Current Approaches to and Future Perspectives on Methomyl Degradation in Contaminated Soil/Water Environments

 ,
,  and
and 
Abstract
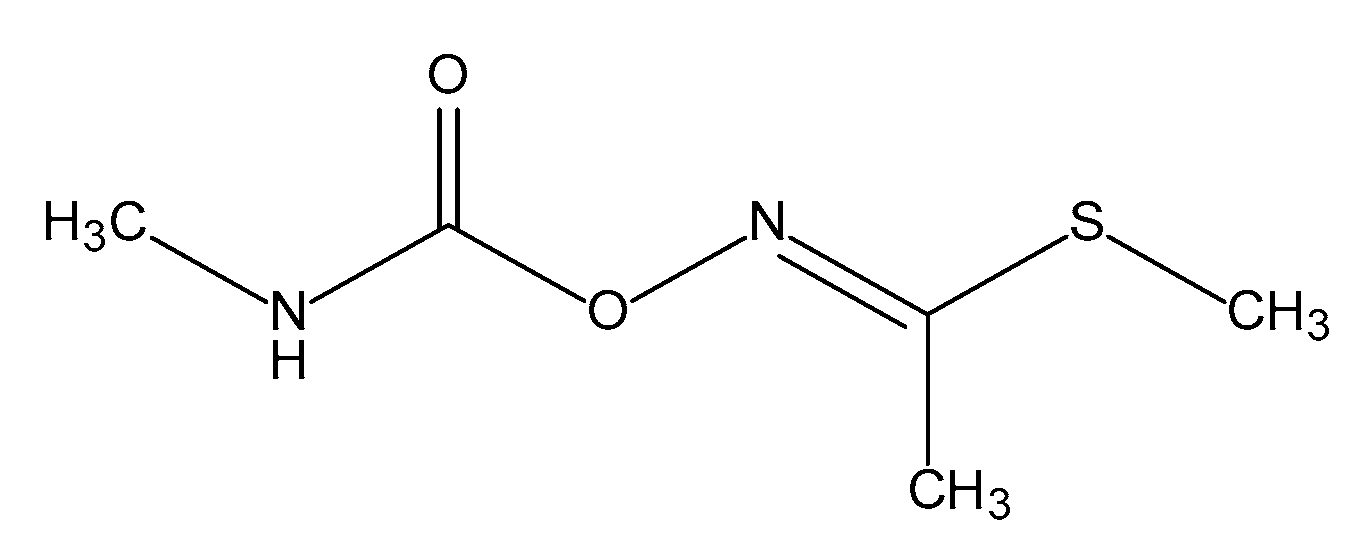
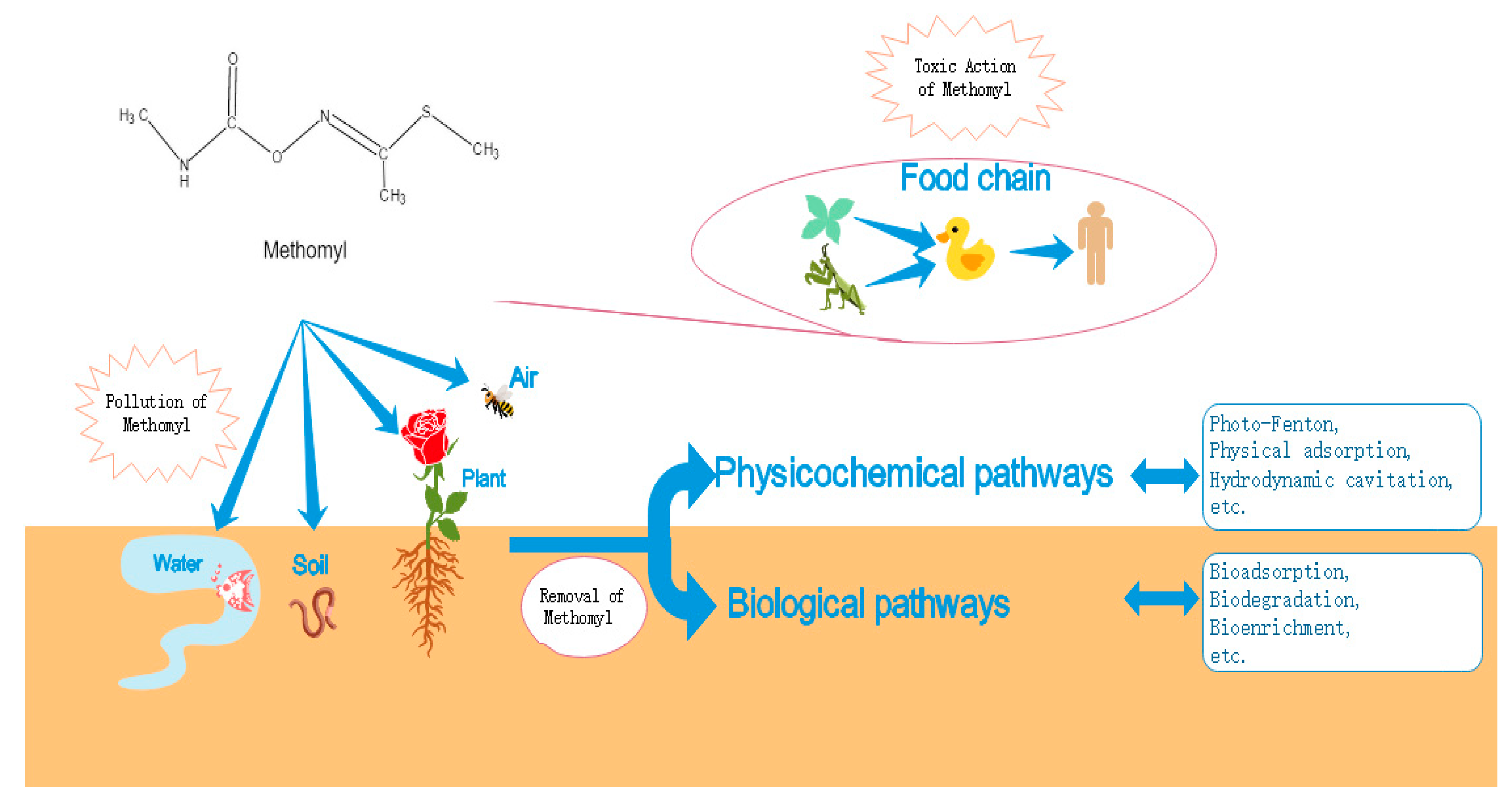
:1. Introduction
2. Toxicological Effects of Methomyl Insecticides
3. Physicochemical Methods for the Remediation of Methomyl-Affected Environments
4. Microbial Degradation of Methomyl
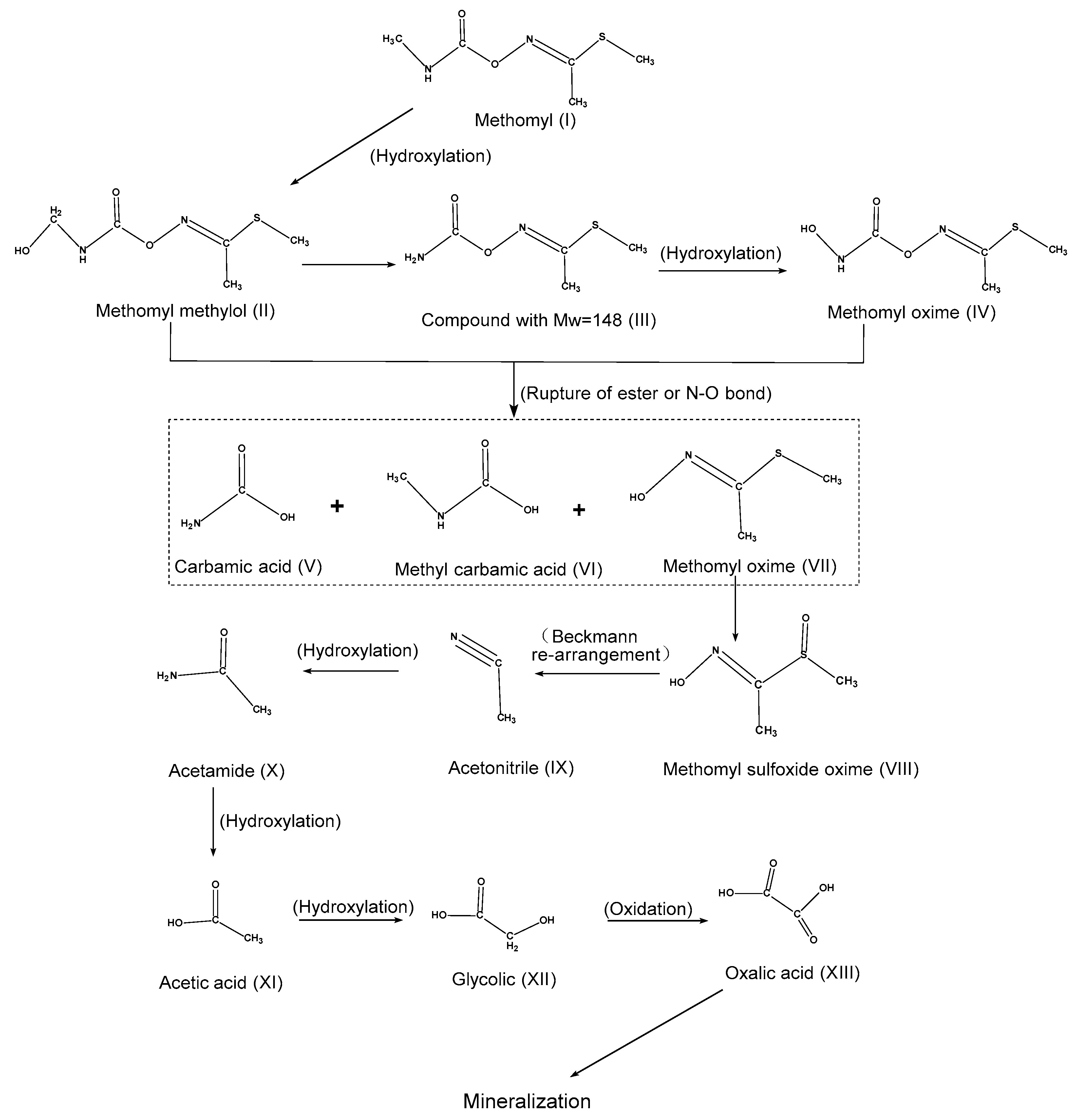
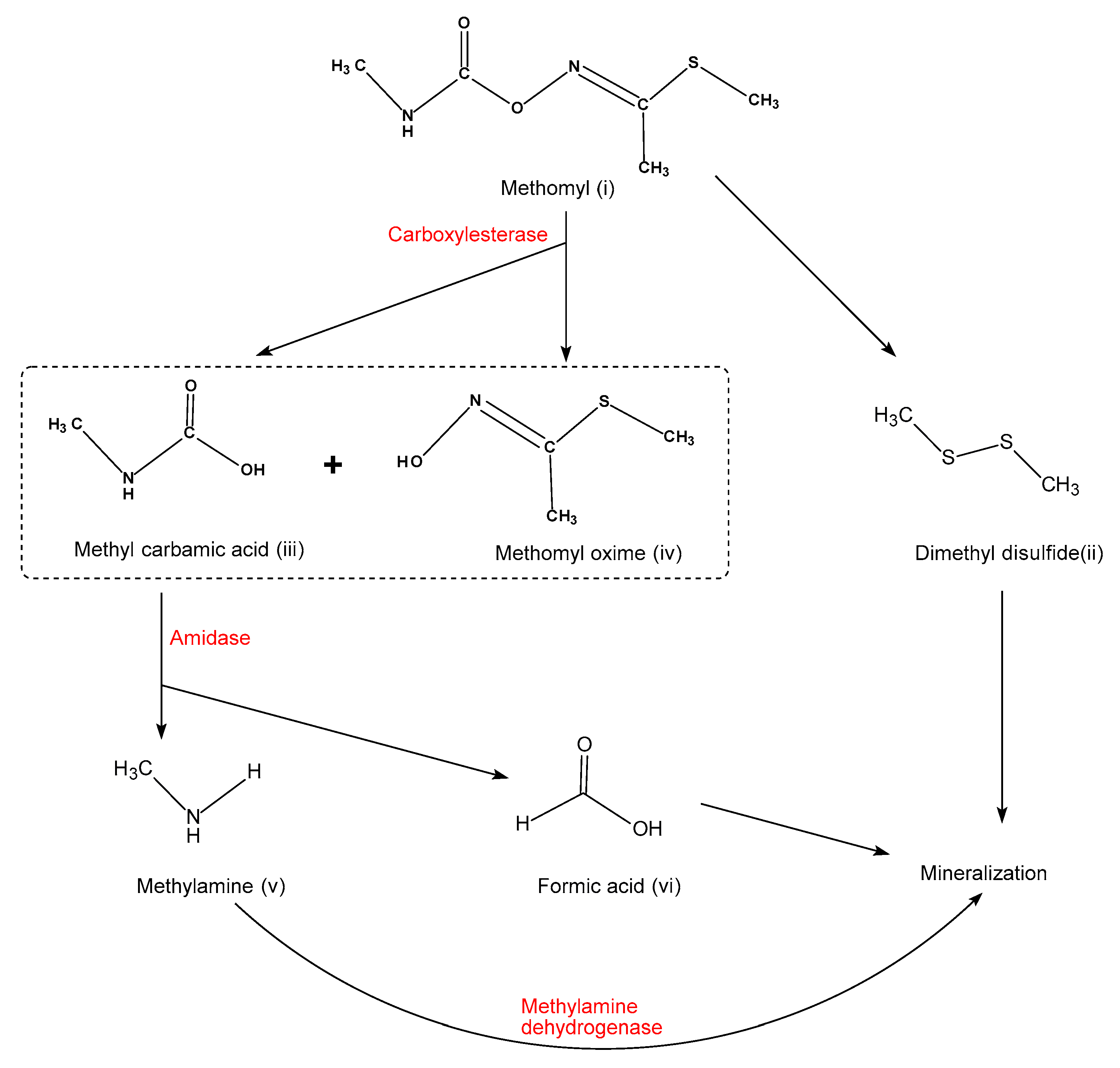
5. Molecular Mechanism of Methomyl Degradation
6. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Ribera, D.; Narbonne, J.F.; Arnaud, C.; Saint-Denis, M. Biochemical responses of the earthworm Eisenia fetida andrei exposed to contaminated articial soil, effects of carbaryl. Soil Biol. Biochem. 2001, 33, 1123–1130. [Google Scholar] [CrossRef]
- Kaur, M.; Sandhir, R. Comparative effects of acute and chronic carbofuran exposure on oxidative stress and drug-metabolizing enzymes in liver. Drug Chem. Toxicol. 2008, 29, 415–421. [Google Scholar] [CrossRef]
- Fernandez-Alba, A.R.; Hernando, D.; Aguera, A.; Caceres, J.; Malato, S. Toxicity assays: A way for evaluating AOPs efficiency. Water Res. 2002, 36, 4255–4262. [Google Scholar] [CrossRef]
- Filho, M.V.S.; Oliveira, M.M.; Salles, J.B.; Bastos, V.L.F.C.; Cassano, V.P.F.; Bastos, J.C. Methyl-paraoxon comparative inhibition kinetics for acetylcholinesterases from brain of neotropical fishes. Toxicol. Lett. 2004, 153, 247–254. [Google Scholar] [CrossRef]
- Yi, M.; Liu, H.; Shi, X.; Liang, P.; Gao, X. Inhibitory effects of four carbamate insecticides on acetylcholinesterase of male and female Carassius auratus in vitro. Comp. Biochem. Phys. C 2006, 143, 113–116. [Google Scholar] [CrossRef]
- Ren, Q.; Zhao, R.; Wang, C.; Li, S.; Zhang, T.; Ren, Z.; Yang, M.; Pan, H.; Xu, S.; Zhu, J.; et al. The role of AChE in swimming behavior of Daphnia magna: Correlation analysis of both parameters affected by deltamethrin and methomyl exposure. J. Toxicol. 2017, 2017, 3265727. [Google Scholar] [CrossRef] [Green Version]
- Abbas, N.; Shad, S.A. Assessment of resistance risk to lambda-cyhalothrin and cross-resistance to four other insecticides in the house fly, Musca domestica L. (Diptera: Muscidae). Parasitol. Res. 2015, 114, 2629–2637. [Google Scholar] [CrossRef] [PubMed]
- Mutunga, J.M.; Anderson, T.D.; Craft, D.T.; Gross, A.D.; Swale, D.R.; Tong, F.; Wong, D.W.; Carlier, P.R.; Bloomquist, J.R. Carbamate and pyrethroid resistance in the akron strain of Anopheles gambiae. Pestic. Biochem. Phys. 2015, 121, 116–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, S.S.; Ndula, M.; Riveron, J.M.; Irving, H.; Wondji, C.S. The P450 CYP6Z1 confers carbamate/pyrethroid cross-resistance in a major African malaria vector beside a novel carbamate-insensitive N485I acetylcholinesterase-1 mutation. Mol. Ecol. 2016, 25, 3436–3452. [Google Scholar] [CrossRef] [PubMed]
- Selvam, A.; Gnana, D.; Thatheyu, A.J.; Vidhya, R. Biodegradation of the synthetic pyrethroid, fenvalerate by Bacillus cereus Mtcc 1305. World J. Environ. Eng. 2013, 1, 21–26. [Google Scholar]
- Malato, S.; Blanco, J.; Cáceres, J.; Fernández-Alba, A.R.; Agüera, A.; Rodríguez, A. Photocatalytic treatment of water-soluble pesticides by photo-Fenton and TiO2 using solar energy. Catal. Today. 2002, 76, 209–220. [Google Scholar] [CrossRef]
- Yang, G.; Zhao, Y.; Lu, X.; Gao, X. Adsorption of methomyl on marine sediments. Colloids Surf. A Physicochem. Eng. Asp. 2005, 264, 179–186. [Google Scholar] [CrossRef]
- Van Scoy, A.R.; Yue, M.; Deng, X.; Tjeerdema, R.S. Environmental fate and toxicology of methomyl. Rev. Environ. Contam. Toxicol. 2013, 222, 93–109. [Google Scholar] [PubMed]
- Wu, X.; Sun, X.; Zhang, C.; Gong, C.; Hu, J. Micro-mechanism and rate constants for OH-initiated degradation of methomyl in atmosphere. Chemosphere 2014, 107, 331–335. [Google Scholar] [CrossRef]
- Meng, S.; Qiu, L.; Hu, G.; Fan, L.; Song, C.; Zheng, Y.; Wu, W.; Qu, J.; Li, D.; Chen, J.; et al. Effect of methomyl on sex steroid hormone and vitellogenin levels in serum of male tilapia (Oreochromis niloticus) and recovery pattern. Environ. Toxicol. 2017, 32, 1869–1877. [Google Scholar] [CrossRef]
