
Medicinal Potential of Isoflavonoids: Polyphenols That May Cure Diabetes

 , ,
, ,  ,
,  ,
,
Abstract
:1. Introduction
1.1. Purpose of the Review
1.2. Dietary Sources and Intake of Isoflavonoids
1.3. Biological Importance of Isoflavonoids
2. Discussion
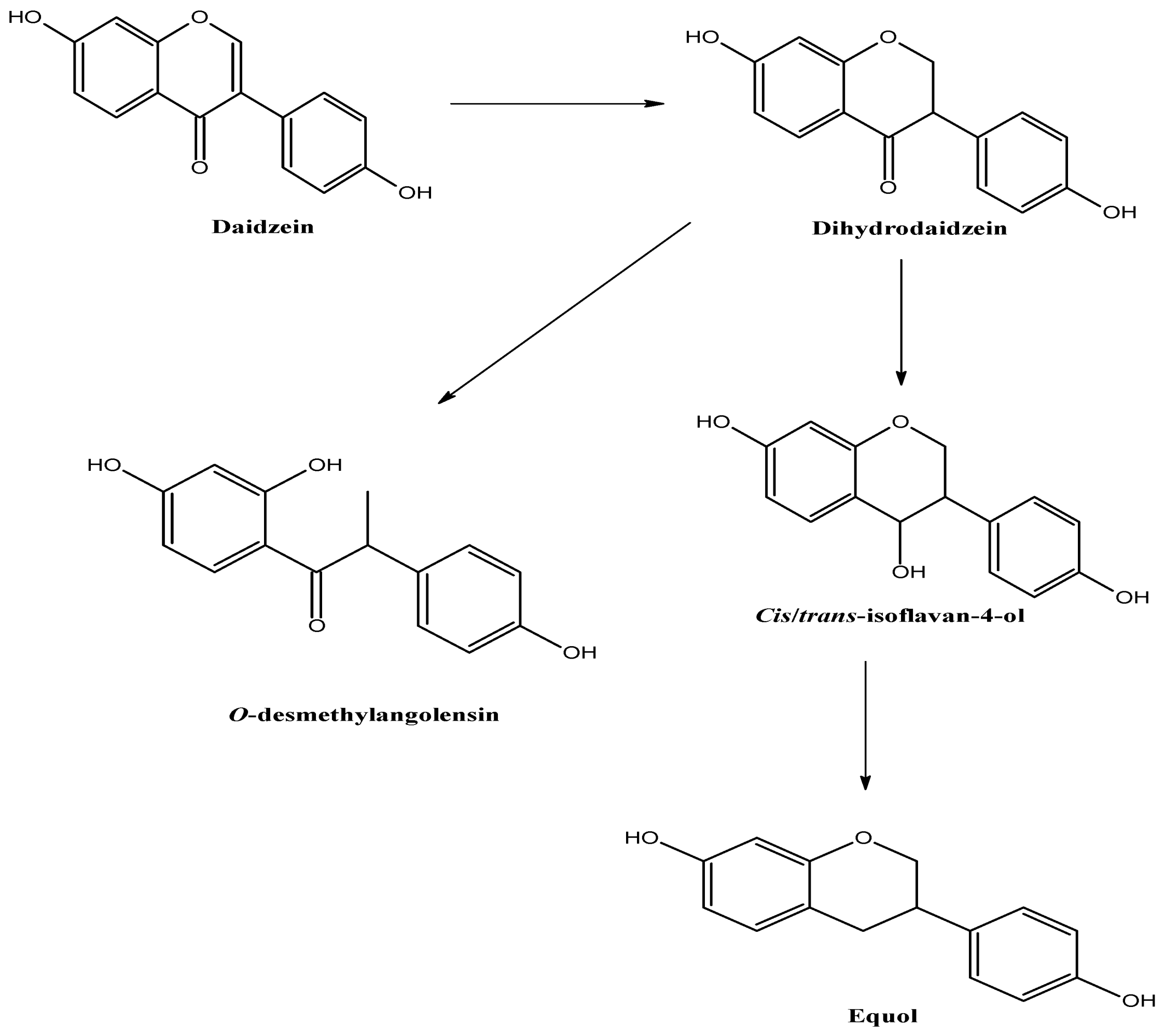
2.1. Metabolism and Bioavailability of Isoflavonoids
2.2. Mode of Action as an Antidiabetic Agent
2.2.1. Peroxisome Proliferator-Activated Receptors α and γ
2.2.2. Antihyperglycemic Effect of Isoflavonoids with Regard to Carbohydrate Metabolism, Insulin Sensitivity and Insulin Resistance, and Preservation of β-Cell Function
2.2.3. Antioxidative Properties of Isoflavonoids in Prevention of Long Term Diabetes Complications
3. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hasan, M.M.; Ahmed, Q.U.; Soad, S.Z.M.; Tunna, T.S. Animal models and natural products to investigate in vivo and in vitro antidiabetic activity. Biomed. Pharm. 2018, 101, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Spranger, J.; Kroke, A.; Möhlig, M.; Hoffmann, K.; Bergmann, M.M.; Ristow, M.; Boeing, H.; Pfeiffer, A.F.H. Inflammatory cytokines and the risk to develop type 2 diabetes: Results of the prospective population-based European Prospective Investigation into Cancer and Nutrition (EPIC)-Potsdam study. Diabetes 2003, 52, 812–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santaguida, P.L.; Balion, C.; Hunt, D. Diagnosis, prognosis, and treatment of impaired glucose tolerance and impaired fasting glucose. Evid. Rep. Technol. Assess. 2008, 12, 1–11. [Google Scholar]
- Mayfield, J. Diagnosis and classification of diabetes mellitus: New criteria. Am. Fam. Physician. 1998, 58, 1355–1362. [Google Scholar]
- Available online: https://www.diabetesatlas.org/en/ (accessed on 14 November 2019).
