
Pharmacological Effects and Potential Clinical Usefulness of Polyphenols in Benign Prostatic Hyperplasia

 ,
,  , , and
, , and
Abstract
:1. Introduction
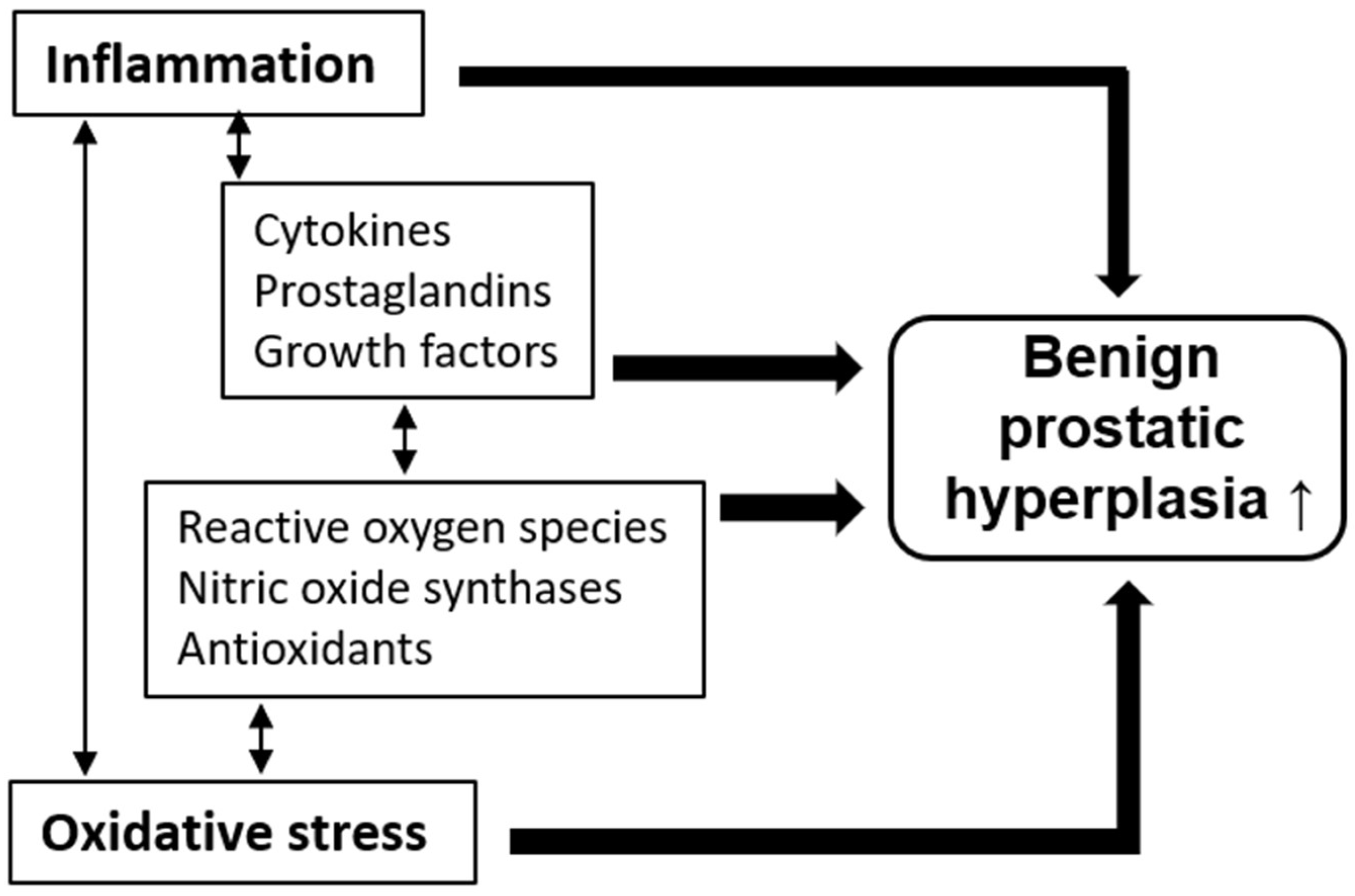
2. A Brief Overview of Inflammation and Oxidative Stress in BPH
2.1. Pathological Roles of Inflammation and Oxidative Stress
2.2. Benign Prostate Hyperplasia and Inflammation
2.3. Benign Prostate Hyperplasia and Oxidative Stress
3. A Brief Overview of Polyphenols in Inflammation and Oxidative Stress
4. Management of Benign Prostate Hyperplasia with Polyphenols
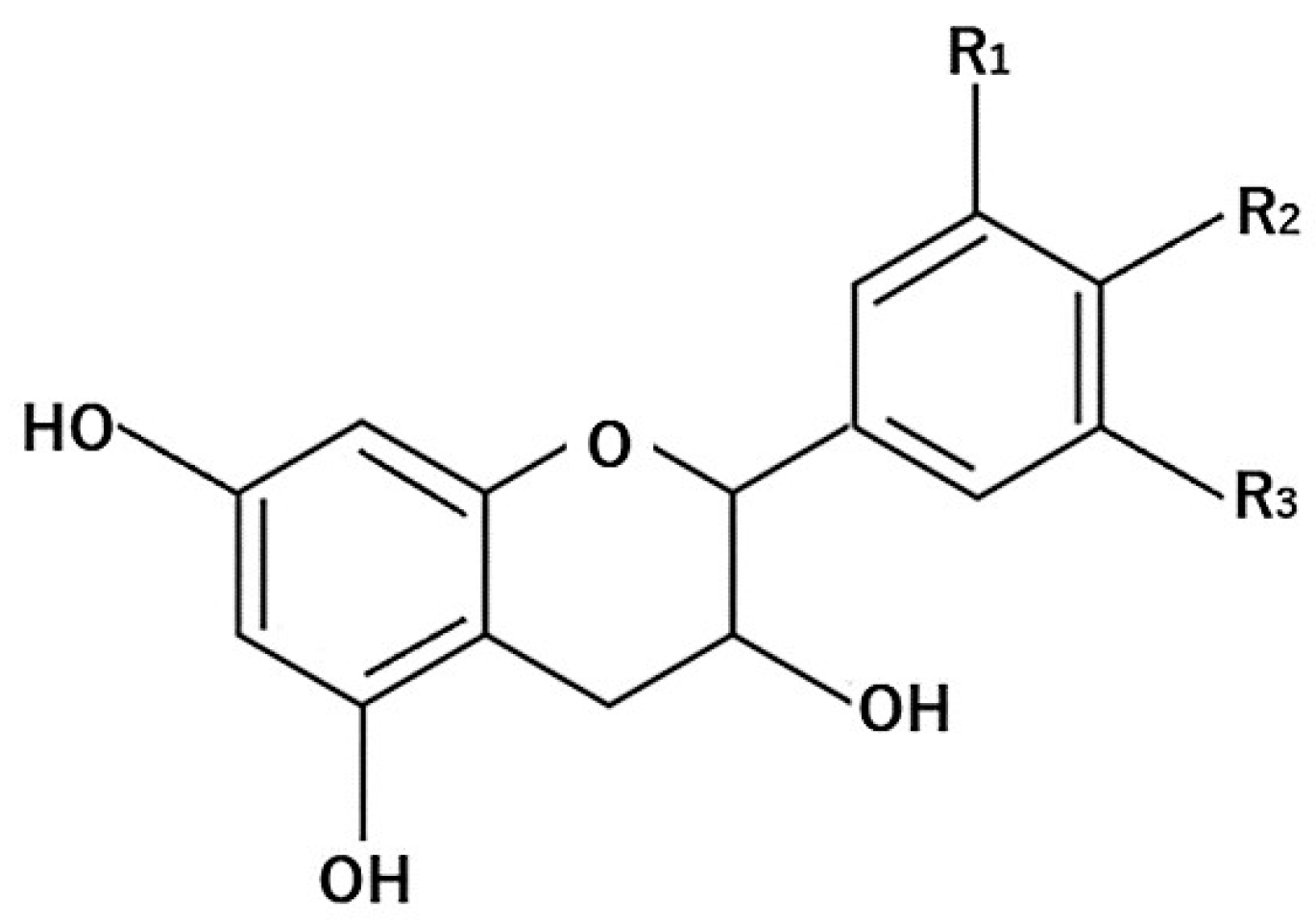
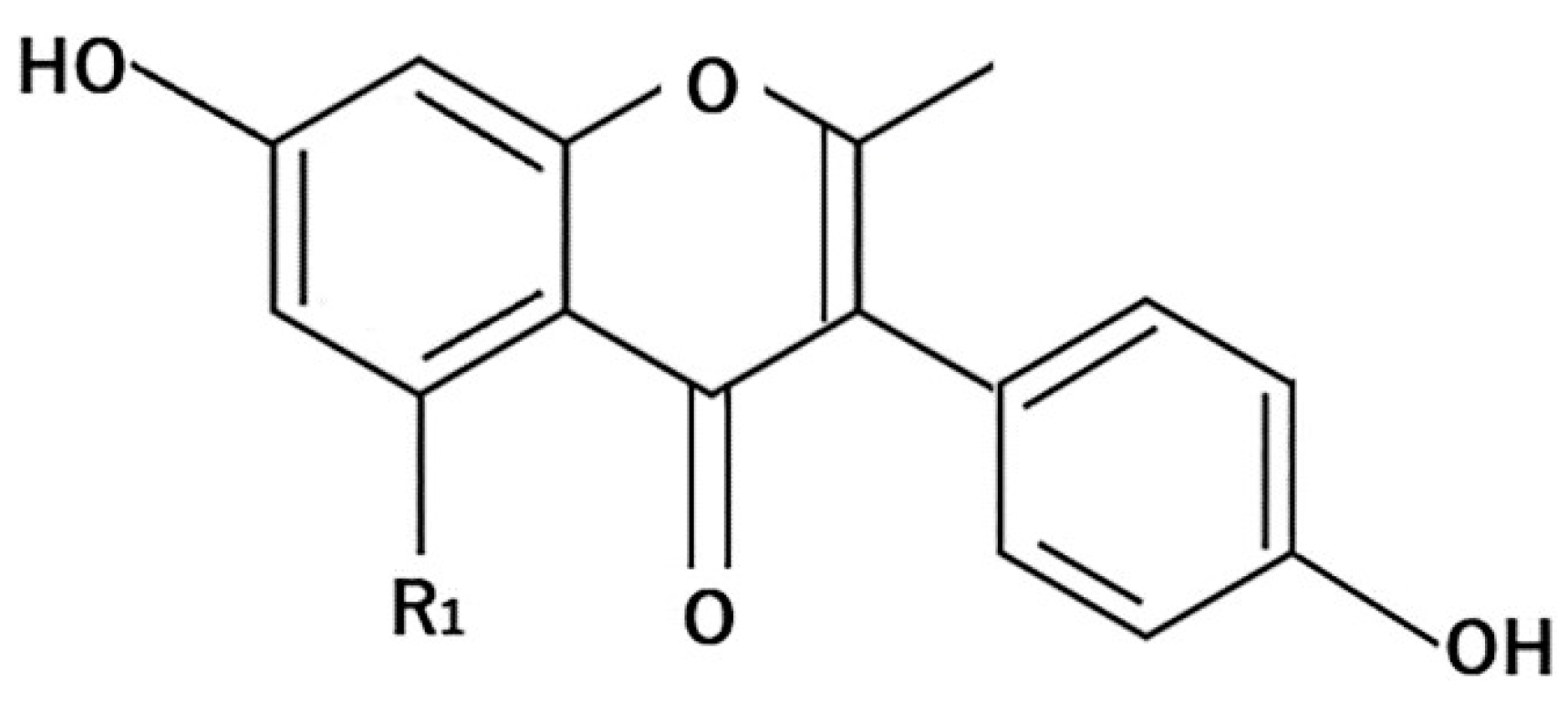
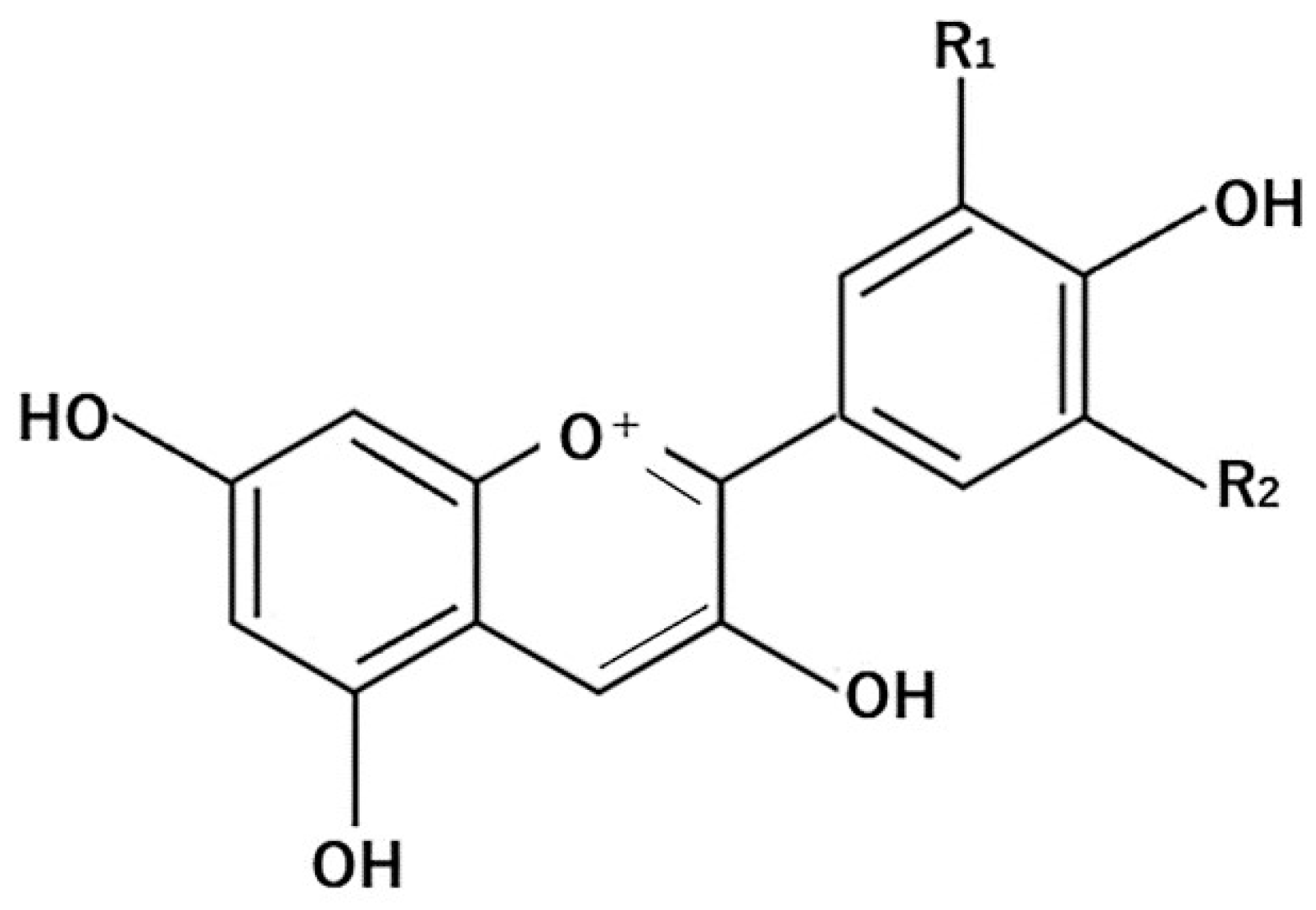
4.1. Flavonoids
4.1.1. Flavanols
4.1.2. Isoflavones
4.1.3. Anthocyanins
4.2. Non-Flavonoids
4.2.1. Green Tea Polyphenol
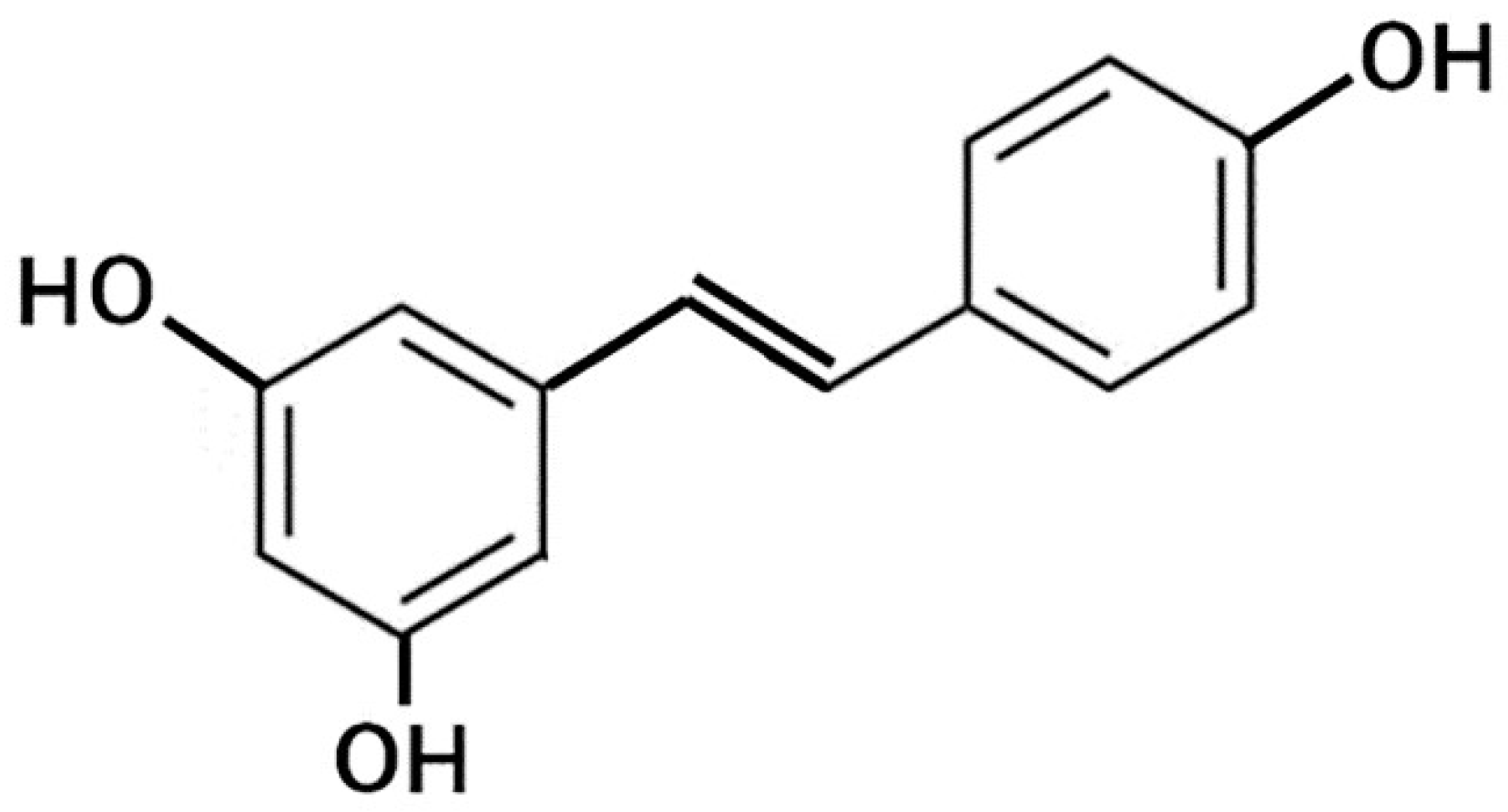
4.2.2. Resveratrol
4.2.3. Cacao Polyphenol
4.3. Metabolites from Flavonoids
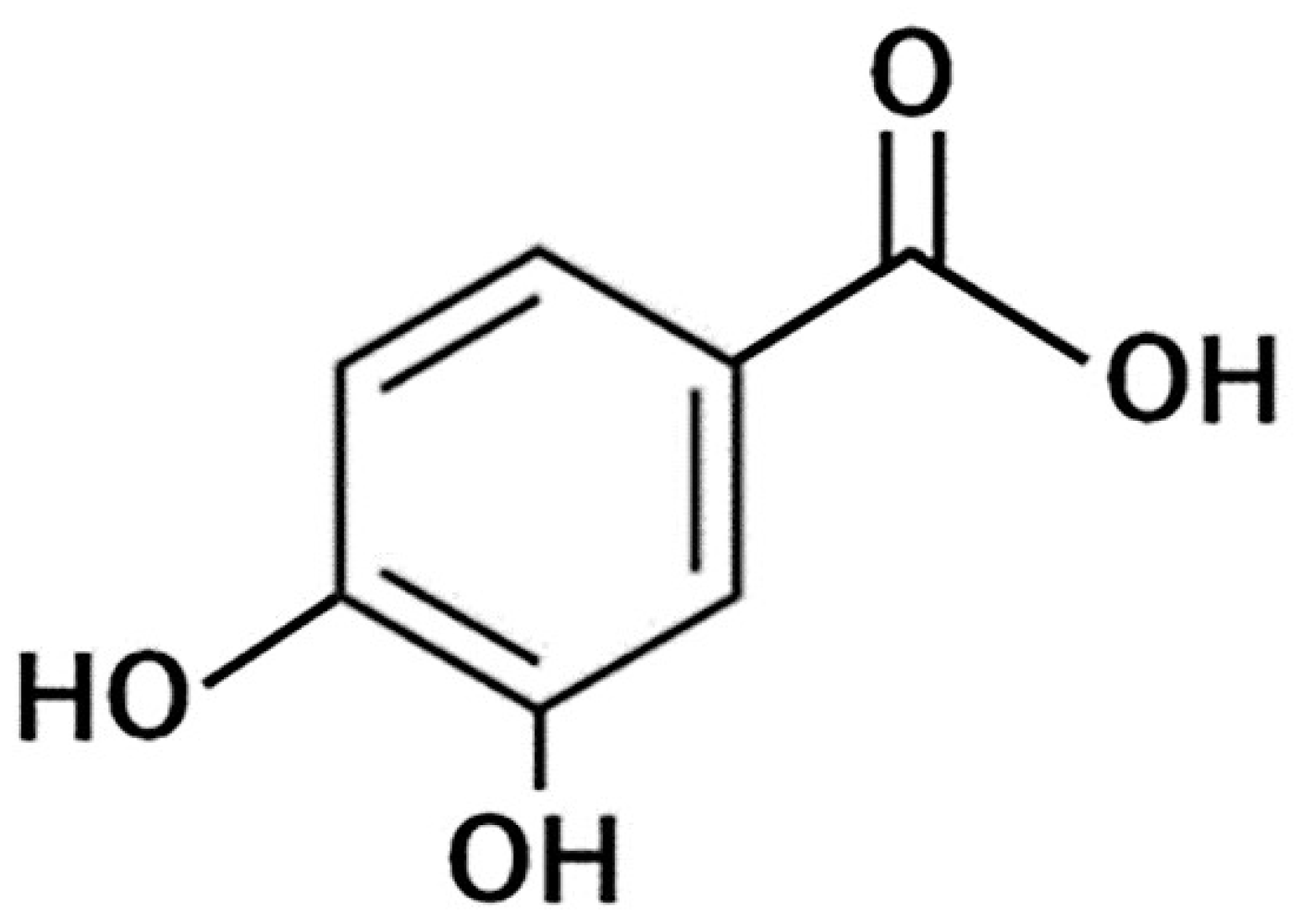
4.3.1. Protocatechuic Acid
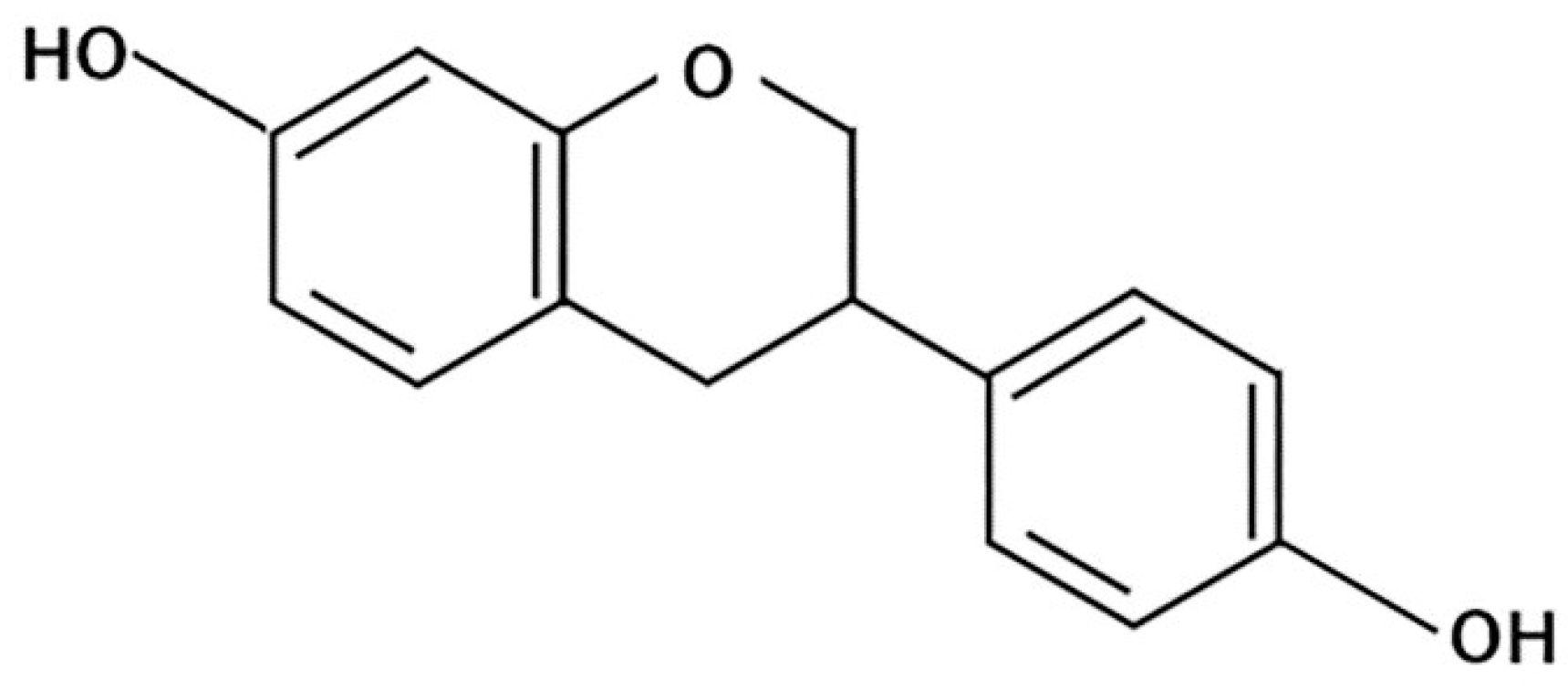
4.3.2. Equol
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Claus, G.R. Male lower urinary tract symptoms (LUTS) and benign prostatic hyperplasia (BPH). Med Clin. North Am. 2011, 95, 87–100. [Google Scholar]
- Dragan, I.; Marie, M. Lycopene for the prevention and treatment of benign prostatic hyperplasia and prostate cancer: A systematic review. Maturitas 2012, 72, 269–276. [Google Scholar]
- Bilal, C.; James, C.F.; Dominique, D.M.T.; Leanna, L.; Tania, H.; Henry, H.W.; Alexis., E.T.; Steven, A.K. Benign prostatic hyperplasia. Nat. Rev. Dis. Primers 2016, 2, 16031. [Google Scholar]
- In, S.S.; Mee, Y.L.; Hye, K.H.; Chang, S.S.; Hyeun-Kyoo, S. Inhibitory effect of Yukmijihwang-tang, a traditional herbal formula against testosterone-induced benign prostatic hyperplasia in rats. BMC Complement Altern. Med. 2012, 2, 48. [Google Scholar]
- Ester, P.; Massimiliano, L.; Michele, G.; Raffaele, C. Phytotherapy of benign prostatic hyperplasia. A minireview. Phytother. Res. 2014, 28, 949–955. [Google Scholar]
- Jack, B. Benign prostatic hyperplasia and lower urinary tract symptoms: Evidence and approaches for best case management. Can. J. Urol. 2011, 18, 14–19. [Google Scholar]
- Aleksandra, R.; Iwona, R.; Tomasz, M.; Marcin, S.; Barbara, D.; Anna, L.; Maria, L. Metabolic syndrome and benign prostatic hyperplasia: Association or coincidence? Diabetol. Metab. Syndr. 2015, 7, 94. [Google Scholar]
- Ponholzer, A.; Temml, C.; Wehrberger, C.; Marszalek, M.; Madersbacher, S. The association between vascular risk factors and lower urinary tract symptoms in both sexes. Eur. Urol. 2006, 50, 581–586. [Google Scholar] [CrossRef]
- McVary, K. Lower urinary tract symptoms and sexual dysfunction: Epidemiology and pathophysiology. BJU Int. 2006, 97, 23–28. [Google Scholar] [CrossRef]
- Gacci, M.; Corona, G.; Vignozzi, L.; Salvi, M.; Serni, S.; De, N.C.; Tubaro, A.; Oelke, M.; Carini, M.; Maggi, M. Metabolic syndrome and benign prostatic enlargement: A systematic review and meta-analysis. BJU Int. 2015, 115, 24–31. [Google Scholar] [CrossRef]
- Vibhash, C.M.; Darrell, J.A.; Christophoros, N.; Haytham, S.; Charles, H.; Omer, M.A.K.; Hanif, G.M.; Marc, E.L. Does intraprostatic inflammation have a role in the pathogenesis and progression of benign prostatic hyperplasia? BJU Int. 2007, 100, 327–331. [Google Scholar]
- Curtis, N.J.; Claus, G.R.; Michael, P.O.; David, G.B.; Matthew, C.S.; Roger, S.R. The relationship between prostate inflammation and lower urinary tract symptoms: Examination of baseline data from the REDUCE trial. Eur. Urol. 2008, 54, 1379–1384. [Google Scholar]
- Dipak, G.; Michael, I. Interleukin-1alpha is a paracrine inducer of FGF7, a key epithelial growth factor in benign prostatic hyperplasia. Am. J. Pathol. 2000, 157, 249–255. [Google Scholar]
- Gero, K.; Dieter, M.; Michael, M. Is benign prostatic hyperplasia (BPH) an immune inflammatory disease? Eur. Urol. 2007, 51, 1202–1216. [Google Scholar]
- Denis, L.; Morton, M.S.; Griffiths, K. Diet and its preventive role in prostatic disease. Eur. Urol. 1999, 35, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Miyanaga, N.; Akaza, H.; Hinotsu, S.; Fujioka, T.; Naito, S.; Namiki, M.; Takahashi, S.; Hirao, Y.; Horie, S.; Tsukamoto, T.; et al. Prostate cancer chemoprevention study: An investigative randomized control study using purified isoflavones in men with rising prostate-specific antigen. Cancer Sci. 2012, 103, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Castelli, T.; Russo, G.I.; Reale, G.; Privitera, S.; Chisari, M.; Fragalà, E.; Favilla, V.; Cimino, S.; Morgia, G. Metabolic syndrome and prostatic disease: Potentially role of polyphenols in preventive strategies. A review. Int. Braz. J. Urol. 2016, 42, 422–430. [Google Scholar] [CrossRef]
