
Anti-Proliferative Activity of A Hydrophilic Extract of Manna from Fraxinus angustifolia Vahl through Mitochondrial Pathway-Mediated Apoptosis and Cell Cycle Arrest in Human Colon Cancer Cells

 ,
,  ,
, 

 and
and 
Abstract
:1. Introduction
2. Results
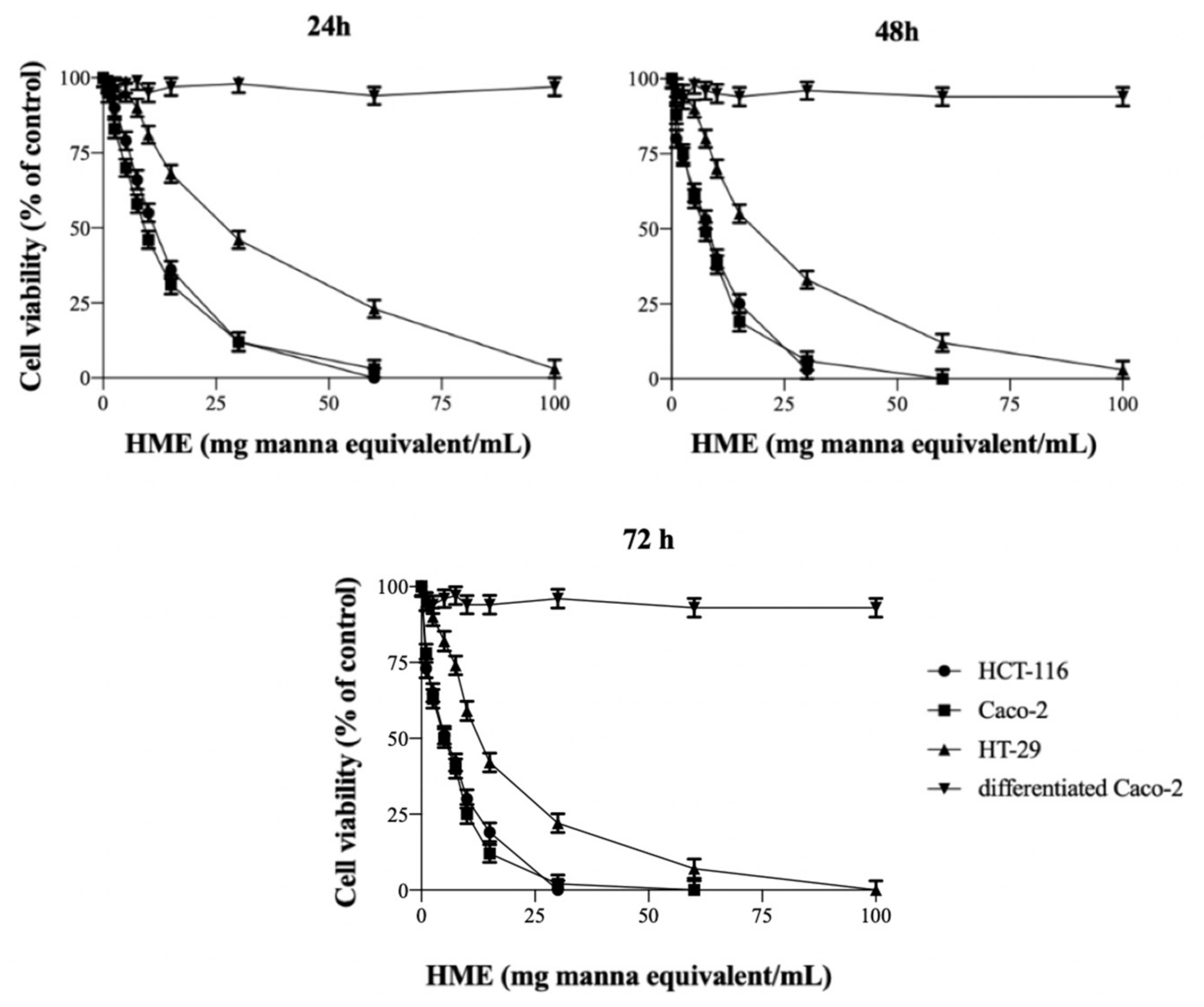
2.1. HME Inhibits the Proliferation of Human Intestinal Cancer Cells
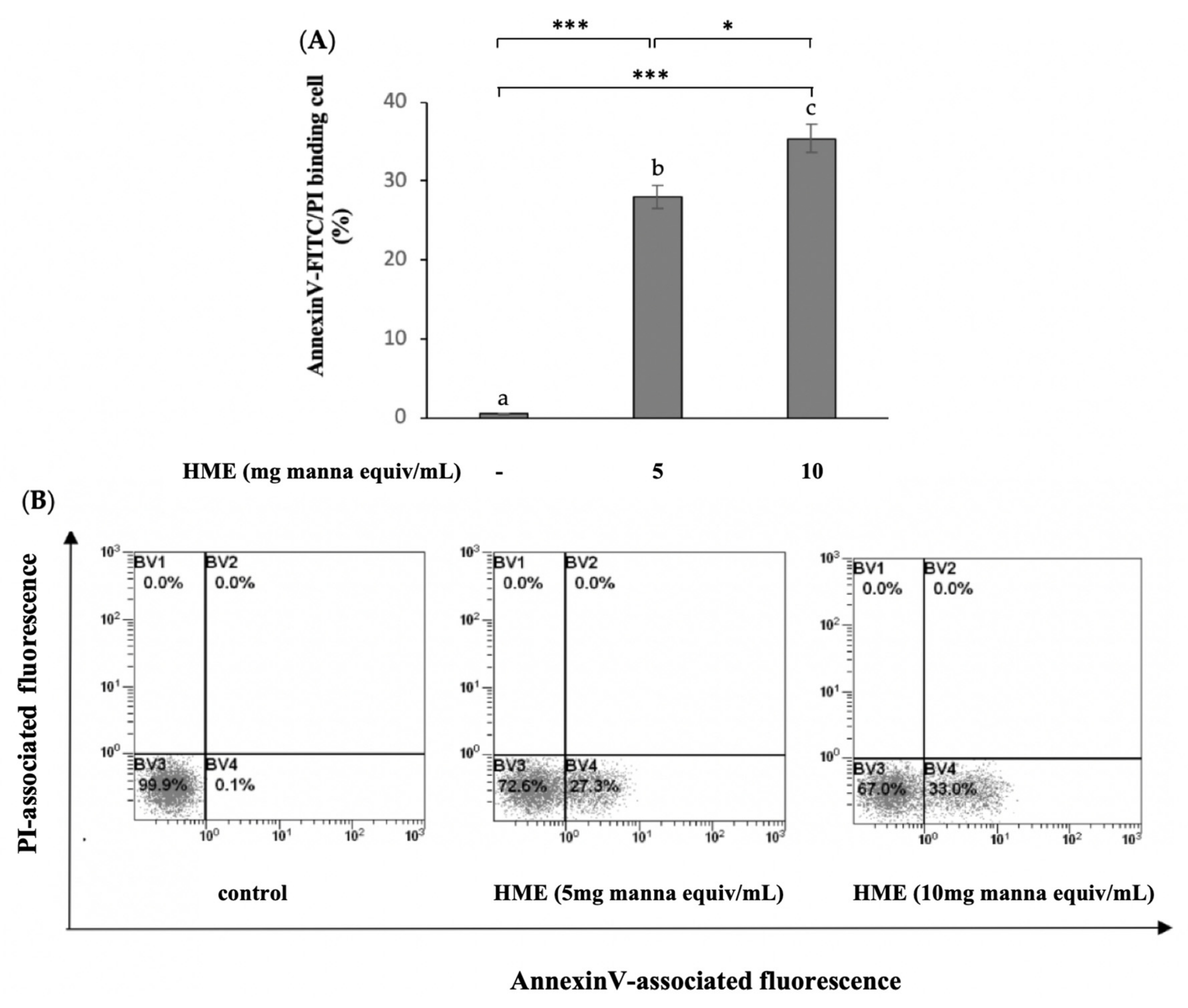
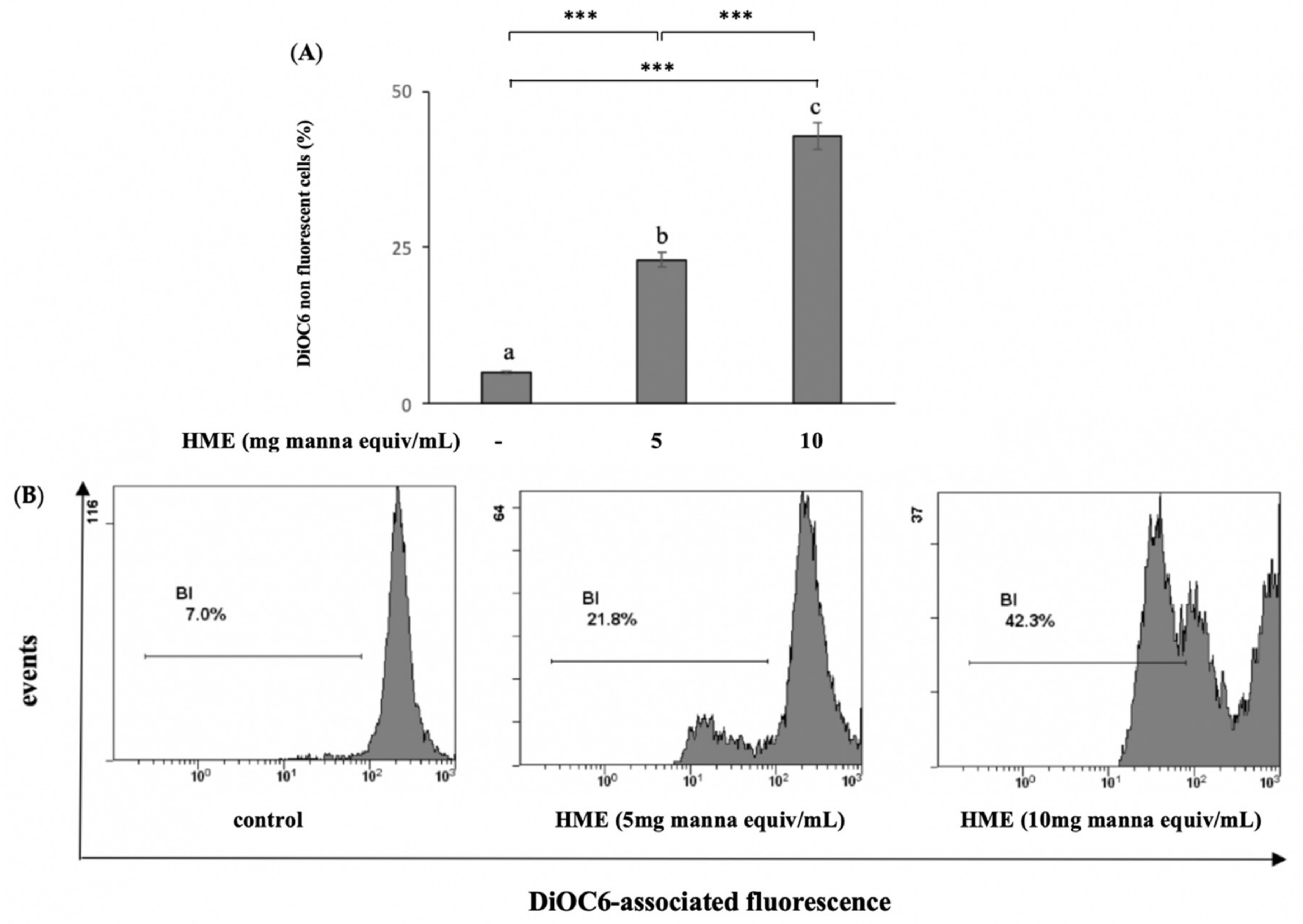
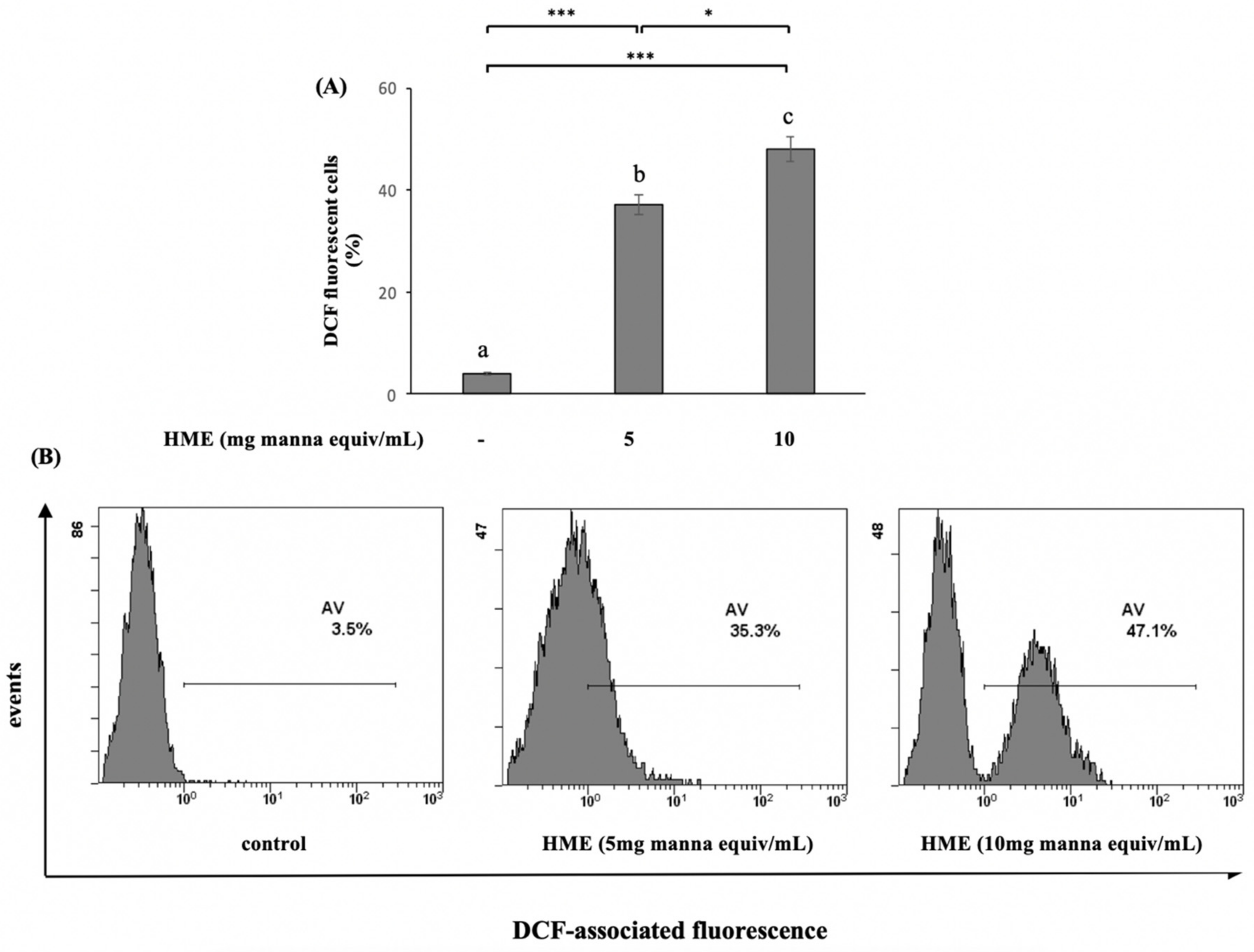
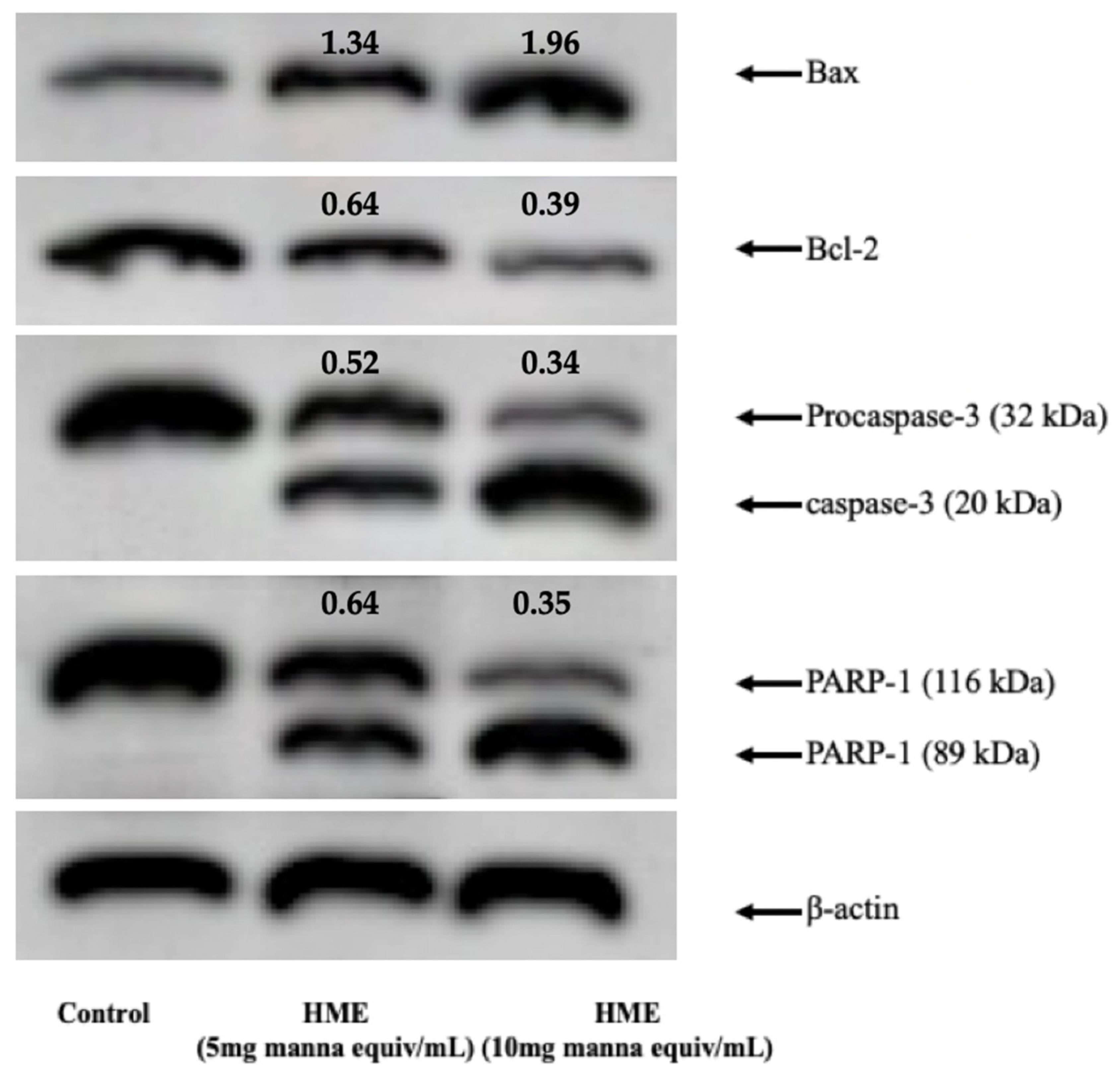
2.2. HME Induces Mitochondrial-Mediated Apoptosis in HCT-116 Cancer Cell Line
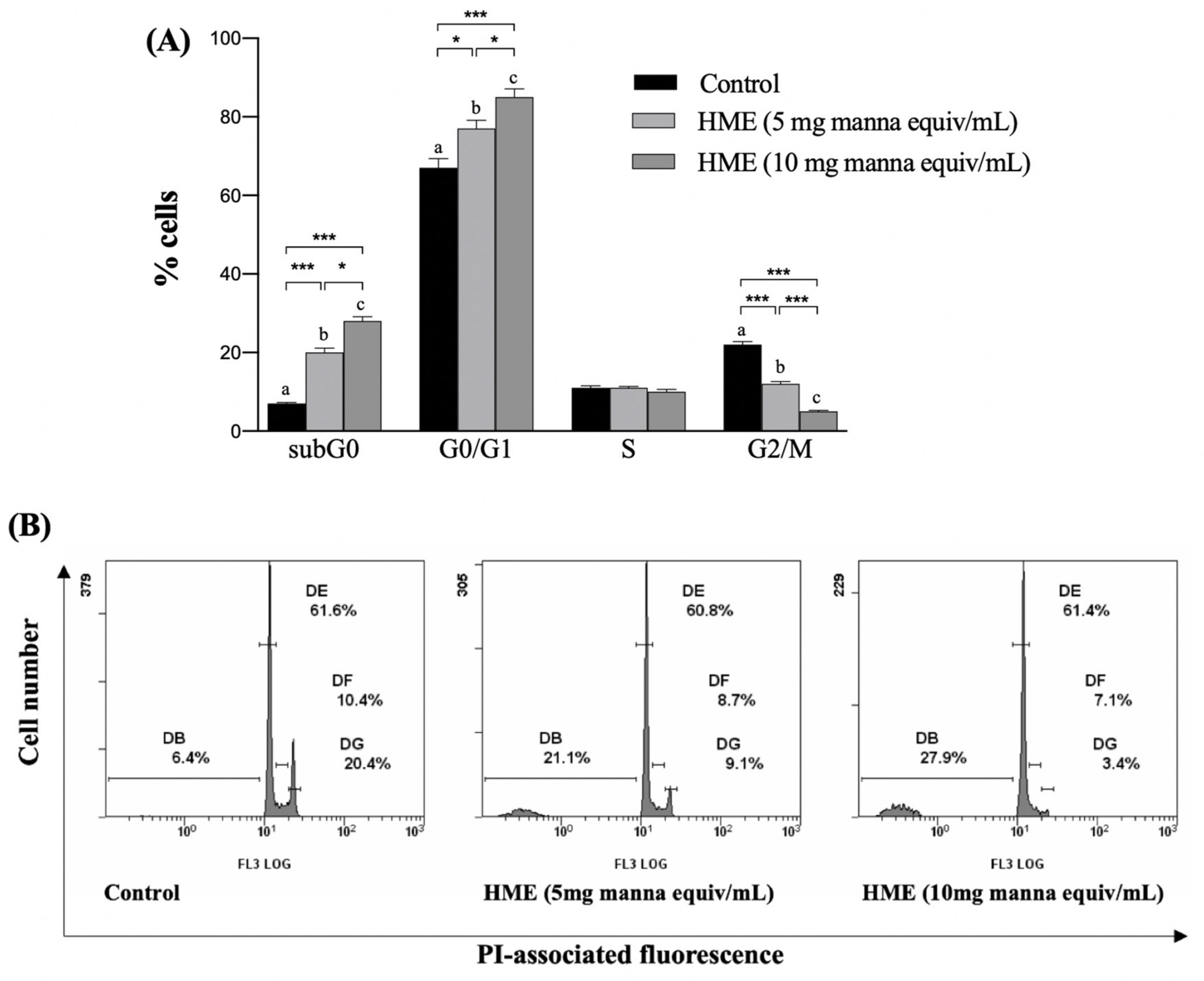
2.3. HME Induces Cell Cycle Arrest in HCT-116 Cancer Cell Line
3. Discussion
4. Materials and Methods
4.1. Hydro-Alcoholic Extract of Manna (HME)
4.2. Cell Culture and Treatment
4.3. Viability Assay
4.4. Flow Cytometry
4.4.1. Measurement of Phosphatidylserine Exposure
4.4.2. Measurement of MMP
4.4.3. Measurement of Intracellular Reactive Oxygen Species (ROS)
4.4.4. Analysis of Cell Cycle
4.5. Western Blotting
4.6. Statistics
Author Contributions
Funding
Conflicts of Interest
References
- Garagnani, P.; Pirazzini, C.; Franceschi, C. Colorectal cancer microenvironment: Among nutrition, gut microbiota, inflammation and epigenetics. Curr. Pharm. Des. 2013, 19, 765–778. [Google Scholar] [CrossRef] [PubMed]
- Ramos, S. Cancer chemoprevention and chemotherapy: Dietary polyphenols and signalling pathways. Mol. Nutr. Food Res. 2008, 52, 507–526. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zheng, J.; Li, Y.; Xu, D.-P.; Li, S.; Chen, Y.-M.; Li, H.-B. Natural polyphenols for prevention and treatment of cancer. Nutrients 2016, 8, 515. [Google Scholar] [CrossRef] [PubMed]