- Available online: https://www.who.int/news-room/fact-sheets/detail/diabetes (accessed on 8 June 2020).
- Alhassan, M.A.; Ahmed, Q.U.; Latip, J.; Shah, S.A.A. A new sulphated flavone and other phytoconstituents from the leaves of Tetracera indica Merr, and their alpha-glucosidase inhibitory activity. Nat. Prod. Res. 2019, 33, 1–8. [Google Scholar] [CrossRef]
- Lee, H.S. Rat lens aldose reductase inhibitory activities of Coptis japonica root-derived isoquinoline alkaloids. J. Agric. Food Chem. 2002, 50, 7013–7016. [Google Scholar] [CrossRef]
- Matsuda, H.; Morikawa, T.; Yoshikawa, M. Antidiabetogenic constituents from several natural medicines. Pure Appl. Chem. 2002, 74, 1301–1308. [Google Scholar] [CrossRef]
- Teles, R.B.A.; Diniz, T.C.; Pinto, T.C.C.; Júnior, R.G.O.; Silva, M.G.; de Lavor, E.M.; Fernandes, A.W.C.; de Oliveira, A.P.; Ribeiro, F.P.R.D.; da Silva, A.A.M.; et al. Flavonoids as therapeutic agents in alzheimer’s and parkinson’s diseases: A systematic review of preclinical evidences. Oxid. Med. Cell. Longev. 2018, 2018, 7043213. [Google Scholar]
- Dixon, R.A.; Summer, L.W. Legume natural products: Understanding and manipulating complex pathways for human and animal health. Plant. Physiol. 2003, 131, 878–885. [Google Scholar] [CrossRef] [Green Version]
- Miadoková, E. Isoflavonoids—an overview of their biological activities and potential health benefits. Interdiscip. Toxicol. 2009, 2, 211–218. [Google Scholar] [CrossRef]
- Promden, W.; Monthakantirat, O.; Umehara, K.; Noguchi, H.; De-Eknamkul, W. Structure and antioxidant activity relationships of isoflavonoids from Dalbergia parviflora. Molecules 2014, 19, 2226–2237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiter, E.; Beck, V.; Medjakovic, S.; Mueller, M.; Jungbauer, A. Comparison of hormonal activity of isoflavone-containing supplements used to treat menopausal complaints. Menopause 2009, 16, 1049–1060. [Google Scholar] [CrossRef]
- Mazur, W.; Adlercreutz, H. Overview of naturally occurring endocrine-active substances in the human diet in relation to human health. Nutrition 2000, 16, 654–658. [Google Scholar] [CrossRef]
- Takata, Y.; Maskarinec, G.; Franke, A.; Nagata, C.; Shimizu, H. A comparison of dietary habits among women in Japan and Hawaii. Public Health Nutr. 2004, 7, 319–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surh, J.; Kim, M.J.; Koh, E.; Kim, Y.K.L.; Kwon, H. Estimated intakes of isoflavones and coumestrol in Korean population. Int. J. Food Sci. Nutr. 2006, 57, 325–344. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.A.; Wen, W.; Xiang, Y.B.; Barnes, S.; Liu, D.; Cai, Q.; Zheng, W.; Xiao, O.S. Assessment of dietary isoflavone intake among middle-aged Chinese men. J. Nutr. 2007, 137, 1011–1016. [Google Scholar] [CrossRef] [Green Version]
- Horn-Ross, P.L.; John, E.M.; Canchola, A.J.; Stewart, S.L.; Lee, M.M. Phytoestrogen intake and endometrial cancer risk. J. Natl. Cancer Inst. 2003, 95, 1158–1164. [Google Scholar] [CrossRef] [Green Version]
- Chun, O.K.; Chung, S.J.; Song, W.O. Estimated dietary flavonoid intake and major food sources of U.S. adults. J. Nutr. 2007, 137, 1244–1252. [Google Scholar] [CrossRef] [Green Version]
- Mulligan, A.A.; Welch, A.A.; McTaggart, A.A.; Bhaniani, A.; Bingham, S.A. Intakes and sources of soya foods and isoflavones in a UK population cohort study (EPIC-Norfolk). Eur. J. Clin. Nutr. 2007, 61, 248–254. [Google Scholar] [CrossRef] [Green Version]
- Rochfort, S.; Panozzo, J. Phytochemicals for health, the role of pulses. J. Agric. Food Chem. 2007, 55, 7381–7994. [Google Scholar] [CrossRef]
- Jang, C.H.; Lim, J.K.; Kim, J.H.; Park, C.S.; Kwon, D.Y.; Kim, Y.S.; Shin, D.H.; Kim, J.S. Change of Isoflavone Content during Manufacturing of Cheonggukjang, a Traditional Korean Fermented Soyfood. Food Sci. Biotechnol. 2006, 15, 643–646. [Google Scholar]
- Iwasaki, M.; Inoue, M.; Otani, T.; Sasazuki, S.; Kurashi, N.; Miura, T.; Yamoto, S.; Tsugane, S. Plasma isoflavone level and subsequent risk of breast cancer among Japanese women: A nested case-control study from Japan Public Health Cen-ter-base prospective study group. J. Clin. Oncol. 2008, 26, 1677–1683. [Google Scholar] [CrossRef] [PubMed]
