
Methods for Studying Endocytotic Pathways of Herpesvirus Encoded G Protein-Coupled Receptors

 , , ,
, , ,
Abstract
:1. Importance of Endocytosis for Viral GPCRs
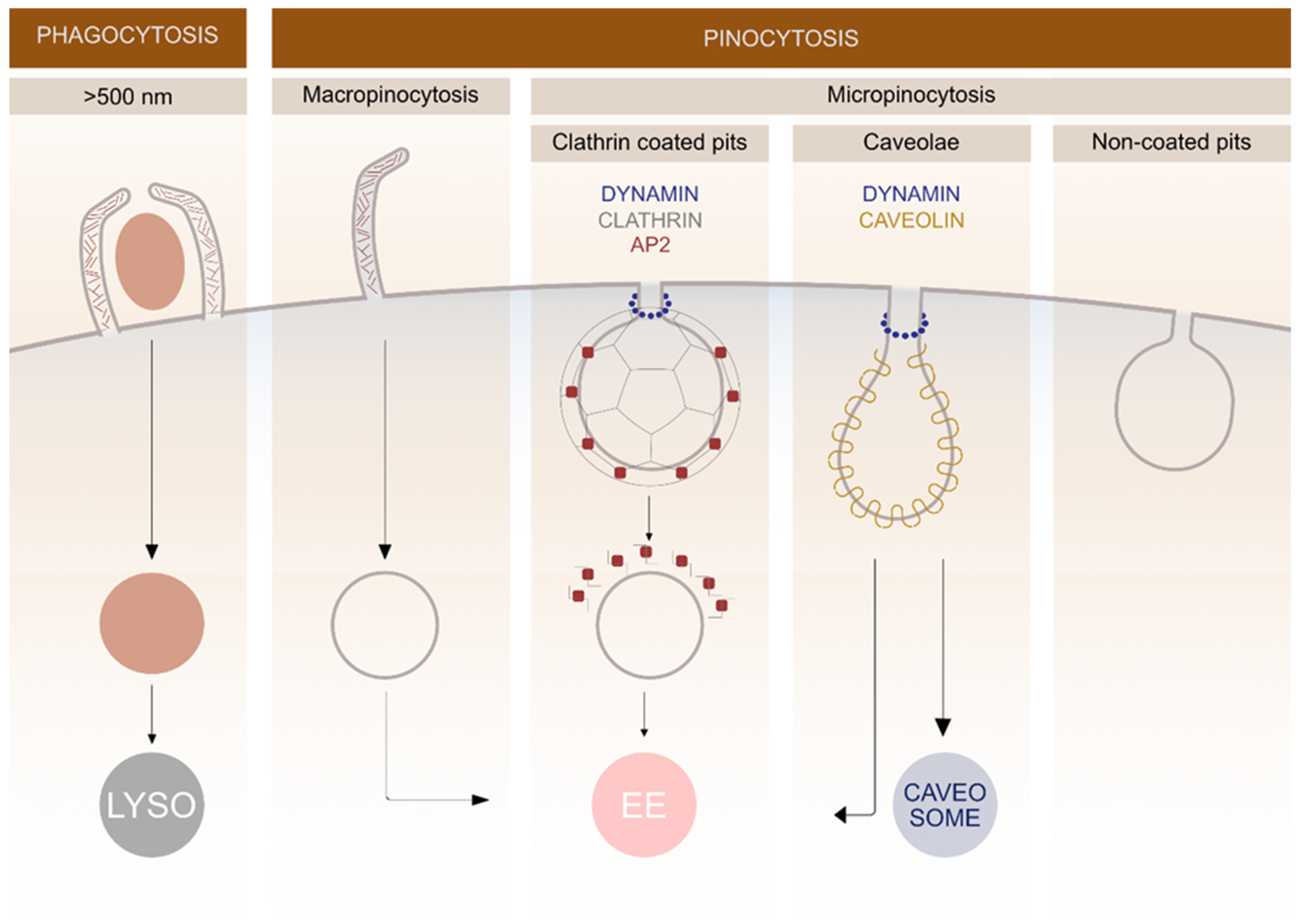
2. Different Endocytic Pathways
2.1. Clathrin-Mediated Pathway
2.2. Caveolae
2.3. Lipid Rafts
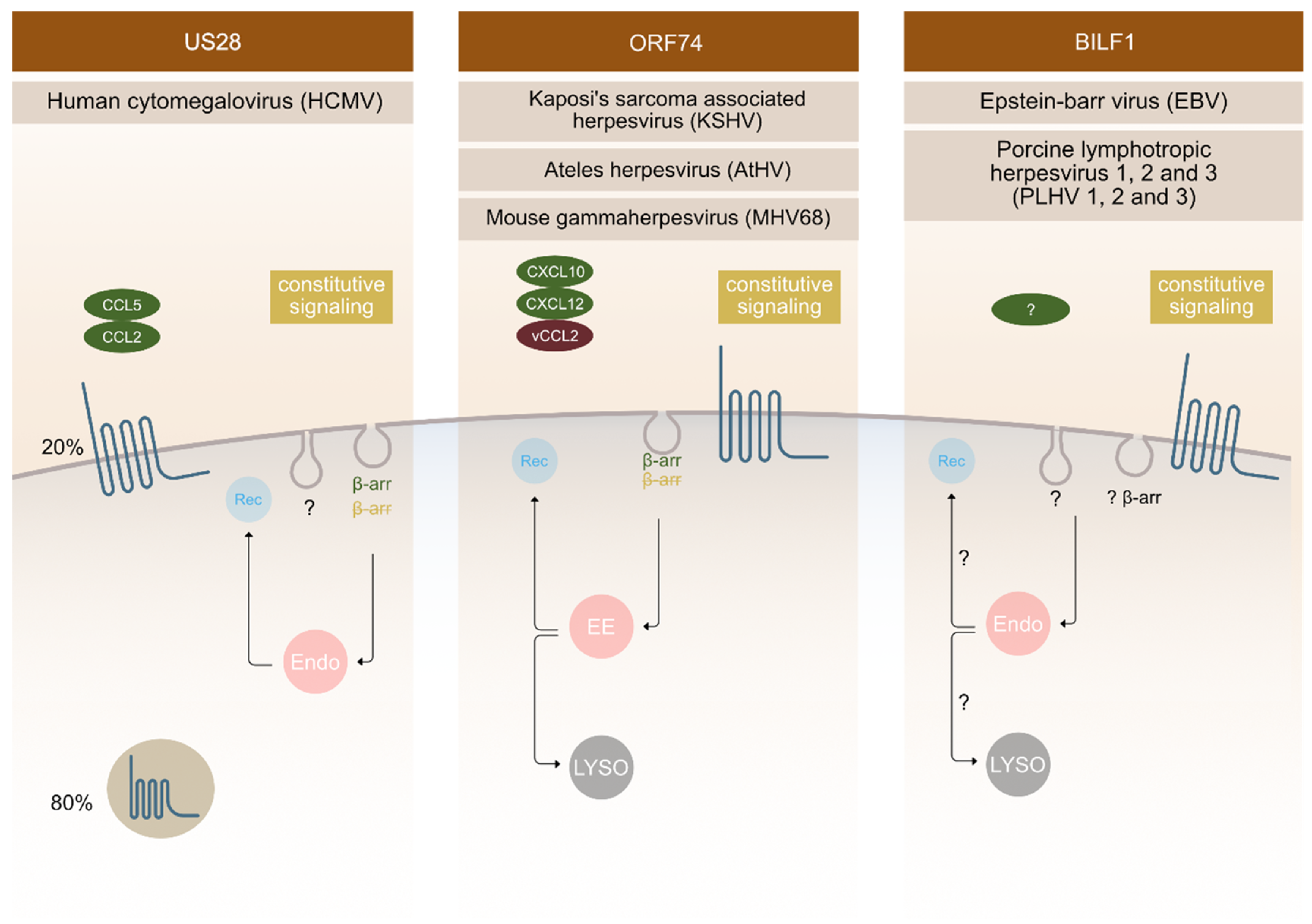
3. Endocytic Properties of The Most Commonly Studied vGPCRs
3.1. Cytomegalovirus (CMV)
3.2. Kaposi’s Sarcoma Associated Herpesvirus (KSHV)
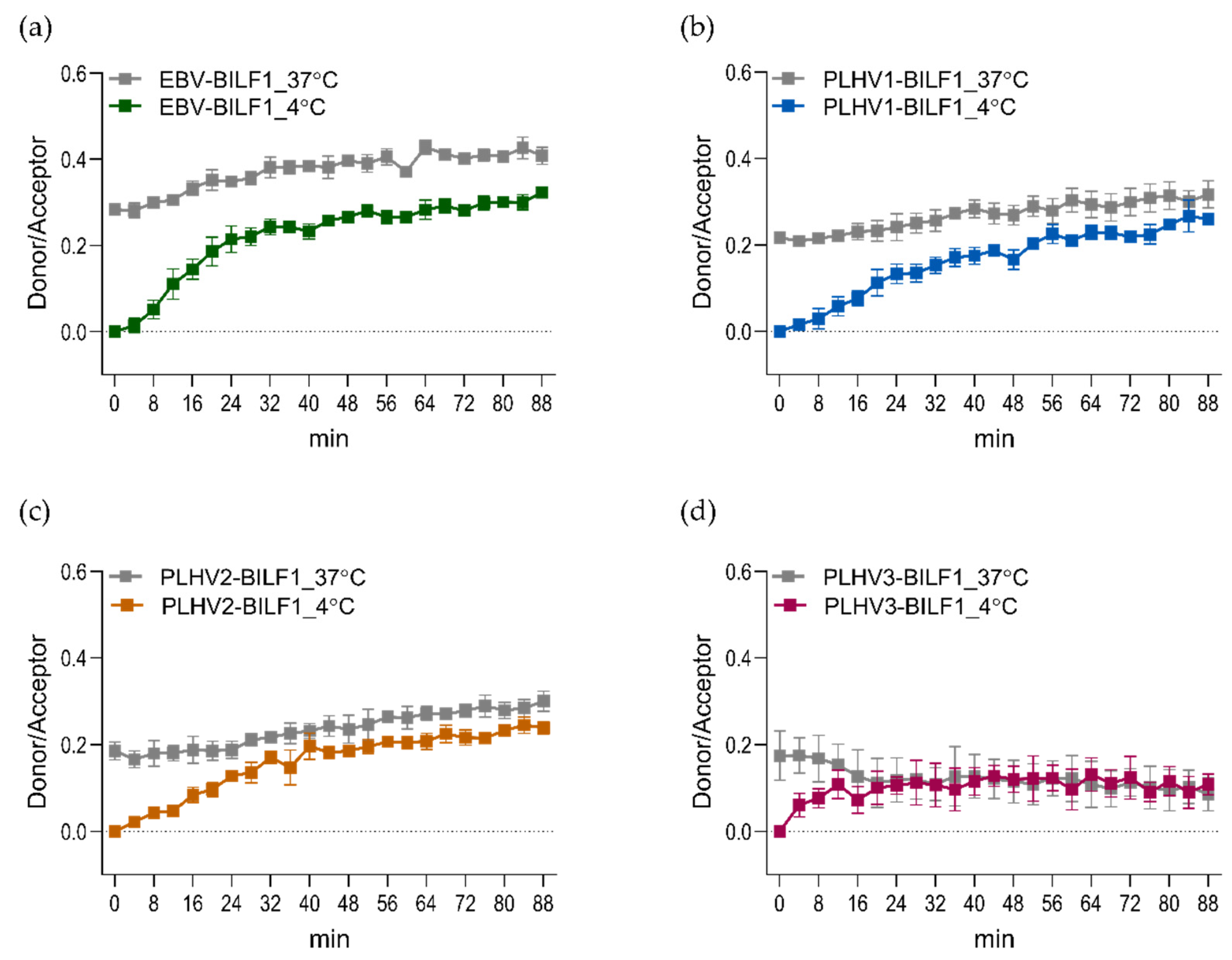
3.3. Epstein-Barr Virus (EBV) and Its Closely Related Lymphocryptoviruses
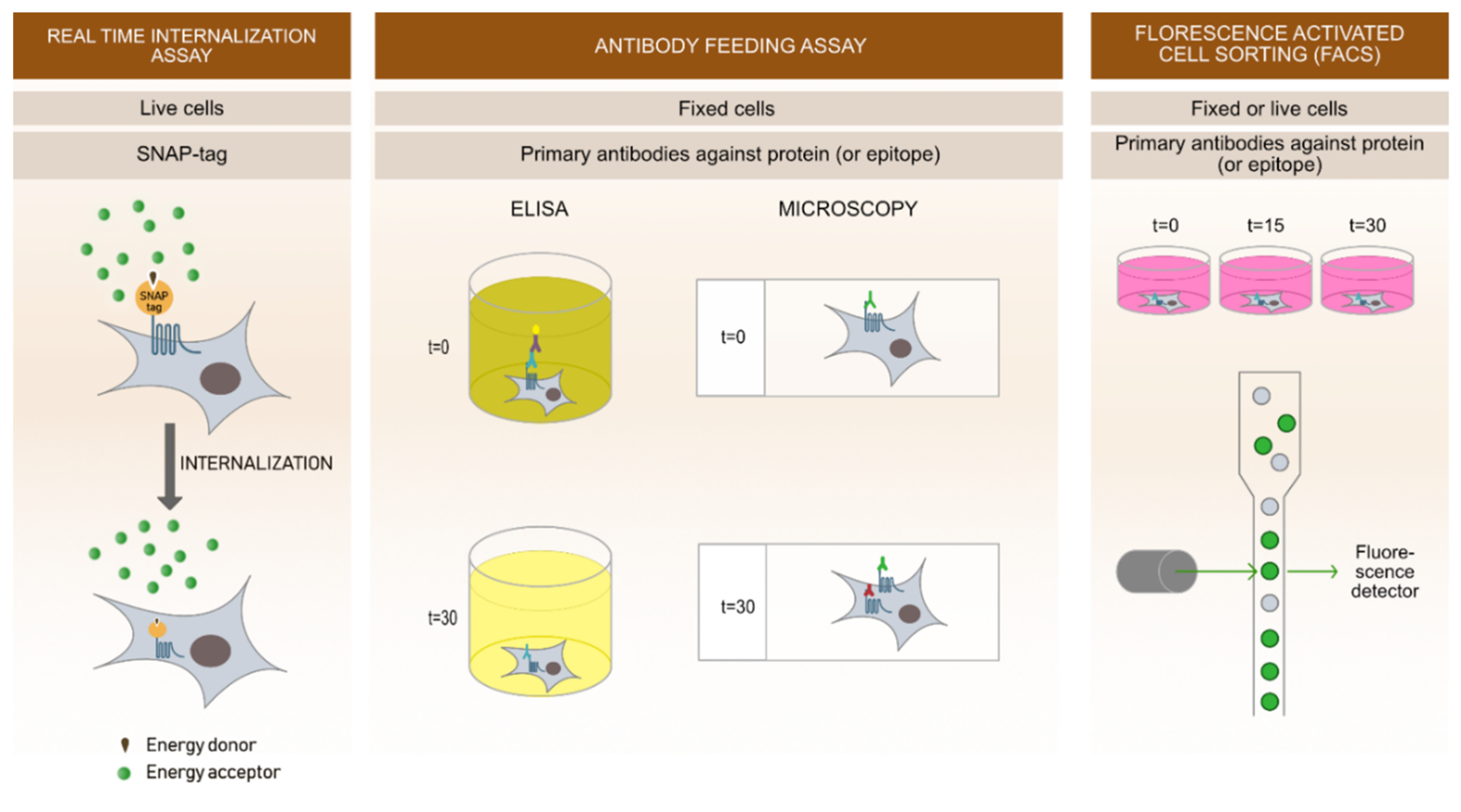
4. Methodological Approaches and Novel Techniques to Study Receptor Mediated Endocytosis
4.1. Real-Time Internalization Assay
4.2. Antibody Feeding Assay
4.3. Fluorescence-Activate Cell Sorting (FACS)
4.4. Microscopy Based Approaches
4.5. Manipulation of Endocytic Pathways
4.5.1. Chemical Inhibitors
4.5.2. Genetic Manipulation
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hanyaloglu, A.C.; von Zastrow, M. Regulation of GPCRs by endocytic membrane trafficking and its potential implications. Annu. Rev. Pharm. Toxicol. 2008, 48, 537–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joseph, J.G.; Liu, A.P. Mechanical Regulation of Endocytosis: New Insights and Recent Advances. Adv. Biosyst. 2020, 4, e1900278. [Google Scholar] [CrossRef] [PubMed]
- Sigismund, S.; Confalonieri, S.; Ciliberto, A.; Polo, S.; Scita, G.; Di Fiore, P.P. Endocytosis and signaling: Cell logistics shape the eukaryotic cell plan. Physiol. Rev. 2012, 92, 273–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbieri, E.; Di Fiore, P.P.; Sigismund, S. Endocytic control of signaling at the plasma membrane. Curr. Opin. Cell Biol. 2016, 39, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Lobingier, B.T.; von Zastrow, M. When trafficking and signaling mix: How subcellular location shapes G protein-coupled receptor activation of heterotrimeric G proteins. Traffic 2019, 20, 130–136. [Google Scholar] [CrossRef] [Green Version]
- Kumari, S.; Mg, S.; Mayor, S. Endocytosis unplugged: Multiple ways to enter the cell. Cell Res. 2010, 20, 256. [Google Scholar] [CrossRef] [Green Version]
- Hanyaloglu, A.C. Advances in Membrane Trafficking and Endosomal Signaling of G Protein-Coupled Receptors. Int. Rev. Cell Mol. Biol. 2018, 339, 93–131. [Google Scholar] [CrossRef]
- Sobhy, H. A comparative review of viral entry and attachment during large and giant dsDNA virus infections. Arch. Virol. 2017, 162, 3567–3585. [Google Scholar] [CrossRef]
- Traylen, C.M.; Patel, H.R.; Fondaw, W.; Mahatme, S.; Williams, J.F.; Walker, L.R.; Dyson, O.F.; Arce, S.; Akula, S.M. Virus reactivation: A panoramic view in human infections. Future Virol. 2011, 6, 451–463. [Google Scholar] [CrossRef] [Green Version]
- Alcami, A.; Lira, S.A. Modulation of chemokine activity by viruses. Curr. Opin. Immunol. 2010, 22, 482–487. [Google Scholar] [CrossRef]
- Raftery, M.; Muller, A.; Schonrich, G. Herpesvirus homologues of cellular genes. Virus Genes 2000, 21, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Rosenkilde, M.M.; Smit, M.J.; Waldhoer, M. Structure, function and physiological consequences of virally encoded chemokine seven transmembrane receptors. Br. J. Pharm. 2008, 153 (Suppl. 1), S154–S166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Senten, J.R.; Fan, T.S.; Siderius, M.; Smit, M.J. Viral G Protein-Coupled Receptors as Modulators of Cancer Hallmarks. Pharmacol. Res. 2020. [Google Scholar] [CrossRef] [PubMed]
- Vischer, H.F.; Siderius, M.; Leurs, R.; Smit, M.J. Herpesvirus-encoded GPCRs: Neglected players in inflammatory and proliferative diseases? Nat. Rev. Drug Discov. 2014, 13, 123–139. [Google Scholar] [CrossRef]
