Anticancer Activities of the Quinone-Methide Triterpenes Maytenin and 22-β-hydroxymaytenin Obtained from Cultivated Maytenus ilicifolia Roots Associated with Down-Regulation of miRNA-27a and miR-20a/miR-17-5p

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
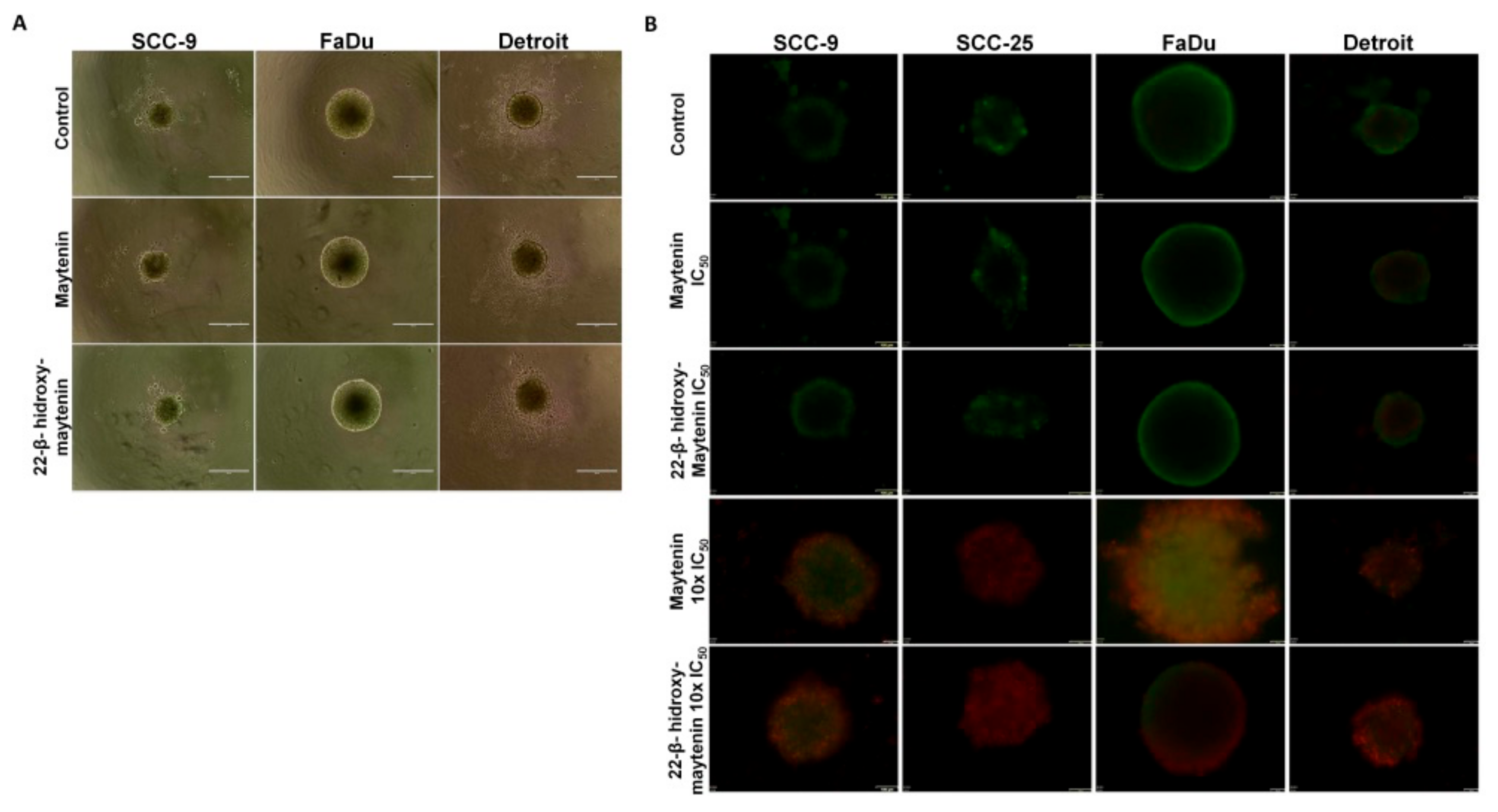
2. Results
2.1. Identification of Maytenin and 22-β-hydroxymaytenin in M. ilicifolia Root Extract
2.2. QMTs Cytotoxic Effects in Squamous Cell Carcinoma Cell Lines and in Oral Keratinocytes
2.3. QMTs and Cisplatin Effects on Cell Proliferation
2.4. Oxidative Stress Induction and Apoptotic Cell Death via Caspase 3/7 Activation
2.5. Maytenin and 22-β-hydroxymaytenin Intefere in miRNA Expression
2.6. Effects of Maytenin on Metastasis and Renal Tissue Toxicity in Human Tumor Xenograft
3. Discussion
4. Material and Methods
4.1. Maytenus Ilicifolia Root Cultivation In Vitro
4.2. Instrumentation and Ultra-High-Performance Liquid Chromatography–Diode Array Detector–Tandem Mass Spectrometry (UPLC-DAD-MS) Analytical Conditions for Raw Extract Analysis
4.3. Quantification of Maytenin in the Raw Extract Using UPLC-MS/MS
4.4. Cell Culture
4.5. Determination of the Half-Maximal Inhibitory Concentration (IC50) for QMT: MTT Assay and Formal Counting
4.6. Analysis of In Vitro Cell Proliferation by EdU Incorporation
4.7. Cellular Oxidative Stress Assay
4.8. Detection of Apoptotic Changes by Annexin V Assay
4.9. Caspase-3/7 Activation Assay
4.10. Formation of Spheroids with Cell Lines and Detection of Live and Dead Cells in their Outer Layers
4.11. miRNA Expression Profiling
4.12. Animal Experiments for Kidney Toxicity Analysis following Cisplatin and Maytenin Treatment
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Basaric, N.; Mlinaric-Majerski, K.; Kralj, M. Quinone methides: Photochemical generation and its application in biomedicine. Curr. Org. Chem. 2014, 18, 3–18. [Google Scholar] [CrossRef]
- Wang, P.; Song, Y.; Zhang, L.; He, H.; Zhou, X. Quinone methide derivatives: Important intermediates to DNA alkylating and DNA cross-linking actions. Curr. Med. Chem. 2005, 12, 2893–2913. [Google Scholar] [CrossRef]
- Shanmugam, M.K.; Nguyen, A.H.; Kumar, A.P.; Tan, B.K.; Sethi, G. Targeted inhibition of tumor proliferation, survival, and metastasis by pentacyclic triterpenoids: Potential role in prevention and therapy of cancer. Cancer Lett. 2012, 320, 158–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paz, T.A.; dos Santos, V.A.; Inácio, M.C.; Pina, E.S.; Pereira, A.M.S.; Furlan, M. Production of the quinone-methide triterpene maytenin by in vitro adventitious roots of Peritassa campestris (Cambess.) AC Sm.(Celastraceae) and rapid detection and identification by APCI-IT-MS/MS. BioMed Res. Int. 2013, 2013, 485837. [Google Scholar] [CrossRef] [PubMed]
