
A Polyphenols-Rich Extract from Moricandia sinaica Boiss. Exhibits Analgesic, Anti-Inflammatory and Antipyretic Activities In Vivo

 ,
,  , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Phytochemical Analysis
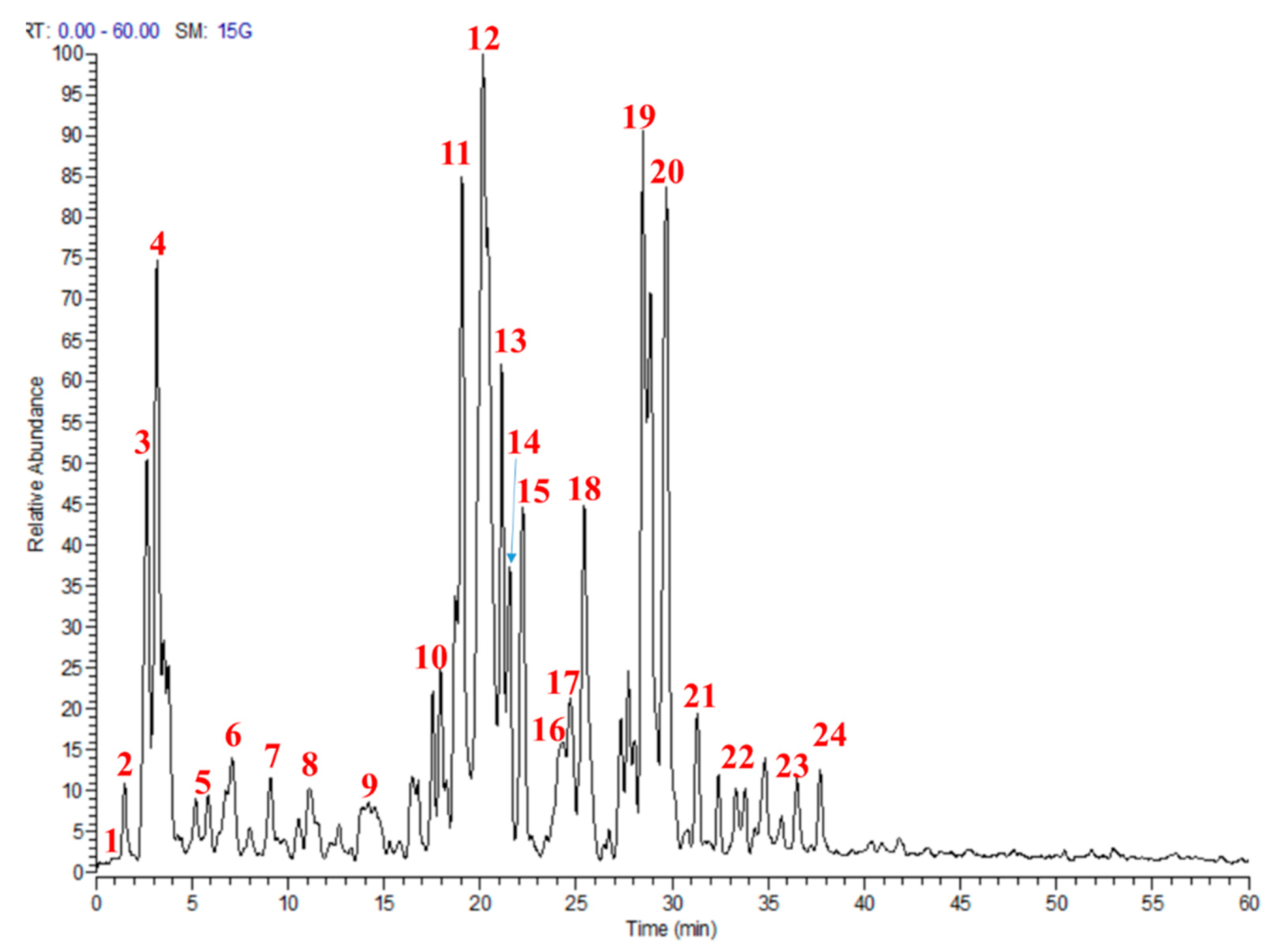
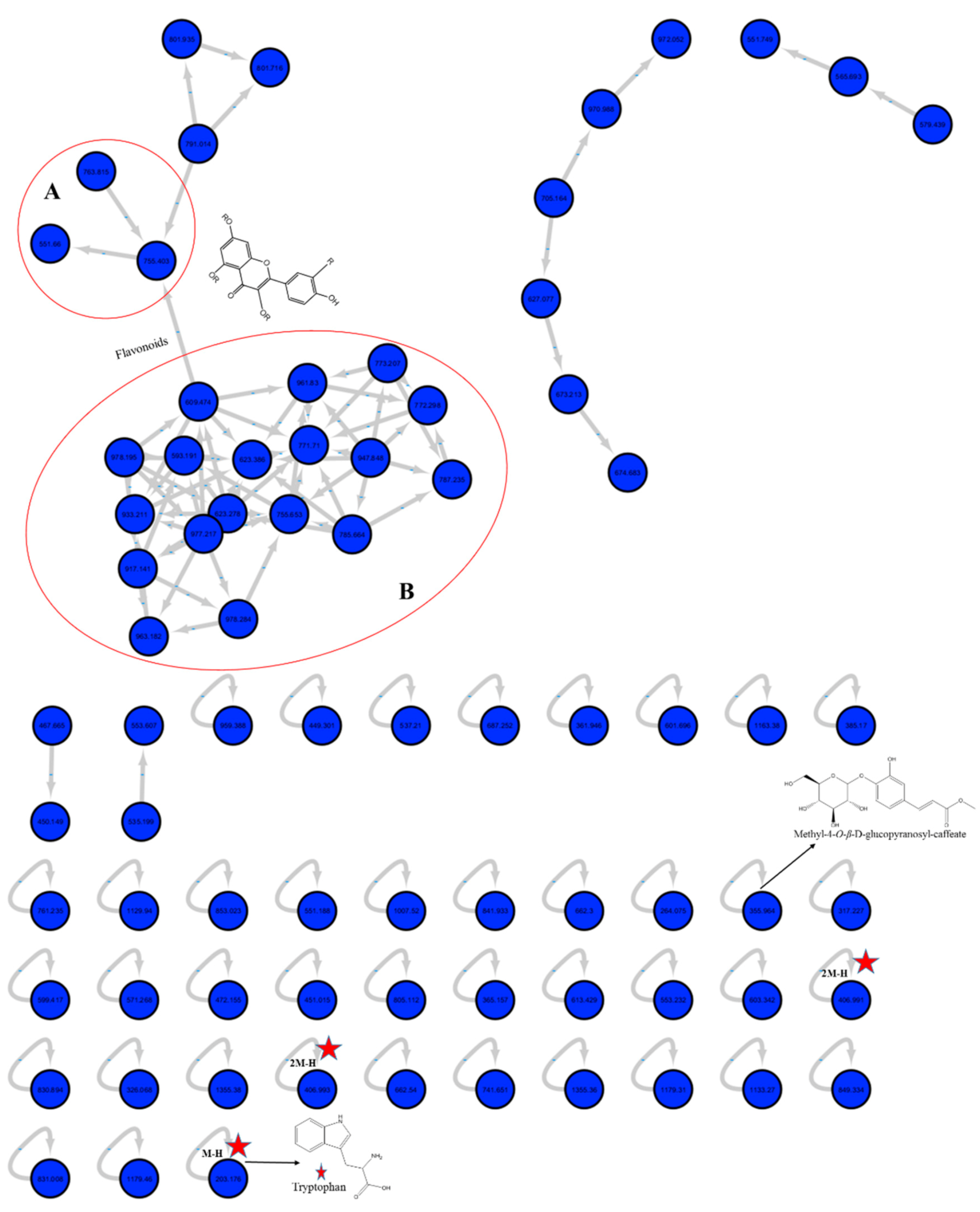
2.1.1. Molecular Networking of M. sinaica Aerial Part (MS-3) Metabolites
2.1.2. LC-MS/MS Metabolites Explanation