- Bezek, S.; Ujhazy, E.; Mach, M.; Navarova, J.; Dubovicky, M. Developmental origin of chronic diseases: Toxicological implication. Interdisc. Toxicol. 2008, 1, 29–31. [Google Scholar] [CrossRef] [Green Version]
- Ørgaard, A.; Jensen, L. The effects of soy isofl avones on obesity. Exp. Biol. Med. 2008, 233, 1066–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabudak, T.; Guler, N.; Trifolium, L. A review on its phytochemical and pharmacological profile. Phytother. Res. 2009, 23, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.J.; Kim, G.H. Antiproliferative activity of daidzein and genistein may be related to ERα/c-erbB-2 expression in human breast cancer cells. Mol. Med. Rep. 2013, 7, 781–784. [Google Scholar] [CrossRef] [Green Version]
- Molteni, A.; Brizio-Molteni, L.; Persky, V. In vitro hormonal effects of soybean isoflavones. J. Nutr. 1995, 125, 751S–756S. [Google Scholar]
- Dixon, R.A. Phytooestrogen. Annu. Rev. Plant. Biol. 2004, 55, 225–261. [Google Scholar] [CrossRef] [Green Version]
- Kawakami, Y.; Tsurugasaki, W.; Nakamura, S.; Osada, K. Comparison of regulative functions between dietary soy isoflavones aglycone and glucoside on lipid metabolism in rats fed cholesterol. J. Nutr. Biochem. 2005, 16, 205–212. [Google Scholar] [CrossRef]
- Yang, W.; Wang, S.; Li, L.; Liang, Z.; Wang, L. Genistein reduces hyperglycemia and islet cell loss in a high-dosage manner in rats with alloxan-induced pancreatic damage. Pancreas 2011, 40, 396–402. [Google Scholar] [CrossRef]
- Cederroth, C.R.; Nef, S. Soy, phytoestrogens and metabolism: A review. Mol. Cell. Endocrinol. 2009, 304, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Setchell, K.D.R.; Brown, N.M.; Zimmer-Nechemias, L.; Brashear, W.T.; Wolfe, B.E.; Kirschner, A.S.; Heubi, J.E. Evidence for lack of absorption of soy isoflavone glycosides in humans, supporting the crucial role of intestinal metabolism for bioavailability. Am. J. Clin. Nutr. 2002, 76, 447–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, V.; Lee, S.O.; Murphy, P.A.; Hendrich, S.; Verbruggen, M.A. The apparent absorptions of isoflavone glucosides and aglucons are similar in women and are increased by rapid gut transit time and low fecal isoflavone degradation. J. Nutr. 2004, 134, 2534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zubik, L.; Meydani, M. Bioavailability of soybean isoflavones from aglycone and glucoside forms in American women. Am. J. Clin. Nutr. 2003, 77, 1459–1465. [Google Scholar] [CrossRef] [PubMed]
- Izumi, T.; Piskula, M.K.; Osawa, S.; Obata, A.; Tobe, K.; Saito, M.; Kataoka, S.; Kubota, Y.; Kikuchi, M. Soy isoflavone aglycones are absorbed faster and in higher amounts than their glucosides in humans. J. Nutr. 2000, 130, 1695–1699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kano, M.; Takayanagi, T.; Harada, K.; Sawada, S.; Ishikawa, F. Bioavailability of isoflavones after ingestion of soy beverages in healthy adults. J. Nutr. 2006, 136, 2291–2296. [Google Scholar] [CrossRef] [PubMed]
- Tsunoda, N.; Pomeroy, S.; Nestel, P. Absorption in humans of isoflavones from soy and red clover is similar. J. Nutr. 2002, 132, 2199. [Google Scholar] [CrossRef] [Green Version]
- Day, A.J.; Dupont, M.S.; Rhodes, M.J.C.; Morgan, M.R.A.; Williamson, G.; Ridley, S.; Rhodes, M. Deglycosylation of flavonoid and isoflavonoid glycosides by human small intestine and liver-glucosidase activity. FEBS Lett. 1998, 436, 71. [Google Scholar] [CrossRef] [Green Version]
- Hwang, C.S.; Kwak, H.S.; Lim, H.J.; Lee, S.H.; Kang, Y.S.; Choe, T.B.; Hur, H.G.; Han, K.O. Isoflavone metabolites and their in vitro dual functions: They can act as an estrogenic agonist or antagonist depending on the estrogen concentration. J. Steroid Biochem. Mol. Biol. 2006, 101, 246–253. [Google Scholar] [CrossRef]
- Heinonen, S.; Wahala, K.; Adlercreutz, H. Identification of isoflavone metabolites dihydrodaidzein, dihydrogenistein, 6′-OH-O-dma, and cis-4-OH-equol in human urine by Gas Chromatography-Mass Spectroscopy using authentic reference compounds. Anal. Biochem. 1999, 274, 211–219. [Google Scholar] [CrossRef]