- Zuo, J.; Currin, A.; Griffin, B.D.; Shannon-Lowe, C.; Thomas, W.A.; Ressing, M.E.; Wiertz, E.J.; Rowe, M. The Epstein-Barr virus G-protein-coupled receptor contributes to immune evasion by targeting MHC class I molecules for degradation. PLoS Pathog. 2009, 5, e1000255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenkilde, M.M. Virus-encoded chemokine receptors—Putative novel antiviral drug targets. Neuropharmacology 2005, 48, 1–13. [Google Scholar] [CrossRef]
- Weeratunga, S.; Paul, B.; Collins, B.M. Recognising the signals for endosomal trafficking. Curr. Opin. Cell Biol. 2020, 65, 17–27. [Google Scholar] [CrossRef]
- McNally, K.E.; Cullen, P.J. Endosomal Retrieval of Cargo: Retromer Is Not Alone. Trends. Cell Biol. 2018, 28, 807–822. [Google Scholar] [CrossRef] [Green Version]
- Huotari, J.; Helenius, A. Endosome maturation. EMBO J. 2011, 30, 3481–3500. [Google Scholar] [CrossRef]
- Wolfe, B.L.; Trejo, J. Clathrin-dependent mechanisms of G protein-coupled receptor endocytosis. Traffic 2007, 8, 462–470. [Google Scholar] [CrossRef]
- Smith, S.M.; Michael, B.; Mary, H.; Corinne, J.M. Weak Molecular Interactions in Clathrin-Mediated Endocytosis. Front. Mol. Biol. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takei, K.; Haucke, V. Clathrin-mediated endocytosis: Membrane factors pull the trigger. Trends Cell Biol. 2001, 11, 385–391. [Google Scholar] [CrossRef]
- Kaksonen, M.; Roux, A. Mechanisms of clathrin-mediated endocytosis. Nat. Rev. Mol. Cell Biol. 2018, 19, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Mettlen, M.; Chen, P.H.; Srinivasan, S.; Danuser, G.; Schmid, S.L. Regulation of Clathrin-Mediated Endocytosis. Annu. Rev. Biochem. 2018, 87, 871–896. [Google Scholar] [CrossRef] [PubMed]
- Fraile-Ramos, A.; Kohout, T.A.; Waldhoer, M.; Marsh, M. Endocytosis of the viral chemokine receptor US28 does not require beta-arrestins but is dependent on the clathrin-mediated pathway. Traffic 2003, 4, 243–253. [Google Scholar] [CrossRef] [PubMed]
- De Munnik, S.M.; Kooistra, A.J.; van Offenbeek, J.; Nijmeijer, S.; de Graaf, C.; Smit, M.J.; Leurs, R.; Vischer, H.F. The Viral G Protein-Coupled Receptor ORF74 Hijacks β-Arrestins for Endocytic Trafficking in Response to Human Chemokines. PLoS ONE 2015, 10, e0124486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ungewickell, E.J.; Hinrichsen, L. Endocytosis: Clathrin-mediated membrane budding. Curr. Opin Cell Biol. 2007, 19, 417–425. [Google Scholar] [CrossRef]
- Kumar, M.; Gouw, M.; Michael, S.; Sámano-Sánchez, H.; Pancsa, R.; Glavina, J.; Diakogianni, A.; Valverde, J.A.; Bukirova, D.; Čalyševa, J.; et al. ELM-the eukaryotic linear motif resource in 2020. Nucleic Acids Res. 2020, 48, D296–D306. [Google Scholar] [CrossRef] [Green Version]
- Azzi, S.; Smith, S.S.; Dwyer, J.; Leclair, H.M.; Alexia, C.; Hebda, J.K.; Dupin, N.; Bidère, N.; Gavard, J. YGLF motif in the Kaposi sarcoma herpes virus G-protein-coupled receptor adjusts NF-κB activation and paracrine actions. Oncogene 2014, 33, 5609–5618. [Google Scholar] [CrossRef] [Green Version]
- Parton, R.G.; Simons, K. The multiple faces of caveolae. Nat. Rev. Mol. Cell Biol. 2007, 8, 185–194. [Google Scholar] [CrossRef]