2.1.3. 1H-NMR Analysis of the M. sinaica Fraction
2.2. Total Phenolic Content (TPC) and Total Flavonoid Content (TFC)
2.3. Analgesic Effects of M. sinaica Extract and Fractions
2.4. Anti-Inflammatory Activity in Carrageenan-Induced Paw Edema
2.5. Antipyretic Activity in Yeast-Induced Hyperthermia in Mice
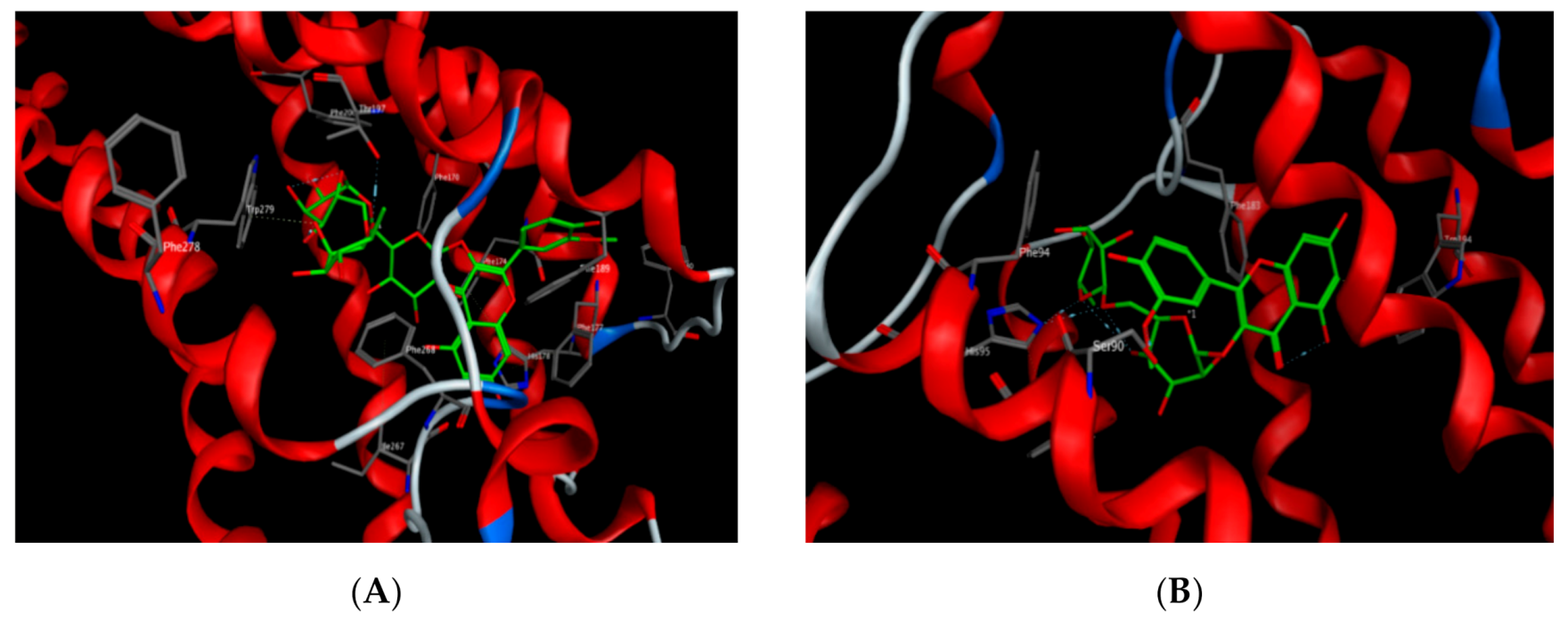
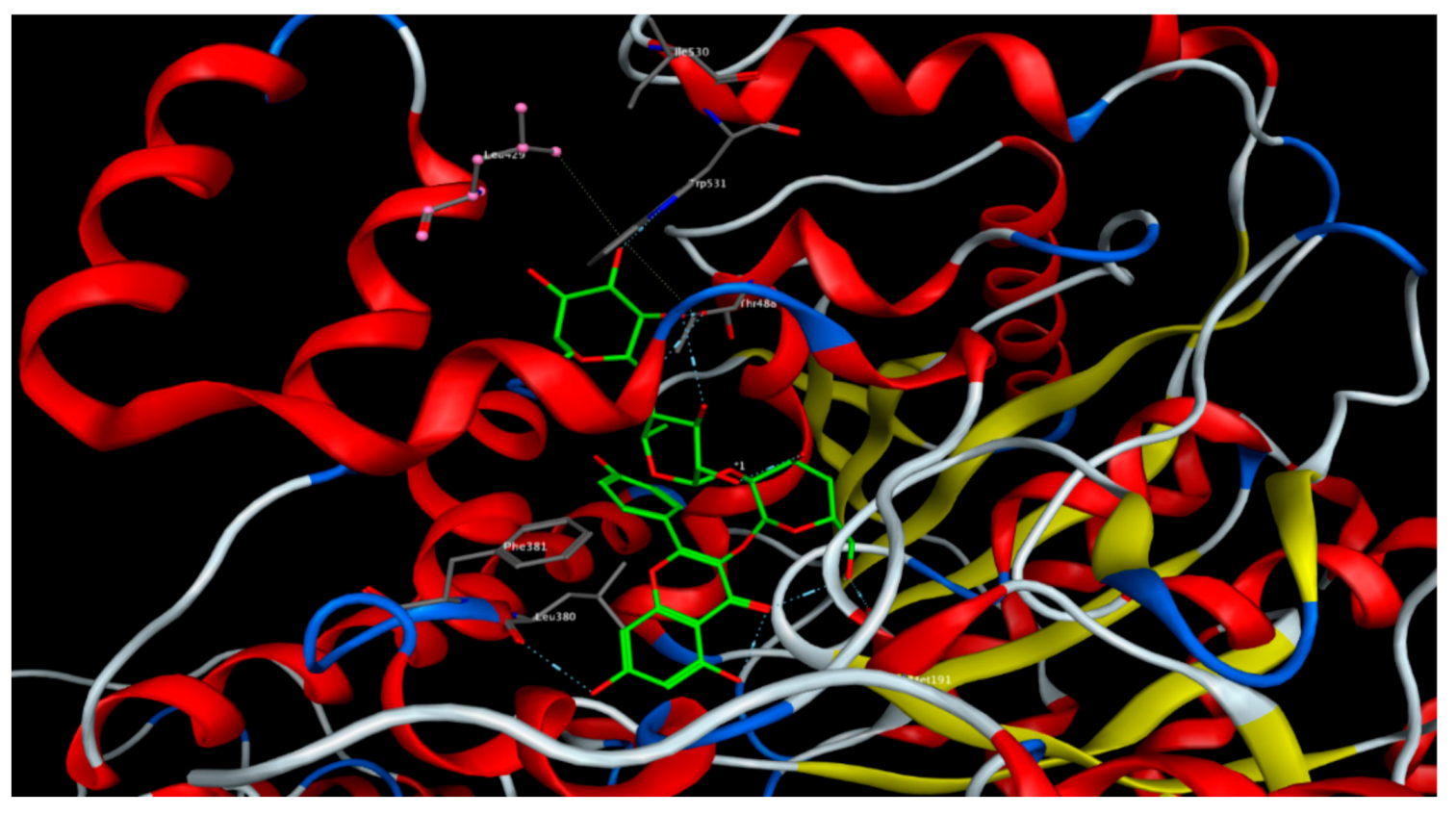
2.6. Molecular Docking
3. Materials and Methods
3.1. Chemicals and Solvents
3.2. Plant Material and Extraction
3.3. Preliminary Qualitative Analysis of the M. sianica Fractions
3.4. HPLC-PDA-MS/MS
3.5. Classical Molecular Networking Workflow Description
3.6. Molecular Modeling
3.7. Total Phenolic Content (TPC) and Total Flavonoid Content (TFC)
3.8. Biological Experiments
3.8.1. Animals
3.8.2. Analgesic Activity
Hot Plate Test
Acetic Acid-Induced Writhing in Mice
number of writhes (control)] × 100.