- Waret, T.; Supap, S.; Kanokporn, S.; Monruedee, C. Lethal and sublethal effects of a methomyl-based insecticide in Hoplobatrachus rugulosus. J. Toxicol. Pathol. 2017, 30, 15–24. [Google Scholar]
- Seleem, A.A. Induction of hyperpigmentation and heat shock protein 70 response to the toxicity of methomyl insecticide during the organ development of the Arabian toad, Bufo arabicus (Heyden, 1827). J. Histotechnol. 2019, 42, 104–115. [Google Scholar] [CrossRef]
- Boucaud-Maitre, D.; Rambourg, M.; Sinno-Tellier, S.; Puskarczyk, E.; Pineau, X.; Kammerer, M.; Bloch, J.; Langrand, J. Human exposure to banned pesticides reported to the French Poison Control Centers: 2012–2016. Environ. Toxicol. Pharmacol. 2019, 69, 51–56. [Google Scholar] [CrossRef]
- Driskell, W.J.; Groce, D.F.; Hill, J.R.H. Methomyl in the blood of a pilot who crashed during aerial spraying. J. Anal. Toxicol. 1991, 15, 339–340. [Google Scholar] [CrossRef]
- Hoizey, G.; Canas, F.; Binet, L.; Kaltenbach, M.L.; Jeunehomme, G.; Bernard, M.; Lamiable, D. Thiodicarb and methomyl tissue distribution in a fatal multiple compounds poisoning. J. Forensic Sci. 2008, 53, 499–502. [Google Scholar] [CrossRef]
- Lin, C. Methomyl poisoning presenting with decorticate posture and cortical blindness. Neurol. Int. 2014, 6, 5307. [Google Scholar] [CrossRef] [PubMed]
- Tamimi, M.; Qourzal, S.; Barka, N.; Assabbane, A.; AitichouI, Y. Methomyl degradation in aqueous solutions by Fenton’s reagent and the photo-Fenton system. Sep. Purif. Technol. 2008, 61, 103–108. [Google Scholar] [CrossRef]
- Sayyaadi, H. Enhanced cavitation-oxidation process of non-VOC aqueous solution using hydrodynamic cavitation reactor. Chem. Eng. J. 2015, 272, 79–91. [Google Scholar] [CrossRef]
- Raut-Jadhav, S.; Saini, D.; Sonawane, S.; Pandit, A. Effect of process intensifying parameters on the hydrodynamic cavitation based degradation of commercial pesticide (methomyl) in the aqueous solution. Ultrason. Sonochem. 2016, 28, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wu, J.; Wang, Z.; Wang, K.; Li, M.; Jiang, J.; He, J.; Li, S. Isolation and characterization of a methomyl-degrading Paracoccus sp. mdw-1. Pedosphere. 2009, 2, 238–243. [Google Scholar] [CrossRef]
- Nyakundi, W.O.; Magoma, G.; Ochora, J.; Nyende, A.B. Biodegradation of diazinon and methomyl pesticides by white rot fungi from selected horticultural farms in rift valley and central Kenya. J. Appl. Tech. Environ. Sanit. 2012, 1, 107–124. [Google Scholar]
- Chen, C.; Wu, T.; Wang, H.; Wu, S.; Tien, C. The ability of immobilized bacterial consortia and strains from river biofilms to degrade the carbamate pesticide methomyl. Int. J. Environ. Sci. Technol. 2015, 12, 2857–2866. [Google Scholar] [CrossRef] [Green Version]
- Kawakami, Y.; Fuke, C.; Fukasawa, M.; Ninomiya, K.; Ihama, Y.; Miyazaki, T. An experimental study of postmortem decomposition of methomyl in blood. Legal Med. Tokyo. 2017, 25, 36–42. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Yang, Z.; Jin, W.; Wang, X.; Zhang, Y.; Zhu, S.; Yu, X.; Hu, G.; Hong, Q. Degradation of methomyl by the combination of Aminobacter sp. MDW-2 and Afipia sp. MDW-3. Lett. Appl. Microbiol. 2017, 64, 289–296. [Google Scholar] [CrossRef]
- Mohamed, M.S. Degradation of methomyl by the novel bacterial strain Stenotrophomonas maltophilia M1. Electron. J. Biotechn. 2009, 12, 6–7. [Google Scholar] [CrossRef] [Green Version]
- Zhan, H.; Feng, Y.; Fan, X.; Chen, S. Recent advances in glyphosate biodegradation. Appl. Microbiol. Biotechnol. 2018, 102, 5033–5043. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, P.; Huang, Y.; Zhan, H.; Chen, S. Insight into microbial applications for the biodegradation of pyrethroid insecticides. Front. Microbiol. 2019, 10, 1778. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zhan, H.; Bhatt, P.; Chen, S. Paraquat degradation from contaminated environments: Current achievements and perspectives. Front. Microbiol. 2019, 10, 1754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mano, H.; Tanaka, Y. Mechanisms of compensatory dynamics in zooplankton and maintenance of food chain efficiency under toxicant stress. Ecotoxicology 2016, 25, 399–411. [Google Scholar] [CrossRef]
