicated that tested compounds exerted a significant cytotoxic effect in higher concentration on both cell lines and effect was concentration-dependent in T24 cells. Combined treatment with CP and QU in all combinations, except for QU1CP1, had a lethal effect on T24 cells under both, physiological and hyperthermic conditions. It is also evident that treatments with Q2 and CP2 alone were lethal for T24 cells. Visual inspection of the plates 14 days after the treatment of UMUC cells, revealed no cells or colonies following exposure to low and high concentration of QU (QU1 and QU2) in the presence of CP2 (QU1CP2 and QU2CP2), under physiological and hyperthermic conditions. Higher concentration of CP also completely prevented colony formation irrespective of the temperature applied.
In order to verify whether the effect of the combined treatment of QU with CP at 37 or 43 °C was due to an antagonistic, additive or synergistic action, combination index values were calculated, as described in the section Materials and Methods and are presented in
Table 1 and
Table 2.
The combination indices (CI) were determined from the data obtained by MTT (
Table 1) and clonogenic assays (
Table 2). The obtained values of CI in MTT analysis for T24 cells suggest that low concentration QU (1 µM) in combination with CP in both concentration induce additive effect under physiological conditions while higher concentration of QU (50 µM) with CP induce synergistic effect under both, physiological and hypertermic conditions (
Table 1). In contrast, both concentrations of QU in combination with CP exert a synergistic effect (
Table 1, UMUC cells).
However, in clonogenic assay, combined effect of QU1CP1 under physiological and hyperthermic conditions may induce additive effect in T24 cells while all other combination were lethal for T24 cells because treatment with 50 µM CP or QU in both condition showed complete lack of cell viability and colony formation, and inability to repair (
Table 2). UMUC cells were also very sensitive to higher concentration of CP (CP2); CP2 combination with QU was lethal to UMUC cells (
Table 1) while combination QU in both concentration with CP1 induce additive effect in hyperthermic conditions.
In Vivo Effects
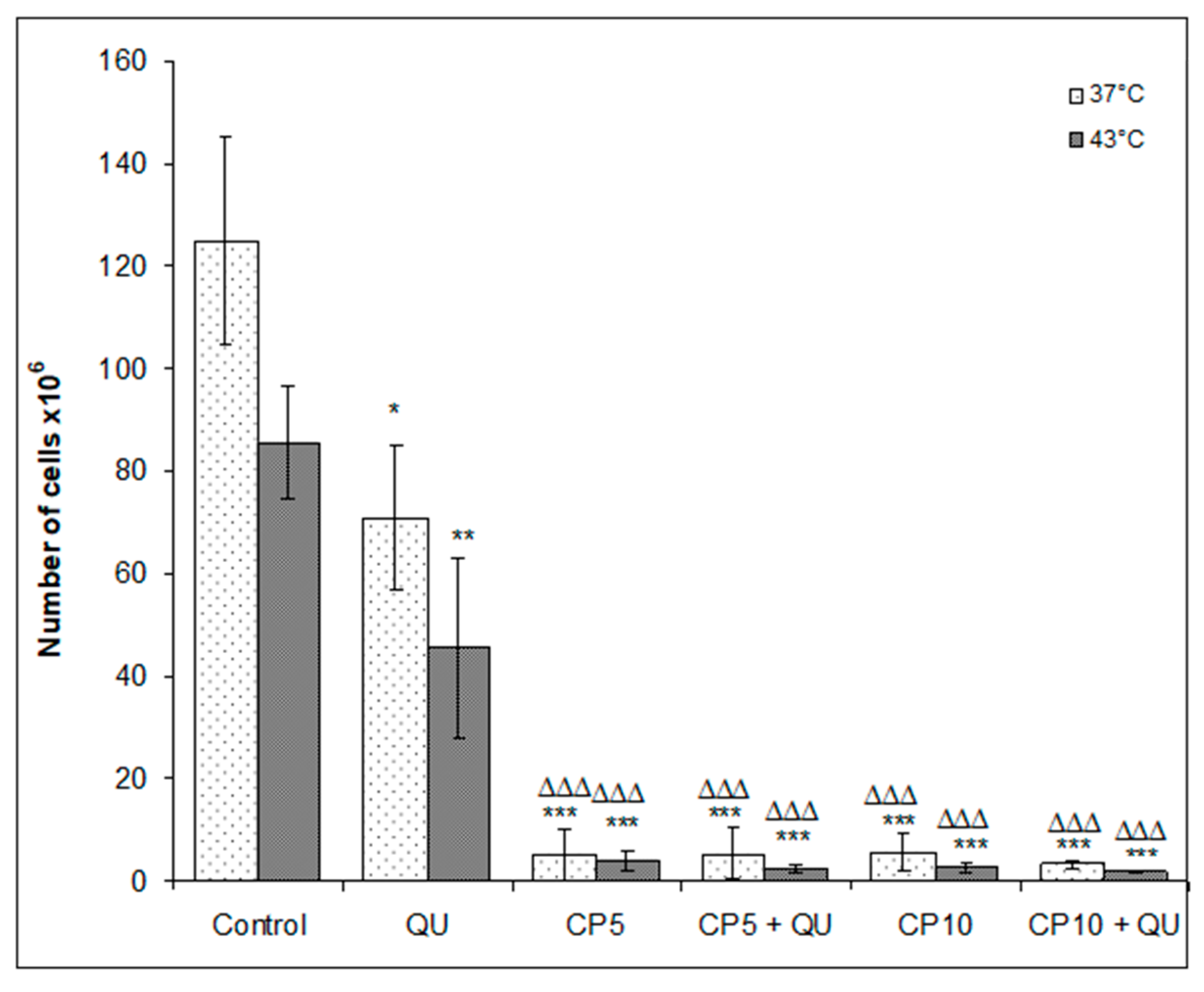
As it is difficult to achieve a specific tumor temperature in the clinic because of the various types of heat conduction due to specific tissue vascularization, we investigated the local effects of QU and CP in vivo on the EAT cells. After disinfection of external abdominal region, each animal was inoculated with 3 mL of saline solution and after gentle agitation of abdominal wall, the solution containing peritoneal cells was removed for cellular evaluation. The following variables were analyzed: the total number of cells and differential count of the cells present in the peritoneal cavity, tumor inhibition (%) and functional activity of macrophages. The remaining animals (n = 9 of each group) were used for the survival analysis.