- Cicero, A.F.G.; Allkanjari, O.; Busetto, G.M.; Cai, T.; Larganà, G.; Magri, V.; Perletti, G.; Robustelli, D.C.F.S.; Russo, G.I.; Stamatiou, K.; et al. Nutraceutical treatment and prevention of benign prostatic hyperplasia and prostate cancer. Arch. Ital. Urol. Androl. 2019, 91, 3. [Google Scholar] [CrossRef]
- Kitamura, M.; Mochizuki, Y.; Miyata, Y.; Obata, Y.; Mitsunari, K.; Matsuo, T.; Ohba, K.; Mukae, H.; Yoshimura, A.; Nishino, T.; et al. Pathological Characteristics of Periodontal Disease in Patients with Chronic Kidney Disease and Kidney Transplantation. Int. J. Mol. Sci. 2019, 20, 3413. [Google Scholar] [CrossRef] [Green Version]
- Kuryłowicz, A.; Koźniewski, K. Anti-Inflammatory Strategies Targeting Metaflammation in Type 2 Diabetes. Molecules 2020, 25, 2224. [Google Scholar] [CrossRef]
- Olivas, A.; Price, R.S. Obesity, Inflammation, and Advanced Prostate Cancer. Nutr. Cancer 2020, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Tarique, H.; Bie, T.; Yulong, Y.; Francois, B.; Myrlene, C.B.T.; Najma, R. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxid. Med. Cell Longev. 2016, 2016, 7432797. [Google Scholar]
- Jean-Marc, C.; Djillali, A. Compartmentalization of the inflammatory response in sepsis and SIRS. J. Endotoxin Res. 2006, 12, 151–170. [Google Scholar]
- Nam, J.K.; Ki, W.L.; Dong, E.L.; Evgeny, A.R.; Ann, M.B.; Hyong, J.L.; Zigang, D. Cocoa procyanidins suppress transformation by inhibiting mitogen-activated protein kinase kinase. J. Biol. Chem. 2008, 283, 20664–20673. [Google Scholar]
- Peter, J.B.; Michael, K. Nuclear factor-kappaB: A pivotal transcription factor in chronic inflammatory diseases. N. Engl. J. Med. 1997, 336, 1066–1071. [Google Scholar]
- Kullisaar, T.; Türk, S.; Punab, M.; Mändar, R. Oxidative stress--cause or consequence of male genital tract disorders? Prostate 2012, 15, 977–983. [Google Scholar] [CrossRef] [PubMed]
- Paola, L.M.; Antonino, I.; Michele, N.; Gioacchino, C.; Carlo, M.; Sebastiano, G. Oxidative stress in benign prostatic hyperplasia: A systematic review. Urol. Int. 2015, 94, 249–254. [Google Scholar]
- Berlett, B.S.; Stadtman, E.R. Protein oxidation in aging, disease, and oxidative stress. J. Biol. Chem. 1997, 272, 20313–20316. [Google Scholar] [CrossRef] [Green Version]
- Salzanoa, S.; Checconia, P.; Hanschmannc, E.M.; Lillig, C.H.; Bowler, L.D.; Chan, P.; Vaudry, D.; Mengozzi, M.; Coppo, L.; Sacre, S.; et al. Linkage of inflammation and oxidative stress via release of glutathionylated peroxiredoxin-2, which acts as a danger signal. Proc. Natl. Acad. Sci. USA 2014, 111, 12157–12162. [Google Scholar] [CrossRef] [Green Version]
- Udensi, K.U.; Paul, B.T. Dual effect of oxidative stress on leukemia cancer induction and treatment. J. Exp. Clin. Cancer Res. 2014, 33, 106. [Google Scholar] [CrossRef] [Green Version]
- Udensi, K.U.; Paul, B.T. Oxidative stress in prostate hyperplasia and Carcinogenesis. J. Exp. Clin. Cancer Res. 2016, 35, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamid, A.R.; Umbas, R.; Mochtar, C.A. Recent role of inflammation in prostate diseases:chemoprevention development opportunity. Acta. Med. Indones. 2011, 43, 59–65. [Google Scholar] [PubMed]
- Lee, K.L.; Peehl, D.M. Molecular and cellular pathogenesis of benign prostatic hyperplasia. J. Urol. 2004, 172, 1784–1791. [Google Scholar] [CrossRef] [PubMed]
- Alessandro, S.; Gianna, M.; Stefano, S.; Ana, A.G.; Salvatore, M.; Vincenzo, T.; Franco, D.S. Prostate growth and inflammation. J. Steroid Biochem. Mol. Biol. 2008, 108, 254–260. [Google Scholar]
- Di Silverio, F.; Gentile, V.; De-Matteis, A. Distribution of inflammation, pre-malignant lesions, incidental carcinoma in histologically confirmed benign prostatic hyperplasia: A retrospective analysis. Eur. Urol. 2003, 43, 164–175. [Google Scholar] [CrossRef]
- Kirschenbaum, A.; Klausner, A.P.; Lee, R.; Unger, P.; Yao, S.; Liu, X.H. Expression of cyclooxygenase-1 and cyclooxygenase-2 in the human prostate. Urology 2000, 56, 671–676. [Google Scholar] [CrossRef]
- Altavilla, D.; Minutoli, L.; Polito, F.; Irrera, N.; Arena, S.; Magno, C. Effects of flavocoxid, a dual inhibitor of COX and 5-lipoxygenase enzymes, on benign prostatic hyperplasia. Br. J. Pharmacol. 2012, 167, 95–108. [Google Scholar] [CrossRef] [Green Version]
- Mbaka, G.; Ogbonnia, S.; Sulaiman, A.; Osiagwu, D. Histomorphological effects of the oil extract of Sphenocentrum jollyanum seed on benign prostatic hyperplasia induced by exogenous testosterone and estradiol in adult Wistar rats. Avicenna J. Phytomed. 2019, 9, 21–33. [Google Scholar]
