Tepary Bean (Phaseolus acutifolius) Lectins Induce Apoptosis and Cell Arrest in G0/G1 by P53(Ser46) Phosphorylation in Colon Cancer Cells

 ,
, 
 ,
, 
Abstract
:
1. Introduction
2. Results
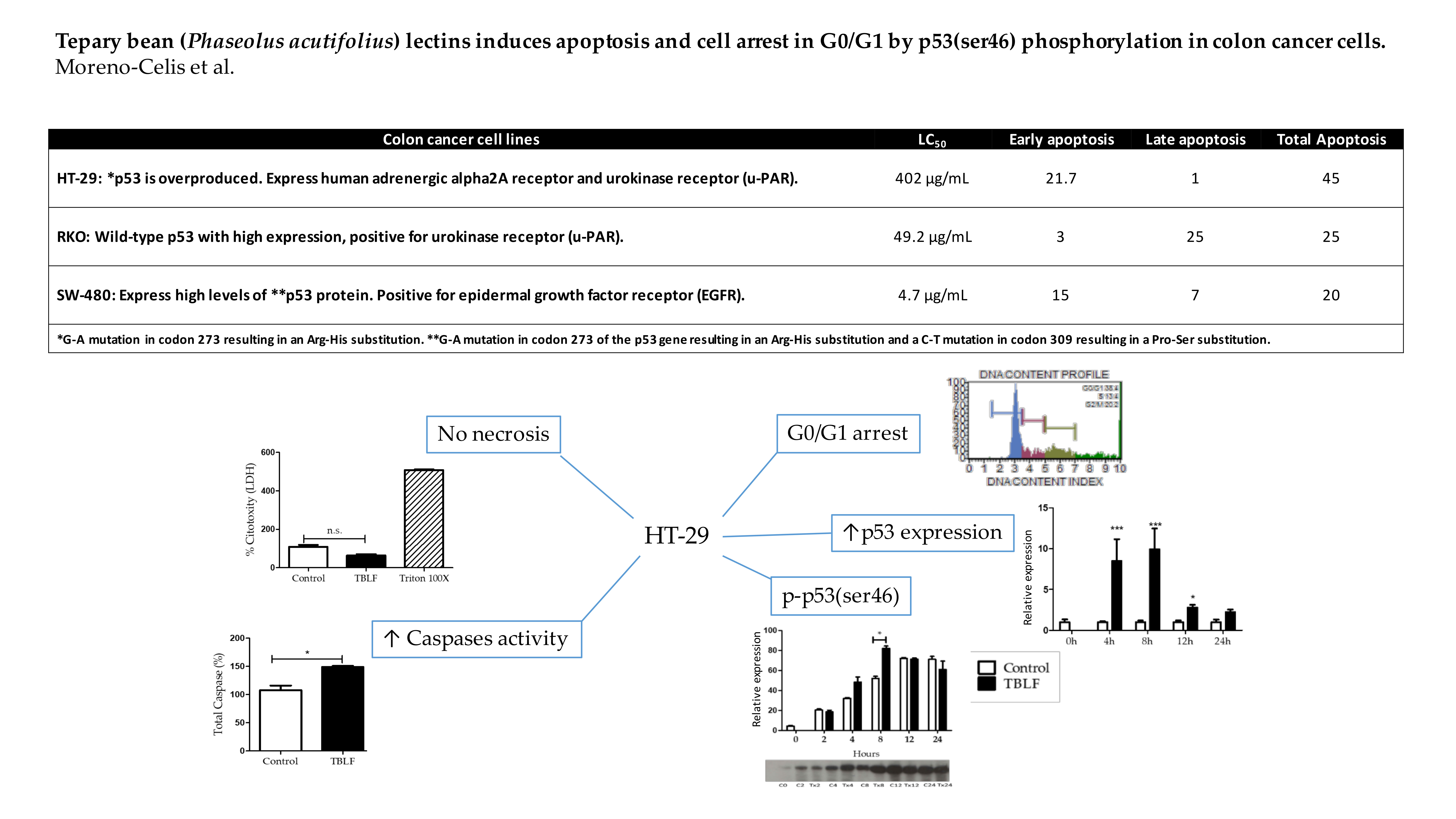
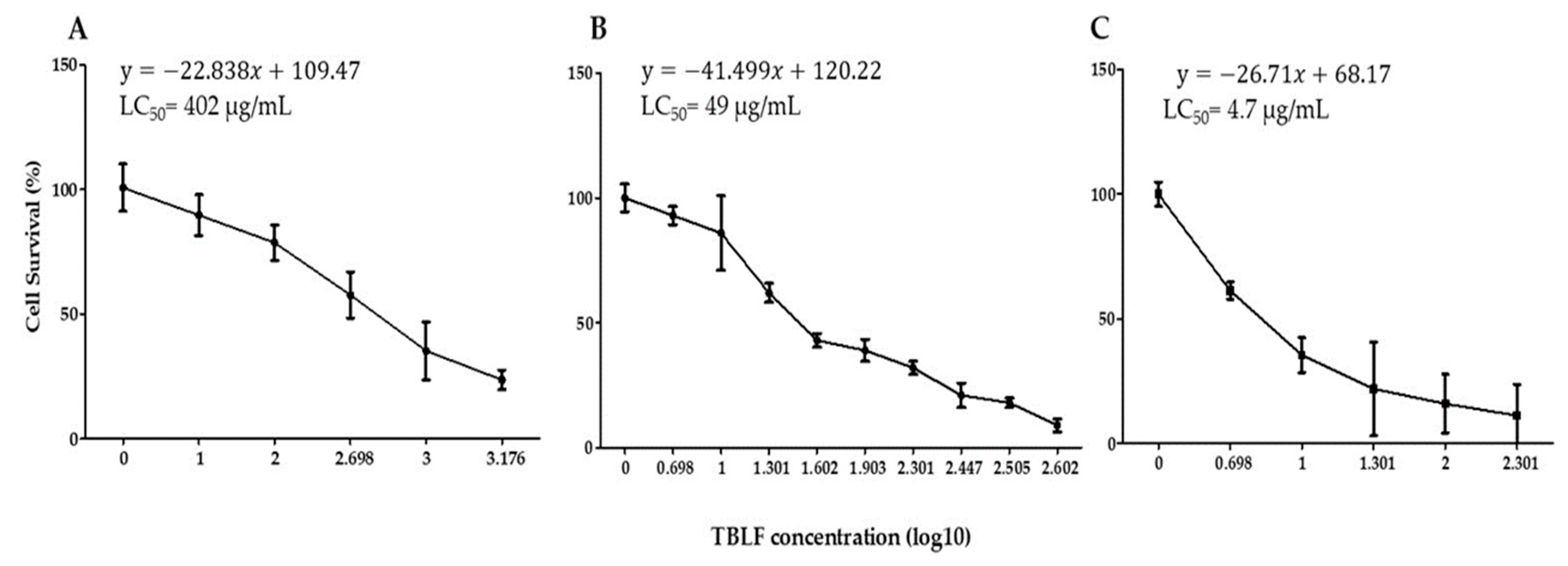
2.1. Concentration-Response Study and LC50 Determination
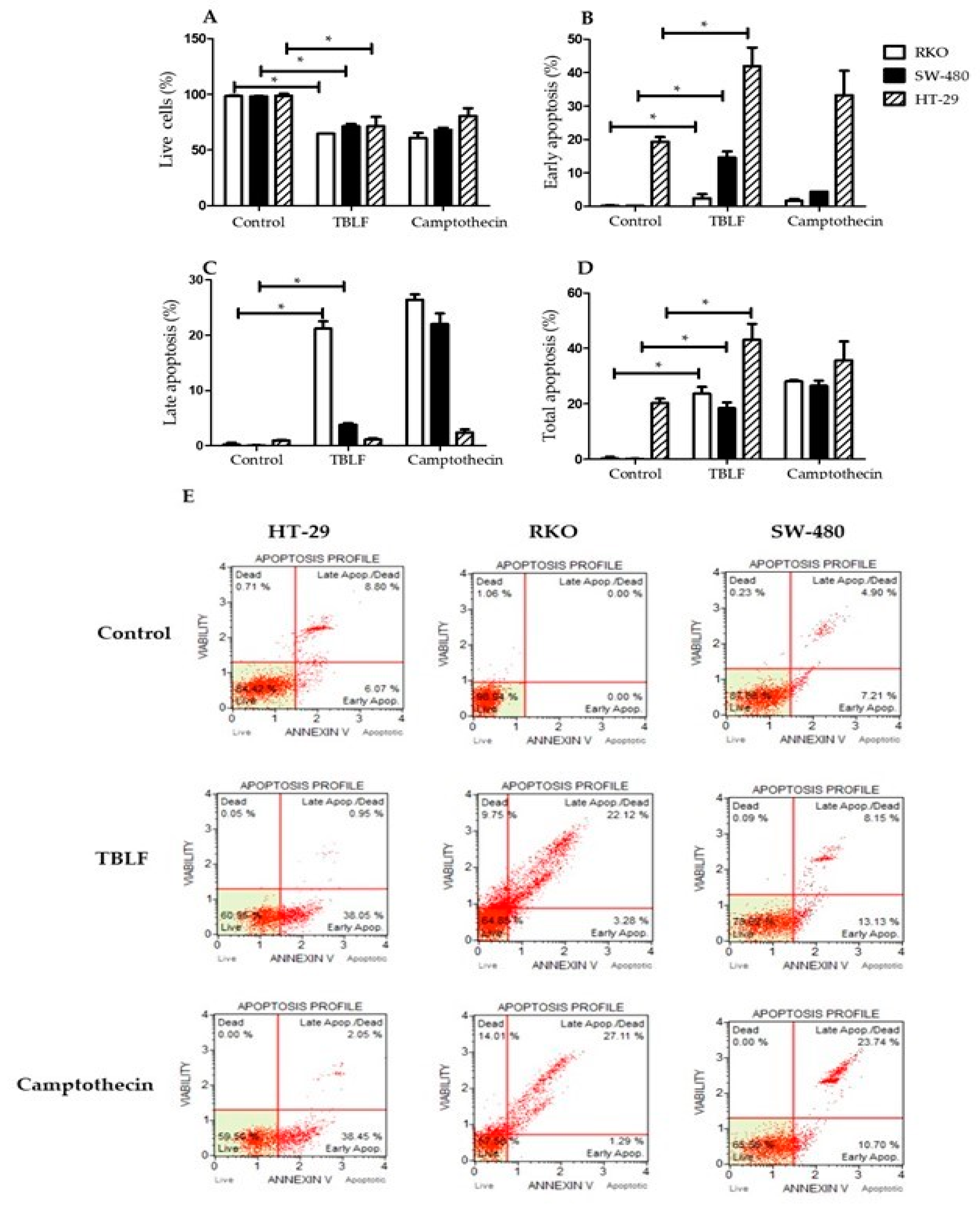
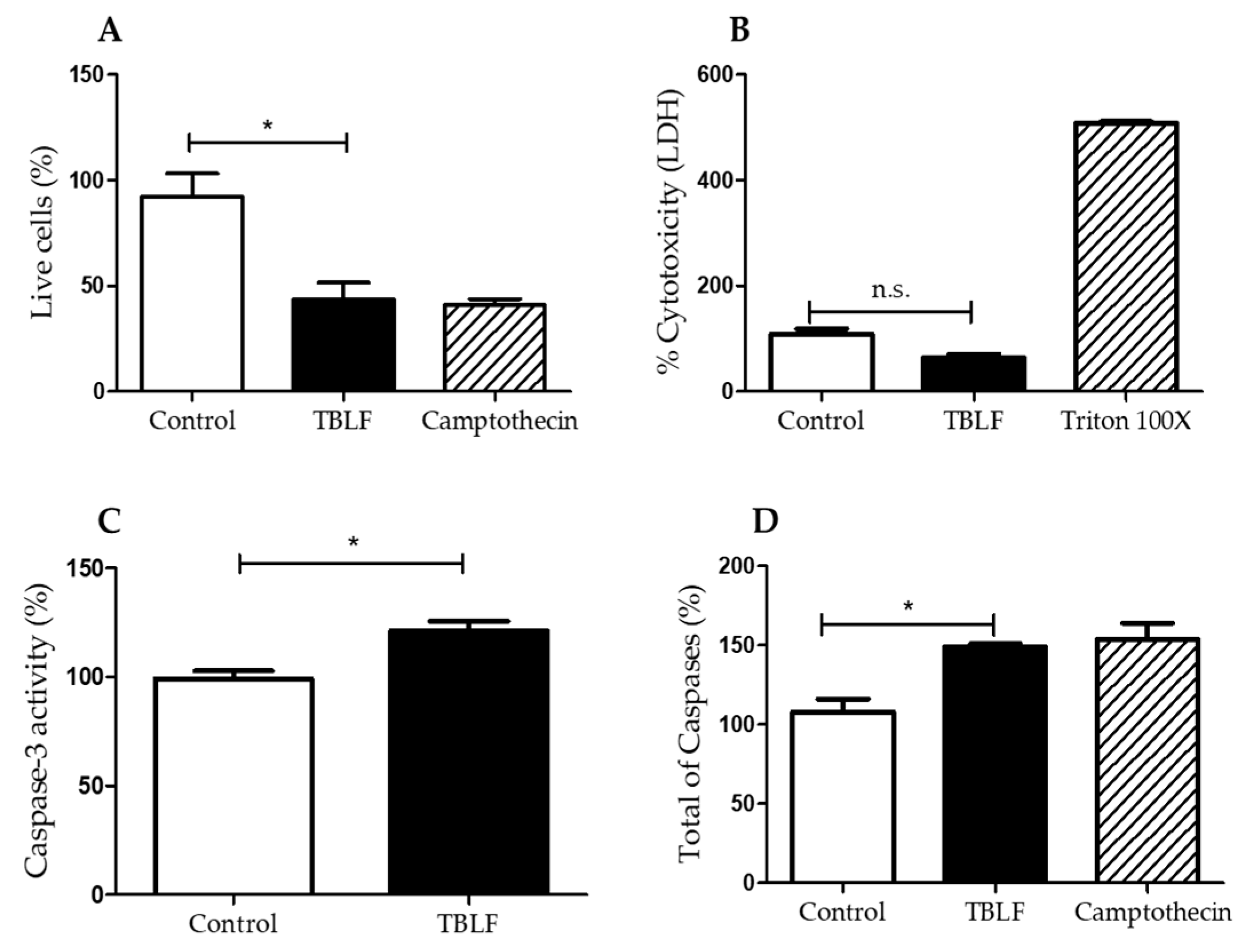
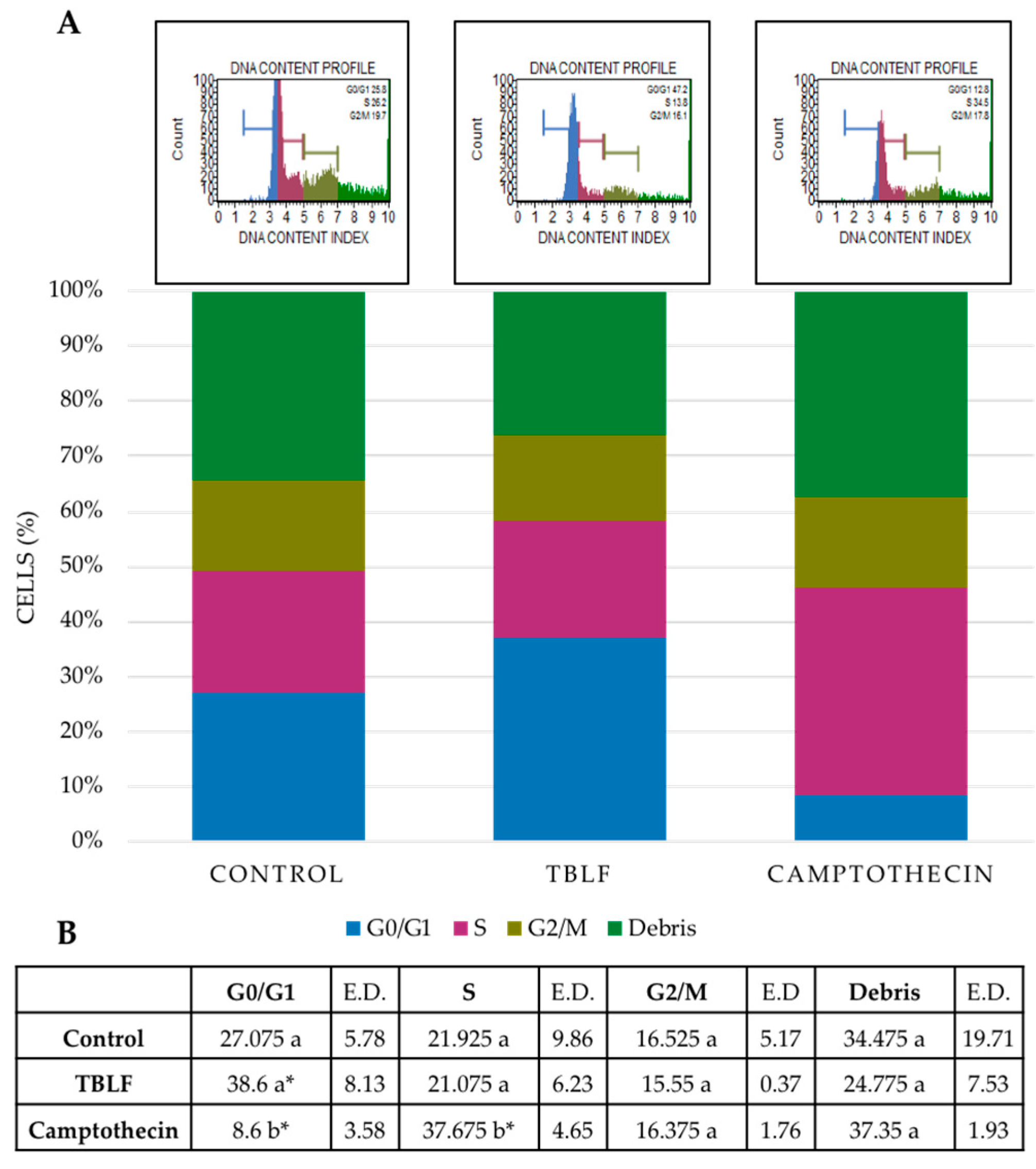
2.2. Effects on Cell Death and Cell Cycle Arrest
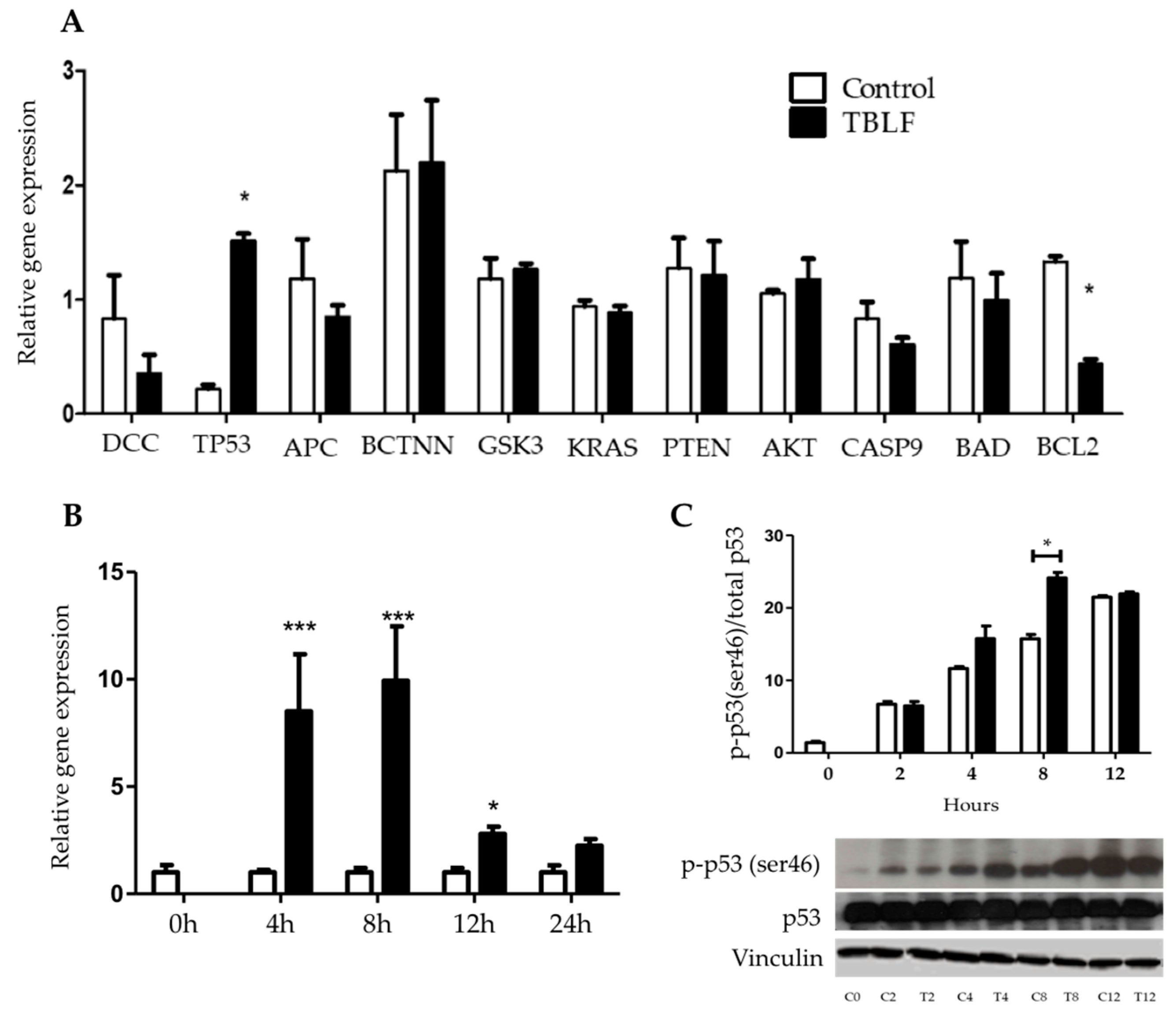
2.3. Apoptotic-Related Gene Expression and Phosphorylation of P53 in Ser46
3. Discussion
4. Materials and Methods
4.1. Obtaining TBLF
4.2. Cell Culture and Concentration–Response Assay
4.3. Evaluation of Apoptosis by Flow Cytometry
4.4. Necrosis Determination by Lactate Dehydrogenase Assay
4.5. Caspase- 3 Activity by Colorimetric Assay and Flow Cytometry Multi-Caspase Assay
4.6. Cell Cycle Analysis
4.7. Gene Expression Evaluation
4.8. Protein Determination by Western Blot
4.9. Statistics
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Estrada-Martínez, L.E.; Moreno-Celis, U.; Cervantes-Jiménez, R.; Ferriz-Martínez, R.A.; Blanco-Labra, A.; García-Gasca, T. Plant lectins as medical tools against digestive system cancers. Int. J. Mol. Sci. 2017, 18, 1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghazarian, H.; Idoni, B.; Oppenheimer, S.B. A glycobiology review: Carbohydrates, lectins and implications in cancer therapeutics. Acta Histochem. 2011, 113, 236–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nonaka, M.; Ma, B.Y.; Murai, R.; Nakamura, N.; Baba, M.; Kawasaki, N.; Hodohara, K.; Asano, S.; Kawasaki, T. Glycosylation-dependent interactions of C-type lectin DC-SIGN with colorectal tumor-associated Lewis glycans impair the function and differentiation of monocyte-derived dendritic cells. J. Immunol. 2008, 180, 3347–3356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferriz-martinez, R.A.; Torres-arteaga, I.C.; Blanco-labra, A.; Garcia-gasca, T. The role of plant lectins in cancer trarment. In New Approaches in the Treatment of Cancer; Mejia-Vazquez, C., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2010; pp. 71–89. ISBN 9781617283048. [Google Scholar]
