
Analysis of Antioxidant Activity and Flavonoids Metabolites in Peel and Flesh of Red-Fleshed Apple Varieties

Abstract
:1. Introduction
2. Results and Discussion
2.1. Phenotypic Characters
2.2. Total Anthocyanins and Total Phenols Content
2.3. Antioxidant Activity of Anthocyanins Extract from Peels and Flesh
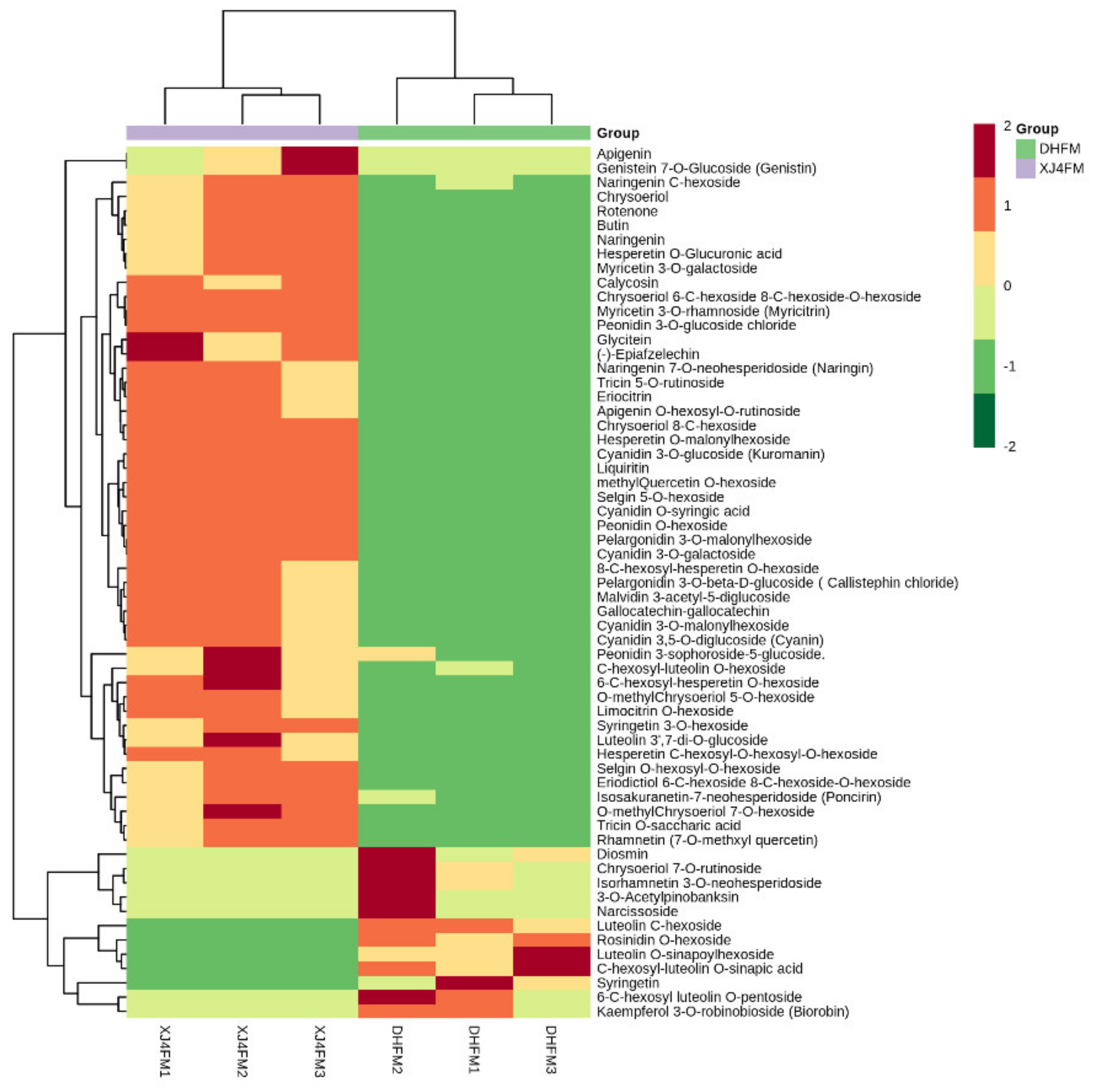
2.4. Distribution and Cluster Analysis of Significantly Differential Metabolites in ‘DH’ vs. ‘XJ4’
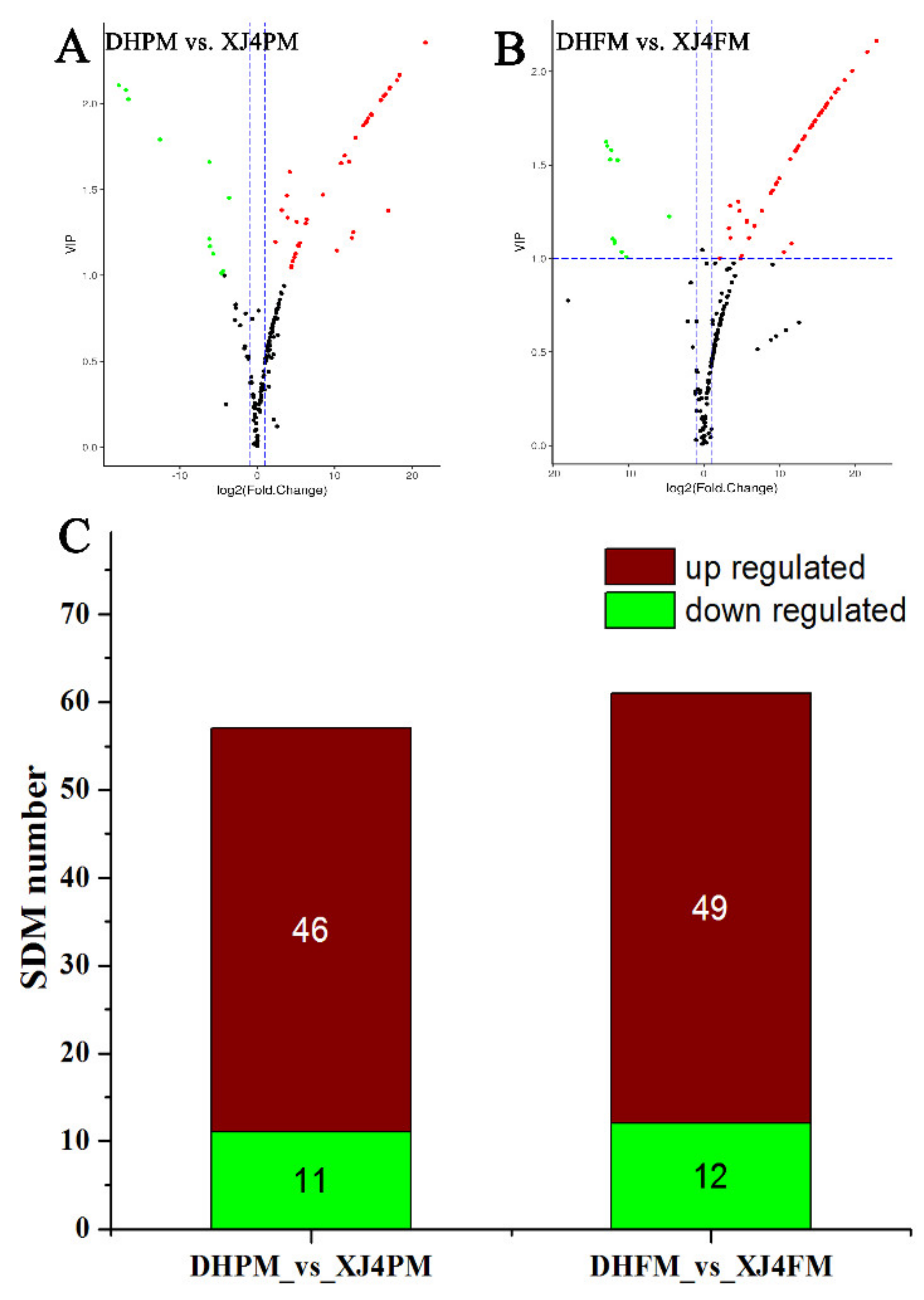
2.5. Significantly Differential Metabolites Assay in DHPM vs. XJ4PM and DHFM vs. XJ4FM
2.5.1. Anthocyanins
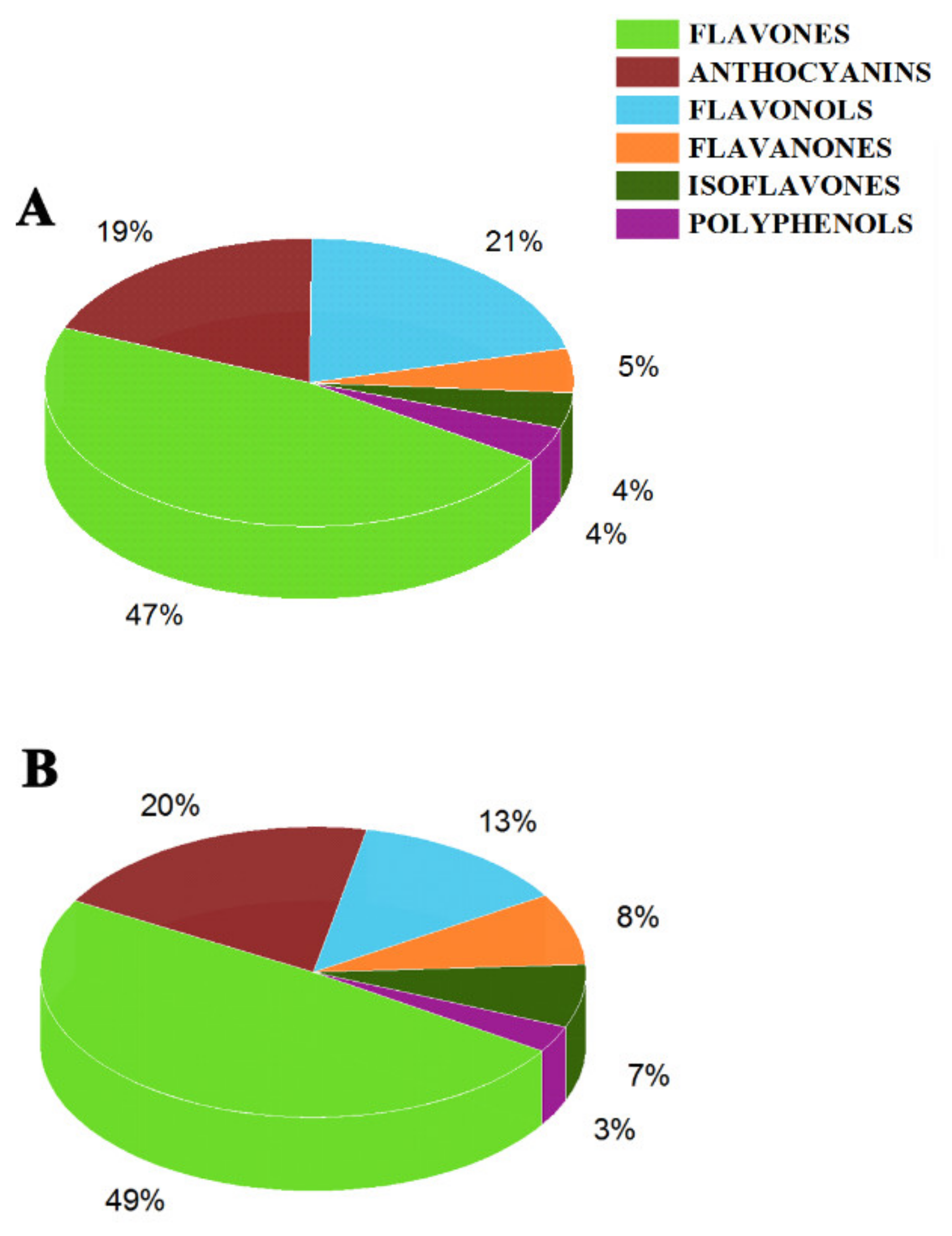
2.5.2. Flavones
2.5.3. Flavonols, Flavanones, Isoflavones and Polyphenols
2.6. KEGG Enrichment Analysis of Significantly Differential Metabolites
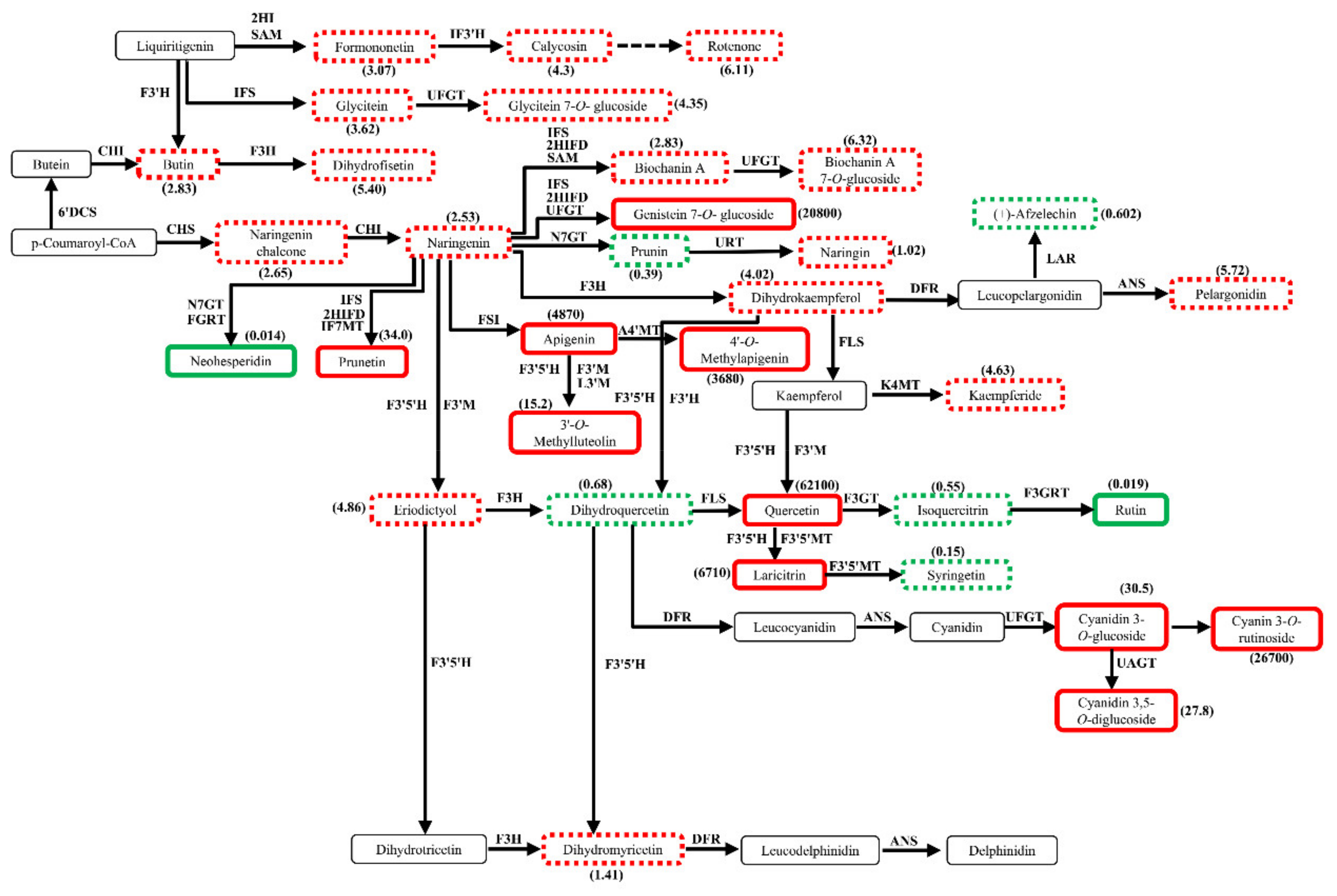
2.7. Profiles of Differential Metabolites in Flavonoid Biosynthetic Pathways
3. Conclusions
4. Materials and Methods
4.1. Plant Materials and Sampling
4.2. Extraction and Determination of Total Anthocyanins Content
4.3. Determination of Total Phenols Content
4.4. Scavenging Capacity of DPPH Radical
4.5. Scavenging Capacity of ·OH
4.6. Scavenging Rate of O2·-
4.7. Sample Preparation and Extraction for Metabolites Study
4.8. Metabolite Separation, Identification and Quantification
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 2HID | 2-Hydroxyisoflavanone dehydratase |
| 2HIFD | 2-Hydroxyisoflavanone dehydratase |
| 6’DCS | 6′-Deoxychalcone synthase |
| ANR | Anthocyanidin reductase |
| ANS | Anthocyanidin synthase |
| CHI | Chalcone isomerase |
| CHS | Chalcone synthase |
| DFR | Dihydroflavonol 4-reductase |
| DH | Daihong |
| DHFM | Flesh of ‘DH’ at mature stage |
| DHPM | Peel of ‘DH’ at mature stage |
| ESI | Electrospray ionization |
| F3GRT | Flavonol-3-O-glucoside L-rhamnosyltransferase |
| F3GT | Flavonol 3-O-glucosyltransferase |
| F3H | Flavanone 3-hydroxylase |
| F3’5’H | Flavanoid 3’,5′-hydroxylase |