- Kalu, W.; Okafor, P.; Ijeh, I.; Eleazu, C. Effect of fractions of kolaviron on some indices of benign prostatic hyperplasia in rats: Identification of the constituents of the bioactive fraction using GC-MS. RSC Adv. 2016, 6, 94352–94360. [Google Scholar]
- Malgorzata, E.Z.; Anna, M.W. Antioxidant Potential and Polyphenol Content of Beverages, Chocolates, Nuts, and Seeds. Int. J. Food Prop. 2014, 17, 86–92. [Google Scholar]
- Aryal, S.; Skinner, T.; Bridges, B.; Weber, J.T. The Pathology of Parkinson’s Disease and Potential Benefit of Dietary Polyphenols. Molecules 2020, 25, 4382. [Google Scholar] [CrossRef] [PubMed]
- Gonzali, S.; Perata, P. Anthocyanins from Purple Tomatoes as Novel Antioxidants to Promote Human Health. Antioxidants 2020, 9, 1017. [Google Scholar] [CrossRef] [PubMed]
- Lecour, S.; Lamont, K.T. Natural polyphenols and cardioprotection. Mini. Rev. Med. Chem. 2011, 11, 1191–1199. [Google Scholar] [PubMed]
- Hatcher, H.; Planalp, R.; Cho, J.; Torti, F.M.; Torti, S.V. Curcumin: From ancient medicine to current clinical trials. Cell. Mol. Life Sci. 2008, 65, 1631–1652. [Google Scholar] [CrossRef] [PubMed]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The Immunomodulatory and Anti-Inflammatory Role of Polyphenols. Nutrients 2018, 2, 1618. [Google Scholar] [CrossRef] [Green Version]
- Subedi, L.; Lee, J.H.; Yumnam, S.; Ji, E.; Kim, S.Y. Anti-Inflammatory Effect of Sulforaphane on LPS-Activated Microglia Potentially through JNK/AP-1/NF-κB Inhibition and Nrf2/HO-1 Activation. Cells 2019, 22, 194. [Google Scholar] [CrossRef] [Green Version]
- Willcox, J.K.; Ash, S.L.; Catignani, G.L. Antioxidants and prevention of chronic disease. Crit. Rev. Food Sci. Nutr. 2004, 44, 275–295. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [Green Version]
- Chanda, S.; Dave, R. In Vitro models for antioxidant activity evaluation and some medicinal plants possessing antioxidant properties: An overview. Afr. J. Microbiol. 2009, 3, 981–996. [Google Scholar]
- Bahare, S.; Elena, A.; Paolo, Z.; Elena, M.V.; Nanjangud, V.A.K.; Lucianan, D.; Elisa, P.; Jovana, R.; Patrick, V.T.F.; IIaria, P.; et al. Plant-Derived Bioactives and Oxidative Stress-Related Disorders: A Key Trend towards Healthy Aging and Longevity Promotion. Appl. Sci. 2020, 10, 947. [Google Scholar]
- Manach, C.; Scalbert, A.; Morand, C.; Remesy, C.; Jimenez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sami, R.; Chun-Juan, L.; Yan, Z.; Ying, L.; Chang-Hao, S. Cabbage (Brassica oleracea L. var. capitata) phytochemicals with antioxidant and anti-inflammatory potential. Asian Pac. J. Cancer Prev. 2014, 14, 6657–6662. [Google Scholar]
- Jie, S.; Yi-Fang, C.; Xianzhong, W.; Rui, H.L. Antioxidant and antiproliferative activities of common fruits. J. Agric. Food Chem. 2002, 50, 7449–7454. [Google Scholar]
- Eleazu, C.O. Characterization of the natural products in cocoyam (Colocasia esculenta) using GC–MS. Pharm. Biol. 2016, 54, 2880–2885. [Google Scholar] [CrossRef] [Green Version]
- Abd, E.M.M.; Marks, J.; Kuhnle, G.; Moore, K.; Debnam, E.; Srai, S.K. Absorption, tissue distribution and excretion of pelargonidin and its metabolites following oral administration to rats. Br. J. Nutr. 2006, 95, 51–58. [Google Scholar]
- Nomiya, M.; Andersson, K.E.; Yamaguchi, O. Chronic bladder ischemia and oxidative stress: New pharmacotherapeutic targets for lower urinary tract symptoms. Int. J. Urol. 2015, 40, 6. [Google Scholar] [CrossRef]
- Miyata, Y.; Matsuo, T.; Mitsunari, K.; Asai, A.; Ohba, K.; Sakai, H. A Review of Oxidative Stress and Urinary Dysfunction Caused by Bladder Outlet Obstruction and Treatments Using Antioxidants. Antioxidants 2019, 8, 132. [Google Scholar] [CrossRef] [Green Version]
- González-Vallinas, M.; González-Castejón, M.; Rodríguez-Casado, A.; Ramírez de Molina, A. Dietary phytochemicals in cancer prevention and therapy; a complementary approach with promising perspectives. Nutr. Rev. 2013, 71, 585–599. [Google Scholar] [CrossRef]
- Khan, N.; Mukhtar, H. Tea polyphenols for health promotion. Life Sci. 2007, 81, 519–533. [Google Scholar] [CrossRef] [Green Version]
- Karen, M.S.; Thomas, J.R.; Andrew, T.S.; Jeffery, S.T.; Matthew, W.H.; Dongmin, L.; Brenda, M.D.; Kevin, P.D.; Andrew, P.N. Mechanisms by which cocoa flavanols improve metabolic syndrome and related disorders. J. Nutr. Biochem. 2016, 35, 1–21. [Google Scholar]
