
The Metabolic Changes of Artesunate and Ursolic Acid on Syrian Golden Hamsters Fed with the High-Fat Diet



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
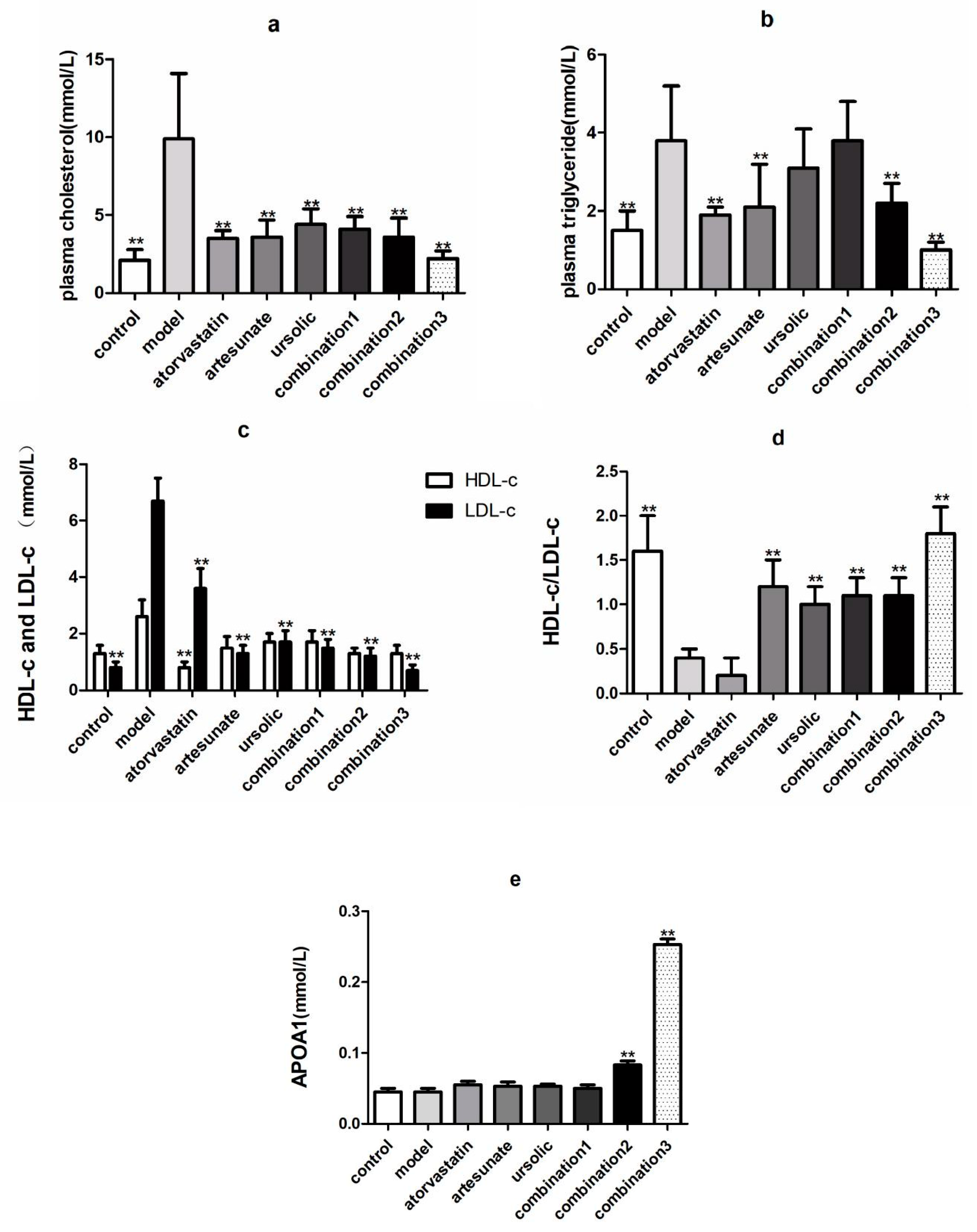
2.1. Artesunate/Ursolic Acid Combination Therapy Has Better Lipid-Lowering Effect than Atorvastatin
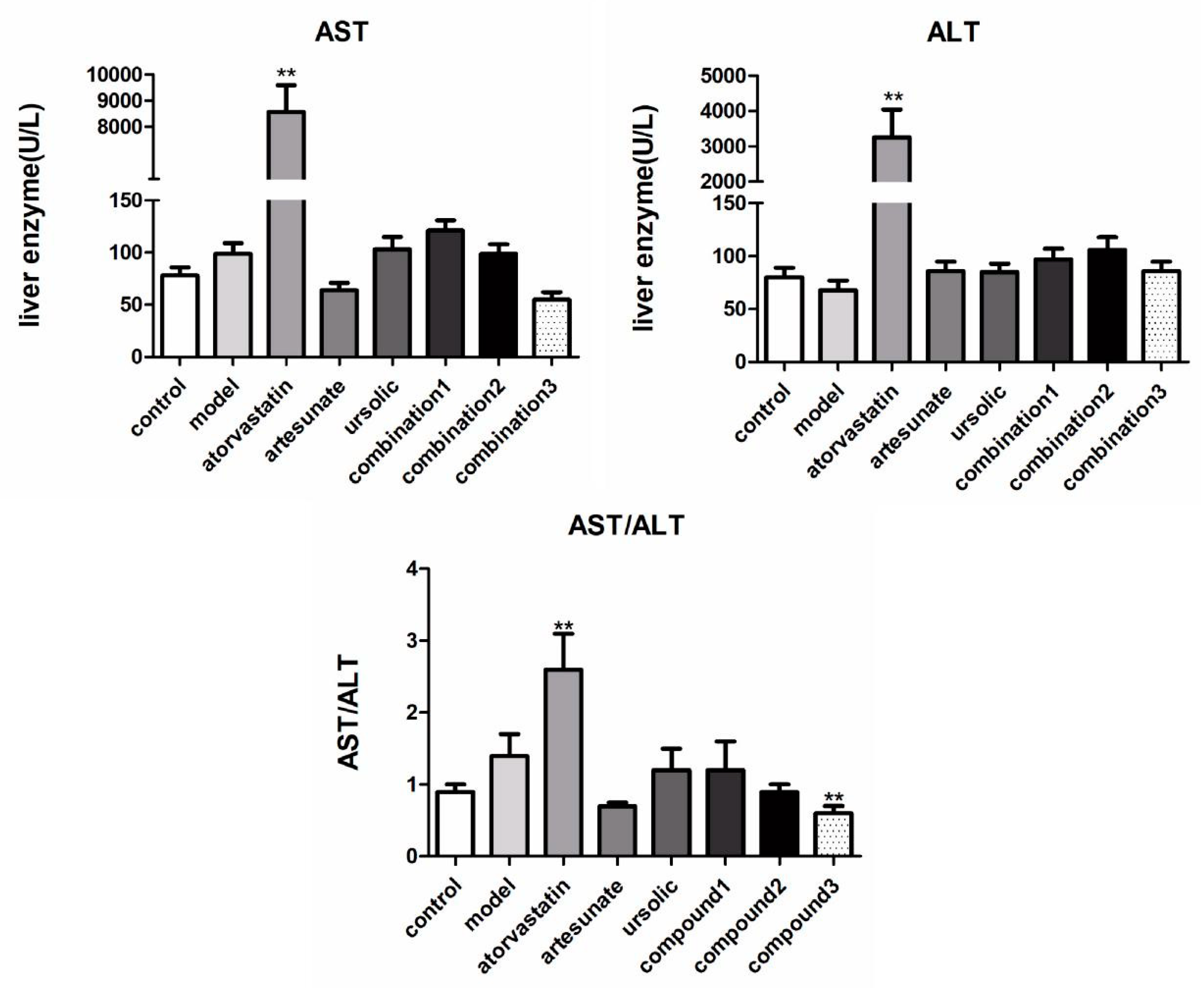
2.2. Artesunate/Ursolic Acid Combination Therapy Has Lower Liver Toxicity Compared with Atorvastatin
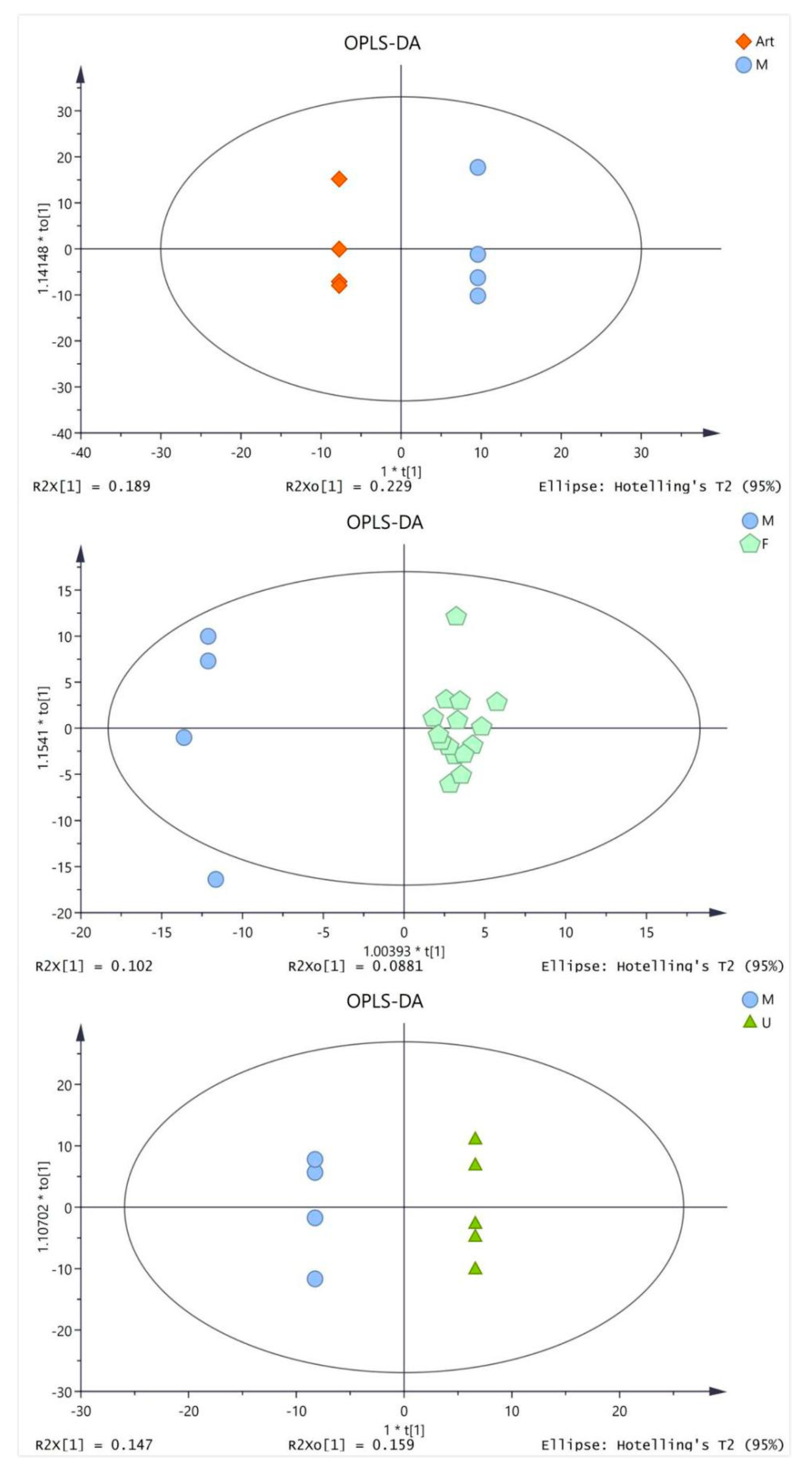
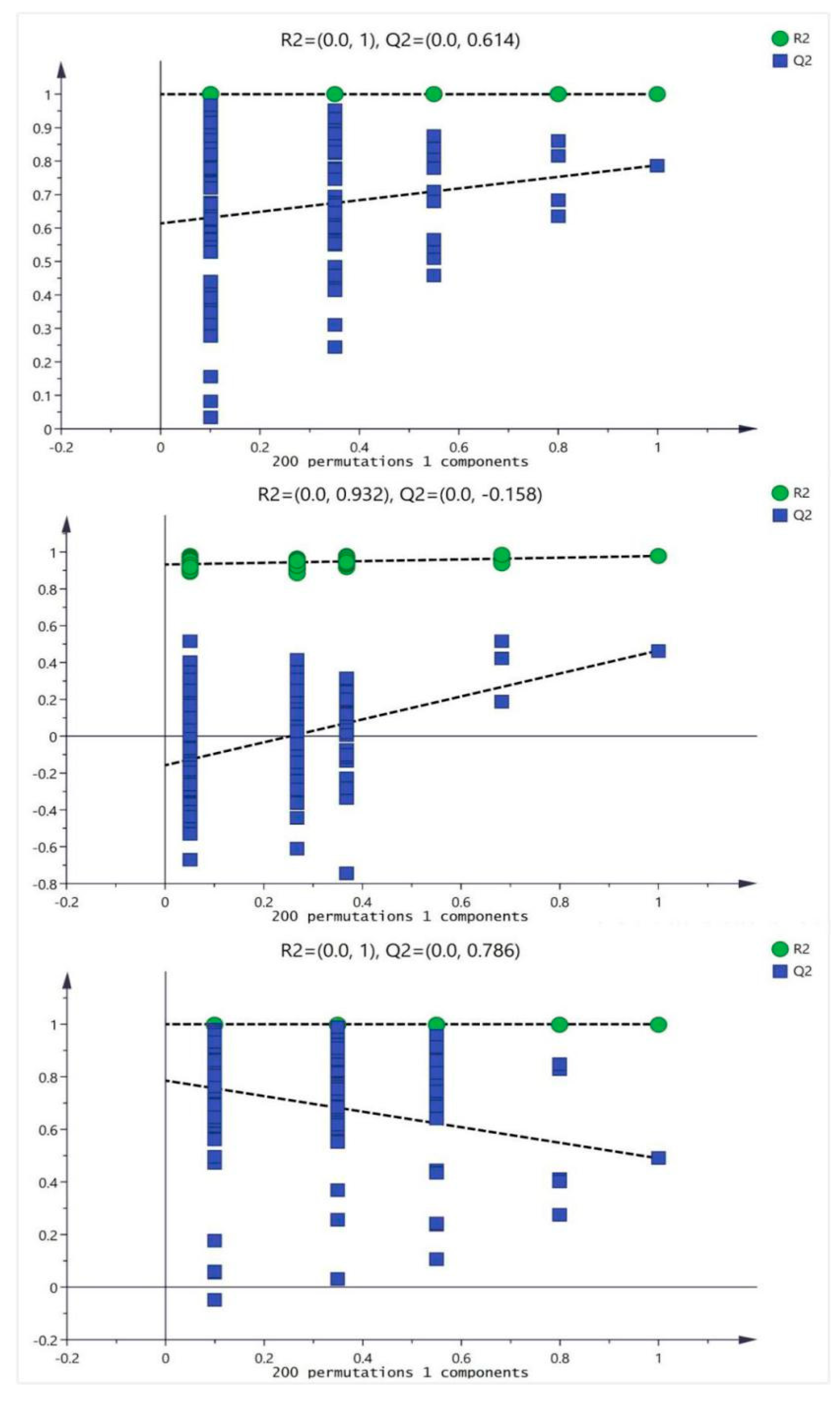
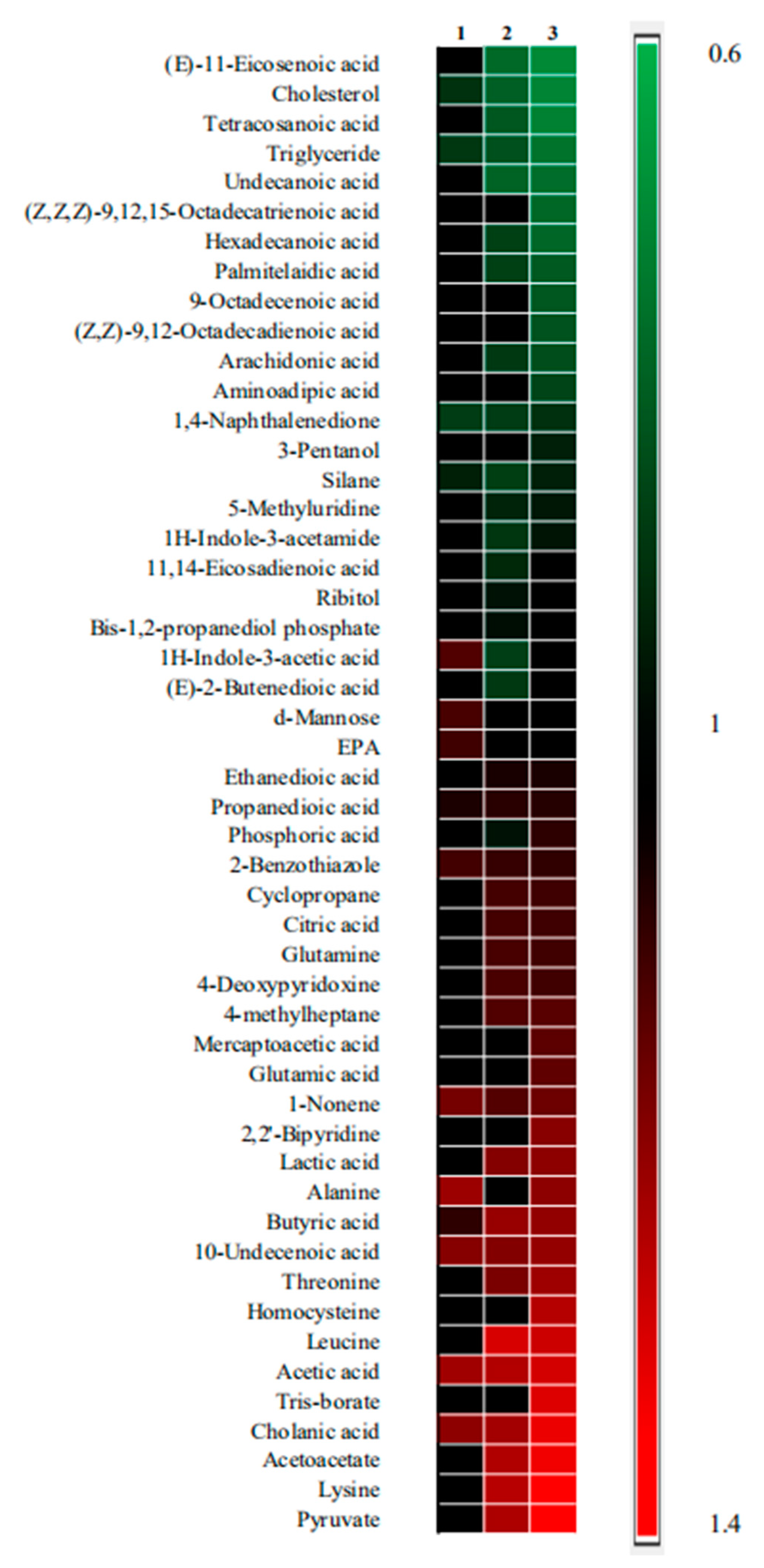
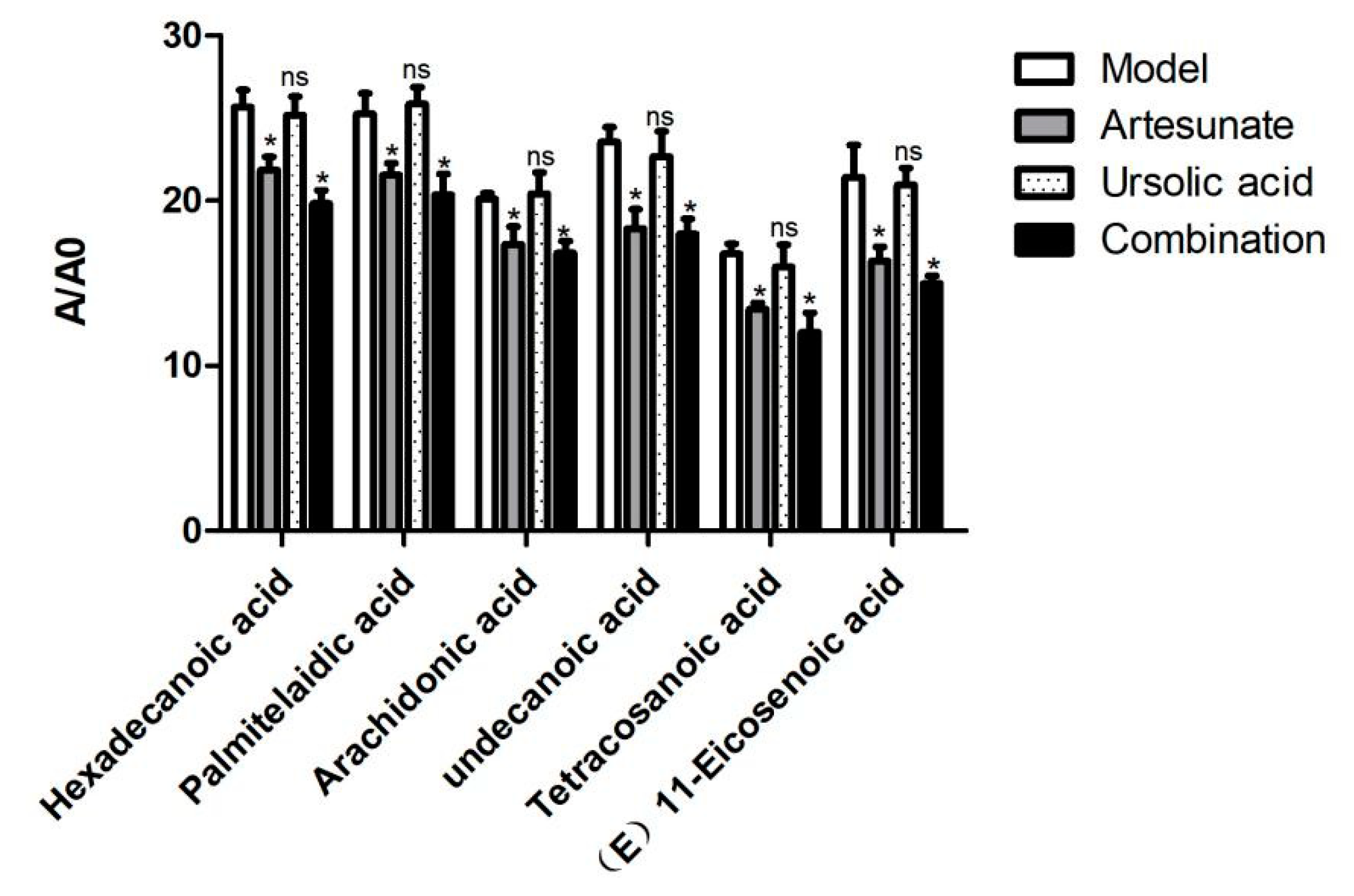
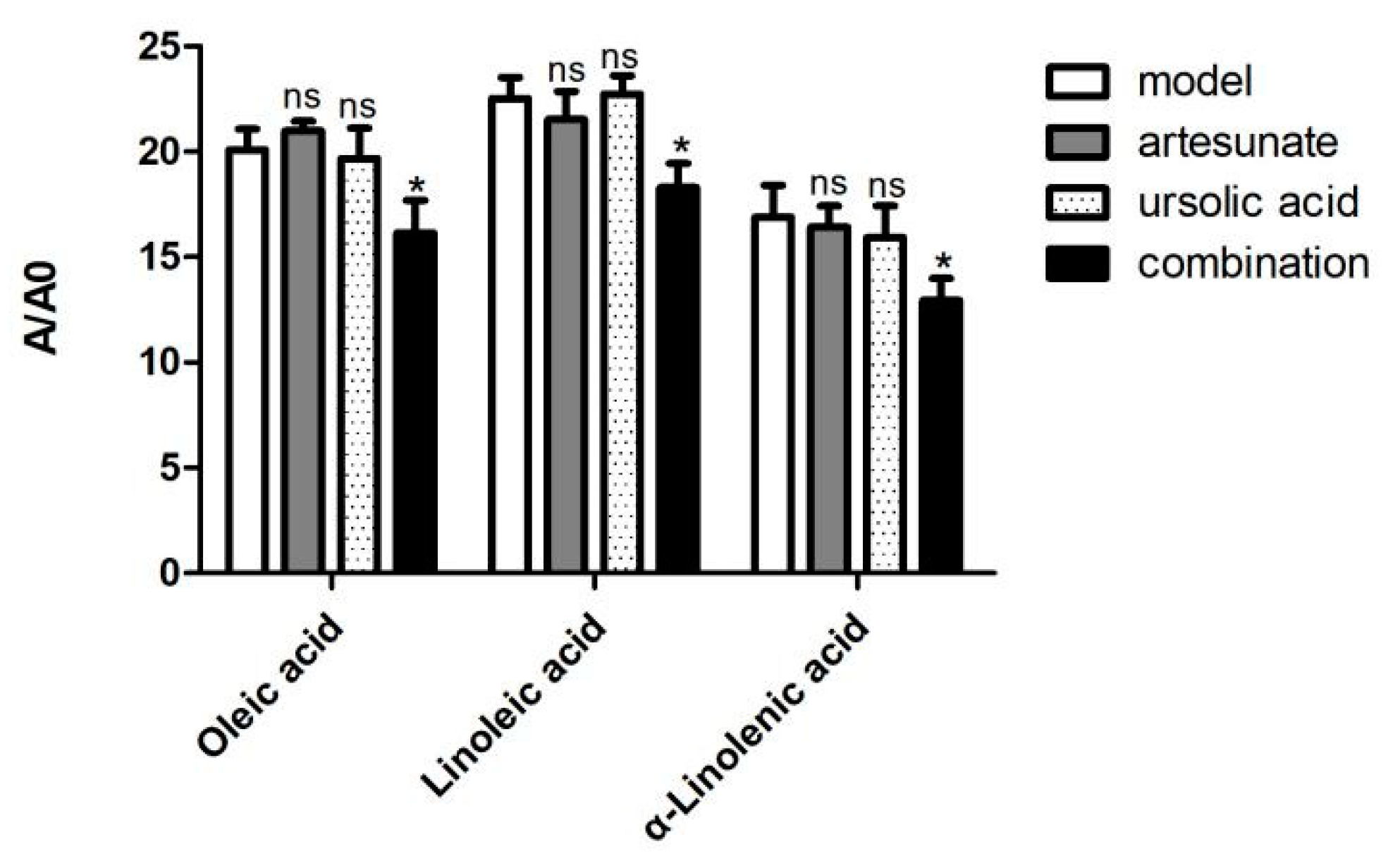
2.3. Metabolic Difference between Drug Treatment and Model Group
3. Discussion
4. Materials and Methods
4.1. Chemical and Reagent
4.2. The Feeding Conditions and Treatments for Syrian Golden Hamsters
4.3. Preparation of Serum Samples
4.4. Preparation of Samples and Analysis by GC/TOF MS
4.5. Identification of Metabolites
4.6. Data Processing and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Marchesini, G.; Brizi, M.; Bianchi, G. Nonalcoholic fatty liver disease: A feature of the metabolic syndrome. Diabetes 2001, 50, 