- Schicchi, R.; Camarda, L.; Spadaro, V.; Pitonzo, R. Caratterizzazione chimica della manna estratta nelle Madonie (Sicilia) da cultivar di Fraxinus angustifolia e di Fraxinus ornus (Oleaceae). Quad. Bot. Ambient. Appl. 2006, 17, 151–174. [Google Scholar]
- Oddo, E.; Saiano, F.; Alonzo, G.; Bellini, E. An investigation of the seasonal pattern of mannitol content in deciduous and evergreen species of the Oleaceae growing in northern Sicily. Ann. Bot. 2002, 90, 239–243. [Google Scholar] [CrossRef] [Green Version]
- Attanzio, A.; D’Anneo, A.; Pappalardo, F.; Bonina, F.P.; Livrea, M.A.; Allegra, M.; Tesoriere, L. Phenolic composition of hydrophilic extract of manna from sicilian fraxinus angustifolia vahl and its reducing, antioxidant and anti-inflammatory activity in vitro. Antioxidants 2019, 8, 494. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, D.; Eide, P.W.; Eilertsen, I.A.; Danielsen, S.A.; Eknaes, M.; Hektoen, M.; Lind, G.E.; Lothe, R.A. Epigenetic and genetic features of 24 colon cancer cell lines. Oncogenesis 2013, 2, e71. [Google Scholar] [CrossRef]
- Terzuoli, E.; Giachetti, A.; Ziche, M.; Donnini, S. Hydroxytyrosol, a product from olive oil, reduces colon cancer growth by enhancing epidermal growth factor receptor degradation. Mol. Nutr. Food Res. 2016, 60, 519–529. [Google Scholar] [CrossRef]