- Yang, X.; Yuan, L.; Xiong, C.; Yin, C.; Ruan, J. Abacopteris penangiana exerts testosterone-induced benign prostatic hyperplasia protective effect through regulating inflammatory responses, reducing oxidative stress and anti-proliferative. J. Ethnopharmacol. 2014, 157, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Apaza, T.L.N.; Tena, P.V.; Bermejo, B.P. Local/traditional uses, secondary metabolites and biological activities of Mashua (Tropaeolum tuberosum Ruíz & Pavón). J. Ethnopharmacol. 2020, 247, 112152. [Google Scholar]
- Eva, B.M.; Maša, K.H.; Mojca, Š.; Željko, K.; Urban, B. Polyphenols: Extraction Methods, Antioxidative Action, Bioavailability and Anticarcinogenic Effects. Molecules 2016, 21, 901. [Google Scholar]
- Cassidy, A.; Hansley, B.; Lamuela-Raventos, R.M. Isoflavones, lignans and stilbenes-origins, metabolism and potential importance to human health. J. Sci. Food Agric. 2000, 80, 1044–1062. [Google Scholar] [CrossRef]
- Yang, C.S.; Landau, J.M.; Huang, M.-T.; Newmark, H.L. Inhibition of carcinogenesis by dietary Polyphenolic compounds. Annu. Rev. Nutr. 2001, 21, 381–406. [Google Scholar] [CrossRef] [Green Version]
- Kris-Etherton, P.M.; Hecker, K.D.; Bonanome, A.; Coval, S.M.; Binkoski, A.E.; Hilpert, K.F.; Griel, A.E.; Etherton, T.D. Bioactive Compounds in Foods: Their Role in the Prevention of Cardiovascular Disease and Cancer. Am. J. Med. 2002, 113, 71S–88S. [Google Scholar] [CrossRef]
- Pejčić, T.; Tosti, T.; Džamić, Z.; Gašić, U.; Vuksanović, A.; Dolićanin, Z.; Tešić, Ž. The Polyphenols as Potential Agents in Prevention and Therapy of Prostate Diseases. Molecules 2019, 24, 3982. [Google Scholar] [CrossRef] [Green Version]
- Sivoňová, M.K.; Kaplán, P.; Tatarková, Z.; Lichardusová, L.; Dušenka, R.; Jurečeková, J. Androgen receptor and soy isoflavones in prostate cancer. Mol. Clin. Oncol. 2019, 10, 191–204. [Google Scholar] [CrossRef] [Green Version]
- Grammatikopoulou, M.G.; Gkiouras, K.; Papageorgiou, S.Τ.; Myrogiannis, I.; Mykoniatis, I.; Papamitsou, T.; Bogdanos, D.P.; Goulis, D.G. Dietary Factors and Supplements Influencing Prostate Specific-Antigen (PSA) Concentrations in Men with Prostate Cancer and Increased Cancer Risk: An Evidence Analysis Review Based on Randomized Controlled Trials. Nutrients 2020, 12, 2985. [Google Scholar] [CrossRef]
- Geller, J.; Sionit, L.; Partido, C.; Li, L.; Tan, X.; Youngkin, T.; Nachtsheim, D.; Hoffman, R.M. Genistein inhibits the growth of human-patient BPH and prostate cancer in histoculture. Prostate 1998, 34, 75–79. [Google Scholar] [CrossRef]
- Olbina, G.; Miljkovic, D.; Hoffman, R.M.; Geller, J. New sensitive discovery histoculture model for growth-inhibition studies in prostate cancer and BPH. Prostate 1998, 37, 126–129. [Google Scholar] [CrossRef]
- Weber, K.S.; Setchell, K.D.; Stocco, D.M.; Lephart, E.D. Dietary soy-phytoestrogens decrease testosterone levels and prostate weight without altering LH, prostate 5alpha-reductase or testicular steroidogenic acute regulatory peptide levels in adult male Sprague-Dawley rats. J. Endocrinol. 2001, 170, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Jarred, R.A.; McPherson, S.J.; Jones, M.E.; Simpson, E.R.; Risbridger, G.P. Anti-androgenic action by red clover-derived dietary isoflavones reduces non-malignant prostate enlargement in aromatase knockout (ArKo) mice. Prostate 2003, 56, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Slater, M.; Brown, D.; Husband, A. In the prostatic epithelium, dietary isoflavones from red clover significantly increase estrogen receptor beta and E-cadherin expression but decrease transforming growth factor beta1. Prostate Cancer Prostatic Dis. 2002, 5, 16–21. [Google Scholar] [CrossRef]
- Masrudin, S.S.; Mohamad, J. Preventive effect of Pueraria mirifica on testosterone-induced prostatic hyperplasia in Sprague Dawley rats. Andrologia 2015, 47, 1153–1159. [Google Scholar] [CrossRef]
- Brandli, A.; Simpson, J.S.; Ventura, S. Isoflavones isolated from red clover (Trifolium pratense) inhibit smooth muscle contraction of the isolated rat prostate gland. Phytomedicine 2010, 17, 895–901. [Google Scholar] [CrossRef]
- Wong, W.C.; Wong, E.L.; Li, H.; You, J.H.; Ho, S.; Woo, J.; Hui, E. Isoflavones in treating watchful waiting benign prostate hyperplasia: A double-blinded, randomized controlled trial. J. Altern. Complement Med. 2012, 18, 54–60. [Google Scholar] [CrossRef] [Green Version]
- Tiscione, D.; Gallelli, L.; Tamanini, I.; Luciani, L.G.; Verze, P.; Palmieri, A.; Mirone, V.; Bartoletti, R.; Malossini, G.; Cai, T. Daidzein plus isolase associated with zinc improves clinical symptoms and quality of life in patients with LUTS due to benign prostatic hyperplasia: Results from a phase I-II study. Arch Ital. Urol. Androl. 2017, 89, 12–16. [Google Scholar] [CrossRef] [Green Version]