- Fu, L.; Zhou, C.; Yao, S.; Yu, J.-Y.; Liu, B.; Bao, J.-K. Plant lectins: Targeting programmed cell death pathways as antitumor agents. Int. J. Biochem. Cell Biol. 2011, 43, 1442–1449. [Google Scholar] [CrossRef] [PubMed]
- Castillo-villanueva, A.; Abdullaev, F. Lectinas vegetales y sus efectos en el cáncer. Rev. Investig. Clínica 2005, 57, 55–64. [Google Scholar]
- Han, S.-Y.; Hong, C.-E.; Kim, H.-G.; Lyu, S.-Y. Anti-cancer effects of enteric-coated polymers containing mistletoe lectin in murine melanoma cells in vitro and in vivo. Mol. Cell. Biochem. 2015, 408, 14–16. [Google Scholar] [CrossRef]
- González De Mejía, E.; Prisecaru, V.I. Lectins as bioactive plant proteins: A potential in cancer treatment. Crit. Rev. Food Sci. Nutr. 2005, 45, 425–445. [Google Scholar] [CrossRef]
- Timoshenko, A.V.; Gorudko, I.V.; Gabius, H.-J. Lectins from Medicinal Plants: Bioeffectors with Diverse Activities. In Phytochemicals. Biosyntehsis, Function and Application; Jetter, R., Ed.; Springer International Publishing: Cham, Switzerland, 2014; pp. 43–57. ISBN 978-3-319-04044-8. [Google Scholar]
- Fang, E.F.; Pan, W.L.; Wong, J.H.; Chan, Y.S.; Ye, X.J.; Ng, T.B. A new Phaseolus vulgaris lectin induces selective toxicity on human liver carcinoma Hep G2 cells. Arch. Toxicol. 2011, 85, 1551–1563. [Google Scholar] [CrossRef]
- Deepa, M.; Sureshkumar, T.; Satheeshkumar, P.K.; Priya, S. Purified mulberry leaf lectin (MLL) induces apoptosis and cell cycle arrest in human breast cancer and colon cancer cells. Chem. Biol. Interact. 2012, 200, 38–44. [Google Scholar] [CrossRef]
- Delebinski, C.I.; Jaeger, S.; Kemnitz-Hassanin, K.; Henze, G.; Lode, H.N.; Seifert, G.J. A new development of triterpene acid-containing extracts from Viscum album L. displays synergistic induction of apoptosis in acute lymphoblastic leukaemia. Cell Prolif. 2012, 45, 176–187. [Google Scholar] [CrossRef]
- Shi, Z.; Chen, J.; Li, C.; An, N.; Wang, Z.; Yang, S.; Huang, K.; Bao, J. Antitumor effects of concanavalin A and Sophora flavescens lectin in vitro and in vivo. Acta Pharmacol. Sin. 2014, 35, 248–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabir, S.R.; Nabi, M.M.; Haque, A.; Zaman, R.U.; Mahmud, Z.H.; Reza, M.A. Pea lectin inhibits growth of Ehrlich ascites carcinoma cells by inducing apoptosis and G2/M cell cycle arrest in vivo in mice. Phytomedicine 2013, 20, 1288–1296. [Google Scholar] [CrossRef] [PubMed]
