
A Unique Gene-Silencing Approach, Using an Intelligent RNA Expression Device (iRed), Results in Minimal Immune Stimulation When Given by Local Intrapleural Injection in Malignant Pleural Mesothelioma


 , , ,
, , ,
Abstract
:1. Introduction
2. Results
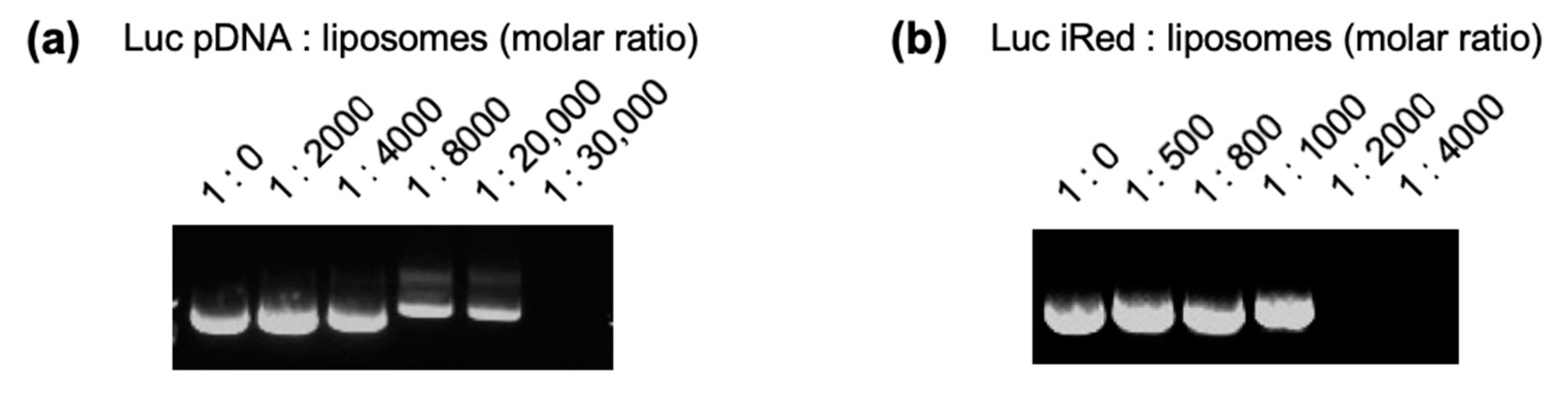
2.1. Preparation of DNAs/Liposome-Complexes
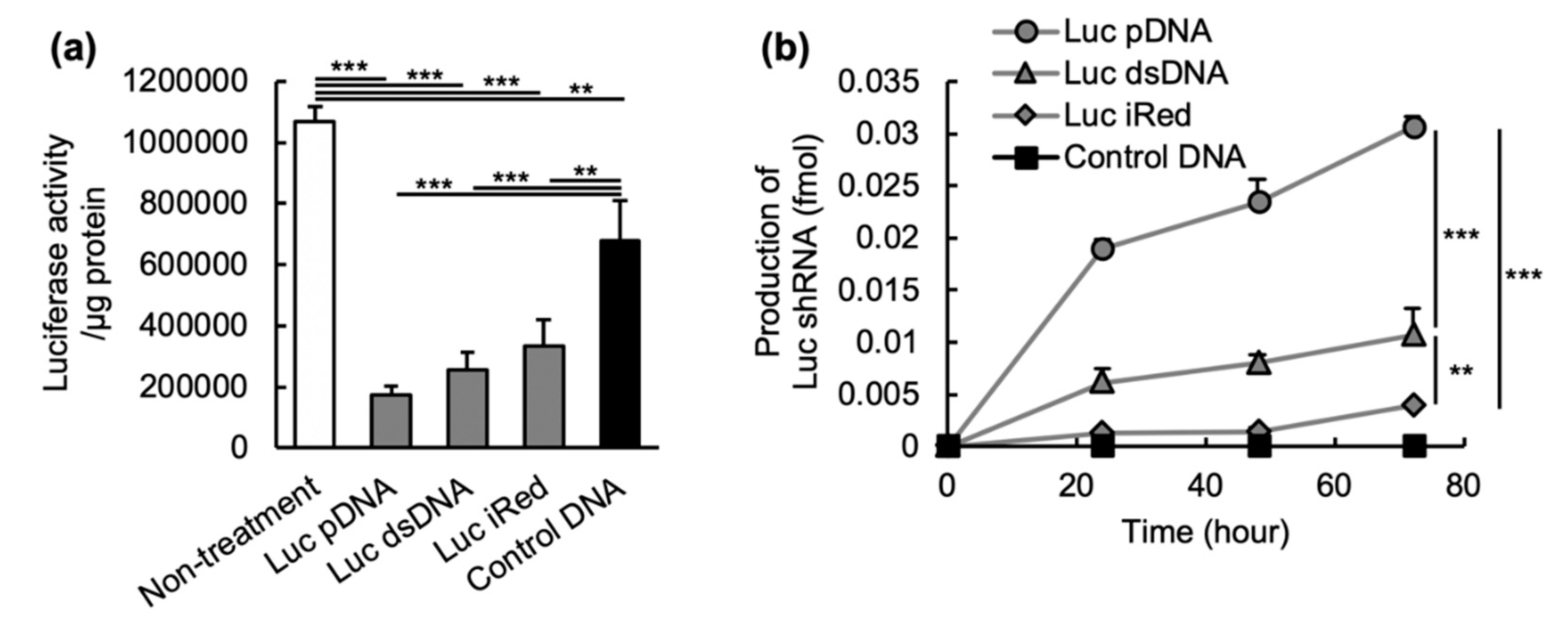
2.2. In Vitro Gene Silencing Effect of shRNA-Expressing DNAs
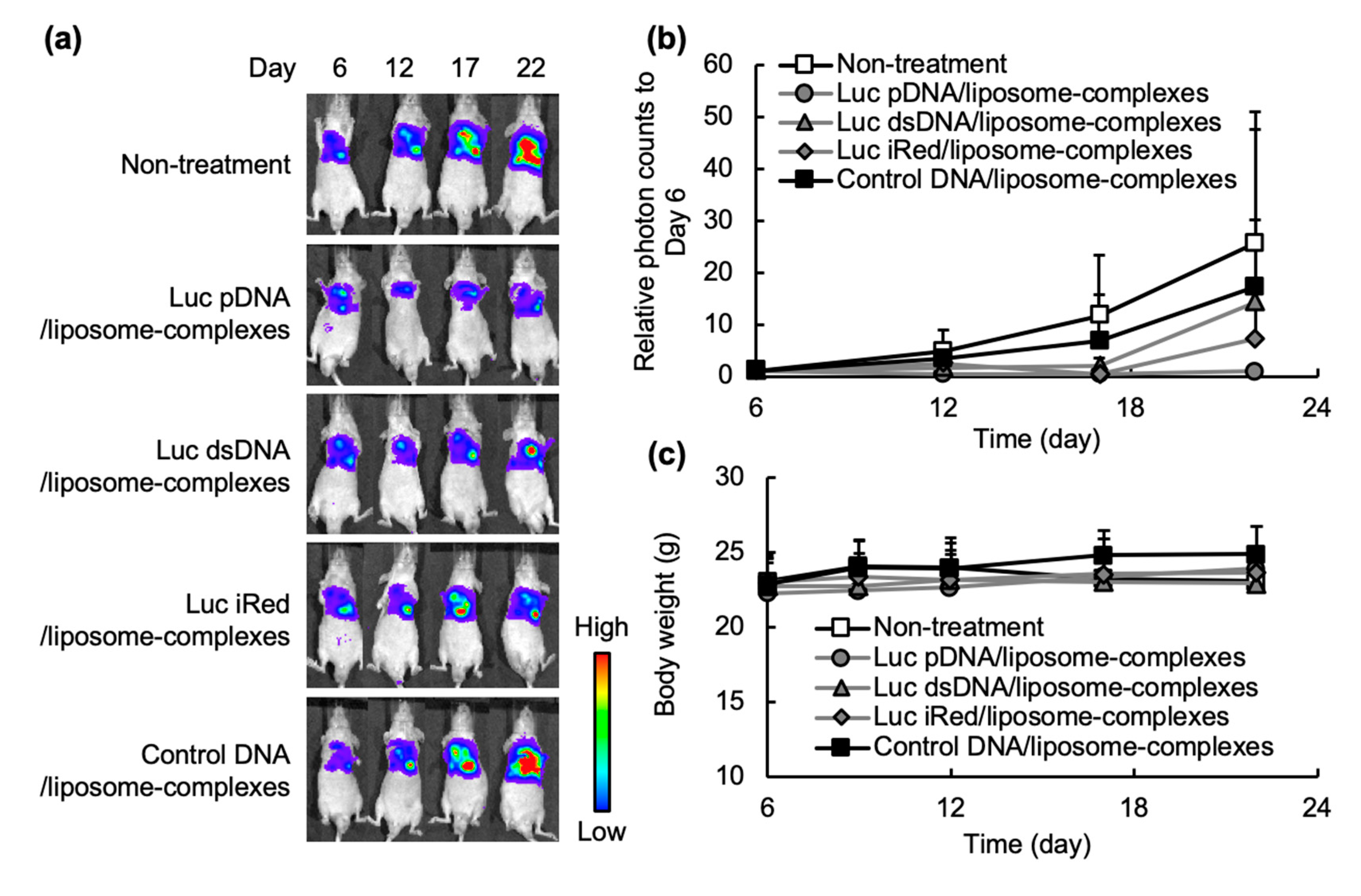
2.3. In Vivo Gene Silencing Effect of shRNA-Expressing DNAs Formulated in DNA/liposome-Complexes
2.4. Induction of Innate Immune Reactions Following Intrapleural Injections of shRNA-Expressing DNAs Formulated in DNA/Liposome-Complexes
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Construction of shRNA-Expression pDNA
4.3. Preparation of Intelligent shRNA-Expression Device (iRed) and shRNA-Expressing Natural Device (Double Strand DNA; dsDNA)
4.4. Preparation of DNAs/Liposome-Complexes
4.5. Electrophoresis
4.6. Cell Culture
4.7. Preparation of the Orthotopic Mesothelioma Mouse Model
4.8. In Vivo Luciferase Reporter Assay
4.9. Determination of IL-6 Production
4.10. In Vitro Luciferase Reporter Assay
4.11. Quantification of Luciferase shRNA by Real Time RT-PCR
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mao, C.P.; Lin, Y.Y.; Hung, C.F.; Wu, T.C. Immunological research using RNA interference technology. Immunology 2007, 121, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Kuwano, K.; Ochiya, T. Development of small RNA delivery systems for lung cancer therapy. Int. J. Mol. Sci. 2015, 16, 5254–5270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, D.D.; Vorhies, J.S.; Senzer, N.; Nemunaitis, J. siRNA vs. shRNA: similarities and differences. Adv. Drug Deliv. Rev. 2009, 61, 746–759. [Google Scholar] [CrossRef] [PubMed]
- Peacock, H.; Kannan, A.; Beal, P.A.; Burrows, C.J. Chemical modification of siRNA bases to probe and enhance RNA interference. J. Org. Chem. 2011, 76, 7295–7300. [Google Scholar] [CrossRef] [Green Version]