Tail Flick Method
3.8.3. Anti-Inflammatory Activity in Carrageenan-Induced Paw Edema
3.8.4. Antipyretic Activity
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Soleimanpour, M.; Imani, F.; Safari, S.; Sanaie, S.; Soleimanpour, H.; Ameli, H.; Alavian, S.M. The role of non-steroidal anti-inflammatory drugs (NSAIDs) in the treatment of patients with hepatic disease: A review article. Anesth. Pain Med. 2016, 6, e37822. [Google Scholar] [CrossRef] [Green Version]
- Emam, M.; El Raey, M.; El-Haddad, A.; El Awdan, S.; Rabie, A.-G.; El-Ansari, M.; Sobeh, M.; Osman, S.; Wink, M. A new polyoxygenated flavonol gossypetin-3-o-β-d-robinobioside from Caesalpinia gilliesii (Hook.) D. Dietr. and in vivo hepatoprotective, anti-inflammatory, and anti-ulcer activities of the leaf methanol extract. Molecules 2019, 24, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinarello, C.A. Anti-inflammatory agents: Present and future. Cell 2010, 140, 935–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaughn, S.F.; Berhow, M.A. Glucosinolate hydrolysis products from various plant sources: pH effects, isolation, and purification. Ind. Crops Prod. 2005, 21, 193–202. [Google Scholar] [CrossRef]
- Skandrani, I.; Bouhlel, I.; Limem, I.; Boubaker, J.; Bhouri, W.; Neffati, A.; Sghaier, M.B.; Kilani, S.; Ghedira, K.; Ghedira-Chekir, L. Moricandia arvensis extracts protect against DNA damage, mutagenesis in bacteria system and scavenge the superoxide anion. Toxicol. In Vitro 2009, 23, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Le Floc’H, E.; Floc’h, L.; Floc’h, E.; Boulous, L. Contribution à une Étude Ethnobotanique de la Flore Tunisienne; Ministry of Republic of Tunisia: Tunis, Tunisia, 1983.
- Skandrani, I.; Boubaker, J.; Bhouri, W.; Limem, I.; Kilani, S.; Sghaier, M.B.; Neffati, A.; Bouhlel, I.; Ghedira, K.; Chekir-Ghedira, L. Leaf extracts from Moricandia arvensis promote antiproliferation of human cancer cells, induce apoptosis, and enhance antioxidant activity. Drug Chem. Toxicol. 2010, 33, 20–27. [Google Scholar] [CrossRef]
- Skandrani, I.; Ben Sghaier, M.; Neffati, A.; Boubaker, J.; Bouhlel, I.; Kilani, S.; Mahmoud, A.; Ghedira, K.; Chekir-Ghedira, L. Antigenotoxic and free radical scavenging activities of extracts from Moricandia arvensis. Drug Chem. Toxicol. 2007, 30, 361–382. [Google Scholar] [CrossRef] [PubMed]
- Skandrani, I.; Limem, I.; Neffati, A.; Boubaker, J.; Sghaier, M.B.; Bhouri, W.; Bouhlel, I.; Kilani, S.; Ghedira, K.; Chekir-Ghedira, L. Assessment of phenolic content, free-radical-scavenging capacity genotoxic and anti-genotoxic effect of aqueous extract prepared from Moricandia arvensis leaves. Food Chem. Toxicol. 2010, 48, 710–715. [Google Scholar] [CrossRef]
- Warwick, S.; Francis, A.; Gugel, R. Guide to wild germplasm of Brassica and allied crops (tribe Brassiceae, Brassicaceae), 3rd ed.; Agriculture and Agri-Food Canada: Ottawa, ON, Canada, 2009. [Google Scholar]
- Contreras, M.d.M.; Algieri, F.; Rodriguez-Nogales, A.; Gálvez, J.; Segura-Carretero, A. Phytochemical profiling of anti-inflammatory Lavandula extracts via RP–HPLC–DAD–QTOF–MS and–MS/MS: Assessment of their qualitative and quantitative differences. Electrophoresis 2018, 39, 1284–1293. [Google Scholar] [CrossRef]
- Sobeh, M.; Esmat, A.; Petruk, G.; Abdelfattah, M.A.; Dmirieh, M.; Monti, D.M.; Abdel-Naim, A.B.; Wink, M. Phenolic compounds from Syzygium jambos (Myrtaceae) exhibit distinct antioxidant and hepatoprotective activities in vivo. J. Funct. Foods 2018, 41, 223–231. [Google Scholar] [CrossRef] [Green Version]