- García-Gasca, T.; García-Cruz, M.; Hernandez-Rivera, E.; López-Matínez, J.; Castañeda-Cuevas, A.L.; Yllescas-Gasca, L.; Rodríguez-Méndez, A.J.; Mendiola-Olaya, E.; Castro-Guillén, J.L.; Blanco-Labra, A. Effects of Tepary bean (Phaseolus acutifolius) protease inhibitor and semipure lectin fractions on cancer cells. Nutr. Cancer 2012, 64, 1269–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yau, T.; Dan, X.; Ng, C.C.W.; Ng, T.B. Lectins with potential for anti-cancer therapy. Molecules 2015, 20, 3791–3810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.H.; Lyu, S.Y.; Park, W.B. Mistletoe lectin induces apoptosis and telomerase inhibition in human A253 cancer cells through dephosphorylation of Akt. Arch. Pharm. Res. 2004, 27, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Rivera, E.H.; Olaya, E.M.; Blanco, A.; García, T. Efecto Citotóxico Diferencial de una Fracción Rica en Lectinas de Frijol Tépari (Phaseolus acutifolius) sobre Células Cancerígenas. In Proceedings of the 10° Verano de la Ciencia de la Región Centro; Univeresidad Autónoma de Querétaro: Queretaro, Mexico, 2008; pp. 1–4. [Google Scholar]
- Valadez-Vega, C.; Morales-González, J.; Sumaya-Martínez, M.; Delgado-Olivares, L.; Cruz-Castañeda, A.; Bautista, M.; Sánchez-Gutiérrez, M.; Zuñiga-Pérez, C. Cytotoxic and Antiproliferative Effect of Tepary Bean Lectins on C33-A, MCF-7, SKNSH, and SW480 Cell Lines. Molecules 2014, 19, 9610–9627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valadez-Vega, C.; Alvarez-Manilla, G.; Riverón-Negrete, L.; García-Carrancá, A.; Morales-González, J.A.; Zuñiga-Pérez, C.; Madrigal-Santillán, E.; Esquivel-Soto, J.; Esquivel-Chirino, C.; Villagómez-Ibarra, R.; et al. Detection of cytotoxic activity of lectin on human colon adenocarcinoma (Sw480) and epithelial cervical carcinoma (C33-A). Molecules 2011, 16, 2107–2118. [Google Scholar] [CrossRef] [Green Version]
- Alatorre-Cruz, J.M.; Pita-López, W.; López-Reyes, R.G.; Ferriz-Martínez, R.A.; Cervantes-Jiménez, R.; de Jesús Guerrero Carrillo, M.; Vargas, P.J.A.; López-Herrera, G.; Rodríguez-Méndez, A.J.; Zamora-Arroyo, A.; et al. Effects of intragastrically-administered Tepary bean lectins on digestive and immune organs: Preclinical evaluation. Toxicol. Reports 2018, 5, 56–64. [Google Scholar] [CrossRef]