1844–1850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drechsler, M.; Megens, R.T.; van Zandvoort, M. Hyperlipidemia-triggered neutrophilia promotes early atherosclerosis. Circulation 2010, 122, 1837–1845. [Google Scholar] [CrossRef] [PubMed]
- DeFaria, Y.D.; Freeman, M.W.; Meigs, J.B.; Grant, R.W. Risk factors for coronary artery disease in patients with elevated high-density lipoprotein cholesterol. Am. J. Cardiol. 2007, 99, 1–4. [Google Scholar] [CrossRef]
- Taskinen, M.R.; Borén, J. New insights into the pathophysiology of dyslipidemia in type 2 diabetes. Atherosclerosis 2015, 239, 483–495. [Google Scholar] [CrossRef]
- Rideout, T.C.; Ramprasath, V.; Griffin, J.D. Phytosterols protect against diet-induced hypertriglyceridemia in Syrian golden hamsters. Lipids Health Dis. 2014, 13, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, H.; Wang, Y. Berberine alleviates hepatic lipid accumulation by increasing ABCA1 through the protein kinase C and pathway. Biochem. Biophys. Res. Commun. 2018, 498, 473–480. [Google Scholar] [CrossRef]
- Ejaz, A.; Wu, D.; Kwan, P.; Meydani, M. Curcumin inhibits adipogenesis in 3T3-L1 adipocytes and angiogenesis and obesity in C57/BL mice. J. Nutr. 2009, 139, 919–925. [Google Scholar] [CrossRef]