- Hernandes, C.; Pereira, A.M.S.; Severino, P. Compounds From Celastraceae Targeting Cancer Pathways and Their Potential Application in Head and Neck Squamous Cell Carcinoma: A Review. Curr. Genom. 2017, 18, 60–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safe, S.H.; Prather, P.L.; Brents, L.K.; Chadalapaka, G.; Jutooru, I. Unifying Mechanisms of Action of the Anticancer Activities of Triterpenoids and Synthetic Analogs. Anticancer Agents Med. Chem. 2012, 12, 1211–1220. [Google Scholar] [CrossRef] [PubMed]
- Vigneswaran, N.; Williams, M.D. Epidemiological Trends in Head and Neck Cancer and Aids in Diagnosis. Oral. Maxillofac Surg. Clin. N. Am. 2014, 26, 123–141. [Google Scholar] [CrossRef] [PubMed]
- Suh, Y.; Amelio, I.; Guerrero Urbano, T.; Tavassoli, M. Clinical update on cancer: Molecular oncology of head and neck cancer. Cell Death Dis. 2014, 5, e1018. [Google Scholar] [CrossRef] [Green Version]
- Pendleton, K.P.; Grandis, J.R. Cisplatin-Based Chemotherapy Options for Recurrent and/or Metastatic Squamous Cell Cancer of the Head and Neck. Clin. Med. Insights 2013, 2013, CMT-S10409. [Google Scholar] [CrossRef] [Green Version]
- Jennette, K.W.; Lippard, S.J.; Vassiliades, G.A.; Bauer, W.R. Metallointercalation reagents. 2-hydroxyethanethiolato(2,2′,2′-terpyridine)-platinum(II) monocation binds strongly to DNA by intercalation. Proc. Natl. Acad. Sci. USA 1974, 71, 3839–3843. [Google Scholar] [CrossRef] [Green Version]
- Basu, A.; Krishnamurthy, S. Cellular responses to Cisplatin-induced DNA damage. J. Nucleic Acids 2010, 2010, 201367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottesman, M.M.; Hall, M.D.; Liang, X.-J.; Shen, D.-W. Resistance to cisplatin results from multiple mechanisms in cancer cells. In Platinum and Other Heavy Metal Compounds in Cancer Chemotherapy; Humana Press, c/o Springer Science + Business Media: New York, NY, USA, 2009; pp. 83–88. [Google Scholar]
- Shen, D.W.; Pouliot, L.M.; Hall, M.D.; Gottesman, M.M. Cisplatin resistance: A cellular self-defense mechanism resulting from multiple epigenetic and genetic changes. Pharm. Rev 2012, 64, 706–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddik, Z.H. Cisplatin: Mode of cytotoxic action and molecular basis of resistance. Oncogene 2003, 22, 7265–7279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.M.; Kim, H.K.; Shim, W.; Anwar, M.A.; Kwon, J.W.; Kwon, H.K.; Kim, H.J.; Jeong, H.; Kim, H.M.; Hwang, D.; et al. Mechanism of Cisplatin-Induced Cytotoxicity Is Correlated to Impaired Metabolism Due to Mitochondrial ROS Generation. PLoS ONE 2015, 10, e0135083. [Google Scholar] [CrossRef] [Green Version]
- Argiris, A.; Harrington, K.J.; Tahara, M.; Schulten, J.; Chomette, P.; Ferreira Castro, A.; Licitra, L. Evidence-Based Treatment Options in Recurrent and/or Metastatic Squamous Cell Carcinoma of the Head and Neck. Front. Oncol. 2017, 7, 72. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, A.C.; Leal, F.; Sasse, A.D. Cost-effectiveness of cetuximab and panitumumab for chemotherapy-refractory metastatic colorectal cancer. PLoS ONE 2017, 12, e0175409. [Google Scholar] [CrossRef] [Green Version]
