
Dependence of Biocatalysis on D/H Ratio: Possible Fundamental Differences for High-Level Biological Taxons

 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
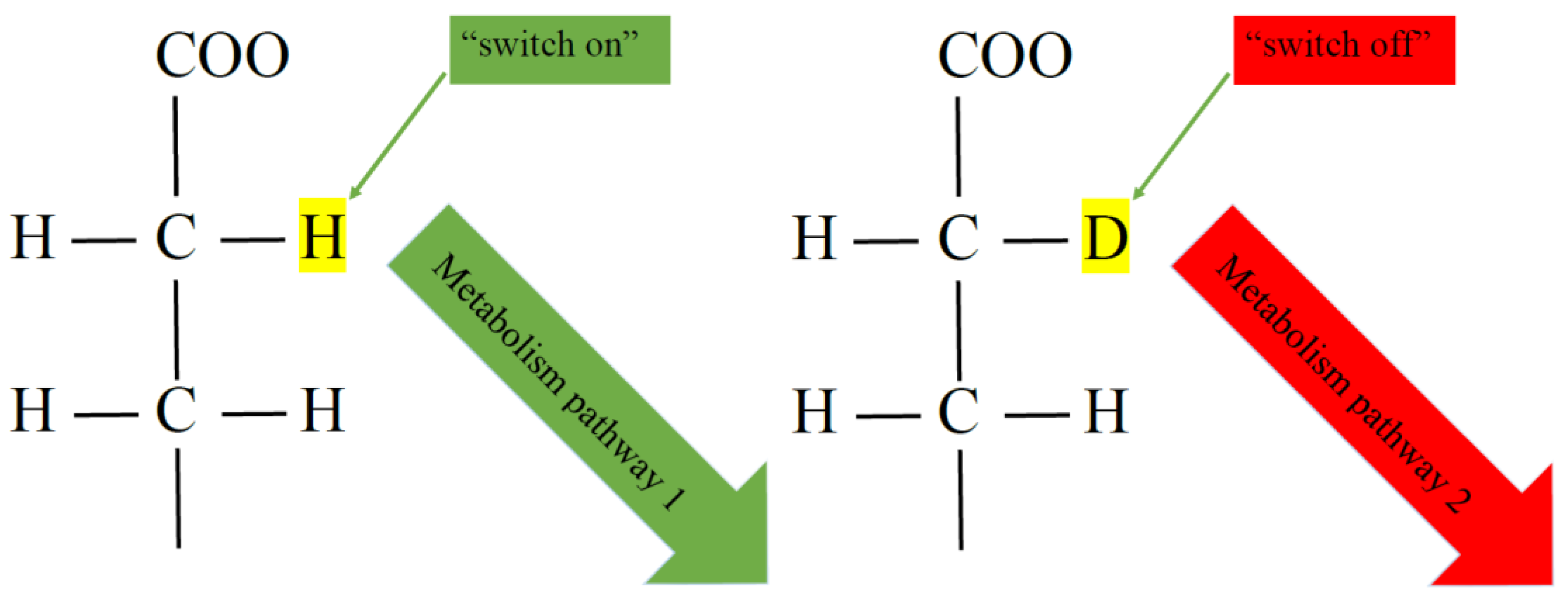
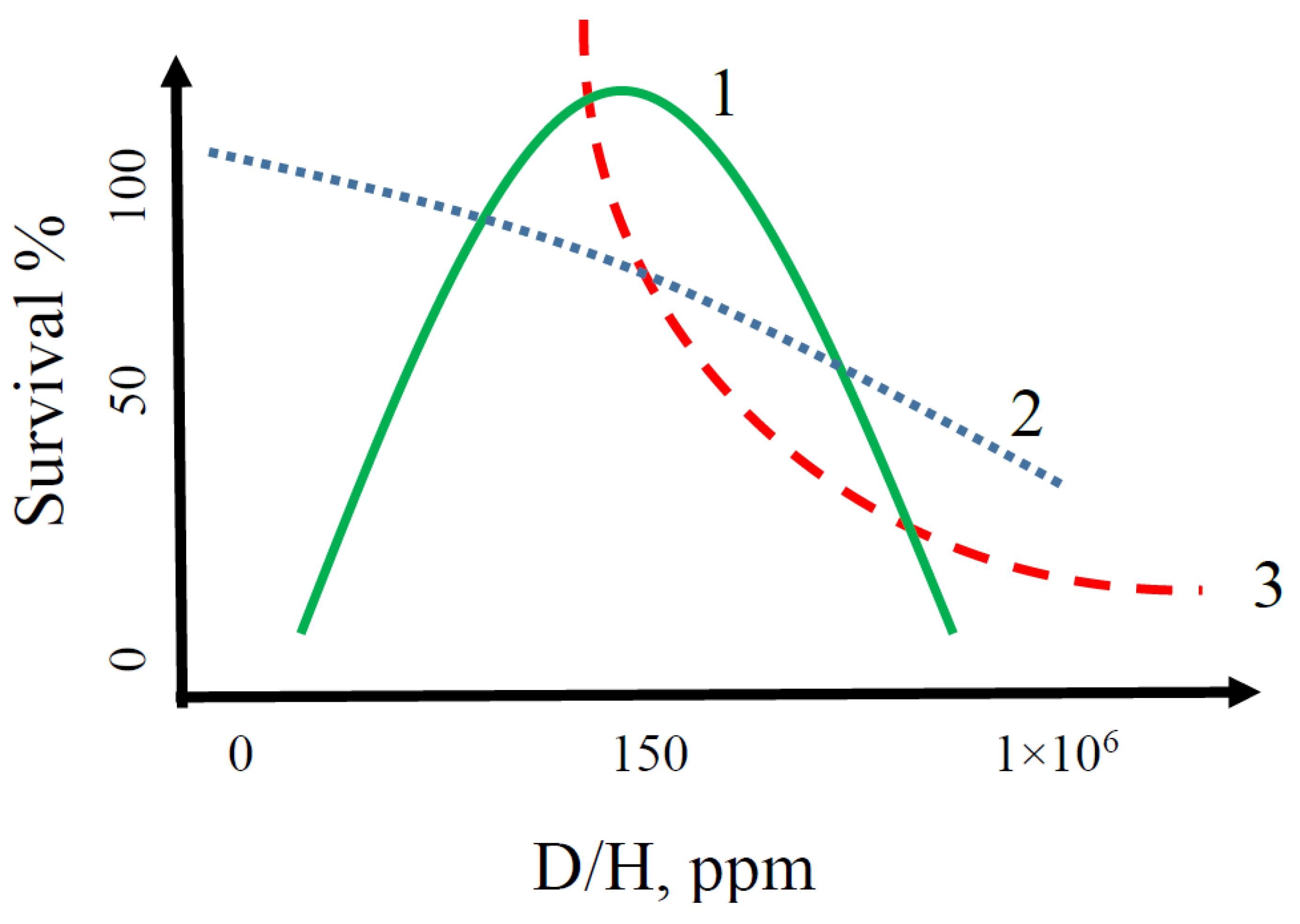
2.1. General Characteristics of Changes in Indicators of Various Taxonomic Groups of Organisms in Water with Different D/H Isotopic Ratios
2.2. Change in CFU of E. Coli in a Medium with Different D/H Isotopic Ratios
2.3. Change in the Accumulation of Minerals during the Germination of C. fragrans L. in Water with Different D/H Isotopic Ratios
2.4. Change in the P. patens Moss Peptidom When Sprouting in Water with Different D/H Isotope Ratios
2.5. Kinetic Model of the Vital Activity of Living Organisms Depending on the D/H Ratio
3. Materials and Methods
3.1. Physico-Chemical Analysis of Water with Different Deuterium Contentss
3.2. Microbiological Methods
3.3. Peptidomics of Moss Physcomitrella Patens
3.3.1. Sprouting of Moss Gametophores
3.3.2. Isolation of Protein from Moss Tissues
3.3.3. 2D Electrophoresis of Proteins
3.3.4. Trypsin Hydrolysis of Proteins
3.3.5. MALDI Mass Spectrometry
3.3.6. Analysis of Mass Spectrometry Results
3.3.7. Liquid Chromatography-Mass Spectrometry (LC-MS/MS)
3.3.8. 2D Electrophoresis of Proteins with Differential Staining (DiGE)
3.4. Germination and Accumulation of Minerals in Callisia fragrans L.
3.4.1. Materials and Method at Germination of C. fragrans L.
3.4.2. Dry Raw Material Analysis
3.5. Determination of Biological Activity Using a Unicellular Biosensor Spirostomum ambiguum
3.6. Determination of Embryotoxicity of Fish
3.7. Ethical Standards of Studies in Animals
3.8. Statistical Data Processing
4. Conclusion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Goncharuk, V.V.; Pleteneva, T.V.; Grebennikova, T.V.; Syroeshkin, A.V.; Uspenskaya, E.V.; Antipova, N.V.; Kovalenko, V.F.; Saprykina, M.N.; Skil’Skaya, M.D.; Zlatskiy, I.A. Determination of Biological Activity of Water Having a Different Isotope Ratio of Protium and Deuterium. J. Water Chem. Technol. 2018, 40, 27–34. [Google Scholar] [CrossRef]
- Lobyshev, V.N.; Kalinichenko, L.P. Isotopic Effects in Biological Systems; Nauka: Moscow, Russia, 1978. (In Russian) [Google Scholar]
- McCartney, D.; Desbrow, B.; Irwin, C. Post-exercise Ingestion of Carbohydrate, Protein and Water: A Systematic Review and Meta-analysis for Effects on Subsequent Athletic Performance. Sports Med. 2017, 48, 379–408. [Google Scholar] [CrossRef] [PubMed]
- Said, H.M. Intestinal absorption of water-soluble vitamins in health and disease. Biochem. J. 2011, 437, 357–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goncharuk, V.V. Science about Water; Naukova Dumka: Kiev, Ukraine, 2010. (In Russian)
- Basov, A.; Fedulova, L.; Baryshev, M.; Dzhimak, S. Deuterium-Depleted Water Influence on the Isotope 2H/1H Regulation in Body and Individual Adaptation. Nutrients 2019, 11, 1903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowen, G.J.; Winter, D.A.; Spero, H.J.; Zierenberg, R.; Reeder, M.D.; Cerling, T.E.; Ehleringer, J.R. Stable hydrogen and oxygen isotope ratios of bottled waters of the world. Rapid Commun. Mass Spectrom. 2005, 19, 3442–3450. [Google Scholar] [CrossRef]
- Goncharuk, V.V.; Lapshin, V.B.; Burdeynaya, T.N.; Pleteneva, T.V.; Chernopyatko, A.S.; Atamanenko, I.D. Physico-chemical properties and biological activity of water, lean on heavy isotope. Chem. Technol. Water 2011, 33, 15–25. [Google Scholar]
- Lewis, G.N. Biology of heavy water. Nature 1934, 133, 620. [Google Scholar] [CrossRef]
- Timmins, G.S. Deuterated drugs: Where are we now? Expert Opin. Ther. Patents 2014, 24, 1067–1075. [Google Scholar] [CrossRef]
- Boros, L.G.; D’Agostino, D.P.; Katz, H.E.; Roth, J.P.; Meuillet, E.J.; Somlyai, G. Submolecular regulation of cell transformation by deuterium depleting water exchange reactions in the tricarboxylic acid substrate cycle. Med. Hypotheses 2016, 87, 69–74. [Google Scholar] [CrossRef]