- Loureiro, G.; Martel, F. The effect of dietary polyphenols on intestinal absorption of glucose and fructose: Relation with obesity and type 2 diabetes. J. Food Rev. Int. 2019, 35, 390–406. [Google Scholar] [CrossRef]
- Sun, L.; Miao, M. Dietary polyphenols modulate starch digestion and glycaemic level: A review. Crit. Rev. Food. Sci. Nutr. 2020, 60, 541–555. [Google Scholar] [CrossRef]
- Svjetlana, M.; Monika, M.; Alois, J. Potential Health-modulating effects of isoflavones and metabolites via activation of PPAR and AhR. Nutrients 2010, 2, 241–279. [Google Scholar]
- Wang, Y.; Han, Y.; Teng, W.; Zhao, X.; Li, Y.; Wu, L.; Li, D.; Li, W. Expression quantitative trait loci infer the regulation of isoflavone accumulation in soybean (Glycine max L. Merr.) seed. BMC Genom. 2014, 15, 680. [Google Scholar] [CrossRef] [Green Version]
- Na, H.K.; Surh, Y.J. Peroxisome proliferator-activated receptor gamma (PPARgamma) ligands as bifunctional regulators of cell proliferation. Biochem. Pharm. 2003, 66, 1381–1391. [Google Scholar] [CrossRef]
- Heikkinen, S.; Auwerx, J.; Argmann, C.A. PPARgamma in human and mouse physiology. Biochim. Biophys. Acta 2007, 1771, 999–1013. [Google Scholar] [CrossRef] [Green Version]
- Tontonoz, P.; Spiegelman, B.M. Fat and beyond: The diverse biology of PPARγ. Annu. Rev. Biochem. 2008, 77, 289–312. [Google Scholar] [CrossRef]
- Shen, P.; Liu, M.H.; Ng, T.Y.; Chan, Y.H.; Yong, E.L. Differential effects of isoflavones, from Astragalus membranaceus and Pueraria thomsonii, on the activation of PPARalpha, PPARgamma, and adipocyte differentiation in vitro. J. Nutr. 2006, 136, 899–905. [Google Scholar] [CrossRef]
- Mezei, O.; Banz, W.J.; Steger, R.W.; Peluso, M.R.; Winters, T.A.; Shay, N. Soy isoflavones exert antidiabetic and hypolipidemic effects through the PPAR pathways in obese Zucker rats and murine RAW 264.7 cells. J. Nutr. 2003, 133, 1238–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, L.; Ye, H.; Chen, L.; Hong, Y.; Zhong, F.; Zhang, T. Red clover extract ameliorates dyslipidemia in streptozotocin-induced diabetic C57BL/6 mice by activating hepatic PPARα. Phytother. Res. 2012, 26, 860–864. [Google Scholar] [CrossRef] [PubMed]
- Hoo, R.L.; Wong, J.Y.; Qiao, C.; Xu, A.; Xu, H.; Lam, K.S. The effective fraction isolated from Radix astragali alleviates glucose intolerance, insulin resistance and hypertriglyceridemia in db/db diabetic mice through its anti-inflammatory activity. Nutr. Metab. 2010, 7, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, M.K.; Roufogalis, B.D.; Huang, T.H.W. Modulation of diabetic retinopathy pathophysiology by natural medicines through PPAR-γ-related pharmacology. Br. J. Pharm. 2012, 165, 4–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, K.W.; Lee, O.H.; Banz, W.J.; Moustaid-Moussa, N.; Shay, N.F.; Kim, Y.C. Daidzein and the daidzein metabolite, equol, enhance adipocyte differentiation and PPARgamma transcriptional activity. J. Nutr. Biochem. 2010, 21, 841–847. [Google Scholar] [CrossRef] [PubMed]
- Cheong, S.H.; Furuhashi, K.; Ito, K.; Nagaoka, M.; Yonezawa, T.; Miura, Y.; Yagasaki, K. Antihyperglycemic effect of equol, a daidzein derivative, in cultured L6 myocytes and ob/ob mice. Mol. Nutr. Food Res. 2014, 58, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.Y.; Hong, S.M.; Ahn, I.S.; Kim, M.J.; Yang, H.J.; Park, S. Isoflavonoids and peptides from meju, long-term fermented soybeans, increase insulin sensitivity and exert insulinotropic effects in vitro. Nutrition 2011, 27, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Carrara, V.S.; Amato, A.A.; Neves, F.A.; Bazotte, R.B.; Mandarino, J.M.; Nakamura, C.V.; Filho, B.P.; Cortez, D.A. Effects of a methanolic fraction of soybean seeds on the transcriptional activity of peroxisome proliferator-activated receptors (PPAR). Braz. J. Med. Biol. Res. 2009, 42, 545–550. [Google Scholar] [CrossRef] [Green Version]