- Denis, M.B.; Tsuyuoka, R.; Poravuth, Y. Surveillance of the efficacy of artesunate and mefloquine combination for the treatment of uncomplicated falciparum malaria in Cambodia. Trop Med. Int. Health 2006, 11, 1360–1366. [Google Scholar] [CrossRef]
- Kyaw, M.P.; Nyunt, M.H.; Chit, K. Reduced susceptibility of Plasmodium falciparum to artesunate in southern Myanmar. PLoS ONE 2013, 8, e57689. [Google Scholar] [CrossRef] [Green Version]
- Pryce, J.; Hine, P. Pyronaridine-artesunate for treating uncomplicated Plasmodium falciparum malaria. Cochrane Database Syst. Rev. 2019, 1, CD006404. [Google Scholar] [CrossRef]
- Wang, L.; Liu, L.; Wang, J.; Chen, Y. Inhibitory Effect of Artesunate on Growth and Apoptosis of Gastric Cancer Cells. Arch. Med. Res. 2017, 48, 623–663. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Liu, L.; Wen, L.; Yu, T.; Wei, F. Artesunate promotes G2/M cell cycle arrest in MCF7 breast cancer cells through ATM activation. Breast Cancer 2018, 25, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, Y.; Murakami, A.; Ohigashi, H. Ursolic acid: An anti- and pro-inflammatory triterpenoid. Mol. Nutr. Food Res. 2008, 52, 26–42. [Google Scholar] [CrossRef] [PubMed]