- Levinger, I.; Ventura, Y.; Vago, R. Life is three dimensional-as in vitro cancer cultures should be. Adv. Cancer Res. 2014, 121, 383–414. [Google Scholar] [CrossRef]
- Pabla, N.; Dong, Z. Cisplatin nephrotoxicity: Mechanisms and renoprotective strategies. Kidney Int. 2008, 73, 994–1007. [Google Scholar] [CrossRef] [Green Version]
- Sahu, B.D.; Rentam, K.K.; Putcha, U.K.; Kuncha, M.; Vegi, G.M.; Sistla, R. Carnosic acid attenuates renal injury in an experimental model of rat cisplatin-induced nephrotoxicity. Food Chem. Toxicol. 2011, 49, 3090–3097. [Google Scholar] [CrossRef]
- Park, W.; Amin, A.R.; Chen, Z.G.; Shin, D.M. New perspectives of curcumin in cancer prevention. Cancer Prev. Res. 2013, 6, 387–400. [Google Scholar] [CrossRef] [Green Version]
- Romero, R.; Erez, O.; Hüttemann, M.; Maymon, E.; Panaitescu, B.; Conde-Agudelo, A.; Pacora, P.; Yoon, B.H.; Grossman, L.I. Metformin, the aspirin of the 21st century: Its role in gestational diabetes mellitus, prevention of preeclampsia and cancer, and the promotion of longevity. Am. J. Obstet. Gynecol. 2017, 217, 282–302. [Google Scholar] [CrossRef] [PubMed]
- Rauf, A.; Imran, M.; Suleria, H.A.R.; Ahmad, B.; Peters, D.G.; Mubarak, M.S. A comprehensive review of the health perspectives of resveratrol. Food Funct. 2017, 8, 4284–4305. [Google Scholar] [CrossRef] [PubMed]
- Nair, V.; Sreevalsan, S.; Basha, R.; Abdelrahim, M.; Abudayyeh, A.; Hoffman, A.R.; Safe, S. Mechanism of metformin-dependent inhibition of mammalian target of rapamycin (mTOR) and Ras activity in pancreatic cancer: Role of specificity protein (Sp) transcription factors. J. Biol. Chem. 2014, 289, 27692–27701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, V.; Pathi, S.; Jutooru, I.; Sreevalsan, S.; Basha, R.; Abdelrahim, M.; Samudio, I.; Safe, S. Metformin inhibits pancreatic cancer cell and tumor growth and downregulates Sp transcription factors. Carcinogenesis 2013, 34, 2870–2879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gandhy, S.U.; Kim, K.; Larsen, L.; Rosengren, R.J.; Safe, S. Curcumin and synthetic analogs induce reactive oxygen species and decreases specificity protein (Sp) transcription factors by targeting microRNAs. BMC Cancer 2012, 12, 564. [Google Scholar] [CrossRef] [Green Version]
- Chadalapaka, G.; Jutooru, I.; Safe, S. Celastrol decreases specificity proteins (Sp) and fibroblast growth factor receptor-3 (FGFR3) in bladder cancer cells. Carcinogenesis 2012, 33, 886–894. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Chadalapaka, G.; Lee, S.-O.; Yamada, D.; Sastre-Garau, X.; Defossez, P.-A.; Park, Y.-Y.; Lee, J.-S.; Safe, S. Identification of oncogenic microRNA-17-92/ZBTB4/specificity protein axis in breast cancer. Oncogene 2011, 31, 1034–1044. [Google Scholar] [CrossRef] [Green Version]