- Ferreira, A.P.A.; Boucrot, E. Mechanisms of Carrier Formation during Clathrin-Independent Endocytosis. Trends Cell Biol. 2018, 28, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Sezgin, E.; Levental, I.; Mayor, S.; Eggeling, C. The mystery of membrane organization: Composition, regulation and roles of lipid rafts. Nat. Rev. Mol. Cell Biol. 2017, 18, 361–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck-García, K.; Beck-García, E.; Bohler, S.; Zorzin, C.; Sezgin, E.; Levental, I.; Alarcón, B.; Schamel, W.W. Nanoclusters of the resting T cell antigen receptor (TCR) localize to non-raft domains. Biochim. Biophys. Acta. 2015, 1853, 802–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sproul, T.W.; Malapati, S.; Kim, J.; Pierce, S.K. Cutting edge: B cell antigen receptor signaling occurs outside lipid rafts in immature B cells. J. Immunol. 2000, 165, 6020–6023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razani, B.; Rubin, C.S.; Lisanti, M.P. Regulation of cAMP-mediated signal transduction via interaction of caveolins with the catalytic subunit of protein kinase A. J. Biol. Chem. 1999, 274, 26353–26360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teissier, E.; Pécheur, E.I. Lipids as modulators of membrane fusion mediated by viral fusion proteins. Eur. Biophys. J. 2007, 36, 887–899. [Google Scholar] [CrossRef]
- Ewers, H.; Helenius, A. Lipid-mediated endocytosis. Cold Spring Harb. Perspect. Biol. 2011, 3, a004721. [Google Scholar] [CrossRef]
- Fecchi, K.; Anticoli, S.; Peruzzu, D.; Iessi, E.; Gagliardi, M.C.; Matarrese, P.; Ruggieri, A. Coronavirus Interplay with Lipid Rafts and Autophagy Unveils Promising Therapeutic Targets. Front. Microbiol. 2020, 11, 1821. [Google Scholar] [CrossRef]
- Dick, R.A.; Goh, S.L.; Feigenson, G.W.; Vogt, V.M. HIV-1 Gag protein can sense the cholesterol and acyl chain environment in model membranes. Proc. Natl. Acad. Sci USA 2012, 109, 18761–18766. [Google Scholar] [CrossRef] [Green Version]
- Nabi, I.R.; Le, P.U. Caveolae/raft-dependent endocytosis. J. Cell Biol. 2003, 161, 673–677. [Google Scholar] [CrossRef]
- Droese, J.; Mokros, T.; Hermosilla, R.; Schülein, R.; Lipp, M.; Höpken, U.E.; Rehm, A. HCMV-encoded chemokine receptor US28 employs multiple routes for internalization. Biochem. Biophys. Res. Commun. 2004, 322, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Limaye, A.P.; Babu, T.M.; Boeckh, M. Progress and Challenges in the Prevention, Diagnosis, and Management of Cytomegalovirus Infection in Transplantation. Clin. Microbiol. Rev. 2020, 34. [Google Scholar] [CrossRef] [PubMed]
- Kledal, T.N.; Rosenkilde, M.M.; Schwartz, T.W. Selective recognition of the membrane-bound CX3C chemokine, fractalkine, by the human cytomegalovirus-encoded broad-spectrum receptor US28. FEBS Lett. 1998, 441, 209–214. [Google Scholar] [CrossRef] [Green Version]
- Neote, K.; DiGregorio, D.; Mak, J.Y.; Horuk, R.; Schall, T.J. Molecular cloning, functional expression, and signaling characteristics of a C-C chemokine receptor. Cell 1993, 72, 415–425. [Google Scholar] [CrossRef]
- McLean, K.A.; Holst, P.J.; Martini, L.; Schwartz, T.W.; Rosenkilde, M.M. Similar activation of signal transduction pathways by the herpesvirus-encoded chemokine receptors US28 and ORF74. Virology 2004, 325, 241–251. [Google Scholar] [CrossRef] [Green Version]
- Krishna, B.A.; Spiess, K.; Poole, E.L.; Lau, B.; Voigt, S.; Kledal, T.N.; Rosenkilde, M.M.; Sinclair, J.H. Targeting the latent cytomegalovirus reservoir with an antiviral fusion toxin protein. Nat. Commun. 2017, 8, 14321. [Google Scholar] [CrossRef] [Green Version]
- Spiess, K.; Jeppesen, M.G.; Malmgaard-Clausen, M.; Krzywkowski, K.; Dulal, K.; Cheng, T.; Hjorto, G.M.; Larsen, O.; Burg, J.S.; Jarvis, M.A.; et al. Rationally designed chemokine-based toxin targeting the viral G protein-coupled receptor US28 potently inhibits cytomegalovirus infection in vivo. Proc. Natl. Acad. Sci. USA 2015, 112, 8427–8432. [Google Scholar] [CrossRef] [Green Version]
- Spiess, K.; Jeppesen, M.G.; Malmgaard-Clausen, M.; Krzywkowski, K.; Kledal, T.N.; Rosenkilde, M.M. Novel Chemokine-Based Immunotoxins for Potent and Selective Targeting of Cytomegalovirus Infected Cells. J. Immunol. Res. 2017, 2017, 4069260. [Google Scholar] [CrossRef]