| F3’5’MT | Flavonoid 3’,5′-methyltransferase |
| F3’H | Flavoniod 3′-hydroxylose |
| F3’H | Flavonoid 3′-hydroxylase |
| F3’M | Flavonoid 3′-monooxygenase |
| FLS | Flavonol synthase |
| FSI | Flavone synthase I |
| HPLC | High performance liquid chromatography |
| HX1 | Hongxun No.1 |
| IF3’H | Isoflavone 3′-hydroxylase |
| IFS | Isoflavonoid synthase |
| K4MT | Kaempferol 4-O-methyltransferase |
| L3’M | Luteolin 3′-O-methyltransferase |
| LAR | Leucoanthocyanin reductase |
| LCR | Leucocyanidin reductase |
| MRM | Multiple-reaction monitoring |
| N7GT | Naringenin 7-O-glucosyltransferase |
| QN-5 | Qingnong No.5 |
| SAM | 2,7,4′-trihydroxyisoflavanone 4′-O-methyltransferase |
| UAGT | UDP-glucose-anthocyanin 5-O-glucosyltransferase |
| UCRGT | UDP-glucose-cyanidin-d-rhamnosyl-glucosyltransferase |
| UFGT | UDP glucose-flavonoid 3-O-glcosyl-transferase |
| URT | UDP-rhamnosetransferase |
| VIP | Variable importance in projection |
References
- Kaisoon, O.; Siriamornpun, S.; Weerapreeyakul, N.; Meeso, N. Phenolic compounds and antioxidant activities of edible flowers from Thailand. J. Funct. Foods 2011, 3, 88–99. [Google Scholar] [CrossRef]
- Sun, Y.; Li, M.; Mitra, S.; Rizwan, H.M.; Debnath, B.; Lu, X.; Jian, H.; Qiu, D. Comparative Phytochemical Profiles and Antioxidant Enzyme Activity Analyses of the Southern Highbush Blueberry (Vaccinium corymbosum) at Different Developmental Stages. Molecules 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, R.H. Dietary Bioactive Compounds and Their Health Implications. J. Food Sci. 2013, 78, A18–A25. [Google Scholar] [CrossRef] [PubMed]
- Jaakola, L. New insights into the regulation of anthocyanin biosynthesis in fruits. Trends Plant Sci. 2013, 18, 477–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grotewold, E. The genetics and biochemistry of floral pigments. Annu. Rev. Plant Boil. 2006, 57, 761–780. [Google Scholar] [CrossRef] [PubMed]