- Ferriz-Martínez, R.; García-García, K.; Torres-Arteaga, I.; Rodriguez-Mendez, A.J.; de Guerrero-Carrillo, M.J.; Moreno-Celis, U.; Ángeles-Zaragoza, M.V.; Blanco-Labra, A.; Gallegos-Corona, M.A.; Robles-Álvarez, J.P.; et al. Tolerability assessment of a lectin fraction from Tepary bean seeds (Phaseolus acutifolius) orally administered to rats. Toxicol. Reports 2015, 2, 63–69. [Google Scholar]
- Moreno-Celis, U.; Lopez-Martinez, J.; Blanco-Labra, A.; Cervantes-Jimenez, R.; Estrada-Martinez, L.E.; Garcia-Pascalin, A.E.; Guerrero-Carrillo, M.D.J.; Rodriguez-Mendez, A.J.; Mejia, C.; Ferriz-Martinez, R.A.; et al. Phaseolus acutifolius Lectin Fractions Exhibit Apoptotic Effects on Colon Cancer: Preclinical Studies Using Dimethilhydrazine or Azoxi-Methane as Cancer Induction Agents. Molecules 2017, 22, 1670. [Google Scholar] [CrossRef] [Green Version]
- García-Gasca, T.; Salazar-Olivo, L.A.; Mendiola-Olaya, E.; Blanco-Labra, A. The effects of a protease inhibitor fraction from tepary bean (Phaseolus acutifolius) on in vitro cell proliferation and cell adhesion of transformed cells. Toxicol. In Vitro 2002, 16, 229–233. [Google Scholar] [CrossRef]
- Jiang, Q.-L.; Zhang, S.; Tian, M.; Zhang, S.-Y.; Xie, T.; Chen, D.-Y.; Chen, Y.-J.; He, J.; Liu, J.; Ouyang, L.; et al. Plant lectins, from ancient sugar-binding proteins to emerging anti-cancer drugs in apoptosis and autophagy. Cell Prolif. 2015, 48, 17–28. [Google Scholar] [CrossRef]
- Chen, D.L.; Engle, J.T.; Griffin, E.A.; Miller, J.P.; Chu, W.; Zhou, D.; Mach, R.H. Imaging Caspase-3 Activation as a Marker of Apoptosis-Targeted Treatment Response in Cancer. Mol. Imaging Biol. 2015, 17, 384–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, A.C.; Ferreira, R.; Freitas, R. Plant Lectins: Bioactivities and Bioapplications. In Studies in Natural Products Chemistry; Atta-ur-Rahman, Ed.; Elsevier B.V.: Amsterdam, The Netherlands, 2018; Volume 58, pp. 1–42. ISBN 9780444640567. [Google Scholar]