- Fraile-Ramos, A.; Pelchen-Matthews, A.; Kledal, T.N.; Browne, H.; Schwartz, T.W.; Marsh, M. Localization of HCMV UL33 and US27 in endocytic compartments and viral membranes. Traffic 2002, 3, 218–232. [Google Scholar] [CrossRef]
- Wagner, S.; Arnold, F.; Wu, Z.; Schubert, A.; Walliser, C.; Tadagaki, K.; Jockers, R.; Mertens, T.; Michel, D. The 7-transmembrane protein homologue UL78 of the human cytomegalovirus forms oligomers and traffics between the plasma membrane and different intracellular compartments. Arch. Virol. 2012, 157, 935–949. [Google Scholar] [CrossRef]
- Ahuja, S.K.; Murphy, P.M. Molecular piracy of mammalian interleukin-8 receptor type B by herpesvirus saimiri. J. Biol. Chem. 1993, 268, 20691–20694. [Google Scholar] [PubMed]
- Rosenkilde, M.M.; Kledal, T.N.; Brauner-Osborne, H.; Schwartz, T.W. Agonists and inverse agonists for the herpesvirus 8-encoded constitutively active seven-transmembrane oncogene product, ORF-74. J. Biol. Chem. 1999, 274, 956–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenkilde, M.M.; Kledal, T.N.; Holst, P.J.; Schwartz, T.W. Selective elimination of high constitutive activity or chemokine binding in the human herpesvirus 8 encoded seven transmembrane oncogene ORF74. J. Biol. Chem. 2000, 275, 26309–26315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenkilde, M.M.; Schwartz, T.W. Potency of ligands correlates with affinity measured against agonist and inverse agonists but not against neutral ligand in constitutively active chemokine receptor. Mol. Pharm. 2000, 57, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Rosenkilde, M.M.; McLean, K.A.; Holst, P.J.; Schwartz, T.W. The CXC chemokine receptor encoded by herpesvirus saimiri, ECRF3, shows ligand-regulated signaling through Gi, Gq, and G12/13 proteins but constitutive signaling only through Gi and G12/13 proteins. J. Biol. Chem. 2004, 279, 32524–32533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.Y.; Chen, S.C.; Leach, M.W.; Manfra, D.; Homey, B.; Wiekowski, M.; Sullivan, L.; Jenh, C.H.; Narula, S.K.; Chensue, S.W.; et al. Transgenic expression of the chemokine receptor encoded by human herpesvirus 8 induces an angioproliferative disease resembling Kaposi’s sarcoma. J. Exp. Med. 2000, 191, 445–454. [Google Scholar] [CrossRef]
- Holst, P.J.; Rosenkilde, M.M.; Manfra, D.; Chen, S.C.; Wiekowski, M.T.; Holst, B.; Cifire, F.; Lipp, M.; Schwartz, T.W.; Lira, S.A. Tumorigenesis induced by the HHV8-encoded chemokine receptor requires ligand modulation of high constitutive activity. J. Clin. Investig. 2001, 108, 1789–1796. [Google Scholar] [CrossRef]
- Grisotto, M.G.; Garin, A.; Martin, A.P.; Jensen, K.K.; Chan, P.; Sealfon, S.C.; Lira, S.A. The Human Herpesvirus 8 Chemokine Receptor vGPCR Triggers Autonomous Proliferation of Endothelial Cells. J. Clin. Investig. 2006, 116. [Google Scholar] [CrossRef]
- De Munnik, S.M.; Smit, M.J.; Leurs, R.; Vischer, H.F. Modulation of cellular signaling by herpesvirus-encoded G protein-coupled receptors. Front. Pharm. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Paulsen, S.J.; Rosenkilde, M.M.; Eugen-Olsen, J.; Kledal, T.N. Epstein-Barr virus-encoded BILF1 is a constitutively active G protein-coupled receptor. J. Virol. 2005, 79, 536–546. [Google Scholar] [CrossRef] [Green Version]
- Chmielewicz, B.; Goltz, M.; Franz, T.; Bauer, C.; Brema, S.; Ellerbrok, H.; Beckmann, S.; Rziha, H.-J.; Lahrmann, K.-H.; Romero, C.; et al. A novel porcine gammaherpesvirus. Virology 2003, 308, 317–329. [Google Scholar] [CrossRef] [Green Version]