- Islamy, R.A.; Yanuhar, U.; Hertika, A.M.S. Assessing the Genotoxic Potentials of Methomyl-based Pesticide in Tilapia (Oreochromis niloticus) Using Micronucleus Assay. J. Exp. Life Sci. 2017, 7, 88–93. [Google Scholar] [CrossRef] [Green Version]
- Meng, S.; Liu, T.; Chen, X.; Qiu, L.; Hu, G.; Song, C.; Xu, P. Effect of chronic exposure to methomyl on tissue damage and apoptosis in testis of tilapia (Oreochromis niloticus) and recovery pattern. Bull. Environ. Contam. Toxicol. 2019, 102, 371–376. [Google Scholar] [CrossRef]
- Meng, S.; Chen, J.; Hu, G.; Song, C.; Fan, L.; Qiu, L.; Xu, P. Effects of chronic exposure of methomyl on the antioxidant system in liver of Nile tilapia (Oreochromis niloticus). Ecotoxicol. Environ. Saf. 2014, 101, 1–6. [Google Scholar] [CrossRef]
- Meng, S.; Qiu, L.; Hu, G.; Fan, L.; Song, C.; Zheng, Y. Effects of methomyl on steroidogenic gene transcription of the hypothalamic-pituitary-gonad-liver axis in male tilapia. Chemosphere 2016, 165, 152–162. [Google Scholar] [CrossRef]
- Lau, E.T.; Karraker, N.E.; Leung, K.M. Temperature-dependent acute toxicity of methomyl pesticide on larvae of 3 Asian amphibian species. Environ. Toxicol. Chem. 2015, 34, 2322–2327. [Google Scholar] [CrossRef]
- Mahgoub, A.A.; El-Medany, A.H. Evaluation of chronic exposure of the male rat reproductive system to the insecticide methomyl. Pharmacol. Res. 2001, 44, 73–80. [Google Scholar] [CrossRef]
- Moser, V.C.; Phillips, P.M.; McDaniel, K.L. Assessment of biochemical and behavioral effects of carbaryl and methomyl in Brown-Norway rats from preweaning to senescence. Toxicology 2015, 331, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Shalaby, M.A.; El Zorba, H.Y.; Ziada, R.M. Reproductive toxicity of methomyl insecticide in male rats and protective effect of folic acid. Food Chem. Toxicol. 2010, 48, 3221–3226. [Google Scholar] [CrossRef] [PubMed]
- El-Fakharany, I.I.; Massoud, A.H.; Derbalah, A.S.; Allah, M.S.S. Toxicological effects of methomyl and remediation technologies of its residues in an aquatic system. J. Environ. Chem. Ecotoxicol. 2011, 3, 332–339. [Google Scholar]
- Xiang, G.; Li, D.; Yuan, J.; Guan, J.; Zhai, H.; Shi, M.; Tao, L. Carbamate insecticide methomyl confers cytotoxicity through DNA damage induction. Food Chem. Toxicol. 2013, 53, 352–358. [Google Scholar]
- Pantani, C.; Pannunzio, G.; De Cristofaro, M.; Novelli, A.A.; Salvatori, M. Comparative acute toxicity of some pesticides, metals, and surfactants to Gammarus italicus Goedm and Echinogammarus tibaldii pink and stock (Crustacea: Amphipoda). Bull. Environ. Contam. Toxicol. 1997, 59, 963–967. [Google Scholar] [CrossRef]
- Bridges, C.M. Long-term effects of pesticide exposure at various life stages of the southern leopard frog (Rana sphenocephala). Arch. Environ. Contam. Toxicol. 2000, 39, 91–96. [Google Scholar] [CrossRef]
- Seleem, A.A. Teratogenicity and neurotoxicity effects induced by methomyl insecticide on the developmental stages of Bufo arabicus. Neurotoxicol. Teratol. 2019, 72, 1–9. [Google Scholar] [CrossRef]
- Tsatsakis, A.M.; Tsakalof, A.K.; Siatitsas, Y.; Michalodimitrakis, E.N. Acute poisoning with carbamate pesticides: The Cretan experience. Sci. Justice 1996, 36, 35–39. [Google Scholar] [CrossRef]
- Sieliechi, J.M.; Thue, P.S. Removal of paraquat from drinking water by activated carbon prepared from waste wood. Desalin. Water Treat. 2014, 4, 986–998. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, L.; Yu, H.; Jiang, B.; Jiang, J. Comparison of sludge treatment by O3 and O3/H2O2. Water Sci. Technol. 2014, 70, 114–119. [Google Scholar]
- Pan, X.; Chen, X.; Yi, Z. Defective, Porous TiO2 nanosheets with Pt decoration as an efficient photocatalyst for ethylene oxidation synthesized by a C3N4 templating method. ACS Appl. Mater. Interfaces 2016, 8, 10104–10108. [Google Scholar] [CrossRef] [PubMed]
- Esplugas, S.; Giménez, J.; Contreras, S.; Pascual, E.; Rodríguez, M. Comparison of different advanced oxidation processes for phenol degradation. Water Res. 2002, 36, 1034–1042. [Google Scholar] [CrossRef]
- Micó, M.M.; Bacardit, J.; Malfeito, J.; Sans, C. Enhancement of pesticide photo-Fenton oxidation at high salinities. Appl. Catal. B Environ. 2013, 132, 162–169. [Google Scholar] [CrossRef]