- Bassani, B.; Rossi, T.; De Stefano, D.; Pizzichini, D.; Corradino, P.; Macrì, M.; Noonan, D.M.; Albini, A.; Bruno, A. Potential chemopreventive activities of polyphenol rich purified exract from olive mill waste-water on colon cancer cells. J. Funct. Food 2016, 27, 236–248. [Google Scholar] [CrossRef]
- Tan, Y.; Demeter, M.R.; Ruan, H.; Comb, M.J. BAD Ser-155 phosphorylation regulates BAD/Bcl-XL interaction and cell survival. J. Biol. Chem. 2000, 275, 25865–25869. [Google Scholar] [CrossRef] [Green Version]
- Kaufmann, S.H.; Desnoyers, S.; Ottaviano, Y.; Davidson, N.E.; Poirier, G.G. Specific proteolytic cleavage of poly(ADP-ribose) polymerase: An early marker of chemotherapy-induced apoptosis. Cancer Res. 1993, 53, 3976–3985. [Google Scholar] [PubMed]
- Lazebnik, Y.A.; Kaufmann, S.H.; Desnoyers, S.; Poirier, G.G.; Earnshaw, W.C. Cleavage of poly(ADP-ribose) polymerase by a proteinase with properties like ICE. Nature 1994, 371, 346–347. [Google Scholar] [CrossRef] [PubMed]
- MacFarlane, M.; Williams, A. Apoptosis and disease: A life or death decision. EMBO Rep. 2004, 5, 674–678. [Google Scholar] [CrossRef] [Green Version]
- Manson, M. Cancer prevention—the potential for diet to modulate molecular signalling. Trends Mol. Med. 2003, 9, 11–18. [Google Scholar] [CrossRef]
- Watson, W.; Cai, J.; Jones, D. Diet and apoptosis. Annu. Rev. Nutr. 2000, 108, 153–164. [Google Scholar]
- Watson, A.J. Apoptosis and colorectal cancer. Gut 2004, 53, 1701–1709. [Google Scholar] [CrossRef] [Green Version]
- Johnson, I.T. Anticarcinogenic effects of diet-related apoptosis in the colorectal mucosa. Food Chem. Toxicol. 2002, 40, 1171–1178. [Google Scholar] [CrossRef]