- Yin, R.; Li, T.; Tian, J.X.; Xi, P.; Liu, R.H. Ursolic acid, a potential anticancer compound for breast cancer therapy. Crit. Rev. Food Sci. Nutr. 2018, 58, 568–574. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Liu, X.; Lee, E.S.; Sun, J.; Feng, Z.; Zhao, L.; Zhao, C. Synthesis of novel oleanolic acid and ursolic acid in C-28 position derivatives as potential anticancer agents. Arch. Pharm. Res. 2017, 40, 458–468. [Google Scholar] [CrossRef] [PubMed]
- Somova, L.O.; Nadar, A.; Rammanan, P.; Shode, F.O. Cardiovascular, antihyperlipidemic and antioxidant effects of oleanolic and ursolic acids in experimental hypertension. Phytomedicine 2003, 10, 115–121. [Google Scholar] [CrossRef]
- Wang, Y.L.; Wang, Z.J.; Shen, H.L.; Yin, M.; Tang, K.X. Effects of artesunate and ursolic acid on hyperlipidemia and its complications in rabbit. Eur. J. Pharm. Sci. 2013, 50, 366–371. [Google Scholar] [CrossRef]
- Wenk, M.R. The emerging field of lipidomics. Nat. Rev. Drug Discov. 2005, 4, 594–610. [Google Scholar] [CrossRef]
- Watson, A.D. Thematic review series: Systems biology approaches to metabolic and cardiovascular disorders. Lipidomics: A global approach to lipid analysis in biological systems. J. Lipid Res. 2006, 47, 2101–2111. [Google Scholar] [CrossRef] [Green Version]