Animals exposed to intraperitoneal hyperthermia had a significant decrease of the proliferative activity of EAT cells. Numbers of EAT cells in control groups under physiological and hyperthermic conditions were (124.88 ± 25.19) × 10
6 and (85.5 ± 11.23) × 10
6, (
Figure 3), while mean survival time was 20.67 and 24.78 days, respectively (
Table 3).
Treatment of mice with QU alone at a dose of 50 mg kg
−1 reduced the number of cells in peritoneal cavity under both physiological and hyperthermic conditions (TI% was 43.27% and 64.39%), but was without the effect on the survival (
Figure 4 and
Table 3). The very pronounced antitumor effect was achieved after treatment with CP at doses of 5 or 10 mg kg
−1 at 37 and 43 °C, and after treatment with CP in combination with QU (
Figure 3,
Table 3). TI% for CP5 or CP10 and its combination with QU were from 95.56 to 98.61%. In addition to significant reduction of tumor cell number compared to the control (
p < 0.001), these groups also had a significant increase of lifespan (
Table 3 and
Figure 4).
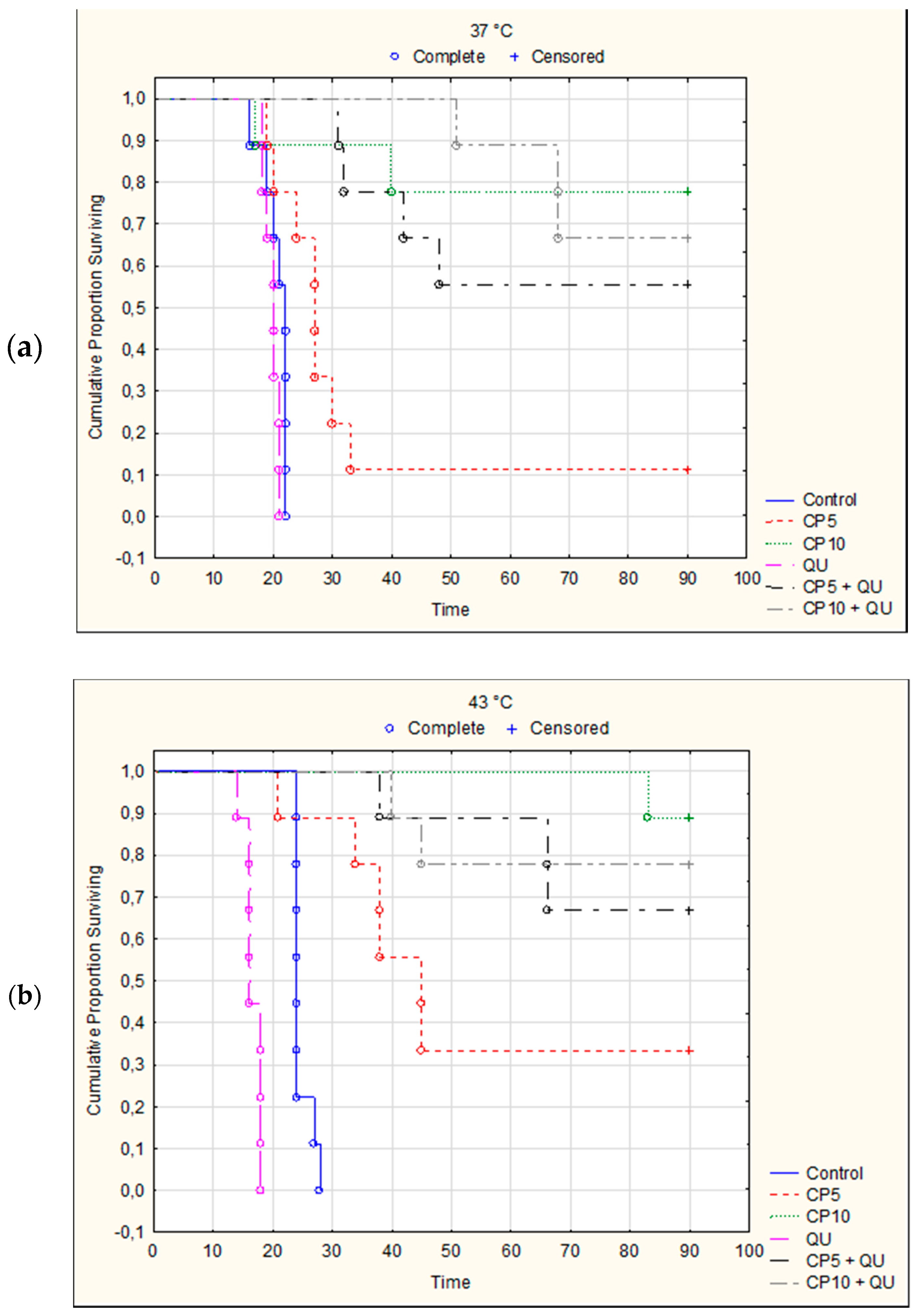
It is important to emphasize that significantly increased survival time was observed after treatment with CP at 37 °C and especially at 43 °C in relation to control group. CP at a dose of 5 mg kg
−1 increased the life span (ILS) at 37 °C for 59.65% and for 163.96% at 43 °C. In the same group, one mouse exposed to physiological and three mice exposed to hyperthermic conditions were long term survivors (
p = 0.0060;
p = 0.0004, Kaplan–Meier analysis). ILS% of mice treated with CP at a dose of 10 mg kg
−1 was 269.28 and 331.64 under physiological and hyperthermic conditions, respectively, whereas seven or eight mice were long term survivors (
p = 0.0060;
p < 0.0001, Kaplan–Meier analysis). Furthermore, significant increase of the survival time was observed after combined treatment with CP5 and QU at 37 °C, in relation to group treated only with CP. Values of ILS% in CP5 + QU group were 224.14 and 59.65, while in CP10 + QU group life span was increased by 290.81% and 269.28% when compared with control group inoculated with EAT cells at 37 °C. Five mice were long-time survivors in CP5 + QU treated group (