- Mertens-Talcott, S.U.; Chintharlapalli, S.; Li, X.; Safe, S. The oncogenic microRNA-27a targets genes that regulate specificity protein transcription factors and the G2-M checkpoint in MDA-MB-231 breast cancer cells. Cancer Res. 2007, 67, 11001–11011. [Google Scholar] [CrossRef] [Green Version]
- Jutooru, I.; Chadalapaka, G.; Abdelrahim, M.; Basha, M.R.; Samudio, I.; Konopleva, M.; Andreeff, M.; Safe, S. Methyl 2-cyano-3,12-dioxooleana-1,9-dien-28-oate decreases specificity protein transcription factors and inhibits pancreatic tumor growth: Role of microRNA-27a. Mol. Pharm. 2010, 78, 226–236. [Google Scholar] [CrossRef] [Green Version]
- Chadalapaka, G.; Jutooru, I.; Burghardt, R.; Safe, S. Drugs that target specificity proteins downregulate epidermal growth factor receptor in bladder cancer cells. Mol. Cancer Res. 2010, 8, 739–750. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Chen, Y.; Tian, R.; Li, J.; Li, H.; Lv, T.; Yao, Q. miRNA-21 enhances chemoresistance to cisplatin in epithelial ovarian cancer by negatively regulating PTEN. Oncol. Lett. 2017, 14, 1807–1810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.M.; Huang, C.; Li, X.F.; Yu, M.Z.; He, Y.; Li, J. miR-21 confers cisplatin resistance in gastric cancer cells by regulating PTEN. Toxicology 2013, 306, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yang, C.; Sun, C.; Sun, Y.; Yang, Z.; Cheng, S.; Zhuge, B. miR-21-5p Suppressed the Sensitivity of Hepatocellular Carcinoma Cells to Cisplatin by Targeting FASLG. DNA Cell Biol. 2019, 38, 865–873. [Google Scholar] [CrossRef] [PubMed]
- Studwell, A.J.; Kotton, D.N. A shift from cell cultures to creatures: In vivo imaging of small animals in experimental regenerative medicine. Molecules 2011, 19, 1933–1941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mir, M.; Arab, M.R.; Shahraki, M.R.; Mashhadi, M.A.; Shahraki, S.M.; Sargolzaei, A.F.; Karimfar, M.H. Toxic effects of cisplatin on hepatocytes and liver enzymes of rats. Anat. Sci. 2015, 12, 171. [Google Scholar]
- El-Sayyad, H.; Ismail, M.F.; Shalaby, F.M.; Abou-El-Magd, R.; Gaur, R.L.; Fernando, A.; Raj, M.H.; Ouhtit, A. Histopathological effects of cisplatin, doxorubicin and 5-fluoracil (5-FU) on the liver of male albino rats. Int. J. Biol. Sci. 2009, 5, 466. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, R.C.; Alexandrino, H.; Cipriano, M.A.; Tralhão, J.G. Liver Metastases and Histological Growth Patterns: Biological Behavior and Potential Clinical Implications—Another Path to Individualized Medicine? J. Oncol. 2019, 25, 6280347. [Google Scholar]
- Kim, J.-S.; Park, S.; Kim, K.H.; Park, J.H.; Kim, W.; Choi, I.S.; Jung, Y.J.; Kim, H.Y. Changes in non-invasive liver fibrosis indices during chemotherapy: Potential marker for oxaliplatin-induced sinusoidal obstruction syndrome. J. Clin. Oncol. 2016, 95, e2454. [Google Scholar] [CrossRef]