- Shimada, Y.; Nakano, S.; Ohbayashi, M. Expression of chimeric P450 genes encoding flavonoid-3′,5′-hydroxylase in transgenic tobacco and petunia plants. FEBS Lett. 1999, 461, 241–245. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Li, X.; Zhu, C.; Sun, J.; Tian, L.; Chen, W.; Bai, W.; Tian, L. The target cells of anthocyanins in metabolic syndrome. Crit. Rev. Food Sci. Nutr. 2018, 59, 921–946. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, J.; Wang, L.; Wang, S.; Li, X.; Zhang, S. Advances in research of red-skinned pear coloration and anthocyanin synthesis. J. Fruit Sci. 2012, 29, 668–675. [Google Scholar]
- Yi, L.; Chen, C.-Y.; Jin, X.; Mi, M.; Yu, B.; Chang, H.; Ling, W.-H.; Zhang, T. Structural requirements of anthocyanins in relation to inhibition of endothelial injury induced by oxidized low-density lipoprotein and correlation with radical scavenging activity. FEBS Lett. 2009, 584, 583–590. [Google Scholar] [CrossRef] [Green Version]
- Xiang, Y.; Zhao, R.X.; Lai, F.N.; Sun, X.; Sun, X.H.; Dai, H.Y.; Zhang, Y.G. Analysis of flavonoid components and antioxidant activity in peel of red-fleshed apple. Plant Physiol. J. 2016, 52, 1353–1360. [Google Scholar]
- Dang, Y.; Liu, S. Study on extraction technology and antioxidant activity of anthocyanin from purple cabbage. North. Hortic. 2015, 1, 128–136. [Google Scholar]
- Yasuko, N.; Takao, K.; Akitane, M.; Lester, P. Antioxidant activities of pomegranate fruit extract and its anthocyanidins: Delphinidin, cyanidin, and pelargonidin. J. Agric. Food Chem. 2002, 50, 166–171. [Google Scholar]
- Li, H.; Deng, Z.; Zhu, H.; Hu, C.; Liu, R.; Young, J.C.; Tsao, R. Highly pigmented vegetables: Anthocyanin compositions and their role in antioxidant activities. Food Res. Int. 2012, 46, 250–259. [Google Scholar] [CrossRef]
- Zhang, Y.G.; Zhu, J.; Dai, H.Y. Morphological characteristics and pollination compatibility of a new red flesh apple, Hongxun No.1. Research on Crops 2013, 14, 199–204. [Google Scholar]