- Robins, R.J.; Remaud, G.S.; Billault, I. Natural mechanisms by which deuterium depletion occurs in specific positions in metabolites. Eur. Chem. Bull. 2012, 1, 39–40. [Google Scholar]
- Cleland, W.W. The use of isotope effects to determine enzyme mechanisms. J. Biol. Chem. 2003, 278, 51975–51984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strekalova, T.; Evans, M.; Chernopiatko, A.; Couch, Y.; Costa-Nunes, J.P.; Cespuglio, R.; Chesson, L.; Vignisse, J.; Steinbusch, H.W.; Anthony, D.C.; et al. Deuterium content of water increases depression susceptibility: The potential role of a serotonin-related mechanism. Behav. Brain Res. 2015, 277, 237–244. [Google Scholar] [CrossRef]
- Cărpinişan, L.; Petcu, M.D.; Petrovici, S.; Chiş, C.; Ghişe, A.; Zehan, R. The Influence of deuterium depleted water on the hematocrit and the leukocyte formula in rats intoxicated with chromium. Sci. Pap. Anim. Sci. Biotechnol. 2010, 43, 464–468. [Google Scholar]
- Huynh, M.H.V.; Meyer, T.J. Colossal kinetic isotope effects in proton-coupled electron transfer. Proc. Natl. Acad. Sci. USA 2004, 101, 13138–13141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syroeshkin, A.; Pleteneva, T.; Uspenskaya, E.; Zlatskiy, I.A.; Antipova, N.; Grebennikova, T.; Levitskaya, O.V. D/H control of chemical kinetics in water solutions under low deuterium concentrations. Chem. Eng. J. 2019, 377, 119827. [Google Scholar] [CrossRef]
- Basov, A.; Fedulova, L.; Vasilevskaya, E.R.; Dzhimak, S. Possible Mechanisms of Biological Effects Observed in Living Systems during 2H/1H Isotope Fractionation and Deuterium Interactions with Other Biogenic Isotopes. Molecules 2019, 24, 4101. [Google Scholar] [CrossRef] [Green Version]
- Goncharuk, V.V.; Syroeshkin, A.V.; Zlatskiy, I.A.; Uspenskaya, E.V.; Orekhova, A.V.; Levitskaya, O.V.; Dobrovolskiy, V.I.; Pleteneva, T. Quasi-chemical description of the kinetics of cell death Spirostomum ambiguum biosensor for biological activity of aqueous solutions. J. Water Chem. Technol. 2017, 39, 97–102. [Google Scholar] [CrossRef]
- Atzrodt, J.; Derdau, V.; Kerr, W.J.; Reid, M. Deuterium- and Tritium-Labelled Compounds: Applications in the Life Sciences. Angew. Chem. Int. Ed. 2018, 57, 1758–1784. [Google Scholar] [CrossRef] [Green Version]
- Farthing, D.E.; Buxbaum, N.P.; Lucas, P.J.; Maglakelidze, N.; Oliver, B.; Wang, J.; Hu, K.; Castro, E.; Bare, C.V.; Gress, R.E. Comparing DNA enrichment of proliferating cells following administration of different stable isotopes of heavy water. Sci. Rep. 2017, 7, 4043. [Google Scholar] [CrossRef]
- Syroeshkin, A.; Antipova, N.; Zlatska, A.; Zlatskiy, I.A.; Skylska, M.; Grebennikova, T.; Goncharuk, V. The effect of the deuterium depleted water on the biological activity of the eukaryotic cells. J. Trace Elem. Med. Boil. 2018, 50, 629–633. [Google Scholar] [CrossRef]
- Zlatska, A.V.; Vasyliev, R.G.; Gordiienko, I.M.; Rodnichenko, A.E.; Morozova, M.A.; Vulf, M.A.; Zubov, D.O.; Novikova, S.N.; Litvinova, L.S.; Grebennikova, T.V.; et al. Effect of the deuterium on efficiency and type of adipogenic differentiation of human adipose-derived stem cells in vitro. Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makarova, M.; Syroeshkin, A.; Maksimova, T.; Pleteneva, T.; Zlatskiy, I.; Antipova, N.; Skripnikov, A.; Dzhavakhyan, M. D/H modification of plant peptides and microelements metabolome. FEBS Open Bio 2019, 9, 287. [Google Scholar]
- Tsisanova, E.S.; Uspenskaya, E.V.; Pleteneva, T.V.; Syroeshkin, A.V. Study of biological activity and D/H ratio of water with the aid of cellular biosensor spirostomum ambiguum. Trace Elem. Med. 2010, 11, 85. [Google Scholar]
- Chuparina, E.V.; Gunicheva, T.N. State and issues of X-ray fluorescence analysis of herbal raw materials. Anal. Control 2004, 8, 211–226. [Google Scholar]
- McCluney, K.E.; Sabo, J.L. Tracing Water Sources of Terrestrial Animal Populations with Stable Isotopes: Laboratory Tests with Crickets and Spiders. PLoS ONE 2010, 5, e15696. [Google Scholar] [CrossRef]