- Katz, A.E. Flavonoid and botanical approaches to prostate health. J. Altern. Complement Med. 2002, 8, 813–821. [Google Scholar] [CrossRef]
- Gaynor, M.L. Isoflavones and the prevention and treatment of prostate disease: Is there a role? Cleve Clin. J. Med. 2003, 70, 203–204. [Google Scholar] [CrossRef]
- Hong, S.J.; Kim, S.I.; Kwon, S.M.; Lee, J.R.; Chung, B.C. Comparative study of concentration of isoflavones and lignans in plasma and prostatic tissues of normal control and benign prostatic hyperplasia. Yonsei Med. J. 2002, 43, 236–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bektic, J.; Berger, A.P.; Pfeil, K.; Dobler, G.; Bartsch, G.; Klocker, H. Androgen receptor regulation by physiological concentrations of the isoflavonoid genistein in androgen-dependent LNCaP cells is mediated by estrogen receptor beta. Eur. Urol. 2004, 45, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Ajdžanovic, V.; Filipovic, B.; Miljic, D.; Mijatovic, S.; Maksimovic-Ivanic, D.; Miler, M.; Živanovic, J.; Miloševic, V. Prostate cancer metastasis and soy isoflavones: A dogfight over a bone. EXCLI J. 2019, 18, 106–126. [Google Scholar] [PubMed]
- Ly, C.; Yockell-Lelièvre, J.; Ferraro, M.Z.; Arnason, J.T.; Ferrier, J.; Gruslin, A. The effects of dietary polyphenols on reproductive health and early development. Hum. Reprod. Update 2015, 21, 228–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, J.; Mumper, J.R. Plant Phenolics: Extraction, Analysis and Their Antioxidant and Anticancer Properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef] [PubMed]
- Konczak, I.; Zhang, W. Anthocyanins—More Than Nature’s Colours. J. Biomed. Biotechnol. 2004, 5, 239–240. [Google Scholar] [CrossRef] [Green Version]
- Lila, M.A. Anthocyanins and Human Health: An in Vitro Investigative Approach. J. Biomed. Biotechnol. 2004, 5, 306–313. [Google Scholar] [CrossRef]
- Hoon, J.; U-Syn, H.; Su-Jin, K.; Byung-I, Y.; Dong-Seok, H.; Seung-Mo, Y. Anthocyanin extracted from black soybean reduces prostate weight and promotes apoptosis in the prostatic hyperplasia-induced rat model. J. Agric. Food Chem. 2010, 58, 12686–12691. [Google Scholar]
- Choi, Y.J.; Fan, M.; Tang, Y.; Yang, H.P.; Hwang, J.Y.; Kim, E.K. In Vivo Effects of Polymerized Anthocyanin from Grape Skin on Benign Prostatic Hyperplasia. Nutrients 2019, 11, 2444. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.F.; Tang, L.P.; He, R.R.; Xu, Z.; He, Q.Q.; Xiang, F.J.; Su, W.W.; Kurihara, H. Anthocyanin extract from bilberry enhances the therapeutic effect of pollen of Brassica napus L. on stress-provoked benign prostatic hyperplasia in restrained mice. J. Funct. Foods 2013, 5, 1357–1365. [Google Scholar] [CrossRef]
- Han, H.Y.; Shan, S.; Zhang, X.; Wang, N.L.; Lu, X.P.; Yao, X.S. Down-regulation of prostate specific antigen in LNCaP cells by flavonoids from the pollen of Brassica napus L. Phytomedicine 2007, 14, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.; Bae, W.J.; Yuk, S.M.; Han, D.S.; Ha, U.S.; Hwang, S.Y.; Yoon, S.H.; Kim, S.W.; Han, C.H. Seoritae extract reduces prostate weight and suppresses prostate cell proliferation in a rat model of benign prostate hyperplasia. Evid. Based Complement. Alternat. Med. 2014, 2014, 475876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, N.; Afaq, F.; Saleem, M.; Ahmad, N.; Mukhtar, H. Targeting multiple signaling pathways by green tea polyphenol (−)-epigallocatechin-3-gallate. Cancer Res. 2006, 66, 2500–2505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weisburger, J.H. Tea and health: A historical perspective. Cancer Lett. 1997, 114, 315–317. [Google Scholar] [CrossRef]
- Tepedelen, B.E.; Soya, E.; Korkmaz, M. Epigallocatechin-3-gallate reduces the proliferation of benign prostatic hyperplasia cells via regulation of focal adhesions. Life Sci. 2017, 191, 74–81. [Google Scholar] [CrossRef]
- Sagara, Y.; Miyata, Y.; Nomata, K.; Hayashi, T.; Kanetake, H. Green tea polyphenol suppresses tumor invasion and angiogenesis in N-butyl-(-4-hydroxybutyl) nitrosamine-induced bladder cancer. Cancer Epidemiol. 2010, 34, 350–354. [Google Scholar] [CrossRef]
- Miyata, Y.; Matsuo, T.; Araki, K.; Nakamura, Y.; Sagara, Y.; Ohba, K.; Sakai, H. Anticancer Effects of Green Tea and the Underlying Molecular Mechanisms in Bladder Cancer. Medicines 2018, 5, 87. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Zhang, L.; Li, C.; Chen, R.; Liu, C.; Chen, M. Lipophilized Epigallocatechin Gallate Derivative Exerts Anti-Proliferation Efficacy through Induction of Cell Cycle Arrest and Apoptosis on DU145 Human Prostate Cancer Cells. Nutrients 2019, 12, 92. [Google Scholar] [CrossRef] [Green Version]