- Chen, J.; Liu, B.; Ji, N.; Zhou, J.; Bian, H.J.; Li, C.Y.; Chen, F.; Bao, J.K. A novel sialic acid-specific lectin from Phaseolus coccineus seeds with potent antineoplastic and antifungal activities. Phytomedicine 2009, 16, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Lv, H.; Wang, Y.; Liu, Y.H.; Li, C.Y.; Meng, L.; Chen, F.; Bao, J.K. Clematis montana lectin, a novel mannose-binding lectin from traditional Chinese medicine with antiviral and apoptosis-inducing activities. Peptides 2009, 30, 1805–1815. [Google Scholar] [CrossRef] [PubMed]
- Asaduzzaman, A.K.M.; Hasan, I.; Chakrabortty, A.; Zaman, S.; Islam, S.S.; Ahmed, F.R.S.; Kabir, K.M.A.; Nurujjaman, M.; Uddin, M.B.; Alam, M.T.; et al. Moringa oleifera seed lectin inhibits Ehrlich ascites carcinoma cell growth by inducing apoptosis through the regulation of Bak and NF-κB gene expression. Int. J. Biol. Macromol. 2018, 107, 1936–1944. [Google Scholar] [CrossRef]
- Hegde, P.; Rajakumar, S.B.; Swamy, B.M.; Inamdar, S.R. A mitogenic lectin from Rhizoctonia bataticola arrests growth, inhibits metastasis, and induces apoptosis in human colon epithelial cancer cells. J. Cell. Biochem. 2018, 119, 5632–5645. [Google Scholar] [CrossRef]
- Islam, F.; Gopalan, V.; Lam, A.K.Y.; Kabir, S.R. Pea lectin inhibits cell growth by inducing apoptosis in SW480 and SW48 cell lines. Int. J. Biol. Macromol. 2018, 117, 1050–1057. [Google Scholar] [CrossRef] [Green Version]
- Lee-Yong, K.; Kim, W.; Lyu, S.; Park, W.-B.; Yoon, J.-W.; Jun, H.-S. Mechanisms involved in Korean mistletoe lectin-induced apoptosis of cancer cells. World J. Gastroenterol. 2007, 13, 2811–2818. [Google Scholar]
- Ingaramo, M.C.; Sánchez, J.A.; Dekanty, A. Regulation and function of p53: A perspective from Drosophila studies. Mech. Dev. 2018, 154, 82–90. [Google Scholar] [CrossRef]
- Fogarty, C.E.; Bergmann, A. Killers creating new life: Caspases drive apoptosis-induced proliferation in tissue repair and disease. Cell Death Differ. 2017, 24, 1390–1400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meulmeester, E.; Jochemsen, A. p53: A Guide to Apoptosis. Curr. Cancer Drug Targets 2008, 8, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Hollstein, M.; Xu, Y. Ser46 phosphorylation regulates p53-dependent apoptosis and replicative senescence. Cell Cycle 2006, 5, 2812–2819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Orazi, G.; Cecchinelli, B.; Bruno, T.; Manni, I.; Higashimoto, Y.; Saito, S.; Gostissa, M.; Coen, S.; Marchetti, A.; Del Sal, G.; et al. Homeodomain-interacting protein kinase-2 phosphorylates p53 at Ser 46 and mediates apoptosis. Nat. Cell Biol. 2002, 4, 11–19. [Google Scholar] [CrossRef]