- Ahmed, K.; Zaidi, S.F.; Cui, Z.G.; Zhou, D.; Saeed, S.A.; Inadera, H. Potential proapoptotic phytochemical agents for the treatment and prevention of colorectal cancer. Oncol. Lett. 2019, 18, 487–498. [Google Scholar] [CrossRef] [Green Version]
- Hengartner, M.O. The biochemistry of apoptosis. Nature 2000, 407, 770–776. [Google Scholar] [CrossRef]
- Kastan, M.B.; Bartek, J. Cell-cycle checkpoints and cancer. Nature 2004, 432, 316–323. [Google Scholar] [CrossRef]
- Alenzi, F.Q.B. Links between apoptosis proliferation and cell cycle. Br. J. Biomed. Sci. 2004, 61, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.N.; Almoyad, M.H. Polyphenols in colorectal cancer: Current state of knowledge including clinical trials and molecular mechanism of action. BioMed Res. Int. 2018, 2018, 4154186. [Google Scholar] [CrossRef] [Green Version]
- de Kok, T.M.; van Breda, S.G.; Manson, M.M. Mechanisms of combined action of different chemopreventicve dietary compounds: A review. Eur. J. Nutr. 2008, 47, 51–59. [Google Scholar] [CrossRef]
- Das, D.; Arber, N.; Jankowski, J.A. Chemoprevention of colorectal cancer. Digestion 2007, 76, 51–67. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Nadeem, M.; Gilani, S.A.; Khan, S.; Sajid, M.W.; Amir, R.M. Antitumor perspectives of oleuropein and its metabolite hydroxytyrosol: Recent updates. J. Food Sci. 2018, 83, 1781–1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosignoli, P.; Fuccelli, R.; Sepporta, M.V.; Fabiani, R. In vitro chemopreventive activities of hydroxytyrosol: The main phenolic compound present in extra-virgin olive oil. Food Funct. 2016, 7, 301–307. [Google Scholar] [CrossRef]