- Hu, T.; Zhang, J.L. Mass-spectrometry-based lipidomics. J. Sep. Sci. 2018, 41, 351–372. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, G.J.; A., J.Y.; Wu, D.; Zhu, L.L.; Ma, B.; Du, Y. Application of GC/MS-based metabonomic profiling in studying the lipid-regulating effects of Ginkgo biloba extract on diet-induced hyperlipidemia in rats. Acta Pharmacol. Sin. 2009, 30, 1674–1687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spady, D.K.; Dietschy, J.M. Sterol synthesis in vivo in 18 tissues of the squirrel, monkey, guinea pig, hamster and rat. J. Lipid Res. 1983, 24, 303–315. [Google Scholar] [CrossRef]
- Khallou, J.; Riottot, M.; Parquet, M.; Verneau, C.; Lutton, C. Biodynamics of cholesterol and bile acids in the lithiasic hamster. Br. J. Nutr. 1991, 66, 479–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goulinet, S.; Chapman, M.J. Plasma lipoproteins in the golden Syrian hamster (Mesocricetus auratus): Heterogeneity of apoBand apoA-I-containing particles. J. Lipid Res. 1993, 34, 943–959. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jang, D.S.; Kim, H.; Kim, J.S. Anti-lipase and lipolytic activities of ursolic acid isolated from the roots of Actinidia arguta. Arch. Pharm. Res. 2009, 32, 983–987. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, P.; Cariou, B.; Lien, F. Role of bile acids and bile acid receptors in metabolic regulation. Physiol. Rev. 2009, 89, 147–191. [Google Scholar] [CrossRef] [Green Version]
- Cortez, M.Y.; Torgan, C.E.; Brozinick, J.T. Effects of pyruvate and dihydroxyacetone consumption on the growth and metabolic state of obese Zucker rats. Am. J. Clin. Nutr. 1991, 53, 847–853. [Google Scholar] [CrossRef]
- Guerranti, R.; Pagani, R.; Neri, S.; Errico, S.V.; Leoncini, R.; Marinello, E. Inhibition and regulation of rat liver L-threonine dehydrogenase by different fatty acids and their derivatives. Biochim. Biophys. Acta 2001, 1568, 45–52. [Google Scholar] [CrossRef]
- Mong, M.C.; Chao, C.Y.; Yin, M.C. Histidine and carnosine alleviated hepatic steatosis in mice consumed high saturated fat diet. Eur. J. Pharmacol. 2011, 653, 82–88. [Google Scholar] [CrossRef]
- Sun, X.; Zemel, M.B. Leucine and calcium regulate fat metabolism and energy partitioning in murine adi-pocytes and muscle cells. Lipids 2007, 42, 297–305. [Google Scholar] [CrossRef]