- Oszmiański, J.; Kolniak-Ostek, J.; Wojdyło, A. Application of ultra performance liquid chromatography-photodiode detector-quadrupole/time of flight-mass spectrometry (UPLC-PDA-Q/TOF-MS) method for the characterization of phenolic compounds of Lepidium sativum L. sprouts. Eur. Food Res. Technol. 2013, 236, 699–706. [Google Scholar] [CrossRef] [Green Version]
- Su, S.; Cui, W.; Zhou, W.; Duan, J.-A.; Shang, E.; Tang, Y. Chemical fingerprinting and quantitative constituent analysis of Siwu decoction categorized formulae by UPLC-QTOF/MS/MS and HPLC-DAD. Chin. Med. 2013, 8, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marrelli, M.; Morrone, F.; Argentieri, M.P.; Gambacorta, L.; Conforti, F.; Avato, P. Phytochemical and biological profile of Moricandia arvensis (L.) DC.: An inhibitor of pancreatic lipase. Molecules 2018, 23, 2829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, R.N.; Mellon, F.A.; Kroon, P.A. Screening crucifer seeds as sources of specific intact glucosinolates using ion-pair high-performance liquid chromatography negative ion electrospray mass spectrometry. J. Agric. Food Chem. 2004, 52, 428–438. [Google Scholar] [CrossRef]
- Vukics, V.; Guttman, A. Structural characterization of flavonoid glycosides by multi-stage mass spectrometry. Mass Spectrom. Rev. 2010, 29, 1–16. [Google Scholar] [CrossRef]
- Braham, H.; Mighri, Z.; Jannet, H.B.; Matthew, S.; Abreu, P.M. Antioxidant Phenolic Glycosides from Moricandia arvensis. J. Nat. Prod. 2005, 68, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Felder, C.C.; Joyce, K.E.; Briley, E.M.; Glass, M.; Mackie, K.P.; Fahey, K.J.; Cullinan, G.J.; Hunden, D.C.; Johnson, D.W.; Chaney, M.O. LY320135, a novel cannabinoid CB1 receptor antagonist, unmasks coupling of the CB1 receptor to stimulation of cAMP accumulation. J. Pharmacol. Exp. Ther. 1998, 284, 291–297. [Google Scholar] [PubMed]
- Williamson, E.M.; Evans, F.J. Cannabinoids in clinical practice. Drugs 2000, 60, 1303–1314. [Google Scholar] [CrossRef]
- Sharma, C.; Sadek, B.; Goyal, S.N.; Sinha, S.; Kamal, M.A.; Ojha, S. Small molecules from nature targeting G-protein coupled cannabinoid receptors: Potential leads for drug discovery and development. Evid. Based Complement. Alternat. Med. 2015, 2015, 238482. [Google Scholar] [CrossRef] [PubMed]
- Rasouli, H.; Farzaei, M.H.; Khodarahmi, R. Polyphenols and their benefits: A review. Int. J. Food Prop. 2017, 20, 1700–1741. [Google Scholar] [CrossRef] [Green Version]
- Korte, G.; Dreiseitel, A.; Schreier, P.; Oehme, A.; Locher, S.; Hajak, G.; Sand, P.G. An examination of anthocyanins’ and anthocyanidins’ affinity for cannabinoid receptors. J. Med. Food 2009, 12, 1407–1410. [Google Scholar] [CrossRef] [PubMed]
- Korte, G.; Dreiseitel, A.; Schreier, P.; Oehme, A.; Locher, S.; Geiger, S.; Heilmann, J.; Sand, P. Tea catechins’ affinity for human cannabinoid receptors. Phytomedicine 2010, 17, 19–22. [Google Scholar] [CrossRef] [PubMed]
- Tarawneh, A.H.; León, F.; Ibrahim, M.A.; Pettaway, S.; McCurdy, C.R.; Cutler, S.J. Flavanones from Miconia prasina. Phytochem. Lett. 2014, 7, 130–132. [Google Scholar] [CrossRef] [Green Version]
- Thors, L.; Belghiti, M.; Fowler, C. Inhibition of fatty acid amide hydrolase by kaempferol and related naturally occurring flavonoids. Br. J. Pharmacol. 2008, 155, 244–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vane, J.R. Inhibition of prostaglandin synthesis as a mechanism of action for aspirin-like drugs. Nat. New Biol. 1971, 231, 232. [Google Scholar] [CrossRef] [PubMed]