- Smeenk, L.; van Heeringen, S.J.; Koeppel, M.; Gilbert, B.; Janssen-Megens, E.; Stunnenberg, H.G.; Lohrum, M. Role of p53 Serine 46 in p53 Target Gene Regulation. PLoS ONE 2011, 6, e17574. [Google Scholar] [CrossRef] [Green Version]
- Liebl, M.C.; Hofmann, T.G. Cell Fate Regulation upon DNA Damage: p53 Serine 46 Kinases Pave the Cell Death Road. BioEssays 2019, 41, 1900127. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.-J.; Jang, Y.-J. Selective apoptotic effect of Zelkova serrata twig extract on mouth epidermoid carcinoma through p53 activation. Int. J. Oral Sci. 2012, 4, 78–84. [Google Scholar] [CrossRef]
- Sun, B.; Ross, S.M.; Joseph Trask, O.; Carmichael, P.L.; Dent, M.; White, A.; Andersen, M.E.; Clewell, R.A. Assessing dose-dependent differences in DNA-damage, p53 response and genotoxicity for quercetin and curcumin. Toxicol. Vitr. 2013, 27, 1877–1887. [Google Scholar] [CrossRef]
- De Lange, J.; Ly, L.V.; Lodder, K.; Verlaan-De Vries, M.; Teunisse, A.F.A.S.; Jager, M.J.; Jochemsen, A.G. Synergistic growth inhibition based on small-molecule p53 activation as treatment for intraocular melanoma. Oncogene 2012, 31, 1105–1116. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Dilly, A.K.; Choudry, H.A.; Bartlett, D.L.; Kwon, Y.T.; Lee, Y.J. Hypoxia promotes synergy between mitomycin c and bortezomib through a coordinated process of Bcl-xL phosphorylation and mitochondrial translocation of p53. Mol. Cancer Res. 2015, 13, 1533–1543. [Google Scholar] [CrossRef] [Green Version]
- Yan, Q.; Li, Y.; Jiang, Z.; Sun, Y.; Zhu, L.; Ding, Z. Antiproliferation and apoptosis of human tumor cell lines by a lectin (AMML) of Astragalus mongholicus. Phytomedicine 2009, 16, 586–593. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, N.; Koyama, Y.; Katsuno, Y.; Hayakawa, S.; Mita, T.; Ohta, T.; Kaji, K.; Isemura, M. Apoptosis induction associated with cell cycle dysregulation by rice bran agglutinin. J. Biochem. 