- Goncharuk, V.V.; Taranov, V.V.; Kurlyantseva, A.Y.; Syroeshkin, A.V. Phase transition in waters with different content of deuterium. J. Water Chem. Technol. 2015, 37, 219–223. [Google Scholar] [CrossRef]
- Rodin, S.; Rebellato, P.; Lundin, A.; Zubarev, R.A. Isotopic resonance at 370 ppm deuterium negatively affects kinetics of luciferin oxidation by luciferase. Sci. Rep. 2018, 8, 16249. [Google Scholar] [CrossRef] [Green Version]
- Dzhimak, S.; Basov, A.A.; Baryshev, M.G. Content of deuterium in biological fluids and organs: Influence of deuterium depleted water on D/H gradient and the process of adaptation. Dokl. Biochem. Biophys. 2015, 465, 370–373. [Google Scholar] [CrossRef]
- Luo, A.L.; Zheng, Y.L.; Cong, F.S. Research progress of biological effects of deuterium-depleted water. J. Shanghai Jiaotong Univ. (Med. Sci.) 2018, 38, 467–471. [Google Scholar]
- Trouiller, B.; Schaefer, D.G.; Charlot, F.; Nogué, F. MSH2 is essential for the preservation of genome integrity and prevents homeologous recombination in the moss Physcomitrella patens. Nucleic Acids Res. 2006, 34, 232–242. [Google Scholar] [CrossRef]
- Schaefer, D.G. Gene targeting in Physcomitrella patens. Curr. Opin. Plant Biol. 2001, 4, 143–150. [Google Scholar] [CrossRef] [Green Version]
- Mittmann, F.; Brücker, G.; Zeidler, M.; Repp, A.; Abts, T.; Hartmann, E.; Hughes, J. Targeted knockout in Physcomitrella reveals direct actions of phytochrome in the cytoplasm. Proc. Natl. Acad. Sci. USA 2004, 101, 13939–13944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rensing, S.A.; Lang, D.; Zimmer, A.; Terry, A.; Salamov, A.; Shapiro, H.; Nishiyama, T.; Perroud, P.-F.; Lindquist, E.; Kamisugi, Y.; et al. The Physcomitrella Genome Reveals Evolutionary Insights into the Conquest of Land by Plants. Science 2007, 319, 64–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarnighausen, E.; Wurtz, V.; Heintz, D.; Van Dorsselaer, A.; Reski, R. Mapping of the Physcomitrella patens proteome. Phytochemisty 2004, 65, 1589–1607. [Google Scholar] [CrossRef]
- Cho, S.H.; Hoang, Q.T.; Kim, Y.Y.; Shin, H.Y.; Ok, S.H.; Bae, J.M.; Shin, J.S. Proteome analysis of gametophores identified a metallothionein involved in various abiotic stress responses in Physcomitrella patens. Plant Cell Rep. 2006, 25, 475–488. [Google Scholar] [CrossRef]
- Avila, D.S.; Somlyai, G.; Somlyai, I.; Aschner, M. Anti-aging effects of deuterium depletion on Mn-induced toxicity in a C. elegans model. Toxicol. Lett. 2012, 211, 319–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somlyai, G. Defeating Cancer. The Biological Effects of Deuterium Depletion; Author House: Bloomington, Indiana, 2002. [Google Scholar]
- Somlyai, G.; Gyöngyi, Z.; Somlyai, I.; Boros, L.G. Pre-clinical and clinical data confirm the anticancer effect of Deuterium depletion. Eur. J. Integr. Med. 2016, 8, 1–7. [Google Scholar] [CrossRef]
- Zhang, K.; Toki, H.; Fujita, Y.; Ma, M.; Chang, L.; Qu, Y.; Harada, S.; Nemoto, T.; Mizuno-Yasuhira, A.; Yamaguchi, J.-I.; et al. Lack of deuterium isotope effects in the antidepressant effects of (R)-ketamine in a chronic social defeat stress model. Psychopharmacology 2018, 235, 3177–3185. [Google Scholar] [CrossRef]
- Yavari, K.; Kooshesh, L. Deuterium Depleted Water Inhibits the Proliferation of Human MCF7 Breast Cancer Cell Lines by Inducing Cell Cycle Arrest. Nutr. Cancer 2019, 71, 1019–1029. [Google Scholar] [CrossRef]
- Halenova, T.; Zlatskiy, I.A.; Syroeshkin, A.V.; Maximova, T.; Pleteneva, T. Deuterium-Depleted Water as Adjuvant Therapeutic Agent for Treatment of Diet-Induced Obesity in Rats. Molecules 2019, 25, 23. [Google Scholar] [CrossRef] [Green Version]
- Goncharuk, V.V.; Syroeshkin, A.V.; Pleteneva, T.V.; Uspenskaya, E.V.; Levitskaya, O.V.; Tverdislov, V.A. On the possibility of chiral structure-density submillimeter inhomogeneities existing in water. J. Water Chem. Technol. 2017, 39, 319–324. [Google Scholar] [CrossRef]