- Zhang, X.; Huo, H.Q.; Sun, X.H.; Zhu, J.; Dai, H.Y.; Zhang, Y.G. Nanocrystallization of Anthocyanin Extract from Red-Fleshed Apple ‘QN-5′ Improved Its Antioxidant Effect through Enhanced Stability and Activity under Stressful Conditions. Molecules 2019. [Google Scholar] [CrossRef] [Green Version]
- Li, C.X.; Zhao, X.H.; Zuo, W.F.; Zhang, T.L.; Zhang, Z.Y.; Chen, X.S. Phytochemical profiles, antioxidant, and antiproliferative activities of four red-fleshed apple varieties in China. J. Food Sci. 2020, 85, 718–726. [Google Scholar] [CrossRef]
- Xiang, Y.; Lai, F.; He, G.; Li, Y.; Yang, L.; Shen, W.; Huo, H.; Zhu, J.; Dai, H.; Zhang, Y. Alleviation of Rosup-induced oxidative stress in porcine granulosa cells by anthocyanins from red-fleshed apples. PLoS ONE 2017. [Google Scholar] [CrossRef]
- Katiyo, W.; Yang, R.; Zhao, W. Phenolic composition and antioxidant activity of Chinese red-fleshed apples (Malus pumila Niedzwetzkyana (Dieck) and effect of different pasteurization treatments on the cloudy juice. Int. Food Res. J. 2018, 25, 2185–2194. [Google Scholar]
- Wang, Y.; Tang, H. Metabolomics: A revolution in progress. Prog. Biochem. Biophys. 2006, 33, 401–417. [Google Scholar]
- Weckwerth, W. Metabolomics: An integral technique in systems biology. Bioanalysis 2010, 2, 829–836. [Google Scholar] [CrossRef]
- Zhang, A.; Sun, H.; Wang, P.; Han, Y.; Wang, X.-J. Recent and potential developments of biofluid analyses in metabolomics. J. Proteom. 2012, 75, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Liu, W.; Zhang, T.; Jiang, S.; Xu, H.; Wang, Y.; Zhang, Z.; Wang, C.; Chen, X. Transcriptomic Analysis of Red-Fleshed Apples Reveals the Novel Role of MdWRKY11 in Flavonoid and Anthocyanin Biosynthesis. J. Agric. Food Chem. 2018, 66, 7076–7086. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Cui, Y.; Vainstein, A.; Chen, S.; Ma, H. Regulation of Fig (Ficus carica L.) Fruit Color: Metabolomic and Transcriptomic Analyses of the Flavonoid Biosynthetic Pathway. Front. Plant Sci. 2017. [Google Scholar] [CrossRef] [Green Version]