- Vitaglione, P.; Savarese, M.; Paduano, A.; Scalfi, L.; Fogliano, V.; Sacchi, R. Healthy virgin olive oil: A matter of bitterness. Crit. Rev. Food Sci. Nutr. 2015, 55, 1808–1818. [Google Scholar] [CrossRef]
- Rahmanian, N.; Jafari, S.M.; Wani, T.A. Bioactive profile, dehydration, extraction and application of the bioactive components of olive leaves. Trends Food Sci. Technol. 2015, 42, 150–172. [Google Scholar] [CrossRef]
- Fabiani, R.; De Bartolomeo, A.; Rosignoli, P.; Servili, M.; Selvaggini, R.; Montedoro, G.F.; Di Saverio, C.; Morozzi, G. Virgin olive oil phenols inhibit proliferation of human promyelocytic leukemia cells (HL60) by inducing apoptosis and differentiation. J. Nutr. 2006, 136, 614–619. [Google Scholar] [CrossRef] [Green Version]
- Fabiani, R.; De Bartolomeo, A.; Rosignoli, P.; Servili, M.; Montedoro, G.F.; Morozzi, G. Cancer chemoprevention by hydroxytyrosol isolated from virgin olive oil through G1 cell cycle arrest and apoptosis. Eur. J. Cancer Prev. 2002, 11, 351–358. [Google Scholar] [CrossRef]
- Sarfraz, A.I.; Rasul, A.; Jabeen, F.; Younis, T.; Zahoor, K.M.; Arshad, M.; Ali, M. Fraxinus: A plant with versatile pharmacological and biological activities. Evid. Based Complement. Altern. Med. 2017, 2017, 4269868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wang, L.; Deng, Y.; Zhao, P.; Deng, W.; Zhang, J.; Luo, J.; Li, R. Fraxetin suppresses proliferation of non-small-cell lung cancer cells via preventing activation of signal transducer and activator of transcription 3. Tohoku