p = 0.0001, Kaplan–Meier analysis), while six mice were long-time survivors in CP10 + QU group (
p = 0.0001, Kaplan–Meier analysis). Interestingly, in hyperthermic conditions the difference between the observed increases of the survival in CP5 + QU and CP5 groups was 117.41% (ILS% for CP5 + QU was 281.36 vs. 163.96 in CP5 group), and 6 mice were long-time survivors in CP5 + QU group
(p < 0.0001, Kaplan–Meier analysis). On the contrary, the lifespan in CP10 + QU group was decreased (the difference was negative, −47.32%) in relation to CP10 group (ILS% for CP10 + QU was 284.32 vs. 331.64 in CP10 group). The number of long-time survivors in CP10 and CP10 + QU groups was eight and seven, respectively (
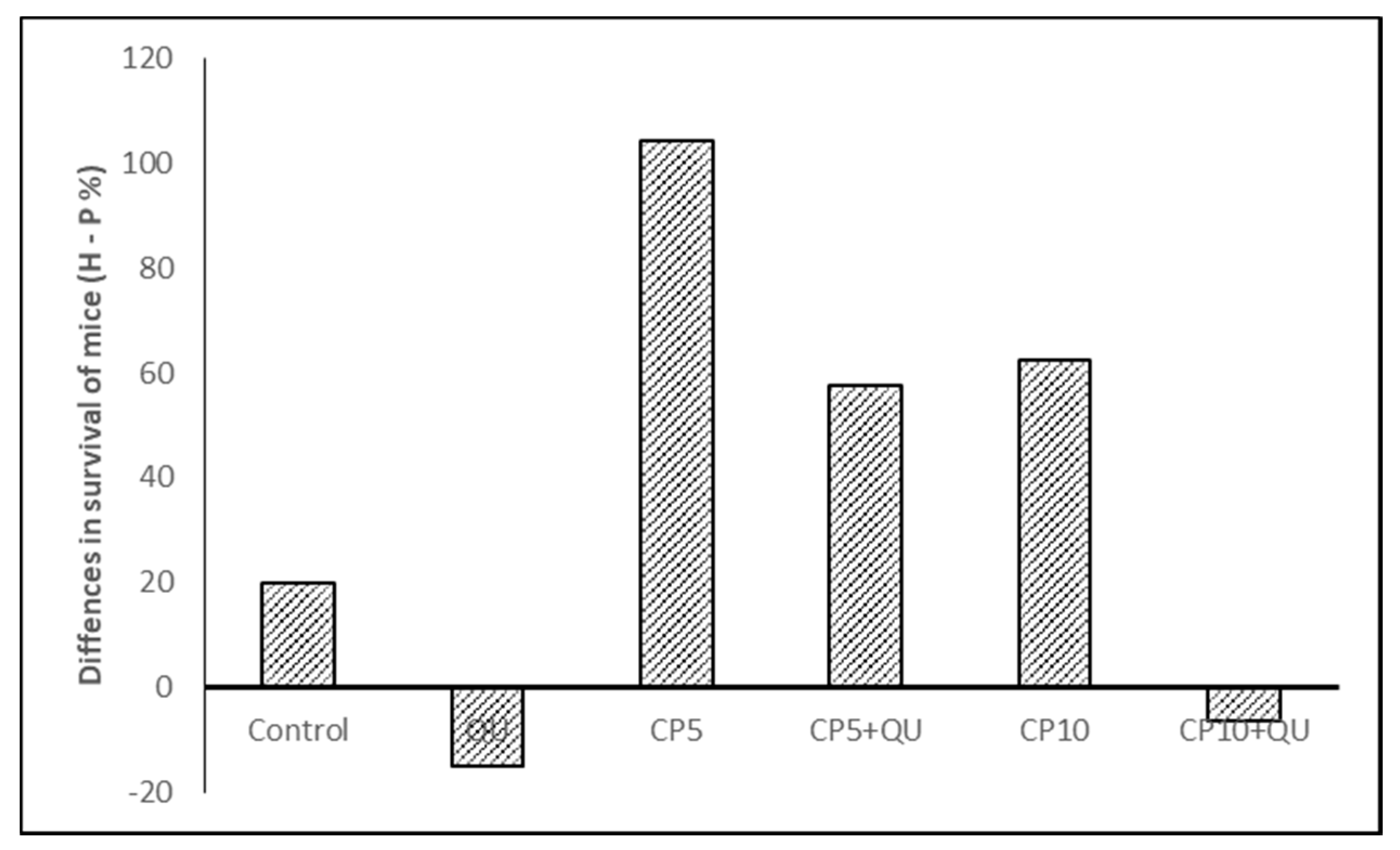
p < 0.0001). Of the nine mice in the CP10 + QU group, two mice died on day 40 and day 45 after implantation due to severe drug toxicity. More exactly, the difference in survival within hyperthermic conditions with respect to physiological conditions (hyperthermic minus physiological treatment) in all experimental groups was as follows: control, 19.88; QU, −15.04; CP5, 104.30; CP5 + QU, 57.52; CP10, 62.36, and CP10 + QU, −6.48, respectively (
Figure 5). Despite the increase in survival rate, there was no statistically significant difference in survival between the corresponding groups treated under physiological conditions compared to hyperthermic conditions, except for treatment with cisplatin at a dose of 5 mg kg
−1.
Combination indices in vivo were calculated from the data obtained by trypan blue (TB) exclusion assay. Analysis of the obtained values suggests that combined treatments CP5 + QU or CP10 + QU may induce synergistic effect (CI = 1.826 ± 0.20 or CI = 2.363 ± 0.13) under physiological and hyperthermic conditions (CI = 1.660 ± 0.23 or CI = 1.857 ± 0.31) (
Table 5).
The analysis was made according to the data from
Figure 4. The combination index was calculated as described in Materials and methods. The combined effects of QU or CP and HT (combination index) was calculated using the formula %AB/(%A × %B), where A and B are the effects of each individual agent and AB is the effect of the combination.