- Basov, A.A.; Elkina, A.A.; Samkov, A.A.; Volchenko, N.N.; Moiseev, A.V.; Fedulova, L.V.; Baryshev, M.G.; Dzhimak, S.S. Influence of Deuterium-Depleted Water on the Isotope D/H Composition of Liver Tissue and Morphological Development of Rats at Different Periods of Ontogenesis. Iran. Biomed. J. 2019, 23, 129–141. [Google Scholar]
- Rasooli, A.; Fatemi, F.; Hajihosseini, R.; Vaziri, A.; Akbarzadeh, K.; Malayeri, M.R.M.; Dini, S.; Foroutanrad, M. Synergistic effects of deuterium depleted water and Mentha longifolia L. essential oils on sepsis-induced liver injuries through regulation of cyclooxygenase-2. Pharm. Boil. 2019, 57, 125–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasilevskaya, E.R.; Akhremko, A. Proteomic study of pig’s spleen. Potravin. Slovak J. Food Sci. 2019, 13, 314–317. [Google Scholar] [CrossRef]
- Pedersen, L.G.; Bartolotti, L.; Li, L. Deuterium and its role in the machinery of evolution. J. Theor. Boil. 2006, 238, 914–918. [Google Scholar] [CrossRef]
- Xie, X.; Zubarev, R.A. On the Effect of Planetary Stable Isotope Compositions on Growth and Survival of Terrestrial Organisms. PLoS ONE 2017, 12, e0169296. [Google Scholar] [CrossRef]
- Zubarev, R.A. Role of stable isotopes in life—Testing isotopic resonance hypothesis. Genom. Proteom. Bioinform. 2011, 9, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Kutyshenko, V.P.; Iurkevich, D.I. Effect of heavy water on metabolism of a symbiotic organism. Biofizika 2003, 48, 690–700. [Google Scholar]
- Gyöngyi, Z.; Budán, F.; Szabó, I.; Ember, I.; Kiss, I.; Krempels, K.; Somlyai, I.; Somlyai, G. Deuterium Depleted Water Effects on Survival of Lung Cancer Patients and Expression of Kras, Bcl2, and Myc Genes in Mouse Lung. Nutr. Cancer 2013, 65, 240–246. [Google Scholar] [CrossRef] [Green Version]
- Kravtsov, A.A.; Kozin, S.V.; Vasilevskaya, E.R.; Elkina, A.A.; Fedulova, L.V.; Popov, K.; Malyshko, V.V.; Moiseev, A.; Shashkov, D.I.; Baryshev, M.G.; et al. Effect of Drinking Ration with Reduced Deuterium Content on Brain Tissue Prooxidant-Antioxidant Balance in Rats with Acute Hypoxia Model. J. Pharm. Nutr. Sci. 2018, 8, 42–51. [Google Scholar] [CrossRef]
- Basov, A.A.; Kozin, S.V.; Bikov, I.M.; Popov, K.A.; Moiseev, A.V.; Elkina, A.A.; Dzhimak, S.S. Changes in Prooxidant-Antioxidant System Indices in the Blood and Brain of Rats with Modelled Acute Hypoxia which Consumed a Deuterium-Depleted Drinking Diet. Biol. Bull. 2019, 46, 531–535. [Google Scholar] [CrossRef]
- Previs, S.F.; Herath, K.; Nawrocki, A.R.; Rodriguez, C.G.; Slipetz, D.; Singh, S.B.; Kang, L.; Bhat, G.; Roddy, T.P.; Conarello, S.; et al. Using [2H]water to quantify the contribution of de novo palmitate synthesis in plasma: Enabling back-to-back studies. Am. J. Physiol. Metab. 2018, 315, E63–E71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kselíková, V.; Vítová, M.; Bišová, K. Deuterium and its impact on living organisms. Folia Microbiol. 2019, 64, 673–681. [Google Scholar] [CrossRef] [PubMed]
- Zlatska, A.; Gordiienko, I.; Vasyliev, R.; Zubov, D.O.; Gubar, O.; Rodnichenko, A.E.; Syroeshkin, A.; Zlatskiy, I.A. In Vitro Study of Deuterium Effect on Biological Properties of Human Cultured Adipose-Derived Stem Cells. Sci. World J. 2018, 2018, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirali, T.; Serafini, M.; Cargnin, S.; Genazzani, A.A. Applications of Deuterium in Medicinal Chemistry. J. Med. Chem. 2019, 62, 5276–5297. [Google Scholar] [CrossRef]
- Tung, R.D. Deuterium medicinal chemistry comes of age. Future Med. Chem. 2016, 8, 491–494. [Google Scholar] [CrossRef] [Green Version]
- Timmins, G.S. Deuterated drugs; updates and obviousness analysis. Expert Opin. Ther. Patents 2017, 27, 1353–1361. [Google Scholar] [CrossRef]
- Russak, E.; Bednarczyk, E.M. Impact of Deuterium Substitution on the Pharmacokinetics of Pharmaceuticals. Ann. Pharmacother. 2018, 53, 211–216. [Google Scholar] [CrossRef]