- Kiwanuka, E.; Lee, C.C.; Hackl, F.; Caterson, E.J.; Junker, J.P.; Gerdin, B.; Eriksson, E. Cdc42 and p190RhoGAP activation by CCN2 regulates cell spreading and polarity and induces actin disassembly in migrating keratinocytes. Int. Wound J. 2016, 13, 372–381. [Google Scholar] [CrossRef]
- Wu, G.S.; Song, Y.L.; Yin, Z.Q.; Guo, J.J.; Wang, S.P.; Zhao, W.W.; Chen, X.P.; Zhang, Q.W.; Lu, J.J.; Wang, Y.T. Ganoderiol A-enriched extract suppresses migration and adhesion of MDA-MB-231 cells by inhibiting FAK-SRC-paxillin cascade pathway. PLoS ONE 2013, 8, e76620. [Google Scholar] [CrossRef] [Green Version]
- de Assis Lima, M.; da Silva, S.V.; Serrano-Garrido, O.; Hülsemann, M.; Santos-Neres, L.; Rodríguez-Manzaneque, J.C.; Hodgson, L.; Freitas, V.M. Metalloprotease ADAMTS-1 decreases cell migration and invasion modulating the spatiotemporal dynamics of Cdc42 activity. Cell Signal. 2021, 77, 109827. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Song, H. Protective potential of epigallocatechin-3-gallate against benign prostatic hyperplasia in metabolic syndrome rats. Environ. Toxicol. Pharmacol. 2016, 45, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Sreenivasulu, K.; Nandeesha, H.; Dorairajan, L.N.; Rajappa, M.; Vinayagam, V.; Cherupanakkal, C. Gene expression of insulin receptor, insulin-like growth factor increases and insulin-like growth factor-binding protein-3 reduces with increase in prostate size in benign prostatic hyperplasia. Aging Male 2018, 21, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Hata, J.; Tanji, R.; Onagi, A.; Honda-Takinami, R.; Matsuoka, K.; Hoshi, S.; Sato, Y.; Akaihata, H.; Haga, N.; Kojima, Y. Morphological change and characteristics of myofibroblasts during the growth process of benign prostatic hyperplasia. Int. J. Urol. 2020, 27, 676–683. [Google Scholar] [CrossRef] [PubMed]
- Ishola, I.O.; Tijani, H.K.; Dosumu, O.O.; Anunobi, C.C.; Oshodi, T.O. Atorvastatin attenuates testosterone-induced benign prostatic hyperplasia in rats: Role of peroxisome proliferator-activated receptor-γ and cyclo-oxygenase-2. Fundam. Clin. Pharmacol. 2017, 31, 652–662. [Google Scholar] [CrossRef]
- Refaie, M.M.M.; Rifaai, R.A.; Zenhom, N.M. Role of PPAR-α agonist fenofibrate in the treatment of induced benign prostatic hyperplasia with dysplastic changes in rats. Fundam. Clin. Pharmacol. 2018, 32, 617–626. [Google Scholar] [CrossRef]
- Zhou, J.; Lei, Y.; Chen, J.; Zhou, X. Potential ameliorative effects of epigallocatechin-3-gallate against testosterone-induced benign prostatic hyperplasia and fibrosis in rats. Int. Immunopharmacol. 2018, 64, 162–169. [Google Scholar] [CrossRef]
- Stefanou, D.; Batistatou, A.; Kamina, S.; Arkoumani, E.; Papachristou, D.J.; Agnantis, N.J. Expression of vascular endothelial growth factor (VEGF) and association with microvessel density in benign prostatic hyperplasia and prostate cancer. In Vivo 2004, 18, 155–160. [Google Scholar]
- Chen, Y.; Xu, H.; Shi, Q.; Gu, M.; Wan, X.; Chen, Q.; Wang, Z. Hypoxia-inducible factor 1α (HIF-1α) mediates the epithelial-mesenchymal transition in benign prostatic hyperplasia. Int. J. Clin. Exp. Pathol. 2019, 12, 295–304. [Google Scholar]
- Chen, W.; Huang, X.; Peng, A.; Chen, T.; Yang, R.; Huang, Y.; Yang, Z.; Xi, S. Kangquan Recipe Regulates the Expression of BAMBI Protein via the TGF- β/Smad Signaling Pathway to Inhibit Benign Prostatic Hyperplasia in Rats. Evid. Based Complement. Alternat. Med. 2019, 2019, 6281819. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.Y.; Jin, B.R.; Chung, T.W.; Bae, S.J.; Park, H.; Ryu, D.; Jin, L.; An, H.J.; Ha, K.T. 6-sialyllactose ameliorates dihydrotestosterone-induced benign prostatic hyperplasia through suppressing VEGF-mediated angiogenesis. BMB Rep. 2019, 52, 560–565. [Google Scholar] [CrossRef] [PubMed]
- Duan, L.J.; Qi, J.; Kong, X.J.; Huang, T.; Qian, X.Q.; Xu, D.; Liang, J.H.; Kang, J. MiR-133 modulates TGF-β1-induced bladder smooth muscle cell hypertrophic and fibrotic response: Implication for a role of microRNA in bladder wall remodeling caused by bladder outlet obstruction. Cell Signal. 2015, 27, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Chen, Q.; Chen, J.; Liu, D. Potential ameliorative effects of grape seed-derived polyphenols against cadmium induced prostatic deficits. Biomed. Pharmacother. 2017, 91, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Liao, S. The medicinal action of androgens and green tea epigallocatechin gallate. Hong Kong Med. J. 2001, 7, 369–374. [Google Scholar]