- Jo, Y.-H.; Park, H.-C.; Choi, S.; Kim, S.; Bao, C.; Kim, H.W.; Choi, H.-K.; Lee, H.J.; Auh, J.-H. Metabolomic Analysis Reveals Cyanidins in Black Raspberry as Candidates for Suppression of Lipopolysaccharide-Induced Inflammation in Murine Macrophages. J. Agric. Food Chem. 2015, 63, 5449–5458. [Google Scholar] [CrossRef]
- Wang, L.; Nägele, T.; Doerfler, H.; Fragner, L.; Chaturvedi, P.; Nukarinen, E.; Bellaire, A.; Huber, W.; Weiszmann, J.; Engelmeier, D.; et al. System level analysis of cacao seed ripening reveals a sequential interplay of primary and secondary metabolism leading to polyphenol accumulation and preparation of stress resistance. Plant J. 2016, 87, 318–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takos, A.; Jaffé, F.W.; Jacob, S.R.; Bogs, J.; Robinson, S.; Walker, A. Light-Induced Expression of a MYB Gene Regulates Anthocyanin Biosynthesis in Red Apples1. Plant Physiol. 2006, 142, 1216–1232. [Google Scholar] [CrossRef] [Green Version]
- Espley, R.; Brendolise, C.; Chagné, D.; Kutty-Amma, S.; Green, S.; Volz, R.; Putterill, J.; Schouten, H.J.; Gardiner, S.E.; Hellens, R.P.; et al. Multiple Repeats of a Promoter Segment Causes Transcription Factor Autoregulation in Red Apples[W]. Plant Cell 2009, 21, 168–183. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Tikunov, Y.; Schouten, R.E.; Marcelis, L.F.M.; Visser, R.G.F.; Bovy, A.G. Anthocyanin Biosynthesis and Degradation Mechanisms in Solanaceous Vegetables: A Review. Front. Chem. 2018. [Google Scholar] [CrossRef]
- Chagné, D.; Lin-Wang, K.; Espley, R.; Volz, R.K.; How, N.M.; Rouse, S.; Brendolise, C.; Carlisle, C.M.; Kumar, S.; De Silva, N.; et al. An Ancient Duplication of Apple MYB Transcription Factors Is Responsible for Novel Red Fruit-Flesh Phenotypes1[C][W]. Plant Physiol. 2012, 161, 225–239. [Google Scholar] [CrossRef] [Green Version]