J. Exp. Med. 2019, 248, 3–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Liu, Z.; Yan, Y.; Wang, H. Effect of fraxetin on proliferation and apoptosis in breast cancer cells. Oncol. Lett. 2017, 14, 7374–7378. [Google Scholar] [CrossRef] [Green Version]
- Maru, G.B.; Hudlikar, R.R.; Kumar, G.; Gandhi, K.; Mahimkar, M.B. Understanding the molecular mechanisms of cancer prevention by dietary phytochemicals: From experimental models to clinical trials. World J. Biol. Chem. 2016, 7, 88–99. [Google Scholar] [CrossRef]
- Kotecha, R.; Takami, A.; Espinoza, J.L. Dietary phytochemicals and cancer chemoprevention: A review of the clinical evidence. Oncotarget 2016, 7, 52517–52529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loeb, L.A.; Harris, C.C. Advances in chemical carcinogenesis: A historical review and prospective. Cancer Res. 2008, 68, 6863–6872. [Google Scholar] [CrossRef] [Green Version]
- Thun, M.; Henley, J.; Gansler, T. Inflammation and cancer: An epidemiological perspective. In Novartis Foundation Symposium; John Wiley: Chichester, UK; New York, NY, USA, 2004; Volume 256, p. 6. [Google Scholar]
- Lu, H.; Ouyang, W.; Huang, C. Inflammation, a key event in cancer development. Mol. Cancer Res. 2006, 4, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.-Y.; Haif, N.; Lotan, R. Apoptosis as a novel target for cancer chemoprevention. J. Nat. Cancer Inst. 2004, 96, 662–672. [Google Scholar] [CrossRef] [Green Version]
- Lukas, M. Inflammatory bowel disease as a risk factor for colorectal cancer. Dig. Dis. 2010, 28, 619–624. [Google Scholar] [CrossRef]