- Gao, Z.; Lin, Y.; Xu, B.; Pan, Y.; Xia, S.; Gao, N.; Zhang, T.; Chen, M. Degradation of acrylamide by the UV/chlorine advanced oxidation process. Chemosphere 2017, 187, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Javier Benitez, F.; Real, F.J.; Acero, J.L.; Casas, F. Assessment of the UV/Cl2 advanced oxidation process for the degradation of the emerging contaminants amitriptyline hydrochloride, methyl salicylate and 2-phenoxyethanol in water systems. Environ. Technol. 2017, 38, 2508–2516. [Google Scholar] [CrossRef] [PubMed]
- Wardenier, N.; Liu, Z.; Nikiforov, A.; Van Hulle, S.W.H.; Leys, C. Micropollutant elimination by O3, UV and plasma-based AOPs: An evaluation of treatment and energy costs. Chemosphere 2019, 234, 715–724. [Google Scholar] [CrossRef]
- Tomašević, A.; Kiss, E.; Petrović, S.; Mijin, D. Study on the photocatalytic degradation of insecticide methomyl in water. Desalination 2010, 262, 228–234. [Google Scholar] [CrossRef]
- El-Geundi, M.S.; Nassar, M.M.; Farrag, T.E.; Ahmed, M.H. Methomyl adsorption onto cotton stalks activated carbon (csac): Equilibrium and process design. Procedia Environ. Sci. 2013, 17, 630–639. [Google Scholar] [CrossRef] [Green Version]
- Grgur, B.N.; Mijin, D. A kinetics study of the methomyl electrochemical degradation in the chloride containing solutions. Appl. Catal. B Environ. 2014, 147, 429–438. [Google Scholar] [CrossRef]
- Tamimi, M.; Qourzal, S.; Assabbane, A.; Chovelon, J.M.; Ferronato, C.; Ait-Ichou, Y. Photocatalytic degradation of pesticide methomyl: Determination of the reaction pathway and identification of intermediate products. Photochem. Photobio. Sci. 2006, 5, 477. [Google Scholar] [CrossRef]
- Chang, C.; Trinh, C.; Chiu, C.; Chang, C.; Chiang, S.; Ji, D.; Tseng, J.; Chang, C.; Chen, Y. UV-C irradiation enhanced ozonation for the treatment of hazardous insecticide methomyl. J. Taiwan Inst. Chem. Eng. 2015, 49, 100–104. [Google Scholar] [CrossRef]
- Malato, S.; Blanco, J.; Vidal, A.; Richter, C. Photocatalysis with solar energy at a pilot-plant scale: An overview. Appl. Catal. B Environ. 2002, 37, 1–15. [Google Scholar] [CrossRef]
- Barakat, N.A.M.; Nassar, M.M.; Farrag, T.E.; Mahmoud, M.S. Effective photodegradation of methomyl pesticide in concentrated solutions by novel enhancement of the photocatalytic activity of TiO2 using CdSO4 nanoparticles. Environ. Sci. Pollut. Res. 2014, 21, 1425–1435. [Google Scholar] [CrossRef]
- Fan, C.; Horng, C.; Li, S. Structural characterization of natural organic matter and its impact on methomyl removal efficiency in Fenton process. Chemosphere 2013, 93, 178–183. [Google Scholar] [CrossRef]
- Juang, R.; Chen, C. Comparative study on photocatalytic degradation of methomyl and parathion over UV-irradiated TiO2 particles in aqueous solutions. J. Taiwan Inst. Chem. E 2014, 45, 989–995. [Google Scholar] [CrossRef]
- Raut-Jadhav, S.; Pinjari, D.V.; Saini, D.R.; Sonawane, S.H.; Pandit, A.B. Intensification of degradation of methomyl (carbamate group pesticide) by using the combination of ultrasonic cavitation and process intensifying additives. Ultrason. Sonochem. 2016, 31, 135–142. [Google Scholar] [CrossRef]
- Gogate, P.R. Treatment of wastewater streams containing phenolic compounds using hybrid techniques based on cavitation: A review of the current status and the way forward. Ultrason. Sonochem. 2008, 15, 1–15. [Google Scholar] [CrossRef]
- Sutkar, V.S.; Gogate, P.R. Design aspects of sonochemical reactors: Techniques for understanding cavitational activity distribution and effect of operating parameters. Chem. Eng. J. 2009, 155, 26–36. [Google Scholar] [CrossRef]
- Villaroel, E.; Silva-Agredo, J.; Petrier, C.; Taborda, G.; Torres-Palma, R.A. Ultrasonic degradation of acetaminophen in water: Effect of sonochemical parameters and water matrix. Ultrason. Sonochem. 2014, 21, 1763–1769. [Google Scholar] [CrossRef]
- Chen, S.; Chang, C.; Deng, Y.; An, S.; Dong, Y.; Zhou, J.; Hu, M.; Zhong, G.; Zhang, L. Fenpropathrin biodegradation pathway in Bacillus sp. DG-02 and its potential for bioremediation of pyrethroid-contaminated soils. J. Agric. Food Chem. 2014, 62, 2147–2157. [Google Scholar] [CrossRef]
- Birolli, W.G.; Alvarenga, N.; Seleghim, M.H.R.; Porto, A.L.M. Biodegradation of the pyrethroid pesticide esfenvalerate by marine-derived fungi. Mar. Biotechnol. 2016, 18, 511–520. [Google Scholar] [CrossRef]