- Lindner, I.; Ehlers, B.; Noack, S.; Dural, G.; Yasmum, N.; Bauer, C.; Goltz, M. The porcine lymphotropic herpesvirus 1 encodes functional regulators of gene expression. Virology 2007, 357, 134–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiess, K.; Fares, S.; Sparre-Ulrich, A.H.; Hilgenberg, E.; Jarvis, M.A.; Ehlers, B.; Rosenkilde, M.M. Identification and functional comparison of seven-transmembrane G-protein-coupled BILF1 receptors in recently discovered nonhuman primate lymphocryptoviruses. J. Virol. 2015, 89, 2253–2267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goltz, M.; Ericsson, T.; Patience, C.; Huang, C.A.; Noack, S.; Sachs, D.H.; Ehlers, B. Sequence analysis of the genome of porcine lymphotropic herpesvirus 1 and gene expression during posttransplant lymphoproliferative disease of pigs. Virology 2002, 294, 383–393. [Google Scholar] [CrossRef] [Green Version]
- Hart, J.; Ackermann, M.; Jayawardane, G.; Russell, G.; Haig, D.M.; Reid, H.; Stewart, J.P. Complete sequence and analysis of the ovine herpesvirus 2 genome. J. Gen. Virol. 2007, 88, 28–39. [Google Scholar] [CrossRef] [Green Version]
- Boudry, C.; Markine-Goriaynoff, N.; Delforge, C.; Springael, J.Y.; de Leval, L.; Drion, P.; Russell, G.; Haig, D.M.; Vanderplasschen, A.F.; Dewals, B. The A5 gene of alcelaphine herpesvirus 1 encodes a constitutively active G-protein-coupled receptor that is non-essential for the induction of malignant catarrhal fever in rabbits. J. Gen. Virol. 2007, 88, 3224–3233. [Google Scholar] [CrossRef]
- Griffin, B.D.; Gram, A.M.; Mulder, A.; Van Leeuwen, D.; Claas, F.H.; Wang, F.; Ressing, M.E.; Wiertz, E. EBV BILF1 evolved to downregulate cell surface display of a wide range of HLA class I molecules through their cytoplasmic tail. J. Immunol. 2013, 190, 1672–1684. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Kim, K.M. Multifactorial Regulation of G Protein-Coupled Receptor Endocytosis. Biomol. Ther. 2017, 25, 26–43. [Google Scholar] [CrossRef] [Green Version]
- Foster, S.R.; Bräuner-Osborne, H. Investigating Internalization and Intracellular Trafficking of GPCRs: New Techniques and Real-Time Experimental Approaches. Handb. Exp. Pharm. 2018, 245, 41–61. [Google Scholar] [CrossRef]
- Arancibia-Cárcamo, I.L.; Fairfax, B.P.; Moss, S.J.; Kittler, J.T. Studying the Localization, Surface Stability and Endocytosis of Neurotransmitter Receptors by Antibody Labeling and Biotinylation Approaches. Available online: https://www.ncbi.nlm.nih.gov/books/NBK2552/ (accessed on 26 November 2020).
- Fraile-Ramos, A.; Kledal, T.N.; Pelchen-Matthews, A.; Bowers, K.; Schwartz, T.W.; Marsh, M. The human cytomegalovirus US28 protein is located in endocytic vesicles and undergoes constitutive endocytosis and recycling. Mol. Biol. Cell 2001, 12, 1737–1749. [Google Scholar] [CrossRef] [Green Version]
- Hislop, J.N.; von Zastrow, M. Analysis of GPCR localization and trafficking. Methods Mol. Biol. 2011, 746, 425–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cossarizza, A.; Chang, H.D.; Radbruch, A.; Acs, A.; Adam, D.; Adam-Klages, S.; Agace, W.W.; Aghaeepour, N.; Akdis, M.; Allez, M.; et al. Guidelines for the use of flow cytometry and cell sorting in immunological studies (second edition). Eur. J. Immunol. 2019, 49, 1457–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado-Peraza, F.; Nogueras-Ortiz, C.; Acevedo Canabal, A.M.; Roman-Vendrell, C.; Yudowski, G.A. Imaging GPCRs trafficking and signaling with total internal reflection fluorescence microscopy in cultured neurons. Methods Cell Biol. 2016, 132, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Shashkova, S.; Leake, M. Single-molecule fluorescence microscopy review: Shedding new light on old problems. Biosci. Rep. 2017, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reck-Peterson, S.L.; Derr, N.D.; Stuurman, N. Imaging single molecules using total internal reflection fluorescence microscopy (TIRFM). Cold Spring Harb. Protoc. 2010, 2010, pdb.top73. [Google Scholar] [CrossRef] [PubMed]