P, statistical significance values of the combination indices (AB) compared with the additive combination index [AB/(A × B)], nonparametric Kruskal–Wallis test,
p < 0.05. Results are reported as the mean ± SD (
n = 6). Abbreviation: QU—treatment with quercetin at a dose of 50 mg kg
−1; CP5 or CP10—treatments with cisplatin at doses of 5 or 10 mg kg
−1.
In addition, during CP and hyperthermia treatment, no significant statistical differences were observed between control and experimental animals regarding behavior and food consumption. No weight loss, diarrhea, skin lesions or other specific abnormalities were noticed.
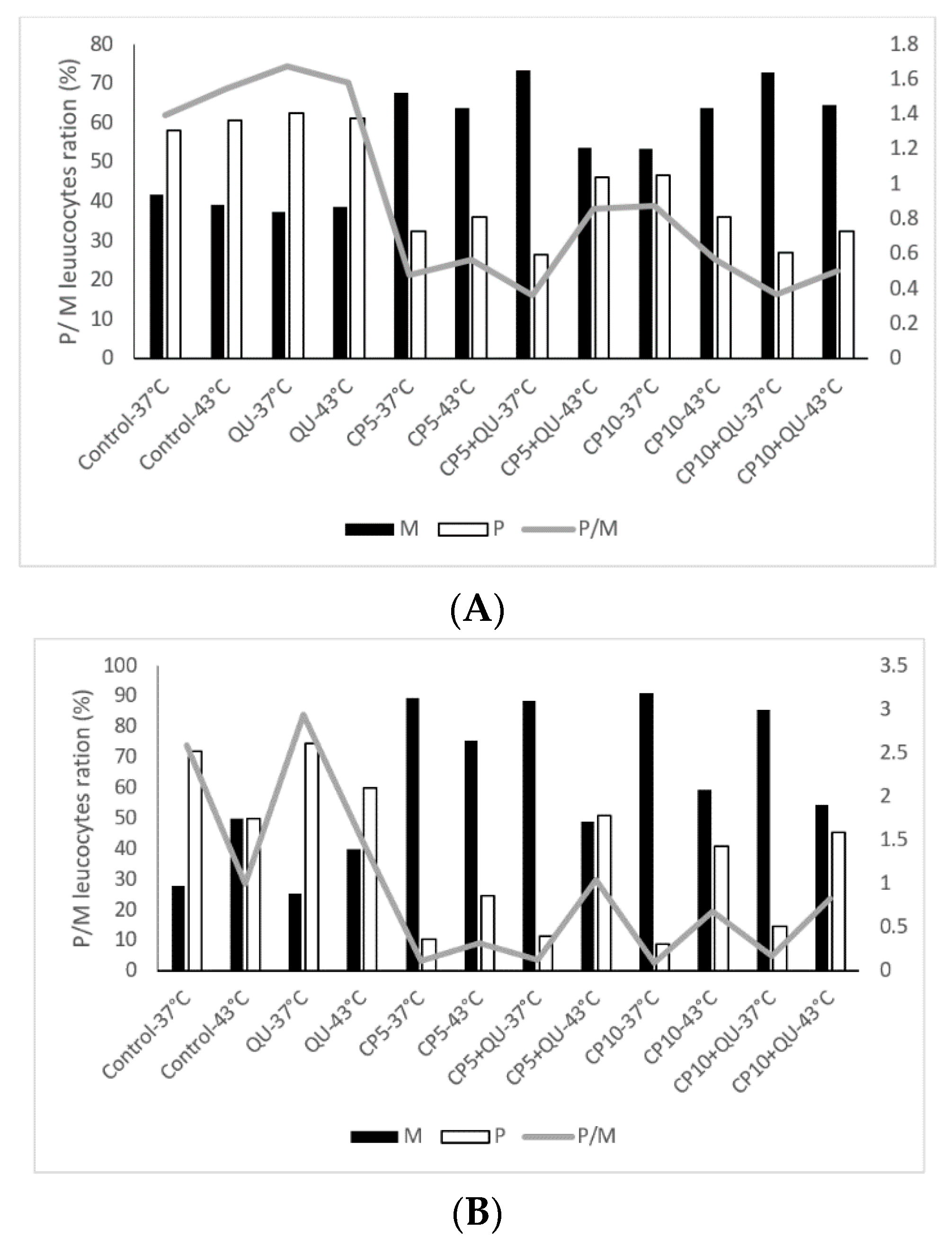
It is known from the literature that many of the anticancer drugs, such as CP, are very toxic. The role of the flavonoids might be important in ameliorating their toxicity on liver, kidney, complete blood count and blood parameters abnormalities. Differential blood analysis clearly demonstrates that hyperthermia in combination with CP changed the ratio between the polymorph nuclear and mononuclear leukocytes (
Figure 6A,B), which coincided with the number of active macrophages in the peritoneal cavity (
Table 6). Macrophage spreading index was increased under hyperthermic conditions (
Table 6).
Finally, hematological and blood parameters were monitored on day 13 and 20 after tumor inoculation. No significant abnormalities in renal (urea, creatinine) and liver biomarkers were observed at day 13 after treatment with CP at both doses (5 or 10 mg kg−1) or during combined treatment with QU (CP + QU). On day 20 after inoculation, we noted improvement in renal and hepatic function (AST, ALT, and LDH) in relation to control group and mice treated only with QU (data not shown), but observed changes did not reach the level of statistical significance (p > 0.05).
3. Discussion
According to previous research, temperatures above 42 °C have shown direct lethal effect on cancer cells in a time- and temperature-dependent manner. These findings were obtained using trypan blue exclusion test of cell viability in vivo and MTT test or clonogenic cell survival assay in vitro [