- Zhao, L.M.; Wu, H.B.; Qin, M.F. Metabolic signatures of kideny Yang deficiency syndrome and protective effects of two herbal extracts in rats using GC/TOFMS. Evid-Based Compl. Alt. 2013, 540957. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Y.; Cai, G.; Su, M. Serum metabolite profiling of human colorectal cancer using GC-TOFMS and UPLCQTOFMS. J. Proteome Res. 2009, 8, 4844–4850. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, P.; Gullberg, J.; Nordström, A.; Kusano, M.; Kowalczyk, M.; Sjöström, M.; Moritz, T. A strategy for identifying diffrences in large series of metabolomic samples analyzed by GC/MS. Anal. Chem. 2004, 76, 1738–1745. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, P.; Johansson, A.I.; Gullberg, J. High-throughput data analysis for detecting and identifying diffrences between samples in GC/MS-based metabolomic analyses. Anal. Chem. 2005, 77, 5635–5642. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Su, M.; Qiu, Y.; Chen, M.; Liu, Y.; Zhao, A.; Jia, W. Metabolic profiing using combined GC-MS and LC-MS provides a systems understanding of aristolochic acid-induced nephrotoxicity in rat. FEBS Lett. 2007, 581, 707–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Sample Availability: Samples of the compounds are available from the authors. |