- Elison, C.; Rapoport, H.; Laursen, R.; Elliott, H.W.; De Vries, H.; Dreimanis, A. Effect of Deuteration of N—CH3 Group on Potency and Enzymatic N-Demethylation of Morphine. Science 1961, 134, 1078–1079. [Google Scholar] [CrossRef]
- Sipes, I.G.; Gandolfi, A.J.; Pohl, L.R.; Krishna, G.; Brown, B.R. Comparison of the biotransformation and hepatotoxicity of halothane and deuterated halothane. J. Pharmacol. Exp. Ther. 1980, 214, 716–720. [Google Scholar]
- Franco, M.I.; Turin, L.; Mershin, A.; Skoulakis, E.M. Molecular vibration-sensing component in Drosophila melanogaster olfaction. Proc. Natl. Acad. Sci. USA 2011, 108, 3797–3802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basov, A.; Drobotenko, M.; Svidlov, A.; Gerasimenko, E.; Malyshko, V.; Elkina, A.; Baryshev, M.; Dzhimak, S. Inequality in the Frequency of the Open States Occurrence Depends on Single 2H/1H Replacement in DNA. Molecules 2020, 25, 3753. [Google Scholar] [CrossRef] [PubMed]
- Coquery, M.; Villeneuve, J.P. Final Report on the Split Sampling Exercises and Quality Assurance Activities; EU Project Number ENVRUS9602; ICWS: Amsterdam, The Netherlands, 2001; 51p. [Google Scholar]
- Skripnikov, A.Y.; Polyakov, N.B.; Tolcheva, E.V.; Velikodvorskaya, V.V.; Dolgov, S.V.; Demina, I.A.; Rogova, M.A.; Govorun, V.M. Proteome Analysis of the Moss Physcomitrella patens (Hedw.) B.S.G.A. Biochemistry (Moscow) 2009, 74, 480–490. [Google Scholar] [CrossRef] [PubMed]
- O’Farrell, P.H. High resolution two-dimensional electrophoresis of proteins. J. Boil. Chem. 1975, 250, 4007–4021. [Google Scholar]
- Blum, H.; Beier, H.; Gross, H.J. Improved silver staining of plant proteins, RNA and DNA in polyacrylamide gels. Electrophoresis 1987, 8, 93–99. [Google Scholar] [CrossRef]
- Shevchenko, A.; Tomas, H.; Havli, J.; Olsen, J.V.; Mann, M. In-gel digestion for mass spectrometric characterization of proteins and proteomes. Nat. Protoc. 2006, 1, 2856–2860. [Google Scholar] [CrossRef]
- Savitzky, A.; Golay, M.J.E. Smoothing and Differentiation of Data by Simplified Least Squares Procedures. Anal. Chem. 1964, 36, 1627–1639. [Google Scholar] [CrossRef]
- Makarova, M.P.; Syroeshkin, A.V.; Maksimova, T.V.; Matveeva, I.S.; Pleteneva, T.V. Features of the rapid determination of trace elements in medicinal and unofficial plants. Dev. Regist. Med. 2019, 8, 93–97. (In Russian) [Google Scholar]
- Syroeshkin, A.; Uspenskaya, E.; Pleteneva, T.; Morozova, M.; Maksimova, T.; Koldina, A.; Makarova, M.; Levitskaya, O.; Zlatskiy, I. Mechanochemical activation of pharmaceutical substances as a factor for modification of their physical, chemical and biological properties. Int. J. Appl. Pharm. 2019, 11, 118–123. [Google Scholar] [CrossRef]
- Kovalenko, V.F.; Zlatskiy, I.A. Toxicity of chlorophenols in embryogenesis of fishes. Hydrobiol. J. 2019, 55, 87–92. [Google Scholar] [CrossRef]
Sample Availability: The experimental data used to support the findings of this study are available from the corresponding author upon request. |



{kind=link}
{kind=link}
| No. | Physico-Chemical Parameter | Water D/H = 4 ± 0.9 ppm (ddw) | High-Resistance Water of Natural Isotopic Composition D/H = 140 ± 0.9 ppm | D2O 99.9% |
|---|---|---|---|---|
| 1 | Surface Tension σ, mN/m | 75.172 | 72.860 | 67.800 |
| 2 | Kinematic Viscosity, mm2/s | 0.987 | 1.012 | 1.274 |
| 3 | Density *, g/cm3 * O18/O16 = 757 ppm; t° = 25 ± 0.05 °C | 0.9969 | 0.9982 | 1.1042 |
| 4 | Freezing Point, T °C | −1.5 | 0 | +3.8 |
| 5 | Dl—self-diffusion coefficient, 109, m2s−1 | 0.63 | 0.46 | 0.52 |
| 6 | Spin-spin Relaxation Time of the Water Proton t, s | 0.347 | 2.000 | − |
| 7 | Volume concentration * of density inhomogeneities, vc (%) | 0.20 | 1.00 | 0.18 |
| 8 | Obscuration * due to laser light scattering, laserobscuration (λ = 633 nm) | 0.003 | 0.020 | 0.005 |
| D/H, ppm | Organisms, Indicators | ||||
|---|---|---|---|---|---|
| Bacteria | Plants | Animals | |||
| Bryopsida | Magnoliophyta | Protozoa | Metazoa (Fish) | ||