- Volz, R.; Chagné, D.; Whitworth, C.; Espley, R.V.; Allan, A.; Carlisle, C.; Oraguzie, N.; Gardiner, S. Breeding for red flesh in apple. In Proceedings of the CF Mercer, 13th Australasian Plant Breeding Conference, Christchurch, New Zealand, 18–21 April 2006; p. 149. [Google Scholar]
- Volz, R.; Oraguzie, N.; Whitworth, C.; How, N.; Chagné, D.; Carlisle, C.; Gardiner, S. Red flesh breeding in apple: Progress and challenges. Acta Hortic. 2009, 814, 337–342. [Google Scholar] [CrossRef]
- Xu, J.; Li, X.; Liu, S.; Zhao, P.; Huo, H.; Zhang, Y. Effect of Nanocrystallization of Anthocyanins Extracted from Two Types of Red-Fleshed Apple Varieties on Its Stability and Antioxidant Activity. Molecules 2019. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.; Ku, K.; Jeong, M.; Kim, S.S.; Mitchell, A.E.; Lee, J. A comparison of the chemical composition and antioxidant activity of several new early- to mid-season apple cultivars for a warmer climate with traditional cultivars. J. Sci. Food Agric. 2019, 99, 4712–4724. [Google Scholar] [CrossRef]
- Telias, A.; Lin-Wang, K.; Stevenson, D.E.; Cooney, J.; Hellens, R.P.; Allan, A.C.; Hoover, E.E.; Bradeen, J. Apple skin patterning is associated with differential expression of MYB10. BMC Plant Boil. 2011. [Google Scholar] [CrossRef] [Green Version]
- El-Sharkawy, I.; Liang, D.; Xu, K. Transcriptome analysis of an apple (Malus x domestica) yellow fruit somatic mutation identifies a gene network module highly associated with anthocyanin and epigenetic regulation. J. Exp. Bot. 2015, 66, 7359–7376. [Google Scholar] [CrossRef] [Green Version]
- Mazza, G.; Velioglu, Y.S. Anthocyanins and other phenolic compounds in fruits of red-flesh apples. Food Chem. 1992, 43, 113–117. [Google Scholar] [CrossRef]
- Lachman, J.; Hamouz, K.; Orsak, M.; Pivec, V.; Hejtmánková, K.; Pazderů, K.; Dvořák, P.; Čepl, J. Impact of selected factors — Cultivar, storage, cooking and baking on the content of anthocyanins in coloured-flesh potatoes. Food Chem. 2012, 133, 1107–1116. [Google Scholar] [CrossRef]