- Coppede, J.S.; Pina, E.S.; Paz, T.A.; Fachin, A.L.; Marins, M.A.; Bertoni, B.W.; França, S.C.; Pereira, A.M.S. Cell cultures of Maytenus ilicifolia Mart. are richer sources of quinone-methide triterpenoids than plant roots in natura. Plant Celltissue Organ Cult. (PCTOC) 2014, 118, 33–43. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Lloyd, G.; McCown, B. Commercially-feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shoot-tip culture. Comb. Proc. Int. Plant Propag Soc. (USA) 1980, 30, 421–427. [Google Scholar]
- Agostini, M.; Almeida, L.Y.; Bastos, D.C.; Ortega, R.M.; Moreira, F.S.; Seguin, F.; Zecchin, K.G.; Raposo, H.F.; Oliveira, H.C.; Amoedo, N.D.; et al. The fatty acid synthase inhibitor orlistat reduces the growth and metastasis of orthotopic tongue oral squamous cell carcinomas. Mol. Cancer 2014, 13, 585–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sant’Anna-Silva, A.C.B.; Santos, G.C.; Campos, S.P.C.; Gomes, A.M.O.; Pérez-Valencia, J.A.; Rumjanek, F.D. Metabolic Profile of Oral Squamous Carcinoma Cell Lines Relies on a Higher Demand of Lipid Metabolism in Metastatic Cells. Front. Oncol. 2018, 8, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- He, Y.; Zhu, Q.; Chen, M.; Huang, Q.; Wang, W.; Li, Q.; Huang, Y.; Di, W. The changing 50% inhibitory concentration (IC50) of cisplatin: A pilot study on the artifacts of the MTT assay and the precise measurement of density-dependent chemoresistance in ovarian cancer. Oncotarget 2016, 7, 70803–70821. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Henkels, K.M.; Turchi, J.J. Cisplatin-induced apoptosis proceeds by caspase-3-dependent and -independent pathways in cisplatin-resistant and -sensitive human ovarian cancer cell lines. Cancer Res. 1999, 59, 3077–3083. [Google Scholar]
- de Andrade, N.P.; Rodrigues, M.F.; Rodini, C.O.; Nunes, F.D. Cancer stem cell, cytokeratins and epithelial to mesenchymal transition markers expression in oral squamous cell carcinoma derived from ortothopic xenoimplantation of CD44(high) cells. Pathol. Res. Pr. 2017, 213, 235–244. [Google Scholar] [CrossRef]
- Guindon, J.; Deng, L.; Fan, B.; Wager-Miller, J.; Hohmann, A.G. Optimization of a cisplatin model of chemotherapy-induced peripheral neuropathy in mice: Use of vitamin C and sodium bicarbonate pretreatments to reduce nephrotoxicity and improve animal health status. Mol. Pain 2014, 10, 56. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Q.W.; Cheng, K.J.; Mei, X.L.; Qiu, J.G.; Zhang, W.J.; Xue, Y.Q.; Qin, W.M.; Yang, Y.; Zheng, D.W.; Chen, Y.; et al. Synergistic anticancer effects of triptolide and celastrol, two main compounds from thunder god vine. Oncotarget 2015, 6, 32790–32804. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds maytenin and 22-β-hydroxymaytenin are available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 (µM) | |||
| Cell Types | Maytenin | 22-β-hydrodymaytenin | Cisplatin |