| E.coli, CFU | P. patens, growth | C. fragrans L., accumulation of Zn | S. ambiguum, survival | B. rerio, embryo toxicity (survival) | |
| 10 | 100% | 70% | 4 × 103% | 5% | 80% |
| 140 | 100% | 100% | 100% | 100% | 100% |
| D2O | 35% | 0% * | 0% * | 1% | 50% |
| Element | k | |
|---|---|---|
| Zn-Gly | Zn-Gly in ddw | |
| Zn | 40 ± 10 | 160 ± 10 * |
| Mn | 1 ± 0.3 | 0.7 ± 0.2 |
| Fe | 3 ± 1 | 10 ± 2 * |
| Item No. | Peptide Code and Characterization of the Proteins | Sprouting on MilliQ | Sprouting on DDW |
|---|---|---|---|
| 1 | A9RAW3_PHYPA Uncharacterized protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_158431 PE = 3; SV = 1 | + | − |
| 2 | A9REY5_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_113064 PE = 3; SV = 1 | + | − |
| 3 | A9RFC9_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_113309 PE = 3; SV = 1; D2XNF3 | + | − |
| 4 | A9RJM3_PHYPA NADH-cytochrome b5 reductase OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_115361 PE = 3; SV = 1 | + | − |
| 5 | A9RKT5_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_175733 PE = 4; SV = 1; A9S7I6 | + | − |
| 6 | A9RLI4_PHYPA Predicted protein OS=Physcomitrella patens subsp. patens GN = PHYPADRAFT_203665 PE = 4; SV = 1 | + | − |
| 7 | A9RQ53_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_62373 PE = 4; SV = 1 | + | − |
| 8 | A9SNF4_PHYPA Serine/threonine-protein phosphatase OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_186751 PE = 3; SV = 1; A9S099 | + | − |
| 9 | A9RR01_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_160535 PE = 4; SV = 1 | + | − |
| 10 | A9RWR3_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_206327 PE = 4; SV = 1 | + | − |
| 11 | A9RXH5_PHYPA Predicted protein (Fragment) OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_20993 PE = 4; SV = 1; A9RXH0 | + | − |
| 12 | A9RXS6_PHYPA ATPase ASNA1 homolog OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_206681 PE = 3; SV = 1 | + | − |
| 13 | A9RY14_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_161687 PE = 3; SV = 1 | + | − |
| 14 | A9RZM5_PHYPA Malic enzyme OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_179808 PE = 3; SV = 1 | + | − |
| 15 | A9S3P3_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_123533 PE = 4; SV = 1; Q05KM6 | + | − |
| 16 | A9S3W0_PHYPA Predicted protein (Fragment) OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_5967 PE = 3; SV = 1 | + | − |
| 17 | A9SVS7_PHYPA Predicted protein (Fragment) OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_19427 PE = 4; SV = 1; A9S6D6 | + | − |
| 18 | A9S6F0_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_163089 PE = 4; SV = 1 | + | − |
| 19 | A9SDC7_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_183678 PE = 4; SV = 1 | + | − |
| 20 | A9SE00_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_164448 PE = 4; SV = 1 | + | − |
| 21 | A9SE49_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_211358 PE = 4; SV = 1 | + | − |
| 22 | A9SJI0_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_213039 PE = 3; SV = 1 | + | − |
| 23 | A9SNN0_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_186902 PE = 4; SV = 1 | + | − |
| 24 | A9SPQ9_PHYPA Acyl carrier protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_110052 PE = 3; SV = 1 | + | − |
| 25 | A9SRJ7_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_134103 PE = 4; SV = 1 | + | − |
| 26 | A9SUV8_PHYPA Beta-amylase OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_107034 PE = 3; SV = 1 | + | − |
| 27 | A9SZ47_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_217333 PE = 3; SV = 1 | + | − |
| 28 | A9SZE9_PHYPA Xyloglucan endotransglucosylase/hydrolase OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_137834 PE = 3; SV = 1 | + | − |
| 29 | A9SZY0_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_167354 PE = 4; SV = 1 | + | − |
| 30 | A9T1V0_PHYPA Predicted protein (Fragment) OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_138980 PE = 4; SV = 1 | + | − |