- Chen, S.; Yang, L.; Hu, M.; Liu, J. Biodegradation of fenvalerate and 3-phenoxybenzoic acid by a novel Stenotrophomonas sp. strain ZS-S-01 and its use in bioremediation of contaminated soils. Appl. Microbiol. Biotechnol. 2011, 90, 755–767. [Google Scholar] [CrossRef]
- Zhan, H.; Wang, H.; Liao, L.; Feng, Y.; Fan, X.; Zhang, L.H.; Chen, S. Kinetics and novel degradation pathway of permethrin in Acinetobacter baumannii ZH-14. Front. Microbiol. 2018, 9, 98. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Rodríguez, C.E.; Madrigal-León, K.; Masís-Mora, M.; Pérez-Villanueva, M.; Chin-Pampillo, J.S. Removal of carbamates and detoxification potential in a biomixture: Fungal bioaugmentation versus traditional use. Ecotoxicol. Environ. Saf. 2017, 135, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Farré, M.; Fernandez, J.; Paez, M.; Granada, L.; Barba, L.; Gutierrez, H.; Pulgarin, C.; Barceló, D. Analysis and toxicity of methomyl and ametryn after biodegradation. Anal. Bioanal. Chem. 2002, 373, 704–709. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Hong, Q.; Han, P.; Dong, X.; Shen, Y.; Li, S. Isolation and characterization of a carbofuran-degrading strain Novosphingobium sp. FND-3. FEMS Microbiol. Lett. 2007, 271, 207–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, X.; Zhang, J.; Li, Y.; Li, W.; Xu, G.; Yan, Y. Biodegradation of insecticide carbofuran by Paracoccus sp. YM3. J. Environ. Sci. Health B 1997, 43, 588–594. [Google Scholar] [CrossRef]
- Omolo, K.M. Characterization of methomyl and carbofuran degrading-bacteria from soils of horticultural farms in Rift Valley and Central Kenya. Afr. J. Environ. Sci. Technol. 2012, 6, 104–114. [Google Scholar]
- Tien, C.J.; Lin, M.; Chiu, W.H.; Chen, C. Biodegradation of carbamate pesticides by natural river biofilms in different seasons and their effects on biofilm community structure. Environ. Pollut. 2013, 179, 95–104. [Google Scholar] [CrossRef]
- Kulkarni, A.G.; Kaliwal, B.B. Bioremediation of methomyl by soil isolate—Pseudomonas Aeruginosa. J. Environ. Sci. Toxicol. Food Technol. 2014, 8, 1–10. [Google Scholar] [CrossRef]
- Ferreira, L.; Rosales, E.; Angeles Sanroman, M.; Pazos, M.M. Scale-up of removal process using a remediating-bacterium isolated from marine coastal sediment. RSC Adv. 2015, 5, 36665–36672. [Google Scholar] [CrossRef]
- Castro-Gutiérrez, V.; Masís-Mora, M.; Caminal, G.; Vicent, T.; Carazo-Rojas, E.; Mora-López, M.; Rodríguez-Rodríguez, C.E. A microbial consortium from a biomixture swiftly degrades high concentrations of carbofuran in fluidized-bed reactors. Process Biochem. 2016, 51, 1585–1593. [Google Scholar] [CrossRef]
- Konstantina, R.; Eleni, C.; Dafne, G.; Eftychia, S.; Demetra, K.; Panagiotis, K.; Spyridon, N.; Maria, T.; Emmanuel, A.T.; Dimitrios, G.K. Isolation of oxamyl-degrading bacteria and identification of cehA as a novel oxamyl hydrolase gene. Front. Microbiol. 2016, 7, 616. [Google Scholar]
- Roy, T.; Das, N. Isolation, characterization, and identification of two methomyl-degrading bacteria from a pesticide-treated crop field in west Bengal, India. Microbiology 2017, 6, 753–764. [Google Scholar] [CrossRef]
- Gong, T.; Xu, X.; Dang, Y.; Kong, A.; Wu, Y.; Liang, P.; Wang, S.; Yu, H.; Xu, P.; Yang, C. An engineered Pseudomonas putida can simultaneously degrade organophosphates, pyrethroids and carbamates. Sci. Total Environ. 2018, 628, 1258–1265. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, A.G.; Kaliwal, B.B. Bioremediation of methomyl by Escherichia coli. In Toxicity and Biodegradation Testing; Humana Press: New York, NY, USA, 2018; pp. 75–86. [Google Scholar]
- Roy, T.; Bandopadhyay, A.; Sonawane, P.J.; Majumdar, S.; Mahapatra, N.R.; Alam, S.; Das, N. Bio-effective disease control and plant growth promotion in lentil by two pesticide degrading strains of Bacillus sp. Biol. Control 2018, 127, 55–63. [Google Scholar] [CrossRef]
- Kaur, P.; Balomajumder, C. Simultaneous biodegradation of mixture of carbamates by newly isolated Ascochyta sp. CBS 237.37. Ecotoxicol. Environ. Saf. 2019, 169, 590–599. [Google Scholar] [CrossRef]