9,
26,
27,
28]. However, mechanisms underlying the effect of hyperthermia are poorly defined. In particular, these include understanding of the differences between in vitro and in vivo activities, a full potential of multimodal approach in tumor therapy by combining chemotherapy and hyperthermia, mutual interactions with chemotherapeutics, as well as advantages and disadvantages of different models in studying effects of hyperthermia. In our previous study, we investigated possible cytotoxic effects of combined chemoimmunotherapy and hyperthermic intraperitoneal chemotherapy (HIPEC). The focus was on direct and indirect cytotoxic and genotoxic effects and the role of different reparatory and immune system mechanisms. The research was performed using the mammary carcinoma cells (MCa) and EAT cells in vivo. Ehrlich ascites tumor is a suitable transplantable tumor model for studying antitumorous, angiogenic, and anti-inflammatory effects of different drugs and natural compounds. More precisely, this model demonstrates a direct interplay between the cells of immune system, angiogenesis and local tumor growth control [
14].
It is known that hyperthermal treatment can enhance the cytotoxicity of some chemotherapeutic agents including CP, bleomycin, cyclophosphamide, mitomycin C, doxorubicin, and l,3-bis (2-chloroethyl)-nitrosourea [
9,
11,
19,
29].
HIPEC treatment may be enhanced with propolis and various flavonoid compounds such as QU, which are used as sensitizers to increase the effect of hyperthermia and reduce kidney and liver damage caused by cytostatics [
9,
28]. Preventive application of QU has shown a beneficial and synergistic effect as QU acted as sensitizer to hyperthermal treatment in many tumors [
9,
23,
24,
27]. However, the therapeutic potential of QU is quite unknown. According to Li et al. [
25], QU at low concentrations reduced the therapeutic effects of CP and other anti-neoplastic drugs in ovarian cancer cells in vitro. In this study, we investigated the ability of QU to act as sensitizer and to enhance effects of hyperthermia and CP on bladder cancer cells in vitro and EAT cells in vivo.