| 31 | A9T469_PHYPA 40S ribosomal protein S8 OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_191463 PE = 3; SV = 1 | + | − |
| 32 | A9T7K4_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_110253 PE = 3; SV = 1 | + | − |
| 33 | A9TE26_PHYPA Cysteine proteinase inhibitor OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_144213 PE = 3; SV = 1 | + | − |
| 34 | A9TF90_PHYPA Predicted protein (Fragment) OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_144707 PE = 4; SV = 1 | + | − |
| 35 | A9THA6_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_222325 PE = 4; SV = 1 | + | − |
| 36 | A9THL7_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_170119 PE = 4; SV = 1 | + | − |
| 37 | A9THY4_PHYPA Predicted protein (Fragment) OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_6064 PE = 4; SV = 1 | + | − |
| 38 | A9TJ89_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_146390 PE = 3; SV = 1 | + | − |
| 39 | A9TLJ0_PHYPA Carboxypeptidase OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_147350 PE = 3; SV = 1 | + | − |
| 40 | A9TMY7_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_223748 PE = 3; SV = 1 | + | − |
| 41 | A9TPH1_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_224194 PE = 4; SV = 1; A9U2L4 | + | − |
| 42 | A9TQB7_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_148985 PE = 3; SV = 1 | + | − |
| 43 | A9TUQ5_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_108826 PE = 4; SV = 1; A9T4Q4 | + | − |
| 44 | Q8GU37_PHYPA Putative phosphatidylcholine-sterol acetyltransferase (Fragment) OS = Physcomitrella patens subsp. patens PE = 2; SV = 1; A9TVL8 | + | − |
| 45 | A9TY79_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_199405 PE = 3; SV = 1 | + | − |
| 46 | A9U536_PHYPA Predicted protein (Fragment) OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_23950 PE = 4; SV = 1; A9U5D9 | + | − |
| 47 | Q1XGA6_PHYPA Cytochrome b OS = Physcomitrella patens subsp. patens GN = cob PE = 3; SV = 1 | + | − |
| 48 | RR19_PHYPA 30S ribosomal protein S19, chloroplastic OS = Physcomitrella patens subsp. patens GN = rps19; PE = 3; SV = 1 | − | + |
| 49 | A9SDA4_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_211044 PE = 3; SV = 1; A9TQF6 | − | + |
| 50 | A9SE53_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_78151 PE = 4 SV = 1; A9SUI0 | − | + |
| 51 | A9RXH3_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_121056 PE = 4; SV = 1 | − | + |
| 52 | A9RQE4_PHYPA 40S ribosomal protein S30 (Fragment) OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_118030 PE = 3; SV = 1; A9RQD4 | − | + |
| 53 | A9SBL7_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_183147 PE = 4; SV = 1 | − | + |
| 54 | A9SF90_PHYPA Predicted protein (Fragment) OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_128900 PE = 4; SV = 1 | − | + |
| 55 | A9T230_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_233069 PE = 1; SV = 1 | − | + |
| 56 | A9SWB0_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_136325 PE = 4; SV = 1 | − | + |
| 57 | A9SUQ5_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_84227 PE = 4; SV = 1 | − | + |
| 58 | A9T6P0_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_168383 PE = 4; SV = 1 | − | + |
| 59 | Q1L642_PHYPA Plasma membrane aquaporin OS = Physcomitrella patens subsp. patens GN = PIP1;1 PE = 3; SV = 1; A9RBK8 | − | + |
| 60 | A9SKG1_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_131352 PE = 4; SV = 1 | − | + |
| 61 | A9RGM6_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_114028 PE = 3; SV = 1 | − | + |
| 62 | RK14_PHYPA 50S ribosomal protein L14, chloroplastic OS = Physcomitrella patens subsp. patens GN = rpl14; PE = 3; SV = 1 | − | + |
| 63 | A9SG27_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_129247 PE = 3; SV = 1; A9RZ43 | − | + |
| 64 | A9TEN6_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_169641 PE = 4; SV = 1 | − | + |