- Triantafillidis, J.K.; Nasioulas, G.; Kosmidis, P.A. Colorectal cancer and inflammatory bowel disease: Epidemiology, risk factors, mechanisms of acrcinogenresis and prevention strategies. Anticancer Res. 2009, 29, 2727–2737. [Google Scholar]
- Itzkovitz, S.H.; Yio, X. Inflammation and cancer IV. Colorectal cancer in inflammatory bowel disease: The role of inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 287, G7–G17. [Google Scholar] [CrossRef]
- Ellis, F.W.; Krantz, J.C. Sugar alcohols: XXII. Metabolism and toxicity studies with mannitol and sorbitol in man and animals. J. Biol. Chem. 1941, 141, 147–154. [Google Scholar]
- Sun, D.X.; Lennernas, H.; Welage, L.S.; Barnett, J.L.; Landowski, C.P.; Foster, D.; Fleischer, D.; Lee, K.D.; Amidon, G.L. Comparison of human duodenum and Caco-2 gene expression profiles for 12,000 gene sequence tags and correlation with permeability of 26 drugs. Pharm. Res. 2002, 19, 1400–1416. [Google Scholar] [CrossRef] [PubMed]
- Allegra, M.; D’Acquisto, F.; Tesoriere, L.; Attanzio, A.; Livrea, M.A. Pro-oxidant activity of indicaxanthin from Opuntia ficus indica modulates arachidonate metabolism and prostaglandin synthesis through lipid peroxide production in LPS-stimulated RAW 264.7 macrophages. Redox Biol. 2014, 2, 892–900. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: Samples of the compounds are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Incubation Time (h) | HCT-116 | Caco-2 | HT-29 |
|---|---|---|---|
| IC50 (mg Manna Equivalent/mL) | |||
| 24 | 10.73 ± 1.22 | 8.51 ± 0.77 | 28.92 ± 1.99 |