- Wandinger-Ness, A.; Zerial, M. Rab proteins and the compartmentalization of the endosomal system. Cold Spring Harb. Perspect. Biol. 2014, 6, a022616. [Google Scholar] [CrossRef]
- Dutta, D.; Donaldson, J.G. Search for inhibitors of endocytosis: Intended specificity and unintended consequences. Cell Logist. 2012, 2, 203–208. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.H.; Rothberg, K.G.; Anderson, R.G. Mis-assembly of clathrin lattices on endosomes reveals a regulatory switch for coated pit formation. J. Cell Biol. 1993, 123, 1107–1117. [Google Scholar] [CrossRef]
- Macia, E.; Ehrlich, M.; Massol, R.; Boucrot, E.; Brunner, C.; Kirchhausen, T. Dynasore, a cell-permeable inhibitor of dynamin. Dev. Cell 2006, 10, 839–850. [Google Scholar] [CrossRef] [Green Version]
- Hall, B.; Limaye, A.; Kulkarni, A.B. Overview: Generation of gene knockout mice. Curr. Protoc. Cell Biol. 2009, 44. [Google Scholar] [CrossRef] [Green Version]
- Kohout, T.A.; Lin, F.T.; Perry, S.J.; Conner, D.A.; Lefkowitz, R.J. β-Arrestin 1 and 2 differentially regulate heptahelical receptor signaling and trafficking. Proc. Natl. Acad. Sci. USA 2001, 98, 1601–1606. [Google Scholar] [CrossRef] [Green Version]
- Williams, B.O.; Warman, M.L. CRISPR/CAS9 Technologies. J. Bone Min. Res. 2017, 32, 883–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reis, C.R.; Chen, P.H.; Srinivasan, S.; Aguet, F.; Mettlen, M.; Schmid, S.L. Crosstalk between Akt/GSK3β signaling and dynamin-1 regulates clathrin-mediated endocytosis. EMBO J. 2015, 34, 2132–2146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Bliek, A.M.; Redelmeier, T.E.; Damke, H.; Tisdale, E.J.; Meyerowitz, E.M.; Schmid, S.L. Mutations in human dynamin block an intermediate stage in coated vesicle formation. J. Cell Biol. 1993, 122, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Krupnick, J.G.; Santini, F.; Gagnon, A.W.; Keen, J.H.; Benovic, J.L. Modulation of the arrestin-clathrin interaction in cells. Characterization of beta-arrestin dominant-negative mutants. J. Biol. Chem. 1997, 272, 32507–32512. [Google Scholar] [CrossRef] [Green Version]
- Shigematsu, S.; Watson, R.T.; Khan, A.H.; Pessin, J.E. The adipocyte plasma membrane caveolin functional/structural organization is necessary for the efficient endocytosis of GLUT4. J. Biol. Chem. 2003, 278, 10683–10690. [Google Scholar] [CrossRef] [Green Version]
- Benmerah, A.; Bayrou, M.; Cerf-Bensussan, N.; Dautry-Varsat, A. Inhibition of clathrin-coated pit assembly by an Eps15 mutant. J. Cell Sci. 1999, 112, 1303–1311. [Google Scholar]
- Chen, C.; Zhuang, X. Epsin 1 is a cargo-specific adaptor for the clathrin-mediated endocytosis of the influenza virus. Proc. Natl. Acad. Sci. USA 2008, 105, 11790–11795. [Google Scholar] [CrossRef] [Green Version]
- Kubale, V.; Abramović, Z.; Pogacnik, A.; Heding, A.; Sentjurc, M.; Vrecl, M. Evidence for a role of caveolin-1 in neurokinin-1 receptor plasma-membrane localization, efficient signaling, and interaction with beta-arrestin 2. Cell Tissue Res. 2007, 330, 231–245. [Google Scholar] [CrossRef]
- Cunningham, C.M.; Bellipanni, G.; Habas, R.; Balciunas, D. Deletion of morpholino binding sites (DeMOBS) to assess specificity of morphant phenotypes. Sci. Rep. 2020, 10, 15366. [Google Scholar] [CrossRef]
- Eisen, J.S.; Smith, J.C. Controlling morpholino experiments: Don’t stop making antisense. Development 2008, 135, 1735–1743. [Google Scholar] [CrossRef] [Green Version]