- Dinneen, S.; Gerich, J.; Rizza, R. Carbohydrate metabolism in non-insulin-dependent diabetes mellitus. N. Engl. J. Med. 1992, 327, 707–713. [Google Scholar] [CrossRef]
- Lee, D.S.; Lee, S.H. Genistein, a soy isoflavone, is a potent alpha-glucosidase inhibitor. FEBS Lett. 2001, 501, 84–86. [Google Scholar] [CrossRef] [Green Version]
- Choi, C.W.; Choi, Y.H.; Cha, M.R.; Yoo, D.S.; Kim, Y.S.; Yon, G.H.; Hong, K.S.; Kim, Y.H.; Ryu, S.Y. Yeast α-glucosidase inhibition by isoflavones from plants of Leguminosae as an in vitro alternative to acarbose. J. Agric. Food Chem. 2010, 58, 9988–9993. [Google Scholar] [CrossRef]
- Asgary, S.; Naderi, G.A.; Zadegan, N.S.; Vakili, R. The Inhibitory effects of pure flavonoids on in vitro protein glycosylation. J. Herb. Pharm. 2002, 2, 47–55. [Google Scholar] [CrossRef]
- Harini, R.; Ezhumalai, M.; Pugalendi, K.V. Antihyperglycemic effect of biochanin A, a soy isoflavone, on streptozotocin-diabetic rats. Eur. J. Pharmacol. 2012, 676, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Azizi, R.; Goodarzi, M.T.; Salemi, Z. Effect of biochanin A on serum visfatin level of streptozocin-induced diabetic rats. Iran. Red Crescent Med. J. 2014, 16, e15424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vedavanam, K.; Srijayanta, S.; O’Reilly, J.; Raman, A.; Wiseman, H. Antioxidant action and potential antidiabetic properties of an isoflavonoid-containing soyabean phytochemical extract (SPE). Phytother. Res. 1999, 13, 601–608. [Google Scholar] [CrossRef]
- Higashi, K.; Ogawara, H. Daidzein inhibits insulin- or insulin-like growth factor-1-mediated signaling in cell cycle progression of Swiss 3T3 cells. Biochim. Biophys. Acta 1994, 1221, 29–35. [Google Scholar] [CrossRef]
- Kwon, D.Y.; Jang, J.S.; Lee, J.E.; Kim, Y.S.; Shin, D.H.; Park, S. The isoflavonoid aglycone-rich fractions of Chungkookjang, fermented unsalted soybeans, enhance insulin signaling and peroxisome proliferator-activated receptor-gamma activity in vitro. Biofactors 2006, 26, 245–258. [Google Scholar] [CrossRef]
- Oh, T.W.; Kim, Y.A.; Jang, W.J.; Byeon, J.I.; Ryu, C.H.; Kim, J.O.; Ha, Y.L. Semipurified fractions from the submerged-culture broth of Agaricus blazei Murill reduce blood glucose levels in streptozotocin-induced diabetic rats. J. Agric. Food Chem. 2010, 58, 4113–4119. [Google Scholar] [CrossRef]
- Cheong, S.H.; Furuhashi, K.; Ito, K.; Nagaoka, M.; Yonezawa, T.; Miura, Y.; Yagasaki, K. Daidzein promotes glucose uptake through glucose transporter 4 translocation to plasma membrane in L6 myocytes and improves glucose homeostasis in Type 2 diabetic model mice. J. Nutr. Biochem. 2014, 25, 136–143. [Google Scholar] [CrossRef]
- Getek, M.; Czech, N.; Muc-Wierzgoń, M.; Grochowska-Niedworok, E.; Kokot, T.; Nowakowska-Zajdel, E. The active role of leguminous plant components in type 2 diabetes. Evid. Based Complement. Alternat. Med. 2014, 2014, 293961. [Google Scholar] [CrossRef]
- Lee, D.; Lee, D.H.; Choi, S.; Lee, J.S.; Jang, D.K.; Kang, K.S. Identification and isolation of active compounds from Astragalus membranaceus that improve insulin secretion by regulating pancreatic β-cell metabolism. Biomolecules 2019, 9, 618. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Shi, X.; Wei, Y.; Qin, L.; Sun, W.; Xu, G.; Xu, T.; Liu, T. Synthesis and biological activity of isoflavone derivatives from chickpea as potent anti-diabetic agents. Molecules 2015, 20, 17016–17040. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.; Li, P.; Li, B.; Gao, J.; Wang, D.; Qin, L.; Sun, W.; Xu, Y.; Shi, H.; Xu, T.; et al. Study of the hypoglycemic activity of derivatives of isoflavones from Cicer arietinum L. Evid. Based Complement. Altern. Med. 2017, 2017, 8746823. [Google Scholar] [CrossRef] [PubMed]