| 65 | A9SZZ7_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_217573 PE = 4; SV = 1 | − | + |
| 66 | A9RS97_PHYPA Predicted protein (Fragment) OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_118507 PE = 4; SV = 1; A9SZB8 | − | + |
| 67 | A9SKW2_PHYPA Peptidylprolyl isomerase (Fragment) OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_49268 PE = 4; SV = 1; A9TGA1 | − | + |
| 68 | A9U1T5_PHYPA Predicted protein (Fragment) OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_5105 PE = 4; SV = 1 | − | + |
| 69 | A9U1Y5_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_173229 PE = 4; SV = 1 | − | + |
| 70 | A9U341_PHYPA Predicted protein (Fragment) OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_8489 PE = 4; SV = 1 | − | + |
| 71 | A9T9M8_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_142289 PE = 3; SV = 1 | − | + |
| 72 | A9U096_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_153718 PE = 4; SV = 1; A9SU96 | − | + |
| 73 | A9T027_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_86316 PE = 3; SV = 1 | − | + |
| 74 | A9TA00_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_220250 PE = 3; SV = 1 | − | + |
| 75 | Q5KSB5_PHYPA Germin-like protein OS = Physcomitrella patens subsp. patens GN = PpGLP6 PE = 2; SV = 1 | − | + |
| 76 | A9S3L2_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_180874 PE = 3; SV = 1 | − | + |
| 77 | A9RGB1_PHYPA Uncharacterized protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_104506 PE = 3; SV = 1 | − | + |
| 78 | A9S1H8_PHYPA Predicted protein (Fragment) OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_44384 PE = 4; SV = 1; A9SQA7 | − | + |
| 79 | A9TNC3_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_196700 PE = 4; SV = 1 | − | + |
| 80 | A9RY38_PHYPA Purple acid phosphatase OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_121352 PE = 3; SV = 1 | − | + |
| 81 | A9TJF9_PHYPA Predicted protein (Fragment) OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_33800 PE = 4; SV = 1 | − | + |
| 82 | A9SKS5_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_186065 PE = 4; SV = 1; A9RPK4 | − | + |
| 83 | A9SIT1_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_165118 PE = 4; SV = 1 | − | + |
| 84 | A9U068_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_227113 PE = 4; SV = 1 | − | + |
| 85 | A9T8V9_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_89614 PE = 4; SV = 1; B7SB99 | − | + |
| 86 | A9TJZ6_PHYPA Predicted protein OS = Physcomitrella patens subsp. patens GN = PHYPADRAFT_146721 PE = 4; SV = 1 | − | + |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zlatskiy, I.; Pleteneva, T.; Skripnikov, A.; Grebennikova, T.; Maksimova, T.; Antipova, N.; Levitskaya, O.; Makarova, M.; Selivanenko, I.; Syroeshkin, A. Dependence of Biocatalysis on D/H Ratio: Possible Fundamental Differences for High-Level Biological Taxons. Molecules 2020, 25, 4173. https://doi.org/10.3390/molecules25184173
Zlatskiy I, Pleteneva T, Skripnikov A, Grebennikova T, Maksimova T, Antipova N, Levitskaya O, Makarova M, Selivanenko I, Syroeshkin A. Dependence of Biocatalysis on D/H Ratio: Possible Fundamental Differences for High-Level Biological Taxons. Molecules. 2020; 25(18):4173. https://doi.org/10.3390/molecules25184173
Chicago/Turabian StyleZlatskiy, Igor, Tatiana Pleteneva, Alexander Skripnikov, Tatiana Grebennikova, Tatiana Maksimova, Nadine Antipova, Olga Levitskaya, Mariia Makarova, Igor Selivanenko, and Anton Syroeshkin. 2020. "Dependence of Biocatalysis on D/H Ratio: Possible Fundamental Differences for High-Level Biological Taxons" Molecules 25, no. 18: 4173. https://doi.org/10.3390/molecules25184173
APA StyleZlatskiy, I., Pleteneva, T., Skripnikov, A., Grebennikova, T., Maksimova, T., Antipova, N., Levitskaya, O., Makarova, M., Selivanenko, I., & Syroeshkin, A. (2020). Dependence of Biocatalysis on D/H Ratio: Possible Fundamental Differences for High-Level Biological Taxons. Molecules, 25(18), 4173. https://doi.org/10.3390/molecules25184173