Our data indicate that hyperthermia alone has a limited effect on T24 and UMUC cells in in vitro conditions. QU at a concentration of 50 µM sensitized these tumor cells in hyperthermia and significantly reduced the viability and clonogenicity (
Figure 1 and
Figure 2). This is consistent with the results obtained in melanoma, colon and HeLa cells, and human acute leukemia blast cells [
23,
28]. Conversely, QU in combination with CP did not additionally sensitize T24 and UMUC cells to heat as revealed by MTT assay in both cell lines. T24 cells are more sensitive to CP and a higher concentration of QU compared to UMUC cells (
Figure 1). However, no statistical (
p ˃ 0.05) difference was observed between the respective groups under physiological and hyperthermic conditions. QU and CP administered at higher concentration (50 µM) caused a lethal effect on T24 cells leading to inability to form colonies, while CP at higher concentration had a lethal effect on UMUC cells (
Figure 2). However, the clonogenic assay showed that treatment of T24 and UMUC cells with high concentrations of QU or CP, as well as their combination, induced significant cell damage that disrupted cell division and colony formation (
Figure 2), possibly due to a higher rate of CP intercalation into the DNA. The increased incorporation of CP into the DNA of the T24 cells was also shown by the lower electrophoretic mobility of the DNA in the agarose gel using a comet assay. Using a comet assay, we observed that the combination of QU and CP at both concentrations under physiological condition reduced the tail length and the amount of DNA in relative to the QU (data not shown). The same observations are described in several papers [
9,
11,
16,
23,
24,
27,
30] with other antioxidants such as vitamin A, C, and E or other flavonoids where the authors suggest that free radicals may participate in the formation of CP-induced DNA-DNA or DNA-protein crosslinks. Under hyperthermic conditions, DNA damage was considerable and difficult to detect.
Obviously, the results of the MTT test, a colorimetric assay for assessing cellular metabolic activity, is less conclusive then the colony formation assay that is based on the ability of a single cell to form a colony. Thus, according to the results of clonogenic assay which is the method of choice for determining cell reproductive death and effectiveness of cytotoxic compounds [
31], it seems that combination of QU and CP can be promising approach in therapy of bladder cancer. Short-term exposure of T24 cells and UMUC cells to QU and CP, which mimics clinical intravesical therapy, may be beneficial for bladder cancer treatment under physiological and hyperthermic conditions. Translation of these facts into in vivo model, may mean, that the local concentration of CP in the bladder would be significantly higher without parallel increase of CP in the serum. In addition to reducing systemic cytotoxicity, hyperthermia may increase chemotherapeutic uptake, and further influences the intracellular distribution and metabolism of drugs, and/or inhibit repair of DNA damage in bladder cancer cells. All these effects lead to an improvement of the effect of chemotherapeutic agents [
8,
9,
10,
11]. Furthermore, QU shows a selective effect; protects normal cells and inhibit proliferation and colony formation of bladder cells [
13,
32]. Furthermore, we also demonstrated that combined treatment with QU or CP and hyperthermia may have an additive, synergistic or lethal effect in vitro (
Table 1 and
Table 2).
In vivo, this multimodal approach exerted the synergistic effect under both temperature related conditions (IC = 1.826 or 2.363 and IC = 1.660 or 1.857 at dose CP5 or CP10) (
Table 5). It seems that additional factors contributed to an increased antitumorogenic effect in vivo, which was manifested through reduced number of cells in the abdominal cavity (
Figure 3), and significantly increased survival of the mice (
Figure 4 and
Table 3). Hyperthermia alone can induce genotoxic effects in vitro and in vivo such as sister chromatid exchanges chromosome aberrations and micronuclei, apoptosis, necrosis, and may damage the plasma membrane and inactivate heat-labile proteins, which lead to protein denaturation and subsequent aggregation [
9,
11,
16,
19]. These effects can change the number of cells in the abdominal cavity leading to increased survival rate of animals (
Figure 4 and
Table 3). On the other hand, hyperthermia treatment increases cytotoxic effect of CP by inducing structural and functional changes in cells. According to obtained results from this study, as well as from our previous data and the data of other authors, treatment with hyperthermia and CP leads to cellular changes and a loss of cellular homeostasis [
9,
23,
33] through different mechanisms: (1) increased accumulation of CP; (2) protein denaturation and aggregation, with consequential cell cycle arrest; (3) disruption of mitotic spindle and enzymes; (4) inactivation of protein synthesis, inhibition of DNA synthesis, transcription, RNA processing and translation; (5) enhanced adduct formation with critical structures; (6) occurrence of different lesions at hyperthermic temperatures and heat-induced inhibition of DNA repair processes; (7) increased degradation of aggregated/misfolded proteins through proteasomal and lysosomal pathways; (8) disruption of the membrane cytoskeleton; (9) effects on tumor oxygenation and tumor micro-environment (e.g., uncoupling of oxidative phosphorylation) that lead to decreased levels of ATP; and (10) alterations in membrane permeability that cause increases in intracellular levels of Na
+, H
+, and Ca
2+ (see [
9,
16]). All of these mechanisms, together with increased active or passive transport across the damaged cell membranes and increased CP solubility and interactions with DNA to form DNA adducts, primarily intra-strand crosslink adducts, contribute to intracellular CP accumulation which culminate in the activation of apoptosis, reduced cell number and increased life span of animals (
Table 2 and
Figure 1) [
9,
23,
24,
27].