- Agyemang, K.; Han, L.; Liu, E.; Zhang, Y.; Wang, T.; Gao, X. Recent advances in Astragalus membranaceus antidiabetic research: Pharmacological effects of its phytochemical constituents. Evid. Based Complement. Altern. Med. 2013, 2013, 654643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazuine, M.; van den Broek, P.J.; Maassen, J.A. Genistein directly inhibits GLUT4-mediated glucose uptake in 3T3-L1 adipocytes. Biochem. Biophys. Res. Commun. 2005, 326, 511–514. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Kim, C.H.; Hoang, D.M.; Kim, B.Y.; Sohn, C.B.; Kim, M.R.; Ahn, J.S. Genistein-derivatives from Tetracera scandens stimulate glucose-uptake in L6 myotubes. Biol. Pharm. Bull. 2009, 32, 504–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, R.M.; Tiesinga, J.J.; Shah, N.; Smith, J.A.; Jarett, L. Genistein inhibits insulin-stimulated glucose transport and decreases immunocytochemical labeling of GLUT4 carboxyl-terminus without affecting translocation of GLUT4 in isolated rat adipocytes: Additional evidence of GLUT4 activation by insulin. Arch. Biochem. Biophys. 1993, 300, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Nomura, M.; Takahashi, T.; Nagata, N.; Tsutsumi, K.; Kobayashi, S.; Akiba, T.; Yokogawa, K.; Moritani, S.; Miyamoto, K. Inhibitory mechanisms of flavonoids on insulin-stimulated glucose uptake in MC3T3-G2/PA6 adipose cells. Biol. Pharm. Bull. 2008, 31, 1403–1409. [Google Scholar] [CrossRef] [Green Version]
- Jonas, J.C.; Plant, T.D.; Gilon, P.; Detimary, P.; Nenquin, M.; Henquin, J.C. Multiple effects and stimulation of insulin secretion by the tyrosine kinase inhibitor genistein in normal mouse islets. Br. J. Pharm. 1995, 114, 872–880. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Zhen, W.; Yang, Z.; Carter, J.D.; Si, H.; Reynolds, K.A. Genistein acutely stimulates insulin secretion in pancreatic beta-cells through a cAMP-dependent protein kinase pathway. Diabetes 2006, 55, 1043–1050. [Google Scholar] [CrossRef] [Green Version]
- Ohno, T.; Kato, N.; Ishii, C.; Shimizu, M.; Ito, Y.; Tomono, S.; Kawazu, S. Genistein augments cyclic adenosine 3′5′-monophosphate (cAMP) accumulation and insulin release in MIN6 cells. Endocr. Res. 1993, 19, 273–285. [Google Scholar] [CrossRef]
- Easom, R.A. CaM kinase II: A protein kinase with extraordinary talents germane to insulin exocytosis. Diabetes 1999, 48, 675–684. [Google Scholar] [CrossRef]
- Fu, Z.; Liu, D. Long-term exposure to genistein improves insulin secretory function of pancreatic beta-cells. Eur. J. Pharmacol. 2009, 616, 321–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neye, H.; Verspohl, E.J. The specificity of tyrosine kinase inhibitors: Their effect on insulin release (short-term effect) and insulin mRNA (long-term effect) in an insulin-secreting cell line (INS-1). Exp. Clin. Endocrinol. Diabetes 1998, 106, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, E.R.; Liu, D. Anti-diabetic functions of soy isoflavone genistein: Mechanisms underlying its effects on pancreatic β-cell function. Food Funct. 2013, 4, 200–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abler, A.; Smith, J.A.; Randazzo, P.A.; Rothenberg, P.L.; Jarett, L. Genistein differentially inhibits postreceptor effects of insulin in rat adipocytes without inhibiting the insulin receptor kinase. J. Biol. Chem. 1992, 267, 3946–3951. [Google Scholar]
- Relic, B.; Zeddou, M.; Desoroux, A.; Beguin, Y.; de Seny, D.; Malaise, M.G. Genistein induces adipogenesis but inhibits leptin induction in human synovial fibroblasts. Lab. Investig. 2009, 89, 811–822. [Google Scholar] [CrossRef] [Green Version]
- Szkudelski, T.; Nogowski, L.; Pruszyńska-Oszmałek, E.; Kaczmarek, P.; Szkudelska, K. Genistein restricts leptin secretion from rat adipocytes. J. Steroid Biochem. Mol. Biol. 2005, 96, 301–307. [Google Scholar] [CrossRef]
- Szkudelska, K.; Nogowski, L.; Szkudelski, T. Genistein, a plant-derived isoflavone, counteracts the antilipolytic action of insulin in isolated rat adipocytes. J. Steroid Biochem. Mol. Biol. 2008, 109, 108–114. [Google Scholar] [CrossRef]
- Chen, Q.C.; Zhang, W.Y.; Jin, W.; Lee, I.S.; Min, B.S.; Jung, H.J.; Na, M.; Lee, S.; Bae, K. Flavonoids and isoflavonoids from Sophorae flos improve glucose uptake in vitro. Planta Med. 2010, 76, 79–81. [Google Scholar] [CrossRef]