- Yonekura-Sakakibara, K.; Higashi, Y.; Nakabayashi, R. The Origin and Evolution of Plant Flavonoid Metabolism. Front. Plant Sci. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB transcription factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Giusti, M.M.; Wrolstad, R.E. Characterization and Measurement of Anthocyanins by UV-Visible Spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, F1.2.1–F1.2.13. [Google Scholar] [CrossRef]
- Cai, W.G.; Wu, W.; Shao, J.F.; Chen, Q.; Wang, Y.; Liu, Z. Determination of polyphenol content in Houttuynia cordata Thunb. by Folin-Ciocalteu colorimetric method. Food Sci. 2010, 31, 201–204. [Google Scholar]
- He, X.H.; Xu, L.; Tan, M.J.; Du, F.L.; Zeng, J.G. DPPH radical scavenging effect of penthorum chinese pursh extract. Shizhen Guoyi Guoyao 2009, 20, 1924–1926. [Google Scholar]
- Ma, T.; Hu, N.; Ding, C.; Zhang, Q.; Li, W.; Suo, Y.; Wang, H.; Bai, B.; Ding, C. In vitro and in vivo biological activities of anthocyanins from Nitraria tangutorun Bobr. fruits. Food Chem. 2016, 194, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, T.; Watanabe, M.; Ohshima, K.; Norinobu, S.; Choi, S.-W.; Kawakishi, S.; Osawa, T. Antioxidative Activity of the Anthocyanin Pigments Cyanidin 3-O-.beta.-d-Glucoside and Cyanidin. J. Agric. Food Chem. 1994, 42, 2407–2410. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety of Red-Fleshed Apple | TAC | TPC | ||
|---|---|---|---|---|
| Peel | Flesh | Peel | Flesh | |
| Young stage | ||||
| XJ4 | 1702.6 ± 75.4a | 516.3 ± 35.6a | 8114.4 ± 152.4a | 3724.4 ± 359.9b |
| QN-5 | 461.3 ± 20.6b | 355.1 ± 118.4b | 3233.6 ± 675.8cd | 2522.6 ± 935.3cd |
| DH | 170.6 ± 47.5d | 99.9 ± 46.1c | 5277.3 ± 1229.1b | 3269.2 ± 288.9bc |
| HX1 | 109.1 ± 25.9de | 66.5 ± 1.9cd | 9082.3 ± 717.1a | 5346.1 ± 450.8a |
| Developmental stage | ||||