Unlike in vitro studies, the in vivo model has an additional advantage, as it depends on various elements of the immune system, thus increasing immune surveillance and offering protection against tumor growth. This model also includes additional mechanisms such as antiestrogenic, antiangiogenic, and pro-apoptotic/necrotic mechanisms, selective destruction of tumor cells in hypoxic and low pH environments, and ability to reverse resistance to certain chemotherapeutic drugs [
16,
20,
28]. According to this and ours previous and literature data, QU interferes with CP by acting on tumor cells through several mechanisms including: (1) increased immunity and direct DNA damage induced by apoptotic processing; (2) microenvironment remodeling to enhance the accumulation and penetration of CP into the tumor site; (3) altered metabolic activity of CP, acting as hyperthermic sensitizer by inducing intracellular lactate accumulation and acidification (increasing lysosomal activity with low pH and heat-induced cell membrane damage); (4) QU also triggered apoptosis and cell cycle arrest in cancer cells to sensitize them to CP chemotherapy; (5) QU enhanced ROS production in tumor cells to activate the intrinsic pathway of apoptosis through the induction of mitochondrial dysfunction and down-regulated anti-apoptotic factors such as Bcl-2 and Bcl-XL while it upregulated apoptotic factor Bax and Bid; (6) QU reversed multidrug resistance; it targets multidrug resistance proteins (MRPs) to inhibit the efflux of CP from cancer cells; (7) QU was capable inhibit HSP27, HSP70, HSP72, and HSP90 and stimulate apoptosis through induction of ER stress and inhibits STAT3 phosphorylation; (8) inhibited angiogenesis; (9) different mechanism which lead to synergistic action QU and CP (QU act on topoisomerase I and II, interact with estrogen binding sites type II and, inhibit different signal pathways involved in cell proliferation while CP act on the formation of DNA adducts and ROS; (10) MiRs, NF-κB, MAPK, AMPK, and HIF are also molecular pathways that are affected by QU with consequential enhancement of the CP chemotherapy; and (11) inhibition of protein kinase δ that enhances cytotoxic effects with CP and hyperthermia while protect kidney cells [
9,
12,
13,
16,
19,
20,
21]. Thus, QU may be included in providing protection against CP mediated renal and liver injury: (1) QU reduces vascular permeability, decreases plasma volume loss after thermal injury and (2) protects against RBC deformity and platelet aggregation, and stabilizes lysosomal membranes [
9,
16,
22,
23,
24,
25,
26,
27]. Antioxidative and antiperoxidative properties of QU, based on free radical scavenger activity, increases the activity of superoxide dismutase, catalase, and glutathione in healthy cells [
9,
16].
We have found that QU increased effect of CP only at lower dose of CP (5 mg kg−1), while higher dose of CP had slightly lower effect as compared to CP10 alone under either condition. Furthermore, treatment of mice with QU alone was ineffective in the advanced stage of tumor growth and the ILS% was negative compared to the control. In general, hyperthermia at 43 °C mainly enhanced the survival of mice when compare to physiological condition. The differences between the ILS% values at 37 and 43 °C were as follows (%): control, 19.88; QU, −15.04; CP5, 104.30; CP5 + QU, 57.52; CP10, 62.36, and CP10 + QU, −6.48, respectively. Despite the increase in survival rate, there was no statistically significant difference in survival between the corresponding groups treated under physiological conditions compared to hyperthermic conditions, except for hyperthermal treatment alone or in combination treatment with cisplatin at a dose of 5 mg kg−1 while QU treatment had a negative effect.
According to this data, it is apparent that higher dose of CP greatly increases the CP’s toxicity under hyperthermic conditions, which is consistent with our previous results regarding the number of micronuclei in peripheral blood reticulocytes in vivo [
28]. Specifically, a lower dose of CP with hyperthermal treatment achieved synergistic anticancer activity and reduced CP toxicity in liver, kidney, and other tissues and organs. This data indicate that under hyperthermic conditions CP alone, or combined with QU, may be the treatment of choice that resembles intraoperative hyperthermic intraperitoneal chemotherapy (HIPEC) or clinical intravesical therapy.
Interestingly, the number of mononuclear leukocyte cells was significantly increased in peripheral bloodstream after treatment with CP, especially at 37 °C, in relation to polymorphonuclear cells. The ratio between mononuclear and polymorphonuclear cells changed in favor of mononuclear leukocyte cells (macrophages and lymphocytes), while the treatment with QU after CP increased the number of neutrophils in the peripheral blood.
The number of active macrophages was also increased in the peritoneal cavity at 43 °C as indicated by the macrophage spreading index (
Table 6). It is possible that an increase of infiltrating monocytes and macrophages in the tumors may be important factor in tumor regression and enhanced survival of the mice [
9,
34,
35,
36]. Tumor regression may be mediated by a complex interaction between the innate and adaptive immunity. The innate mechanism is based on soluble and cellular components that trigger inflammatory events in tumor microenvironment and cytokine production that stimulates dendritic cells and enhances the capacity of peritoneal macrophages to kill tumor cells [
9,
36]. This can be achieved via direct cytotoxic effect or via an indirect cytotoxic activity by “preparing” tumor cells to their own elimination by immune cells such as natural killer (NK) cells or cytotoxic T lymphocytes using a Fast or TNF-related apoptosis-inducing ligand (TRAIL)-dependent pathway [
37]. Furthermore, chemotherapy together with hyperthermia can modify interaction between tumor and dendritic cells, enhance maturation of dendritic cells, and enhance endocytosis of tumor cells by dendritic cells as well as increase the presentation of tumor antigens to T cells, especially by absorption of neoplastic tumor fragments after tumor cell destruction with HIPEC treatment [
37,
38,
39]. In addition, HIPEC treatment increases the sensitivity of tumor cells to activate antitumor effector cells. Thus, hyperthermia may have a direct suppressive effect on tumor cells, or it may act indirectly via enhanced host immune function [
9].