| 48 | 6.38 ± 0.51 | 5.88 ± 0.43 | 18.68 ± 1.54 |
| 72 | 4.25 ± 0.31 | 3.84 ± 0.29 | 13.07 ± 1.12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Restivo, I.; Tesoriere, L.; Frazzitta, A.; Livrea, M.A.; Attanzio, A.; Allegra, M. Anti-Proliferative Activity of A Hydrophilic Extract of Manna from Fraxinus angustifolia Vahl through Mitochondrial Pathway-Mediated Apoptosis and Cell Cycle Arrest in Human Colon Cancer Cells. Molecules 2020, 25, 5055. https://doi.org/10.3390/molecules25215055
Restivo I, Tesoriere L, Frazzitta A, Livrea MA, Attanzio A, Allegra M. Anti-Proliferative Activity of A Hydrophilic Extract of Manna from Fraxinus angustifolia Vahl through Mitochondrial Pathway-Mediated Apoptosis and Cell Cycle Arrest in Human Colon Cancer Cells. Molecules. 2020; 25(21):5055. https://doi.org/10.3390/molecules25215055
Chicago/Turabian StyleRestivo, Ignazio, Luisa Tesoriere, Anna Frazzitta, Maria Antonia Livrea, Alessandro Attanzio, and Mario Allegra. 2020. "Anti-Proliferative Activity of A Hydrophilic Extract of Manna from Fraxinus angustifolia Vahl through Mitochondrial Pathway-Mediated Apoptosis and Cell Cycle Arrest in Human Colon Cancer Cells" Molecules 25, no. 21: 5055. https://doi.org/10.3390/molecules25215055
APA StyleRestivo, I., Tesoriere, L., Frazzitta, A., Livrea, M. A., Attanzio, A., & Allegra, M. (2020). Anti-Proliferative Activity of A Hydrophilic Extract of Manna from Fraxinus angustifolia Vahl through Mitochondrial Pathway-Mediated Apoptosis and Cell Cycle Arrest in Human Colon Cancer Cells. Molecules, 25(21), 5055. https://doi.org/10.3390/molecules25215055