| SCC-9 | 1.5 ± 0.2 | 1.9 ± 0.2 | 59.1 ± 9.1 |
| SCC-25 | 1.5 ± 0.3 | 1.9 ± 0.1 | 21 ± 3 |
| FaDu | 1.6 ± 0.2 | 2.5 ± 0.3 | 33.5 ± 0.6 |
| keratinocytes | 6 ± 0.2 | 10.2 ± 0.4 | 85.6 ± 2.1 |
| Systematic miRNA Name | FC (SCC25 22b vs. SCC25 Untreated) | FC (SCC25 Maytenin vs. SCC25 Untreated) | FC (SCC25 Cisplatin vs. SCC25 Untreated) | miRBase Accession |
| miR-17-5p | −36.4 | −36.4 | −1.3 | MIMAT0000070 |
| miR-20a-5p | −88.1 | −88.1 | −1.1 | MIMAT0000075 |
| miR-27a-3p | −62.0 | −62.0 | −1.2 | MIMAT0000084 |
| Systematic miRNA Name | FC (Keratinocyte 22b vs. Keratinocyte Untreated) | FC (Keratinocyte Maytenin vs. Keratinocyte Untreated) | FC (Keratinocyte Cisplatin vs. Keratinocyte Untreated) | miRBase Accession |
| miR-17-5p | −1.0 | 55.4 | −1.0 | MIMAT0000070 |
| miR-20a-5p | 1.7 | 8.3 | 3.4 | MIMAT0000075 |
| miR-27a-3p | 1.9 | 7.9 | 2.9 | MIMAT0000084 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernandes, C.; Miguita, L.; de Sales, R.O.; Silva, E.d.P.; Mendonça, P.O.R.d.; Lorencini da Silva, B.; Klingbeil, M.d.F.G.; Mathor, M.B.; Rangel, E.B.; Marti, L.C.; et al. Anticancer Activities of the Quinone-Methide Triterpenes Maytenin and 22-β-hydroxymaytenin Obtained from Cultivated Maytenus ilicifolia Roots Associated with Down-Regulation of miRNA-27a and miR-20a/miR-17-5p. Molecules 2020, 25, 760. https://doi.org/10.3390/molecules25030760
Hernandes C, Miguita L, de Sales RO, Silva EdP, Mendonça PORd, Lorencini da Silva B, Klingbeil MdFG, Mathor MB, Rangel EB, Marti LC, et al. Anticancer Activities of the Quinone-Methide Triterpenes Maytenin and 22-β-hydroxymaytenin Obtained from Cultivated Maytenus ilicifolia Roots Associated with Down-Regulation of miRNA-27a and miR-20a/miR-17-5p. Molecules. 2020; 25(3):760. https://doi.org/10.3390/molecules25030760
Chicago/Turabian StyleHernandes, Camila, Lucyene Miguita, Romario Oliveira de Sales, Elisangela de Paula Silva, Pedro Omori Ribeiro de Mendonça, Bruna Lorencini da Silva, Maria de Fatima Guarizo Klingbeil, Monica Beatriz Mathor, Erika Bevilaqua Rangel, Luciana Cavalheiro Marti, and et al. 2020. "Anticancer Activities of the Quinone-Methide Triterpenes Maytenin and 22-β-hydroxymaytenin Obtained from Cultivated Maytenus ilicifolia Roots Associated with Down-Regulation of miRNA-27a and miR-20a/miR-17-5p" Molecules 25, no. 3: 760. https://doi.org/10.3390/molecules25030760
APA StyleHernandes, C., Miguita, L., de Sales, R. O., Silva, E. d. P., Mendonça, P. O. R. d., Lorencini da Silva, B., Klingbeil, M. d. F. G., Mathor, M. B., Rangel, E. B., Marti, L. C., Coppede, J. d. S., Nunes, F. D., Pereira, A. M. S., & Severino, P. (2020). Anticancer Activities of the Quinone-Methide Triterpenes Maytenin and 22-β-hydroxymaytenin Obtained from Cultivated Maytenus ilicifolia Roots Associated with Down-Regulation of miRNA-27a and miR-20a/miR-17-5p. Molecules, 25(3), 760. https://doi.org/10.3390/molecules25030760