- Ranjan, P.; Dalela, D.; Sankhwar, S.N. Diet and benign prostatic hyperplasia: Implications for prevention. Urology 2006, 68, 470–476. [Google Scholar] [CrossRef]
- Ciesek, S.; von Hahn, T.; Colpitts, C.C.; Schang, L.M.; Friesland, M.; Steinmann, J.; Manns, M.P.; Ott, M.; Wedemeyer, H.; Meuleman, P.; et al. The green tea polyphenol, epigallocatechin-3-gallate, inhibits hepatitis C virus entry. Hepatology 2011, 54, 1947–1955. [Google Scholar] [CrossRef]
- Wang, L.C.; Pan, T.M.; Tsai, T.Y. Lactic acid bacteria-fermented product of green tea and Houttuynia cordata leaves exerts anti-adipogenic and anti-obesity effects. J. Food Drug Anal. 2018, 26, 973–984. [Google Scholar] [CrossRef]
- Miyata, Y.; Shida, Y.; Hakariya, T.; Sakai, H. Anti-Cancer Effects of Green Tea Polyphenols Against Prostate Cancer. Molecules 2019, 24, 193. [Google Scholar] [CrossRef] [Green Version]
- Calmasini, F.B.; de Oliveira, M.G.; Alexandre, E.C.; Silva, F.H.; Tavares, E.B.G.; André, D.M.; Zapparoli, A.; Antunes, E. Obesity-induced mouse benign prostatic hyperplasia (BPH) is improved by treatment with resveratrol: Implication of oxidative stress, insulin sensitivity and neuronal growth factor. J. Nutr. Biochem. 2018, 55, 53–58. [Google Scholar] [CrossRef]
- Guan, Z.; Shen, L.; Liang, H.; Yu, H.; Hei, B.; Meng, X.; Yang, L. Resveratrol inhibits hypoxia-induced proliferation and migration of pulmonary artery vascular smooth muscle cells by inhibiting the phosphoinositide 3-kinase/protein kinase B signaling pathway. Mol. Med. Rep. 2017, 16, 1653–1660. [Google Scholar] [CrossRef] [Green Version]
- Chao, L.; Wan-Li, H.; Meng-Xin, L.; Guan-Fa, X. Resveratrol induces apoptosis of benign prostatic hyperplasia epithelial cell line (BPH-1) through p38 MAPK-FOXO3a pathway. BMC Complement Altern. Med. 2019, 19, 233. [Google Scholar]
- Kyung-Sook, C.; Se-Yun, C.; Hyo-Jin, A. Effects of resveratrol on benign prostatic hyperplasia by the regulation of inflammatory and apoptotic proteins. J. Nat. Prod. 2015, 78, 689–694. [Google Scholar]
- Kjaer, T.N. Resveratrol reduces the levels of circulating androgen precursors but has no effect on, testosterone, dihydrotestosterone, PSA levels or prostate volume. A 4-month randomised trial in middle-aged men. Prostate 2015, 75, 1255–1263. [Google Scholar] [CrossRef] [PubMed]
- Shalaby, A.M.; Alabiad, M.A.; El, S.D.F. Resveratrol Ameliorates the Seminiferous Tubules Damages Induced by Finasteride in Adult Male Rats. Microsc. Microanal. 2020, 26, 1176–1186. [Google Scholar] [CrossRef]
- Jean-François, B.; Sophie, H.; Pascale, R.; Michaël, M. Therapeutic effect of ACTICOA powder, a cocoa polyphenolic extract, on experimentally induced prostate hyperplasia in Wistar-Unilever rats. J. Med. Food. 2007, 10, 628–635. [Google Scholar]
- Adedara, I.A.; Omole, O.; Okpara, E.S.; Fasina, O.B.; Ayeni, M.F.; Ajayi, O.M.; Busari, E.O.; Farombi, E.O. Impact of prepubertal exposure to dietary protocatechuic acid on the hypothalamic-pituitary-testicular axis in rats. Chem. Biol. Interact. 2018, 290, 99–109. [Google Scholar] [CrossRef]
- Tsuda, T.; Horio, F.; Osawa, T. Absorption and metabolism of cyanidin 3-O-beta-D-glucoside in rats. FEBS Lett. 1999, 449, 179–182. [Google Scholar] [CrossRef] [Green Version]
- Olubukola, O.A.; Solomon, E.O.; Olusegun, G.O.; Adebukola, A.A.; Olusoji, J.A.; Oluwatosin, A.A. Protocatechuic acid ameliorates testosterone-induced benign prostatic hyperplasia through the regulation of inflammation and oxidative stress in castrated rats. J. Biochem. Mol. Toxicol. 2020, 34, e22502. [Google Scholar]
- Tao, R.; Miao, L.; Yu, X.; Orgah, J.O.; Barnabas, O.; Chang, Y.; Liu, E.; Fan, G.; Gao, X. Cynomorium songaricum Rupr demonstrates phytoestrogenic or phytoandrogenic like activities that attenuates benign prostatic hyperplasia via regulating steroid 5-α-reductase. J. Ethnopharmacol. 2019, 235, 65–74. [Google Scholar] [CrossRef]
- Lephart, E.D. Skin aging and oxidative stress: Equol’s anti-aging effects via biochemical and molecular mechanisms. Ageing Res Rev. 2016, 31, 36–54. [Google Scholar] [CrossRef]
- Křížová, L.; Dadáková, K.; Kašparovská, J.; Kašparovský, T. Isoflavones. Molecules 2019, 24, 1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trent, D.L.; Crystal, B.; Lihong, B.; Amy, N.H.; Edwin, D.L. Equol an isoflavonoid: Potential for improved prostate health, in vitro and in vivo evidence. Reprod. Biol. Endocrinol. 2011, 9, 4. [Google Scholar]