- Cong, H.; Khaziakhmetova, V.; Zigashina, L. Rat paw oedema modeling and NSAIDs: Timing of effects. Int. J. Risk Saf. Med. 2015, 27, S76–S77. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.S.; Mann, M.; DuBois, R.N. The role of cyclooxygenases in inflammation, cancer, and development. Oncogene 1999, 18, 7908–7916. [Google Scholar] [CrossRef] [Green Version]
- Kokate, C.; Purohit, A.; Gokhale, S. Carbohydrate and derived Products, drugs containing glycosides, drugs containing tannins, lipids and protein alkaloids. Text Book Pharmacogn. 2001, 7, 133–166. [Google Scholar]
- Harborne, A. Phytochemical Methods A Guide to Modern Techniques of Plant Analysis, 3rd ed.; Springer: Heidelberg, Germany, 1998. [Google Scholar]
- Tiwari, P.; Kumar, B.; Kaur, M.; Kaur, G.; Kaur, H. Phytochemical screening and extraction: A review. Int. J. Pharm. Sci. 2011, 1, 98–106. [Google Scholar]
- Sobeh, M.; Mahmoud, M.F.; Abdelfattah, M.A.; El-Beshbishy, H.A.; El-Shazly, A.M.; Wink, M. Hepatoprotective and hypoglycemic effects of a tannin rich extract from Ximenia americana var. caffra root. Phytomedicine 2017, 33, 36–42. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatatikun, M.; Chiabchalard, A. Phytochemical screening and free radical scavenging activities of orange baby carrot and carrot (Daucus carota Linn.) root crude extracts. J. Chem. Pharm. Res. 2013, 5, 97–102. [Google Scholar]
- Saboo, S.; Tapadiya, R.; Khadabadi, S.; Deokate, U. In vitro antioxidant activity and total phenolic, flavonoid contents of the crude extracts of Pterospermum acerifolium wild leaves (Sterculiaceae). J. Chem. Pharm. Res. 2010, 2, 417–423. [Google Scholar]
- Kiranmai, M.; Kumar, C.M.; Ibrahim, M. Comparison of total flavanoid content of Azadirachta indica root bark extracts prepared by different methods of extraction. Res. J. Pharm. Biol. Chem. Sci. 2011, 2, 254–261. [Google Scholar]
- Turner, R. The organization of screening. In Screening Methods in Pharmacology, 1st ed.; Elsevier: Cambridge, MA, USA, 1965. [Google Scholar]
- Awe, E.; Adeloye, A.; Idowu, T.; Olajide, O.A.; Makinde, J. Antinociceptive effect of Russelia equisetiformis leave extracts: Identification of its active constituents. Phytomedicine 2008, 15, 301–305. [Google Scholar] [CrossRef]
- D’amour, F.E.; Smith, D.L. A method for determining loss of pain sensation. J. Pharmacol. Exp. Ther. 1941, 72, 74–79. [Google Scholar]
- Winter, C.A.; Risley, E.A.; Nuss, G.W. Carrageenin-induced edema in hind paw of the rat as an assay for antiinflammatory drugs. Proc. Soc. Exp. Biol. Med. 1962, 111, 544–547. [Google Scholar] [CrossRef]
- Loux, J.; DePalma, P.; Yankell, S. Antipyretic testing of aspirin in rats. Toxicol. Appl. Pharmacol. 1972, 22, 672–675. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | tR | (M − H)− | MS/MS | Identified Secondary Metabolites | References |
|---|---|---|---|---|---|
| 1 | 0.92 | 317 | 225, 165, 125 | Galloyl phloretic acid | |
| 2 | 2.66 | 341 | 179 | Caffeoyl glucose | [11] |
| 3 | 3.17 | 372 | 292, 259, 194, 163 | Gluconapin | [16] |
| 4 | 3.53 | 133 | 114.9, 115, 87, 71 | Malic acid | [12] |
| 5 | 5.55 | 385 | 223, 205 | Sinapic acid 3-O-glucoside | [13] |
| 6 | 7.11 | 163 | 147, 119, 106, 72 | p-Coumaric acid | [14] |
| 7 | 9.08 | 263 | 245, 179 | Caffeic acid derivative | |
| 8 | 11.12 | 355 | 338, 309, 193 | Methyl-4-O-β-D-glucopyranosyl-caffeate | [15] |
| 9 | 14.20 | 203 | 186, 159, 143, 116 | Tryptophan | |
| 10 | 17.94 | 771 | 301, 447, 609, 625 | Quercetin 3,4‘-di-O-β-d-glucopyranoside-7-O-α-l-rhamnopyranoside # | [18] |
| 11 | 19.06 | 755 | 285, 447, 609 | Kaempferol-3-O-β-(2″-O-galactosyl)-rutinoside | |