- Wen, X.; Jia, Y.; Li, J. Enzymatic degradation of tetracycline and oxytetracycline by crude manganese peroxidase prepared from Phanerochaete chrysosporium. J. Hazard. Mater. 2010, 177, 924–928. [Google Scholar] [CrossRef]
- Harms, H.; Schlosser, D.; Wick, L.Y. Untapped potential: Exploiting fungi in bioremediation of hazardous chemicals. Nat. Rev. Microbiol. 2011, 9, 177–192. [Google Scholar] [CrossRef]
- Chen, S.; Liu, C.; Peng, C.; Liu, H.; Hu, M.; Zhong, G. Biodegradation of chlorpyrifos and its hydrolysis product 3, 5, 6-trichloro-2-pyridinol by a new fungal strain Cladosporium cladosporioides Hu-01. PLoS ONE 2012, 7, e47205. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Hu, Q.; Hu, M.; Luo, J.; Weng, Q.; Lai, K. Isolation and characterization of a fungus able to degrade pyrethroids and 3-phenoxybenzaldehyde. Bioresour. Technol. 2011, 102, 8110–8116. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Zeng, G.; Chen, G.; Fan, J.; Zou, Z.; Li, H.; Hu, X.; Long, F. Simultaneous cadmium removal and 2,4-dichlorophenol degradation from aqueous solutions by Phanerochaete chrysosporium. Appl. Microbiol. Biotechnol. 2011, 91, 811–821. [Google Scholar] [CrossRef] [PubMed]
- Fragoeiro, S.; Magan, N. Impact of Trametes versicolor and Phanerochaete chrysosporium on differential breakdown of pesticide mixtures in soil microcosms at two water potentials and associated respiration and enzyme activity. Int. Biodeterior. Biodegrad. 2008, 62, 376–383. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Liang, M.; Liu, Y.; Fan, X. Directed evolution and secretory expression of a pyrethroid-hydrolyzing esterase with enhanced catalytic activity and thermostability. Microb. Cell Factories 2017, 16, 81. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Hu, M.; Liu, J.; Zhong, G.; Yang, L.; Rizwan-ul-Haq, M.; Han, H. Biodegradation of beta-cypermethrin and 3-phenoxybenzoic acid by a novel Ochrobactrum lupini DG-S-01. J. Hazard. Mater. 2011, 187, 433–440. [Google Scholar] [CrossRef]
- Yang, J.; Feng, Y.; Zhan, H.; Liu, J.; Zhang, K.; Zhang, L.H.; Chen, S. Characterization of a pyrethroid-degrading Pseudomonas fulva strain P31 and biochemical degradation pathway of D-phenothrin. Front. Microbiol. 2018, 9, 1003. [Google Scholar] [CrossRef]
- Chen, S.; Deng, Y.; Chang, C.; Lee, J.; Cheng, Y.; Cui, Z.; Zhou, J.; He, F.; Hu, M.; Zhang, L.H. Pathway and kinetics of cyhalothrin biodegradation by Bacillus thuringiensis strain ZS-19. Sci. Rep. 2015, 5, 8784. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Dong, Y.H.; Chang, C.; Deng, Y.; Zhang, X.F.; Zhong, G.; Song, H.; Hu, M.; Zhang, L.H. Characterization of a novel cyfluthrin-degrading bacterial strain Brevibacterium aureum and its biochemical degradation pathway. Bioresour. Technol. 2013, 132, 16–23. [Google Scholar] [CrossRef]
- Bhatt, P.; Bhatt, K.; Huang, Y.; Lin, Z.; Chen, S. Esterase is a powerful tool for the biodegradation of pyrethroid insecticides. Chemosphere 2020, 244, 125507. [Google Scholar] [CrossRef]
- Zhan, H.; Huang, Y.; Lin, Z.; Bhatt, P.; Chen, S. New insights into the microbial degradation and catalytic mechanism of synthetic pyrethroids. Environ. Res. 2020, 182, 109138. [Google Scholar] [CrossRef]
- Nguyen, T.P.O.; Helbling, D.E.; Bers, K.; Fida, T.T.; Wattiez, R.; Kohler, H.E.; Springael, D.; Mot, R.D. Genetic and metabolic analysis of the carbofuran catabolic pathway in Novosphingobium sp. KN65.2. Appl. Microbiol. Biotechnol. 2014, 98, 8235–8252. [Google Scholar] [CrossRef] [PubMed]
- Öztürk, B.; Ghequire, M.; Nguyen, T.P.O.; De Mot, R.; Wattiez, R.; Springael, D. Expanded insecticide catabolic activity gained by a single nucleotide substitution in a bacterial carbamate hydrolase gene. Environ. Microbiol. 2016, 18, 4878–4887. [Google Scholar] [CrossRef] [PubMed]
- Fareed, A.; Zaffar, H.; Rashid, A.; Maroof Shah, M.; Naqvi, T.A. Biodegradation of N-methylated carbamates by free and immobilized cells of newly isolated strain Enterobacter cloacae strain TA7. Bioremediat. J. 2017, 21, 119–127. [Google Scholar] [CrossRef]
- Yang, L.; Chen, S.; Hu, M.; Liu, J. Biodegradation of carbofuran by Pichia anomala strain HQ-C-01 and its application for bioremediation of contaminated soils. Biol. Fert. Soils 2011, 47, 917–923. [Google Scholar] [CrossRef]