- Lephart, E.D. Severe and moderate BPH symptoms in mid-aged men improved with isoflavonoid-equol treatment: Pilot intervention study. Open J. Urol. 2013, 3, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Edwin, D.L. Anti-oxidant and anti-aging properties of equol in prostate health. Open J. Endocr. Metab. Dis. 2014, 4, 1–12. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecules | Sample | Change | Author/Year/Reference |
|---|---|---|---|
| Anti-oxidants | |||
| CAT | Tissue | ↑ | Zhou, J./2018/[107] |
| GPX | Blood | ↑ | Chen, J./2016/[102] |
| Tissue | ↑ | Chen, J./2016/[102] | |
| Tissue | ↑ | Zhou, J./2018/[107] | |
| GSH | Tissue | ↑ | Zhou, J./2018/[107] |
| SOD | Blood | ↑ | Chen, J./2016/[102] |
| Tissue | ↑ | Chen, J./2016/[102] | |
| Tissue | ↑ | Zhou, J./2018/[107] | |
| TSH | Tissue | ↑ | Zhou, J./2018/[107] |
| Oxidative marker | |||
| MDA | Blood | ↓ | Chen, J./2016/[102] |
| Tissue | ↓ | Chen, J./2016/[102] | |
| Tissue | ↓ | Zhou, J./2018/[107] | |
| Pro-inflammation | |||
| COX-2 | Tissue | ↓ | Zhou, J./2018/[107] |
| IL-1β | Blood | ↓ | Chen, J./2016/[102] |
| Tissue | ↓ | Chen, J./2016/[102] | |
| Tissue | ↓ | Zhou, J./2018/[107] | |
| Cell line | ↓ | Cicero, A.F.G./2019/[18] | |
| IL-6 | Blood | ↓ | Chen, J./2016/[102] |
| Tissue | ↓ | Chen, J./2016/[102] | |
| Tissue | ↓ | Zhou, J./2018/[107] | |
| IL-16 | Cell line | ↓ | Cicero, A.F.G./2019/[18] |
| P65; total | Tissue | → | Zhou, J./2018/[107] |
| P65; phosphorylated | Tissues | ↓ | Zhou, J./2018/[107] |
| TNF-α | Blood | ↓ | Chen, J./2016/[102] |
| Tissue | ↓ | Chen, J./2016/[102] | |
| Tissue | ↓ | Zhou, J./2018/[107] | |
| Cell line | ↓ | Cicero, A.F.G./2019/[18] |
| Molecules | Sample | Change | Author/Year/Reference |
|---|---|---|---|
| Hormonal | |||
| 5α-reductase | Tissues | ↓ | Liao, S./2001/[114] |
| AR | Tissues | ↓ | Zhou, J./2018/[107] |
| ER-α | Tissues | ↓ | Zhou, J./2018/[107] |
| ER-β | Tissues | ↑ | Zhou, J./2018/[107] |
| Growth factors | |||
| Basic FGF | Tissue | ↓ | Zhou, J./2018/[107] |
| EGF | Tissue | ↓ | Zhou, J./2018/[107] |
| IGF-1 | Tissue | ↓ | Chen, J./2016/[102] |
| Cell line | ↓ | Cicero, A.F.G./2019/[18] | |
| IGF-2 | Tissue | ↓ | Chen, J./2016/[102] |
| Cell line | ↓ | Cicero, AFG/2019/[18] | |
| IGFBP-3 | Tissue | ↑ | Chen, J./2016/[102] |
| TGF-β1 | Tissue | ↓ | Zhou, J./2018/[107] |
| TGF-β1 receptor | Tissue | ↓ | Zhou, J./2018/[107] |
| VEGF | Blood | ↓ | Chen, J./2016/[102] |
| Tissue | ↓ | Chen, J./2016/[102] | |
| Tissue | ↓ | Zhou, J./2018/[107] |
| Molecules | Sample | Change | Author/Year/Reference |
|---|---|---|---|
| α-SMA | Tissue | ↓ | Zhou, J./2018/[107] |
| Cdc42 | Cell line | ↓ | Tepedelen, B.E./2017/[95] |
| E-cadherin | Tissue | ↑ | Zhou, J./2018/[107] |
| FAK; total | Cell line | → | Tepedelen, B.E./2017/[95] |
| FAK; phosphorylated | Cell line | ↓ | Tepedelen, B.E./2017/[95] |
| Fibronectin | Tissue | ↓ | Zhou, J./2018/[107] |
| HIF-1α | Tissue | ↓ | Zhou, J./2018/[107] |
| miR-133a | Tissue | ↑ | Zhou, J./2018/[107] |
| miR-133b | Tissue | ↑ | Zhou, J./2018/[107] |
| PAK | Cell line | ↓ | Tepedelen, B.E./2017/[95] |
| PPAR-α | Tissue | ↑ | Chen, J./2016/[102] |
| Cell line | ↓ | Cicero, A.F.G./2019/[18] | |
| PPAR-γ | Tissue | ↑ | Chen, J./2016/[102] |
| Cell line | ↓ | Cicero, A.F.G./2019/[18] | |
| Paxillin | Cell line | ↓ | Tepedelen, B.E./2017/[95] |
| Rho A | Cell line | ↓ | Tepedelen, B.E./2017/[95] |
| Smad3; total | Cell line | → | Zhou, J./2018/[107] |
| Smad3; phosphorylated | Cell line | ↓ | Zhou, J./2018/[107] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mitsunari, K.; Miyata, Y.; Matsuo, T.; Mukae, Y.; Otsubo, A.; Harada, J.; Kondo, T.; Matsuda, T.; Ohba, K.; Sakai, H. Pharmacological Effects and Potential Clinical Usefulness of Polyphenols in Benign Prostatic Hyperplasia. Molecules 2021, 26, 450. https://doi.org/10.3390/molecules26020450
Mitsunari K, Miyata Y, Matsuo T, Mukae Y, Otsubo A, Harada J, Kondo T, Matsuda T, Ohba K, Sakai H. Pharmacological Effects and Potential Clinical Usefulness of Polyphenols in Benign Prostatic Hyperplasia. Molecules. 2021; 26(2):450. https://doi.org/10.3390/molecules26020450
Chicago/Turabian StyleMitsunari, Kensuke, Yasuyoshi Miyata, Tomohiro Matsuo, Yuta Mukae, Asato Otsubo, Junki Harada, Tsubasa Kondo, Tsuyoshi Matsuda, Kojiro Ohba, and Hideki Sakai. 2021. "Pharmacological Effects and Potential Clinical Usefulness of Polyphenols in Benign Prostatic Hyperplasia" Molecules 26, no. 2: 450. https://doi.org/10.3390/molecules26020450
APA StyleMitsunari, K., Miyata, Y., Matsuo, T., Mukae, Y., Otsubo, A., Harada, J., Kondo, T., Matsuda, T., Ohba, K., & Sakai, H. (2021). Pharmacological Effects and Potential Clinical Usefulness of Polyphenols in Benign Prostatic Hyperplasia. Molecules, 26(2), 450. https://doi.org/10.3390/molecules26020450