- Lundin, K.E.; Gissberg, O.; Smith, C.I. Oligonucleotide Therapies: The Past and the Present. Hum. Gene Ther. 2015, 26, 475–485. [Google Scholar] [CrossRef] [Green Version]
- Shirota, H.; Tross, D.; Klinman, D.M. CpG Oligonucleotides as Cancer Vaccine Adjuvants. Vaccines 2015, 3, 390–407. [Google Scholar] [CrossRef] [Green Version]
- Chiu, Y.L.; Rana, T.M. siRNA function in RNAi: A chemical modification analysis. RNA 2003, 9, 1034–1048. [Google Scholar] [CrossRef] [Green Version]
- Valenzuela, R.A.; Onizuka, K.; Ball-Jones, A.A.; Hu, T.; Suter, S.R.; Beal, P.A. Guide Strand 3’-End Modifications Regulate siRNA Specificity. Chembiochem 2016, 17, 2340–2345. [Google Scholar] [CrossRef] [Green Version]
- Strapps, W.R.; Pickering, V.; Muiru, G.T.; Rice, J.; Orsborn, S.; Polisky, B.A.; Sachs, A.; Bartz, S.R. The siRNA sequence and guide strand overhangs are determinants of in vivo duration of silencing. Nucleic Acids Res. 2010, 38, 4788–4797. [Google Scholar] [CrossRef] [Green Version]
- Tarashima, N.; Ando, H.; Kojima, T.; Kinjo, N.; Hashimoto, Y.; Furukawa, K.; Ishida, T.; Minakawa, N. Gene Silencing Using 4’-thioDNA as an Artificial Template to Synthesize Short Hairpin RNA Without Inducing a Detectable Innate Immune Response. Mol. Ther. Nucleic Acids 2016, 5, e274. [Google Scholar] [CrossRef]
- Inoue, N.; Shionoya, A.; Minakawa, N.; Kawakami, A.; Ogawa, N.; Matsuda, A. Amplification of 4‘-ThioDNA in the Presence of 4‘-Thio-dTTP and 4‘-Thio-dCTP, and 4‘-ThioDNA-Directed Transcription in Vitro and in Mammalian Cells. J. Am. Chem. Soc. 2007, 129, 15424–15425. [Google Scholar] [CrossRef]
- Hoshika, S.; Minakawa, N.; Matsuda, A. Synthesis and physical and physiological properties of 4’-thioRNA: Application to post-modification of RNA aptamer toward NF-kappaB. Nucleic Acids Res. 2004, 32, 3815–3825. [Google Scholar] [CrossRef]
- Abu Lila, A.S.; Kato, C.; Fukushima, M.; Huang, C.L.; Wada, H.; Ishida, T. Downregulation of thymidylate synthase by RNAi molecules enhances the antitumor effect of pemetrexed in an orthotopic malignant mesothelioma xenograft mouse model. Int. J. Oncol. 2016, 48, 1399–1407. [Google Scholar] [CrossRef] [Green Version]