| 12 | 20.15 | 755 | 285, 447, 609 | Kaempferol-3-O-β-(2″-O-glucosyl)-rutinoside * | [15] |
| 13 | 21.13 | 785 | 315, 461, 623, 639 | Isorhamnetin-3-O-(2-glucosyl) rutinoside | |
| 14 | 21.19 | 785 | 315, 461, 623, 639 | Isorhamnetin-3-O-(2-glucosyl) rutinoside | |
| 15 | 22.04 | 947 | 315, 609, 771, 801 | Isorhamnetin-3-O-rutinoside-7-O-diglucoside | |
| 16 | 24.02 | 977 | 315, 639, 771, 785, 831 | Isorhamnetin-3-O-feruloyl glucoside-7-O-diglucoside | |
| 17 | 24.71 | 609 | 179, 301, 447, 463 | Quercetin-3-O-β-glucosyl-7-O-α-rhamnoside * | [15] |
| 18 | 25.41 | 771 | 301, 447, 625 | Quercetin-3-O-β-sophoroside-7-O-α- rhamnoside | [15] |
| 19 | 28.47 | 785 | 315, 461, 639 | Isorhamnetin-3-O-(2-glucosyl) rutinoside | |
| 20 | 29.69 | 623 | 315, 461, 477 | Isorhamnetin 3-O-β-glucopyranoside-7-O-α-rhamnopyranoside | |
| 21 | 31.33 | 977 | 315, 639, 771, 785, 831 | Isorhamnetin-3-O-feruloyl glucoside-7-O-diglucoside | |
| 22 | 32.34 | 947 | 315, 639, 785, 801 | Isorhamnetin 7-O-dicaffeoyl-3-O-rutinoside | |
| 23 | 36.54 | 755 | 285, 593, 431 | Kaempferol-3-O-β-sophoroside-7-O-α-rhamnoside | [15] |
| 24 | 37.71 | 593 | 285, 431, 447 | Kaempferol-3-O-β-glucosyl-7-O-α -rhamnoside | [15] |
| Treatment | Dose (mg/kg) | Reaction Time (Seconds) Pretreatment | Reaction Time (Seconds) Post-Treatment | ||
|---|---|---|---|---|---|
| 30 min | 60 min | 120 min | |||
| MS-1 | 250 | 8.00 ± 0.36 | 7.00 ± 0.44 | 8.16 ± 0.30 | 8.16 ± 0.30 |
| MS-1 | 500 | 7.50 ± 0.42 | 8.16 ± 0.47 | 8.50 ± 0.42 | 8.66 ± 0.42 |
| MS-2 | 250 | 7.66 ± 0.49 | 8.83 ± 0.30 | 9.00 ± 0.36 | 9.50 ± 0.42 * |
| MS-2 | 500 | 6.83 ± 0.30 | 10.00± 0.36 *** | 11.00 ± 0.51 *** | 10.33 ± 0.49 *** |
| MS-3 | 250 | 7.33 ± 0.42 | 9.66 ± 0.49 ** | 10.66 ± 0.49 *** | 10.83 ± 0.30 *** |
| MS-3 | 500 | 7.16 ± 0.30 | 10.50 ± 0.42 *** | 11.83 ± 0.30 *** | 11.66 ± 0.42 *** |
| Indomethacin | 4 | 7.33 ± 0.42 | 12.16 ± 0.47 *** | 14.00 ± 0.36 *** | 14.50 ± 0.42 *** |
| Treatments | Dose (mg/kg) | Number of Writhing in 20 min. | % Inhibition |
|---|---|---|---|
| Control (Acetic acid) | 0.1 mL of 20% | 36.83 ± 1.53 | - |
| MS-1 | 250 | 34.50 ± 1.17 | 6.33 |
| MS-1 | 500 | 36.66 ± 2.21 * | 16.74 |
| MS-2 | 250 | 29.00 ± 1.06 ** | 21.26 |
| MS-2 | 500 | 21.66 ± 0.71 *** | 41.17 |
| MS-3 | 250 | 20.50 ± 1.17 *** | 44.34 |
| MS-3 | 500 | 14.16 ± 0.60 *** | 61.53 |
| Indomethacin | 4 | 6.83 ± 0.60 *** | 81.44 |
| Treatment | Dose (mg/kg) | Pre-Drug | Response Time Duration (Seconds) Post-Drug | ||
|---|---|---|---|---|---|
| 30 m | 60 m | 120 m | |||
| MS-1 | 250 | 3.66 ± 0.33 | 4.16 ± 0.30 | 3.83 ± 0.30 | 4.00 ± 0.36 |
| MS-1 | 500 | 4.16 ± 0.30 | 4.83 ± 0.30 | 5.50 ± 0.22 ** | 4.83 ± 0.40 |
| MS-2 | 250 | 3.83 ± 0.30 | 5.83 ± 0.30 *** | 6.33 ± 0.33 *** | 6.33 ± 0.33 *** |
| MS-2 | 500 | 4.66 ± 0.21 | 6.83 ± 0.30 *** | 7.16 ± 0.40 *** | 7.50 ± 0.56 *** |
| MS-3 | 250 | 4.50 ± 0.22 | 6.50 ± 0.34 *** | 7.33 ± 0.33 *** | 7.5 ± 0.42 *** |
| MS-3 | 500 | 5.16 ± 0.30 | 7.83 ± 0.30 *** | 9.16 ± 0.30 *** | 8.66 ± 0.49 *** |
| Indomethacin | 4 | 4.50 ± 0.42 | 9.33 ± 0.55 *** | 10.50 ± 0.42 *** | 11.00 ± 0.36 *** |
| Extract | Dose | Before Carrageenan | Carrageenan | Change | Inhibition |
|---|---|---|---|---|---|
| (mg/kg) | After 3 h | (%) | |||
| Carrageenan | 0.05 mL of 1% | 0.98 ± 0.03 | 1.57 ± 0.02 | 0.58 ± 0.01 | - |
| MS-1 | 250 | 0.98 ± 0.04 | 1.54 ± 0.02 | 0.56 ± 0.03 | 3.70 |
| MS-1 | 500 | 1.00 ± 0.02 | 1.54 ± 0.01 | 0.53 ± 0.01 | 8.54 |
| MS-2 | 250 | 1.03 ± 0.02 | 1.43 ± 0.01 | 0.40± 0.01 *** | 31.33 |
| MS-2 | 500 | 1.01 ± 0.03 | 1.30 ± 0.02 | 0.29 ± 0.20 *** | 50.42 |
| MS-3 | 250 | 0.95 ± 0.04 | 1.27 ± 0.04 | 0.32 ± 0.01 *** | 45.00 |