- Tomasek, P.H.; Karns, J.S. Cloning of a carbofuran hydrolase gene from Achromobacter sp. strain WM111 and its expression in gram-negative bacteria. J. Bacteriol. 1989, 171, 4038–4044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desaint, S.; Hartmann, A.; Parekh, N.R.; Fournier, J. Genetic diversity of carbofuran-degrading soil bacteria. FEMS Microbiol. Ecol. 2000, 34, 173–180. [Google Scholar] [CrossRef]
- Naqvi, T.; Cheesman, M.J.; Williams, M.R.; Campbell, P.M.; Ahmed, S.; Russell, R.J.; Scott, C.; Oakeshott, J.G. Heterologous expression of the methyl carbamate-degrading hydrolase MCD. J. Biotechnol. 2009, 144, 89–95. [Google Scholar] [CrossRef]
- Bhatt, P.; Huang, Y.; Zhang, W.; Sharma, A.; Chen, S. Enhanced cypermethrin degradation kinetics and metabolic pathway in Bacillus thuringiensis strain SG4. Microorganisms 2020, 8, 223. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.No. | Study Sample/Sample Sources | Concentration/Volume of Methomyl | Specific Statement | References |
|---|---|---|---|---|
| 1 | Tilapia | 3.2-10 mg L−1 | Genotoxicity caused by methomyl | [35] |
| 2 | Tilapia | 0.2-200 µg L−1 | Injury to and apoptosis of testicular tissue | [36] |
| 3 | Tilapia | 0.2-200 μg L−1 | Inhibition of the antioxidant system | [37] |
| 4 | Tilapia | 0.2-200 μg L−1 | Disruption of the endocrine system and genetic variation | [38] |
| 5 | Frogs | 8.69 mg L−1 | Reduced growth rates and tissue damage | [16] |
| 6 | Frogs | 10 mg L−1 | Methomyl induces teratogenicity and neurotoxicity | [17] |
| 7 | Frogs | 15.43 mg L−1 | Death of or deformations in tadpoles | [39] |
| 8 | Rats | 17 mg kg−1 | Inhibition of the reproductive system | [40] |
| 9 | Rats | 0.25-2.5 mg kg−1 | Inhibited activity of brain ChE and RBC ChE | [41] |
| 10 | Rats | 0.5-20 mg kg−1 | Inhibition of the reproductive system | [42] |
| 11 | Rats | 10 mg kg−1 | Inhibition of liver function and enzyme activity | [43] |
| 12 | Human | unknown | 17 people poisoned (2012–2016, France) | [18] |
| 13 | Human | 570 μg L−1 | Death by inhalation of too much methomyl | [19] |
| 14 | Human | Unknown | The person died after swallowing methomyl | [20] |
| 15 | Human | 300 cm3 | Reversible cortical blindness and continuous peeling | [21] |
| 16 | Cells | 6-30 mmol L−1 | DNA damage and apoptosis induced by methomyl | [44] |
| 17 | Zooplankton and fish | 8 μg L−1 | Reduction in the efficiency of the food chain in a Cr/Dg system | [34] |
| S.No. | Study Sample/Sample Sources | Physicochemical Method Used | Specific Statement | References |
|---|---|---|---|---|
| 1 | UV/TiO2 | Photocatalysis | 100% methomyl was degraded in 45 min | [60] |
| 2 | Photo-Fenton | AOPS | 100% methomyl was degraded in an hour | [22] |
| 3 | Fenton/Fe-ZSM-5 | Photocatalysis | Methomyl was completely degraded | [57] |
| 4 | Fenton/H2O2/UV | AOPS | Methomyl was degraded within 320 min | [43] |
| 5 | Fenton/HA | Photocatalysis | HA promotes the degradation of methomyl | [64] |
| 6 | Activated Carbon | Adsorption | Methomyl was removed in 2.5 h | [58] |
| 7 | US/Photo-Fenton | AOPS | Promotion of the degradation ability | [24] |
| 8 | TiO2 nanoparticles | Photocatalysis | Pesticide was removed in 1 h | [63] |
| 9 | UV/TiO2 | AOPS | Promotion of the degradation ability | [65] |
| 10 | HC/H2O2 | AOPS | Promotion of hydrodynamic cavitation | [23] |
| 11 | O3/UV | AOPS | UV can promote the degradation effect | [61] |
| 12 | HC/Fenton/O3 | AOPS | Promotion of methomyl degradation | [66] |
| 13 | DSA Ti/RuO2 electrode | Electrocatalysis | 90% methomyl was degraded within 0.5 h | [59] |
| S.No. | Strain Or Community | Sample Source | Detected Metabolites | Comments | References |
|---|---|---|---|---|---|
| 1 | Mixed microbial community | Activated sludge from a domestic wastewater treatment plant | Methomyl oxime | Methomyl and its intermediates were completely degraded on the 12th and the 28th day, respectively | [75] |
| 2 | Novosphingobium SP. FND3 | No data | No data | Degraded 63% methomyl within 16 h | [76] |
| 3 | Paracoccus sp. YM3 | Sludge from a wastewater treatment facility | No data | Strain removed more than 80% of methomyl (50 mg L−1) in 7 days | [77] |
| 4 | Stenotrophomonas maltophilia M1 | Irrigation sites in Egypt | No data | Bacteria can grow on methomyl (100 mg L−1) and can tolerate up to 1000 mg L−1 of methomyl in the presence of 0.05% glucose | [30] |