- Fu, Z.; Zhang, W.; Zhen, W.; Lum, H.; Nadler, J.; Bassaganya-Riera, J.; Jia, Z.; Wang, Y.; Misra, H.; Liu, D. Genistein induces pancreatic beta-cell proliferation through activation of multiple signaling pathways and prevents insulin-deficient diabetes in mice. Endocrinology 2010, 151, 3026–3037. [Google Scholar] [CrossRef] [Green Version]
- Matveyenko, A.V.; Butler, P.C. Relationship between β-cell mass and diabetes onset. Diabetes Obes. Metab. 2008, 10, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.; Gilbert, E.R.; Pfeiffer, L.; Zhang, Y.; Fu, Y.; Liu, D. Genistein ameliorates hyperglycemia in a mouse model of nongenetic type 2 diabetes. Appl. Physiol. Nutr. Metab. 2012, 37, 480–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Gao, X.J.; Zhao, W.W.; Zhao, W.J.; Jiang, C.H.; Huang, F.; Kou, J.P.; Liu, B.L.; Liu, K. Opposite effects of genistein on the regulation of insulin-mediated glucose homeostasis in adipose tissue. Br. J. Pharm. 2013, 170, 328–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, M.S.; Jung, U.J.; Yeo, J.; Kim, M.J.; Lee, M.K. Genistein and daidzein prevent diabetes onset by elevating insulin level and altering hepatic gluconeogenic and lipogenic enzyme activities in non-obese diabetic (NOD) mice. Diabetes Metab. Res. Rev. 2008, 24, 74–81. [Google Scholar] [CrossRef]
- Moran, J.; Garrido, P.; Cabello, E.; Alonso, A.; González, C. Effects of estradiol and genistein on the insulin signaling pathway in the cerebral cortex of aged female rats. Exp. Gerontol. 2014, 58, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Rauter, A.P.; Martins, A.; Borges, C.; Mota-Filipe, H.; Pinto, R.; Sepodes, B.; Justino, J. Antihyperglycaemic and protective effects of flavonoids on streptozotocin-induced diabetic rats. Phytother. Res. 2010, 24, S133–S138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jesus, A.R.; Dias, C.; Matos, A.M.; de Almeida, R.F.; Viana, A.S.; Marcelo, F.; Ribeiro, R.T.; Macedo, M.P.; Airoldi, C.; Nicotra, F.; et al. Exploiting the therapeutic potential of 8-β-D-glucopyranosylgenistein: Synthesis, antidiabetic activity, and molecular interaction with islet amyloid polypeptide and amyloid β-peptide (1-42). J. Med. Chem. 2014, 57, 9463–9472. [Google Scholar] [CrossRef]
- Al-Nakkash, L.; Markus, B.; Batia, L.; Prozialeck, W.C.; Broderick, T.L. Genistein induces estrogen-like effects in ovariectomized rats but fails to increase cardiac GLUT4 and oxidative stress. J. Med. Food 2010, 13, 1369–1375. [Google Scholar] [CrossRef] [Green Version]
- Nogowski, L.; Nowak, K.W.; Kaczmarek, P.; Maćkowiak, P. The influence of coumestrol, zearalenone, and genistein administration on insulin receptors and insulin secretion in ovariectomized rats. J. Recept. Signal. Transduct. Res. 2002, 22, 449–457. [Google Scholar] [CrossRef]
- Mackowiak, P.; Nogowski, L.; Nowak, K.W. Effect of isoflavone genistein on insulin receptors in perfused liver of ovariectomized rats. J. Recept. Signal. Transduct. Res. 1999, 19, 283–292. [Google Scholar] [CrossRef]
- Ae Park, S.; Choi, M.S.; Cho, S.Y.; Seo, J.S.; Jung, U.J.; Kim, M.J.; Sung, M.K.; Park, Y.B.; Lee, M.K. Genistein and daidzein modulate hepatic glucose and lipid regulating enzyme activities in C57BL/KsJ-db/db mice. Life Sci. 2006, 79, 1207–1213. [Google Scholar] [CrossRef]
- Weigt, C.; Hertrampf, T.; Kluxen, F.M.; Flenker, U.; Hülsemann, F.; Fritzemeier, K.H.; Diel, P. Molecular effects of ER alpha- and beta-selective agonists on regulation of energy homeostasis in obese female Wistar rats. Mol. Cell. Endocrinol. 2013, 377, 147–158. [Google Scholar] [CrossRef]
- Arunkumar, E.; Anuradha, C.V. Genistein promotes insulin action through adenosine monophosphate-activated protein kinase activation and p70 ribosomal protein S6 kinase 1 inhibition in the skeletal muscle of mice fed a high energy diet. Nutr. Res. 2012, 32, 617–625. [Google Scholar] [CrossRef]
- Park, M.H.; Ju, J.W.; Park, M.J.; Han, J.S. Daidzein inhibits carbohydrate digestive enzymes in vitro and alleviates postprandial hyperglycemia in diabetic mice. Eur. J. Pharm. 2013, 712, 48–52. [Google Scholar] [CrossRef]