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pu, S.; Liu, Y.; Liang, S.; Liu, P.; Qian, H.; Wu, Q.; Wang, Y. The Metabolic Changes of Artesunate and Ursolic Acid on Syrian Golden Hamsters Fed with the High-Fat Diet. Molecules 2020, 25, 1392. https://doi.org/10.3390/molecules25061392
Pu S, Liu Y, Liang S, Liu P, Qian H, Wu Q, Wang Y. The Metabolic Changes of Artesunate and Ursolic Acid on Syrian Golden Hamsters Fed with the High-Fat Diet. Molecules. 2020; 25(6):1392. https://doi.org/10.3390/molecules25061392
Chicago/Turabian StylePu, Shichen, Yumin Liu, Shan Liang, Pin Liu, Hongmei Qian, Qian Wu, and Yuliang Wang. 2020. "The Metabolic Changes of Artesunate and Ursolic Acid on Syrian Golden Hamsters Fed with the High-Fat Diet" Molecules 25, no. 6: 1392. https://doi.org/10.3390/molecules25061392
APA StylePu, S., Liu, Y., Liang, S., Liu, P., Qian, H., Wu, Q., & Wang, Y. (2020). The Metabolic Changes of Artesunate and Ursolic Acid on Syrian Golden Hamsters Fed with the High-Fat Diet. Molecules, 25(6), 1392. https://doi.org/10.3390/molecules25061392