| 5 | Paracoccus sp. mdw-1 | Methomyl wastewater treatment plant | Methomyl oxime | 100 mg L−1 of methomyl was transformed into an unknown metabolite within 10 h | [25] |
| 6 | White-rot fungal isolates WR1, WR2, WR4, WR9, and WR15 | Rift-valley region and a Mountain region in Kenya | No data | Complete degradation of 50 mg L−1 of methomyl by a single strain in 100 days whereas mixed strains took only 50–60 days | [26] |
| 7 | Pseudomonas sp. EB20 | Water polluted by persistent organic pollutants in Egypt | No data | 77% of 10 mg L−1 of methomyl was degraded within 2 weeks | [43] |
| 8 | Flavobacterium, Alcaligenes | Horticultural farms in Rift Valley and Central Kenya | No data | Strains completely degraded methomyl and its metabolites within 40 days as compared to the control | [78] |
| 9 | A consortium of Gomphonema parvulum, Cymbella silesiaca, and Nitzschia dissipata | Tseng-Wen River | No data | Methomyl was efficiently removed by biofilms containing degrading micro-organisms and diatoms | [79] |
| 10 | Microbial communities | Natural river biofilms | No data | 91% of added methomyl (50 mg L−1) was removed in 7 days | [27] |
| 11 | Pseudomonas aeruginosa | Soil samples from Dharwad | No data | Methomyl was significantly decreased | [80] |
| 12 | Serratia plymuthica | Marine coastal sediment | No data | Bacterium showed an excellent ability to remove imidacloprid, methomyl, and fenamiphos | [81] |
| 13 | Bacillus cereus, Bacillus safensis | Pesticide-treated crop field in India | No data | B. cereus and B. safensis showed 88.25% and 77.5% of methomyl degradation, respectively, within 96 h | [82] |
| 14 | Pseudomonas | Banana plantation, Greece | No data | Transformed all tested carbamates including aldicarb and methomyl | [83] |
| 15 | Bacillus cereus, Pseudomonas aeruginosa | Human stool samples provided by volunteers | Dimethyl disulfide | Strains can generate large quantities of DMDS | [28] |
| 16 | Trametes versicolor | No data | No data | More than 99% methomyl was removed by the bioaugmentation of the strain | [74] |
| 17 | A consortium of Cupriavidus, Achromobacter and Pseudomonas genera | Biopurificati-on system | No data | Methomyl was completely degraded within 7 days | [84] |
| 18 | Aminobacter sp. MDW-2 and Afipia sp. MDW-3 | Wastewater treatment system of a pesticide manufacturer | Methomyl oxime, methyl carbamic acid | Strains MDW-2 and MDW-3 co-existed and completely degraded 50 mg L−1 of methomyl within 3 days | [29] |
| 19 | Pseudomonas putida KT2440 | Genome editing | No data | Strain simultaneously degraded organophosphates, pyrethroids, and carbamates | [85] |
| 20 | Escherichia coli | India | No data | Methomyl was efficiently degraded by Escherichia coli with a plasmid | [86] |
| 21 | Bacillus cereus, Bacillus safensis | No data | No data | Strains degraded methomyl, carbendazim, and imidacloprid in NB medium | [87] |
| 22 | Ascochyta sp. CBS 237.37 | Paddy and maize cultivated fields, India | No data | Strain removed 90.15% of 85 mg L−1 of carbamates in 40 days | [88] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Z.; Zhang, W.; Pang, S.; Huang, Y.; Mishra, S.; Bhatt, P.; Chen, S. Current Approaches to and Future Perspectives on Methomyl Degradation in Contaminated Soil/Water Environments. Molecules 2020, 25, 738. https://doi.org/10.3390/molecules25030738
Lin Z, Zhang W, Pang S, Huang Y, Mishra S, Bhatt P, Chen S. Current Approaches to and Future Perspectives on Methomyl Degradation in Contaminated Soil/Water Environments. Molecules. 2020; 25(3):738. https://doi.org/10.3390/molecules25030738
Chicago/Turabian StyleLin, Ziqiu, Wenping Zhang, Shimei Pang, Yaohua Huang, Sandhya Mishra, Pankaj Bhatt, and Shaohua Chen. 2020. "Current Approaches to and Future Perspectives on Methomyl Degradation in Contaminated Soil/Water Environments" Molecules 25, no. 3: 738. https://doi.org/10.3390/molecules25030738
APA StyleLin, Z., Zhang, W., Pang, S., Huang, Y., Mishra, S., Bhatt, P., & Chen, S. (2020). Current Approaches to and Future Perspectives on Methomyl Degradation in Contaminated Soil/Water Environments. Molecules, 25(3), 738. https://doi.org/10.3390/molecules25030738