- Seo, S.G.; Yang, H.; Shin, S.H.; Min, S.; Kim, Y.A.; Yu, J.G.; Lee, D.E.; Chung, M.; Heo, Y.; Kwon, J.Y.; et al. A metabolite of daidzein, 6,7,4′-trihydroxyisoflavone, suppresses adipogenesis in 3T3-L1 preadipocytes via ATP-competitive inhibition of PI3K. Mol. Nutr. Food Res. 2013, 57, 1446–1455. [Google Scholar] [CrossRef]
- Bhathena, S.J.; Velasquez, M.T. Beneficial role of dietary phytoestrogens in obesity and diabetes. Am. J. Clin. Nutr. 2002, 76, 1191–1201. [Google Scholar] [CrossRef] [Green Version]
- Zang, Y.; Igarashi, K.; Yu, C. Anti-obese and anti-diabetic effects of a mixture of daidzin and glycitin on C57BL/6J mice fed with a high-fat diet. Biosci. Biotechnol. Biochem. 2015, 79, 117–123. [Google Scholar] [CrossRef]
- Kruk, I.; Aboul-Enein, H.Y.; Michalska, T.; Lichszteld, K.; Kładna, A. Scavenging of reactive oxygen species by the plant phenols genistein and oleuropein. Luminescence 2005, 20, 81–89. [Google Scholar] [CrossRef]
- Persaud, S.J.; Harris, T.E.; Burns, C.J.; Jones, P.M. Tyrosine kinases play a permissive role in glucose-induced insulin secretion from adult rat islets. J. Mol. Endocrinol. 1999, 22, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.Y.; Shaw, N.S.; Tsai, K.S.; Chen, C.Y. The hypoglycemic effects of soy isoflavones on postmenopausal women. J. Womens Health 2004, 13, 1080–1086. [Google Scholar] [CrossRef]
- Bitto, A.; Altavilla, D.; Bonaiuto, A.; Polito, F.; Minutoli, L.; Di Stefano, V.; Giuliani, D.; Guarini, S.; Arcoraci, V.; Squadrito, F. Effects of aglycone genistein in a rat experimental model of postmenopausal metabolic syndrome. J. Endocrinol. 2009, 200, 367–376. [Google Scholar] [CrossRef] [Green Version]
- Salih, M.; Nallasamy, P.; Muniyandi, P.; Periyasami, V.; Venkatraman, A.C. Genistein improves liver function and attenuates non-alcoholic fatty liver disease in a rat model of insulin resistance. J. Diabetes. 2009, 1, 278–287. [Google Scholar] [CrossRef]
- Lee, J.S. Effects of soy protein and genistein on blood glucose, antioxidant enzyme activities, and lipid profile in streptozotocin-induced diabetic rats. Life Sci. 2006, 79, 1578–1584. [Google Scholar] [CrossRef]
- Horiuchi, H.; Harada, N.; Adachi, T.; Nakano, Y.; Inui, H.; Yamaji, R. S-equol enantioselectively activates cAMP-protein kinase A signaling and reduces alloxan-induced cell death in INS-1 pancreatic β-cells. J. Nutr. Sci. Vitaminol. 2014, 60, 291–296. [Google Scholar] [CrossRef] [Green Version]
- Ateba, S.B.; Mvondo, M.A.; Djiogue, S.; Zingué, S.; Krenn, L.; Njamen, D. A Pharmacological overview of alpinumisoflavone, a natural prenylated isoflavonoid. Front. Pharm. 2019, 10, 952. [Google Scholar] [CrossRef]


{kind=link}
| No | Isoflavonoids | Biological Structure |
|---|---|---|
| 1 | Biochanin A |  |
| 2 | Genistein |  |
| 3 | Daidzein |  |
| 4 | Glycitein |  |
| 5 | Formononetin |  |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, Q.U.; Ali, A.H.M.; Mukhtar, S.; Alsharif, M.A.; Parveen, H.; Sabere, A.S.M.; Nawi, M.S.M.; Khatib, A.; Siddiqui, M.J.; Umar, A.; et al. Medicinal Potential of Isoflavonoids: Polyphenols That May Cure Diabetes. Molecules 2020, 25, 5491. https://doi.org/10.3390/molecules25235491
Ahmed QU, Ali AHM, Mukhtar S, Alsharif MA, Parveen H, Sabere ASM, Nawi MSM, Khatib A, Siddiqui MJ, Umar A, et al. Medicinal Potential of Isoflavonoids: Polyphenols That May Cure Diabetes. Molecules. 2020; 25(23):5491. https://doi.org/10.3390/molecules25235491
Chicago/Turabian StyleAhmed, Qamar Uddin, Abdul Hasib Mohd Ali, Sayeed Mukhtar, Meshari A. Alsharif, Humaira Parveen, Awis Sukarni Mohmad Sabere, Mohamed Sufian Mohd. Nawi, Alfi Khatib, Mohammad Jamshed Siddiqui, Abdulrashid Umar, and et al. 2020. "Medicinal Potential of Isoflavonoids: Polyphenols That May Cure Diabetes" Molecules 25, no. 23: 5491. https://doi.org/10.3390/molecules25235491
APA StyleAhmed, Q. U., Ali, A. H. M., Mukhtar, S., Alsharif, M. A., Parveen, H., Sabere, A. S. M., Nawi, M. S. M., Khatib, A., Siddiqui, M. J., Umar, A., & Alhassan, A. M. (2020). Medicinal Potential of Isoflavonoids: Polyphenols That May Cure Diabetes. Molecules, 25(23), 5491. https://doi.org/10.3390/molecules25235491