After heat stress, the heat shock proteins (HSPs) can activate tumor immunity by increasing antigen presentation through CD91 receptors, or via toll-like receptors (TLRs). In addition, tumor cells are involved in the production of “danger signals” that are required for dendritic cell maturation and tumor antigen presentation to T cells [
40,
41]. Some authors have also shown that intraperitoneal application of chemotherapeutic agents such as anthracycline (doxorubicin), mitomycin C, or CP may enhance the capacity of peritoneal macrophages to destroy tumor cells [
37,
38,
39]. The underlying mechanisms of CP chemotherapy in advanced tumor stage may be: (1) overcoming the immunosuppression induced by the regulatory T cells (Treg) and myeloid derived suppressor cells (MDSC); (2) enhancing the maturation of dendritic cells and their function by displaying tumor antigens; (3) recruitment and proliferation of effector cells and strengthening the activity of cytotoxic T cells; (4) enhancing the penetration of immune cells into the tumor nucleus; (5) upregulation of MHC class I expression; (6) upregulation of the lytic activity of cytotoxic effectors; and (7) treatment of the cells by anticancer drugs at the temperature of 43 °C that increase tumor cell sensitivity to activate lymphocytes [
9,
37,
38,
39].
According to the mentioned data, the immunogenic effect of hyperthermia together with cisplatin can significantly increase therapeutic efficacy to tumor cells with low normal tissue damage leading to long-term tumor regression and increased animal survival (
Figure 4 and
Table 4 and
Table 6).
EAT is a rapidly growing tumor with aggressive behavior, meaning that the innate immune response is important for controlling its growth, especially the inflammatory response. Thus, the neutrophilic inflammatory response is essential for control of Ehrlich tumor growth. Activated neutrophils may induce tumor destruction by releasing a variety of factors (cytokines, enzymes, chlorinated oxidants, etc.) that are involved in direct tumor destruction, extracellular lysis, inhibition of angiogenesis and activation of other reactive cells, resulting in NK cell, T cell, and antibody-dependent cytotoxicity. However, the high influx of these cells promotes tumor development [
42] as observed in control and QU groups (
Figure 6). This effect is probably related to angiogenesis and releasing of growing factors induced by inflammation that are necessary for tumor development. However, it should be noted that macrophages can exert opposite effects, antitumor as well as pro-tumor activity. In addition to the macrophage-mediated bystander effect initiated by tumor cell apoptosis and the facilitation of antigen recognition, such as phosphatidylserine on the cell surface and enhanced killing of non-apoptotic bystander tumor cells, there are also different tumor-promoting functions of macrophages after chemotherapy that can misdirect macrophage-orchestrated tissue repair response resulting in chemo resistance [
43]. According to that, it seems that chemotherapeutics-induced reduction of the macrophage infiltration into tumors may help in avoiding the pro-tumor activity of macrophages. Therefore, it is very important to investigate the role of macrophage and neutrophils in tumor growth and angiogenesis in relation to tumor microenvironment that affects their polarization and leads to the breakdown of phagocytic and tumoricidal activity of macrophage to pro-tumor activity. It is possible that CP plays a role in immunomodulation, specifically by altering the ratio of M2/M1-like macrophages [
44,
45] and in this way can achieve not only the therapeutic synergism, but also a long-term immunity against tumor growth, leading to increased survival of the animal (
Figure 6).
Furthermore, we have noted the improvement of hepatorenal function after 20 days in relation to the control group and mice treatment only with QU, which is consistent with the increased survival of animals after treatment with CP or combination of CP and QU (
Figure 6).
In conclusion, our results suggest that combined treatment with CP and QU may increase the death of tumor cells under physiological and hyperthermic conditions and could be of clinical relevance in locoregional chemotherapy without systemic toxicity. According to our results, QU in combination with a lower dose of CP in vivo was more effective in anti-cancer therapy than in vitro treatment, especially under hyperthermic condition. HIPEC treatment probably induces local inflammation and recruits effectors of antitumor immunity in vivo, thus increasing cytotoxic effect of CP and resulting in a long-term survival benefit for animals with EAT. The advantages of HIPEC or bladder intracavitary hyperthermic chemotherapy over single chemotherapy can be attributed to the synergy of local hyperthermia treatment, chemotherapy and immunomodulation. It appears that QU could increase sensitivity of chemotherapy and reduce toxicity to healthy cells. However, it should be noted that we can’t exclude the possible role of QU in reducing the toxicity of CP in therapeutic treatment to tumor cells due to possibility of collecting reactive radicals. Moreover, according to the obtained results, further studies are needed to examine the in vivo effects co-administration of quercetin with cisplatin and, the complex network of interactions between macrophages, neutrophils and the activation of effective adaptive responses and chemotherapy treatment in different contexts, including different microenvironment, pH, inflammation, cytokines, tumor immunogenicity, tissue of origin, characteristics of different drugs, dose as well as tumor types.













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}