- Ando, H.; Abu Lila, A.S.; Fukushima, M.; Matsuoka, R.; Shimizu, T.; Okuhira, K.; Ishima, Y.; Huang, C.L.; Wada, H.; Ishida, T. A simplified method for manufacturing RNAi therapeutics for local administration. Int. J. Pharm. 2019, 564, 256–262. [Google Scholar] [CrossRef]
- Kojima, T.; Furukawa, K.; Maruyama, H.; Inoue, N.; Tarashima, N.; Matsuda, A.; Minakawa, N. PCR amplification of 4’-thioDNA using 2’-deoxy-4’-thionucleoside 5’-triphosphates. ACS Synth. Biol. 2013, 2, 529–536. [Google Scholar] [CrossRef]
- Ishida, T.; Maeda, R.; Ichihara, M.; Irimura, K.; Kiwada, H. Accelerated clearance of PEGylated liposomes in rats after repeated injections. J. Control. Release 2003, 88, 35–42. [Google Scholar] [CrossRef]
- Bartlett, G.R. Colorimetric assay methods for free and phosphorylated glyceric acids. J. Biol. Chem. 1959, 234, 469–471. [Google Scholar]
- Zhang, X.X.; McIntosh, T.J.; Grinstaff, M.W. Functional lipids and lipoplexes for improved gene delivery. Biochimie 2012, 94, 42–58. [Google Scholar] [CrossRef] [Green Version]
- Shen, W.G. RNA interference and its current application in mammals. Chin. Med. J. (Engl.) 2004, 117, 1084–1091. [Google Scholar]
- Ambesajir, A.; Kaushik, A.; Kaushik, J.J.; Petros, S.T. RNA interference: A futuristic tool and its therapeutic applications. Saudi. J. Biol. Sci. 2012, 19, 395–403. [Google Scholar] [CrossRef] [Green Version]
- Juliano, R.; Alam, M.R.; Dixit, V.; Kang, H. Mechanisms and strategies for effective delivery of antisense and siRNA oligonucleotides. Nucleic Acids Res. 2008, 36, 4158–4171. [Google Scholar]
- Kanasty, R.L.; Whitehead, K.A.; Vegas, A.J.; Anderson, D.G. Action and reaction: the biological response to siRNA and its delivery vehicles. Mol. Ther. 2012, 20, 513–524. [Google Scholar]
- Mizrahy, S.; Hazan-Halevy, I.; Dammes, N.; Landesman-Milo, D.; Peer, D. Current Progress in Non-viral RNAi-Based Delivery Strategies to Lymphocytes. Mol. Ther. 2017, 25, 1491–1500. [Google Scholar]
- Oliveira, S.; Storm, G.; Schiffelers, R.M. Targeted delivery of siRNA. J Biomed Biotechnol. 2006, 2006, 63675. [Google Scholar]