2001, 130, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Bhutia, S.K.; Panda, P.K.; Sinha, N.; Praharaj, P.P.; Bhol, C.S.; Panigrahi, D.P.; Mahapatra, K.K.; Saha, S.; Patra, S.; Mishra, S.R.; et al. Plant lectins in cancer therapeutics: Targeting apoptosis and autophagy-dependent cell death. Pharmacol. Res. 2019, 144, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Wu, J.M.; Li, J.; Liu, J.J.; Li, W.W.; Li, C.Y.; Xu, H.L.; Bao, J.K. Polygonatum cyrtonema lectin induces murine fibrosarcoma L929 cell apoptosis and autophagy via blocking Ras-Raf and PI3K-Akt signaling pathways. Biochimie 2010, 92, 1934–1938. [Google Scholar] [CrossRef] [PubMed]
- Hostanska, K.; Vuong, V.; Rocha, S.; Soengas, M.S.; Glanzmann, C.; Saller, R.; Bodis, S.; Pruschy, M. Recombinant mistletoe lectin induces p53-independent apoptosis in tumour cells and cooperates with ionising radiation. Br. J. Cancer 2003, 88, 1785–1792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takumi, S.; Komatsu, M.; Furukawa, T.; Ikeda, R.; Sumizawa, T.; Akenaga, H.; Maeda, Y.; Aoyama, K.; Arizono, K.; Ando, S.; et al. P53 plays an important role in cell fate determination after exposure to Microcystin-LR. Environ. Health Perspect. 2010, 118, 1292–1298. [Google Scholar] [CrossRef] [Green Version]
- Bundy, H.F.; Westberg, N.J.; Dummel, B.M.; Becker, C.A. Purification and Partial Characterization of Prorennin. Biochemistry 1964, 3, 923–926. [Google Scholar] [CrossRef]
- Torres Arteaga, I.; Castro Guillen, J.L.; Mendiola-Olaya, E.; Garcia-Gasca, T.; Angeles Zaragoza, M.; Garcia Santoyo, V.; Torres Castillo, J.A.; Aguirre, C.; Phinney, B.; Blanco-Labra, A. Characterization of Two Non-Fetuin-Binding Lectins from Tepary Bean (Phaseolus acutifolius) Seeds with Differential Cytotoxicity on Colon Cancer Cells. J. Glycobiol. 2016, 5, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Jaffé, W. Hemagglutinins (Lectins). In Toxic Constituents of Plant Foodstuffs; Academic Press: New York, NY, USA, 1980; pp. 73–102. [Google Scholar]
- Turner, R.H.; Liener, I.E. The use of glutaraldehyde-treated erythrocytes for assaying the agglutinating activity of lectins. Anal. Biochem. 1975, 68, 651–653. [Google Scholar] [CrossRef]
- Sanger Institute Genome Research Limited Cell Lines Project v90. Available online: http://cancer.sanger.ac.uk/cosmic/ (accessed on 2 March 2018).