- Leung, T.; Humbert, J.E.; Stauffer, A.M.; Giger, K.E.; Chen, H.; Tsai, H.J.; Wang, C.; Mirshahi, T.; Robishaw, J.D. The orphan G protein-coupled receptor 161 is required for left-right patterning. Dev. Biol. 2008, 323, 31–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryja, V.; Gradl, D.; Schambony, A.; Arenas, E.; Schulte, G. Beta-arrestin is a necessary component of Wnt/beta-catenin signaling in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2007, 104, 6690–6695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, P.S.; Boshoff, C.; Weiss, R.A.; Chang, Y. Molecular mimicry of human cytokine and cytokine response pathway genes by KSHV. Science 1996, 274, 1739–1744. [Google Scholar] [CrossRef]
- Maussang, D.; Verzijl, D.; van Walsum, M.; Leurs, R.; Holl, J.; Pleskoff, O.; Michel, D.; van Dongen, G.A.; Smit, M.J. Human cytomegalovirus-encoded chemokine receptor US28 promotes tumorigenesis. Proc. Natl. Acad. Sci. USA 2006, 103, 13068–13073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, W.E.; Zagorski, W.A.; Brenneman, J.D.; Avery, D.; Miller, J.L.; O’Connor, C.M. US28 is a potent activator of phospholipase C during HCMV infection of clinically relevant target cells. PLoS ONE 2012, 7, e50524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyngaa, R.; Norregaard, K.; Kristensen, M.; Kubale, V.; Rosenkilde, M.M.; Kledal, T.N. Cell transformation mediated by the Epstein-Barr virus G protein-coupled receptor BILF1 is dependent on constitutive signaling. Oncogene 2010, 29, 4388–4398. [Google Scholar] [CrossRef] [Green Version]
- Miller, W.E.; Houtz, D.A.; Nelson, C.D.; Kolattukudy, P.E.; Lefkowitz, R.J. G-protein-coupled receptor (GPCR) kinase phosphorylation and beta-arrestin recruitment regulate the constitutive signaling activity of the human cytomegalovirus US28 GPCR. J. Biol. Chem. 2003, 278, 21663–21671. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Virus | Receptor | Preferred Endocytic Pathway | Signaling Pathways | G Protein Coupling |
|---|---|---|---|---|---|
| β-Herpesviruses | Human cytomegalovirus (CMV) | US27 | - | - | - |
| US28 | β-arrestin independent clathrin-mediated, partly through lipid rafts | Constitutive NFκB, NFAT, CREB, PLC, SRF, STAT3, TCF/LEF, Ligand induced PLC, MAPK | Gαq, Gαi/o, Gα12/13 | ||
| UL33 | - | Constitutive SRC, CREB | Gαq, Gαi, Gαs | ||
| UL78 | - | - | - | ||
| Human herpesvirus 6 | U12 | - | - | - | |
| U51 | - | - | - | ||
| Human herpesvirus 7 | U12 | - | - | - | |
| U51 | - | - | - | ||
| Mouse cytomegalovirus | M33 | - | Constitutive PLC, NFκB, CREB | Gαs | |
| M78 | - | - | - | ||
| Rat cytomegalovirus | R33 | - | Constitutive PLC, NFκB | Gαq, Gαi | |
| R78 | - | - | - | ||
| γ-Herpesviruses | Human herpesvirus 8 (HHV8 or Kaposi’s sarcoma virus (KSHV)) | ORF74 | β-arrestin independent clathrin mediated constitutive endocytosis, β-arrestin dependent clathrin-mediated ligand dependent endocytosis | Constitutive and ligand induced RAC1, PLC, PKC, AKT, JNK-SAPK, LYN-SRC, GSK3, JAK2-STAT3, HIF1α, PI3Kγ, calcineurin | Gαq, Gαi,Gα12/13 |
| Ateles herpesvirus (AtHV) | ORF74-AtHV | - | - | - | |
| MouseHV68 | ORF74-MHV68 | - | not constitutively active PLC, MAPK, Akt, NFκB | Gαi | |
| Equine HV2 (EHV2) | E1 | - | - | - | |
| E6 | - | - | - | ||
| ORF74-EHV2 | - | - | Gαi | ||
| Herpesvirus Saimiri (HVS) | ECRF3 | - | - | - | |
| Human Epstein barr virus (EBV) | BILF1 | - | Constitutive NFκB, NFAT, CREB | Gαi | |
| Rhesus lymphocryptovirus (RhLCV) | BILF1 | - | Constitutive NFκB | Gαi | |
| Callitrichine herpesvirus 3 (CalHv3) | BILF1 | - | Constitutive NFκB, NFAT | Gαi | |
| Pan troglodytes lymphocryptovirus 1 (PtroLCV1) | BILF1 | - | Constitutive NFκB | Gαi | |
| Gorilla gorilla lymphocryptovirus 1 (GgorLCV1) | BILF1 | - | - | - | |