- Xin, Y.; Huang, M.; Guo, W.W.; Huang, Q.; Zhang, L.Z.; Jiang, G. Nano-based delivery of RNAi in cancer therapy. Mol. Cancer 2017, 16, 134. [Google Scholar]
- Walther, W.; Stein, U. Viral Vectors for Gene Transfer. Drugs 2000, 60, 249–271. [Google Scholar]
- Zuhorn, I.S.; Engberts, J.B.; Hoekstra, D. Gene delivery by cationic lipid vectors: Overcoming cellular barriers. Eur. Biophys. J. 2007, 36, 349–362. [Google Scholar]
- Ando, H.; Kobayashi, S.; Abu Lila, A.S.; Eldin, N.E.; Kato, C.; Shimizu, T.; Ukawa, M.; Kawazoe, K.; Ishida, T. Advanced therapeutic approach for the treatment of malignant pleural mesothelioma via the intrapleural administration of liposomal pemetrexed. J. Control. Release 2015, 220, 29–36. [Google Scholar]
- Hutvagner, G.; Simard, M.J. Argonaute proteins: Key players in RNA silencing. Nat. Rev. Mol. Cell Biol. 2008, 9, 22–32. [Google Scholar]
- Marques, J.T.; Williams, B.R. Activation of the mammalian immune system by siRNAs. Nat. Biotechnol. 2005, 23, 1399–1405. [Google Scholar]
- Meng, Z.; Lu, M. RNA Interference-Induced Innate Immunity, Off-Target Effect, or Immune Adjuvant? Front. Immunol. 2017, 8, 331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robbins, M.; Judge, A.; MacLachlan, I. siRNA and innate immunity. Oligonucleotides 2009, 19, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Lacy, P.; Stow, J.L. Cytokine release from innate immune cells: Association with diverse membrane trafficking pathways. Blood 2011, 118, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Striz, I.; Brabcova, E.; Kolesar, L.; Sekerkova, A. Cytokine networking of innate immunity cells: a potential target of therapy. Clin. Sci. (Lond.) 2014, 126, 593–612. [Google Scholar]
Sample Availability: Samples of the all compounds that we used in this study are available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Complexes | Particle Size (nm) | Zeta Potential (mV) |
|---|---|---|
| Luc pDNA/liposome-complexes (molar ratio = 1:30,000) | 197.1 ± 0.4 | +23.0 ± 2.8 |
| Luc dsDNA/liposome-complexes (molar ratio = 1:2000) | 160.9 ± 1.2 | +38.0 ± 0.7 |