- American Type Culture Collection American Type Culture Collection (ATCC): The Global Bioresource Center. Available online: http://www.atcc.org/ (accessed on 3 May 2018).
- University of California. UCSC Genome Browser Home. Available online: https://genome.ucsc.edu/ (accessed on 10 August 2018).
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3 Input. Available online: http://primer3.ut.ee/ (accessed on 8 October 2018).
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Characteristics |
|---|---|
| HT-29 (ATCC® HTB-38™) | Colorectal adenocarcinoma cells. Positive for c-myc, K-ras, H-ras, N-ras, Myb, sis and fos oncogenes expression, N-myc oncogene expression was not detected. p53 protein is overproduced with a G-A mutation in codon 273 resulting in an Arg-His substitution. Express human adrenergic alpha2A receptor, urokinase receptor (u-PAR) and moderate expression of vitamin D receptor |
| RKO (ATCC® CRL-2577™) | Colon carcinoma cells. Wild-type p53 with high expression, positive for urokinase receptor (u-PAR), but lack endogenous human thyroid receptor nuclear receptor (h-TRbeta1). |
| SW-480 (ATCC® CCL-228™) | Colorectal adenocarcinoma cells. Positive for the expression of c-myc, K-ras, H-ras, N-ras, myb, sis and fos oncogenes, negative for N-myc oncogene expression and for Matrilysin (a metalloproteinase associated with tumor invasiveness), express high levels of p53 protein with a G-A mutation in codon 273 of the p53 gene resulting in an Arg-His substitution and a C-T mutation in codon 309 resulting in a Pro-Ser substitution. Positive for epidermal growth factor receptor (EGFR). |
| Gen Target | Forward | Reverse |
|---|---|---|
| B-CTNN | TGGACTTGATATTGGTGCCCA | GCCACCCATCTCATGTTCCA |
| DCC | CCCCTGAAGTGTCTGAGGAG | AGCTGCTTCATGAGTCCTTCC |
| PI3K | TGGAGCTGACCCAAATCCAT | TTCAAAGGCAGGGTTACTCC |
| GSK3 | CTCCATCCAACCGTCTCTCA | GGTAGGTGTGGCATCGGTC |
| CAS9 | CAAGAGTGGCTCCTGGTACG | TCCCTTTCACCGAAACAGCA |
| BAD | TTCGGAGGATGAGTGACGAG | CAAGTTCCGATCCCACCAGG |
| PTEN | GCCGTCAAATCCAGAGGCTA | GGATCAGAGTCAGTGGTGTCA |
| AKT | CCTTCAAGCCCCAGGTCAC | CGCTCGCTGTCCACACAC |
| TP53 | CCAACAACACCAGCTCCTCT | TCAGGAAGTAACACCATCGTAAG |
| BCL-2 | GACTGAGTACCTGAACCGGC | GGCCAAACTGAGCAGAGTCT |
| KRAS | TGTGATTTGCCTTCTAGAACAGT | ACACCCTGTCTTGTCTTTGCT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno-Celis, U.; López-Martínez, F.J.; Cervantes-Jiménez, R.; Ferríz-Martínez, R.A.; Blanco-Labra, A.; García-Gasca, T. Tepary Bean (Phaseolus acutifolius) Lectins Induce Apoptosis and Cell Arrest in G0/G1 by P53(Ser46) Phosphorylation in Colon Cancer Cells. Molecules 2020, 25, 1021. https://doi.org/10.3390/molecules25051021
Moreno-Celis U, López-Martínez FJ, Cervantes-Jiménez R, Ferríz-Martínez RA, Blanco-Labra A, García-Gasca T. Tepary Bean (Phaseolus acutifolius) Lectins Induce Apoptosis and Cell Arrest in G0/G1 by P53(Ser46) Phosphorylation in Colon Cancer Cells. Molecules. 2020; 25(5):1021. https://doi.org/10.3390/molecules25051021
Chicago/Turabian StyleMoreno-Celis, Ulisses, F. Josué López-Martínez, Ricardo Cervantes-Jiménez, Roberto Augusto Ferríz-Martínez, Alejandro Blanco-Labra, and Teresa García-Gasca. 2020. "Tepary Bean (Phaseolus acutifolius) Lectins Induce Apoptosis and Cell Arrest in G0/G1 by P53(Ser46) Phosphorylation in Colon Cancer Cells" Molecules 25, no. 5: 1021. https://doi.org/10.3390/molecules25051021
APA StyleMoreno-Celis, U., López-Martínez, F. J., Cervantes-Jiménez, R., Ferríz-Martínez, R. A., Blanco-Labra, A., & García-Gasca, T. (2020). Tepary Bean (Phaseolus acutifolius) Lectins Induce Apoptosis and Cell Arrest in G0/G1 by P53(Ser46) Phosphorylation in Colon Cancer Cells. Molecules, 25(5), 1021. https://doi.org/10.3390/molecules25051021