| Gorilla gorilla lymphocryptovirus 2 (GgorLCV2) | BILF1 | - | - | - | |
| Pongo pygmaeus lymphocryptovirus 1 (PpygLCV1) | BILF1 | - | Constitutive NFκB | Gαi | |
| Pongo pygmaeus lymphocryptovirus 2 (PpygLCV2) | BILF1 | - | - | - | |
| Symphalangus syndactylus lymphocryptovirus 1 (SsynLCV1) | BILF1 | - | Constitutive NFκB, NFAT | Gαi | |
| Symphalangus syndactylus lymphocryptovirus 2 (SsynLCV2) | BILF1 | - | - | - | |
| Macaca fascicularis lymphocryptovirus 1 (MfasLCV1) | BILF1 | - | - | - | |
| Erythrocebus patas lymphocryptovirus 1 (EpatLCV1) | BILF1 | - | - | - | |
| Piliocolobus badius lymphocryptovirus 1 (PbadLCV1) | BILF1 | - | - | - | |
| Ateles paniscus lymphocryptovirus 1 (ApanLCV1) | BILF1 | - | - | - | |
| Pithecia pithecia lymphocryptovirus 1 (Ppit LCV1) | BILF1 | - | - | - | |
| Porcine lymphotropic herpesvirus 1, 2 and 3 (PLHV1-3) | BILF1 | - | - | - |
| Receptor | Elm Name | Instances | Positions | Elm Description | Cell Compartment |
|---|---|---|---|---|---|
| (Matched Sequence) | |||||
| EBV-BILF1 | TRG_ENDOCYTIC_2 | YSAF | 32–35 [A] | Tyrosine-based sorting signal responsible for the interaction with µ2 subunit of AP (Adaptor Protein) complex | plasma membrane, |
| clathrin-coated endocytic vesicle, | |||||
| cytosol | |||||
| PLHV1-BILF1 | TRG_ENDOCYTIC_2 | YTTL | 179–182 [A] | Tyrosine-based sorting signal responsible for the interaction with µ2 subunit of AP (Adaptor Protein) complex | plasma membrane, |
| clathrin-coated endocytic vesicle, | |||||
| cytosol | |||||
| PLHV2-BILF1 | TRG_ENDOCYTIC_2 | YAVL | 159–162 [A] | Tyrosine-based sorting signal responsible for the interaction with µ2 subunit of AP (Adaptor Protein) complex | plasma membrane, |
| clathrin-coated endocytic vesicle, | |||||
| cytosol | |||||
| PLHV3-BILF1 | TRG_ENDOCYTIC_2 | YAAL | 194–197 [A] | Tyrosine-based sorting signal responsible for the interaction with µ2 subunit of AP (Adaptor Protein) complex | plasma membrane, |
| clathrin-coated endocytic vesicle, | |||||
| cytosol | |||||
| US28 | TRG_ENDOCYTIC_2 | YYAI | 130–133 [A] | Tyrosine-based sorting signal responsible for the interaction with µ2 subunit of AP (Adaptor Protein) complex | plasma membrane, |
| YAIV | 131–134 [A] | clathrin-coated endocytic vesicle, | |||
| YRPV | 138–141 [A] | cytosol | |||
| YDYL | 177–180 [A] | ||||
| YLEV | 179–182 [A] | ||||
| YHSM | 321–324 [A] | ||||
| ORF74 | TRG_ENDOCYTIC_2 | YGLF | 326–329 [A] | Tyrosine-based sorting signal responsible for the interaction with µ2 subunit of AP (Adaptor Protein) complex | plasma membrane, |
| clathrin-coated endocytic vesicle, | |||||
| cytosol |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mavri, M.; Spiess, K.; Rosenkilde, M.M.; Rutland, C.S.; Vrecl, M.; Kubale, V. Methods for Studying Endocytotic Pathways of Herpesvirus Encoded G Protein-Coupled Receptors. Molecules 2020, 25, 5710. https://doi.org/10.3390/molecules25235710
Mavri M, Spiess K, Rosenkilde MM, Rutland CS, Vrecl M, Kubale V. Methods for Studying Endocytotic Pathways of Herpesvirus Encoded G Protein-Coupled Receptors. Molecules. 2020; 25(23):5710. https://doi.org/10.3390/molecules25235710
Chicago/Turabian StyleMavri, Maša, Katja Spiess, Mette Marie Rosenkilde, Catrin Sian Rutland, Milka Vrecl, and Valentina Kubale. 2020. "Methods for Studying Endocytotic Pathways of Herpesvirus Encoded G Protein-Coupled Receptors" Molecules 25, no. 23: 5710. https://doi.org/10.3390/molecules25235710
APA StyleMavri, M., Spiess, K., Rosenkilde, M. M., Rutland, C. S., Vrecl, M., & Kubale, V. (2020). Methods for Studying Endocytotic Pathways of Herpesvirus Encoded G Protein-Coupled Receptors. Molecules, 25(23), 5710. https://doi.org/10.3390/molecules25235710