| Luc iRed/liposome-complexes (molar ratio = 1:2000) | 159.1 ± 4.1 | +35.5 ± 1.5 |
| Cationic liposomes (DOPE:cholesterol:DOTAP = 3:3:4 molar ratio) | 108.1 ± 3.4 | +44.0 ± 0.6 |
| Complexes | Inhibition Rate (%/mol) | Significance |
|---|---|---|
| Luc pDNA/liposome-complexes | 40.4 ± 1.54 | *** p < 0.001 vs. Luc dsDNA/liposome-complexes *** p < 0.001 vs. Luc iRed/liposome-complexes |
| Luc dsDNA/liposome-complexes | 1.0 ± 1.42 | |
| Luc iRed/liposome-complexes | 1.65 ± 0.56 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ando, H.; Saito-Tarashima, N.; Lila, A.S.A.; Kinjo, N.; Shimizu, T.; Ishima, Y.; Minakawa, N.; Ishida, T. A Unique Gene-Silencing Approach, Using an Intelligent RNA Expression Device (iRed), Results in Minimal Immune Stimulation When Given by Local Intrapleural Injection in Malignant Pleural Mesothelioma. Molecules 2020, 25, 1725. https://doi.org/10.3390/molecules25071725
Ando H, Saito-Tarashima N, Lila ASA, Kinjo N, Shimizu T, Ishima Y, Minakawa N, Ishida T. A Unique Gene-Silencing Approach, Using an Intelligent RNA Expression Device (iRed), Results in Minimal Immune Stimulation When Given by Local Intrapleural Injection in Malignant Pleural Mesothelioma. Molecules. 2020; 25(7):1725. https://doi.org/10.3390/molecules25071725
Chicago/Turabian StyleAndo, Hidenori, Noriko Saito-Tarashima, Amr S. Abu Lila, Nozomi Kinjo, Taro Shimizu, Yu Ishima, Noriaki Minakawa, and Tatsuhiro Ishida. 2020. "A Unique Gene-Silencing Approach, Using an Intelligent RNA Expression Device (iRed), Results in Minimal Immune Stimulation When Given by Local Intrapleural Injection in Malignant Pleural Mesothelioma" Molecules 25, no. 7: 1725. https://doi.org/10.3390/molecules25071725
APA StyleAndo, H., Saito-Tarashima, N., Lila, A. S. A., Kinjo, N., Shimizu, T., Ishima, Y., Minakawa, N., & Ishida, T. (2020). A Unique Gene-Silencing Approach, Using an Intelligent RNA Expression Device (iRed), Results in Minimal Immune Stimulation When Given by Local Intrapleural Injection in Malignant Pleural Mesothelioma. Molecules, 25(7), 1725. https://doi.org/10.3390/molecules25071725