| MS-3 | 500 | 0.91 ± 0.02 | 1.19 ± 0.01 | 0.27 ± 0.01 *** | 52.42 |
| Oxyphenbutazone | 100 | 1.00 ± 0.04 | 1.19 ± 0.04 | 0.19 ± 0.01 *** | 66.66 |
| Treatment | Dose (mg/kg) | Normal Rectal Temperature | Rectal Temperature after Yeast Administration | Rectal Temperature °C Post-Treatment | ||
|---|---|---|---|---|---|---|
| 30 min | 60 min | 120 min | ||||
| MS-1 | 250 | 35.3 ± 0.09 | 38.46 ± 0.16 *** | 38.15 ± 0.12 | 38.18 ± 0.15 | 38.10 ± 0.12 * |
| MS-1 | 500 | 35.31 ± 0.10 | 38.21 ± 0.12 *** | 37.88 ± 0.07 * | 37.88 ± 0.09 | 37.98 ± 0.14 |
| MS-2 | 250 | 35.28 ± 0.11 | 38.31 ± 0.17 *** | 37.88 ± 0.14 | 37.43 ± 0.16 ** | 37.50 ± 0.15 ** |
| MS-2 | 500 | 35.46 ± 0.15 | 38.56 ± 0.16 *** | 37.55 ± 0.17 ** | 37.25 ± 0.21 *** | 37.26 ± 0.11 *** |
| MS-3 | 250 | 35.33 ± 0.10 | 38.60 ± 0.15 *** | 37.61 ± 0.13 *** | 37.05 ± 0.09 *** | 36.86 ± 0.14 *** |
| MS-3 | 500 | 35.20 ± 0.12 | 38.50 ± 0.15 *** | 37.46 ± 0.20 ** | 36.56 ± 0.11 *** | 36.35 ± 0.08 *** |
| Indomethacin | 4 | 35.51 ± 0.10 | 38.86 ± 0.10 *** | 36.51 ± 0.19 *** | 36.08 ± 0.08 *** | 35.70 ± 0.10 *** |
| Compound | Scoring Function (kcal/mol) | ||||
|---|---|---|---|---|---|
| CB1 | CB2 | FAAH | COX-1 | COX-2 | |
| Quercetin 3,4‘-di-O-β-d-glucopyranoside-7-O-α-l-rhamnopyranoside | −23.51 | Failed | −23.52 | −25.43 | −23.75 |
| Kaempferol-3-O-β-(2″-O-glucosyl)-rutinoside | −21.46 | Failed | −27.03 | −23.04 | −27.49 |
| Isorhamnetin-3-O-(2-glucosyl) rutinoside | −24.66 | Failed | −27.11 | −21.85 | −27.23 |
| Quercetin-3-O-β-glucosyl-7-O-α-rhamnoside | −22.58 | −17.66 | −26.20 | −24.23 | −23.94 |
| Isorhamnetin 3-O-β-glucopyranoside-7-O-α-rhamnopyranoside | −22.45 | −20.10 | −25.19 | −21.69 | −27.94 |
| Quercetin | −12.02 | −13.80 | −16.04 | −16.18 | −16.07 |
| Kaempferol | −11.20 | −12.76 | −13.51 | −14.90 | −13.05 |
| Isorhamnetin | −12.20 | −12.06 | −15.31 | −14.40 | −15.12 |
| Diclofenac | −10.36 | −12.30 | |||
| AM11542 (CB1 agonist) | −14.22 | ||||
| HU308 (CB2 agonist) | −13.10 | ||||
| Ketobenzimidazole derivative (FAAH inhibitor) | −14.18 | ||||
Sample Availability: Samples of the plant material is available from the authors. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-mekkawy, S.; Shahat, A.A.; Alqahtani, A.S.; Alsaid, M.S.; Abdelfattah, M.A.O.; Ullah, R.; Emam, M.; Yasri, A.; Sobeh, M. A Polyphenols-Rich Extract from Moricandia sinaica Boiss. Exhibits Analgesic, Anti-Inflammatory and Antipyretic Activities In Vivo. Molecules 2020, 25, 5049. https://doi.org/10.3390/molecules25215049
El-mekkawy S, Shahat AA, Alqahtani AS, Alsaid MS, Abdelfattah MAO, Ullah R, Emam M, Yasri A, Sobeh M. A Polyphenols-Rich Extract from Moricandia sinaica Boiss. Exhibits Analgesic, Anti-Inflammatory and Antipyretic Activities In Vivo. Molecules. 2020; 25(21):5049. https://doi.org/10.3390/molecules25215049
Chicago/Turabian StyleEl-mekkawy, Sahar, Abdelaaty A. Shahat, Ali S. Alqahtani, Mansour S. Alsaid, Mohamed A.O. Abdelfattah, Riaz Ullah, Mahmoud Emam, Abdelaziz Yasri, and Mansour Sobeh. 2020. "A Polyphenols-Rich Extract from Moricandia sinaica Boiss. Exhibits Analgesic, Anti-Inflammatory and Antipyretic Activities In Vivo" Molecules 25, no. 21: 5049. https://doi.org/10.3390/molecules25215049
APA StyleEl-mekkawy, S., Shahat, A. A., Alqahtani, A. S., Alsaid, M. S., Abdelfattah, M. A. O., Ullah, R., Emam, M., Yasri, A., & Sobeh, M. (2020). A Polyphenols-Rich Extract from Moricandia sinaica Boiss. Exhibits Analgesic, Anti-Inflammatory and Antipyretic Activities In Vivo. Molecules, 25(21), 5049. https://doi.org/10.3390/molecules25215049