| XJ4 | 1506 ± 101.4a | 631 ± 38.7a | 5460.3 ± 532.3b | 2397.1 ± 699.5cd |
| QN-5 | 253.1 ± 46.9c | 361 ± 45.5b | 2724.6 ± 247.7cde | 1198.8 ± 181.2e |
| DH | 35 ± 15f | 54.2 ± 34.5cd | 3171.2 ± 528.1cd | 1127.1 ± 163.1e |
| HX1 | 111 ± 71.1de | 71.5 ± 11.7c | 2636.5 ± 154.5cde | 1974.9 ± 373.7d |
| Mature stage | ||||
| XJ4 | 441 ± 76.3b | 401 ± 13.5b | 3333.9 ± 432.2c | 2283.6 ± 520.2cd |
| QN-5 | 93.4 ± 3.3de | 76.6 ± 16.1c | 1239.6 ± 481.4f | 327.6 ± 54.4f |
| DH | 12.1 ± 3.4f | 24.3 ± 6.7d | 2069.3 ± 276de | 1010.2 ± 280.6e |
| HX1 | 80.1 ± 13.4e | 57.3 ± 1.1cd | 2691.9 ± 239.7cde | 1598.8 ± 287.3de |
| Variety of Red-Fleshed Apple | Scavenging Rate of DPPH/% | Scavenging Rate of OH/% | Scavenging Rate of O2·-/% | Dilution Ratio | ||||
|---|---|---|---|---|---|---|---|---|
| Peel | Flesh | Peel | Flesh | Peel | Flesh | Peel | Flesh | |
| Young stage | ||||||||
| XJ4 | 57.7 ± 2.8de | 80.9 ± 7.8ab | 14.3 ± 1.5e | 31.4 ± 8.0c | 22.1 ± 1.0d | 12 ± 4.6f | 34.0 | 10.3 |
| QN-5 | 88.1 ± 0.8a | 85.7 ± 1.0ab | 46.1 ± 2.9c | 49.5 ± 8.1b | 11.1 ± 1.3e | 18.4 ± 2.9ef | 9.2 | 7.1 |
| DH | 84.7 ± 6.1ab | 81.2 ± 3.8ab | 49.2 ± 5.6bc | 38.5 ± 1.7c | 27.2 ± 0.7cd | 27.6 ± 0.7cde | 3.4 | 2.0 |
| HX1 | 70.9 ± 1.7bc | 81.4 ± 3.6ab | 81.5 ± 7.3a | 51.6 ± 11.2b | 21.7 ± 0.8d | 15.3 ± 1.6ef | 2.1 | 1.3 |
| Developmental stage | ||||||||
| XJ4 | 90.7 ± 9.0a | 84.5 ± 1.6ab | 33.7 ± 3.1d | 37.3 ± 3.5c | 47.7 ± 12.6ab | 31.7 ± 2.4abc | 30.1 | 12.6 |
| QN-5 | 94.7 ± 0.2a | 94.6 ± 0.2ab | 43 ± 4.0cd | 54 ± 3.9b | 52.7 ± 4.8a | 25.9 ± 10.1cde | 5.0 | 7.2 |
| DH | 48.5 ± 10.2e | 80 ± 3.2b | 40.1 ± 6.0cd | 13.8 ± 3.1e | 29.4 ± 7.4cd | 21.1 ± 4.1def | −1.4 | 1.0 |
| HX1 | 65.8 ± 1.3cd | 83.5 ± 0.7ab | 41.8 ± 3.6cd | 34.2 ± 6.1c | 31.1 ± 3.2bcd | 17.1 ± 3.1ef | 2.2 | 1.4 |
| Mature stage | ||||||||
| XJ4 | 90.6 ± 2.1a | 97.1 ± 1.2a | 59.4 ± 8.6b | 75.1 ± 0.71a | 39.7 ± 2.7abc | 39.9 ± 2.5ab | 8.8 | 8.0 |
| QN-5 | 93.3 ± 2.5a | 94.7 ± 0.4ab | 2.4 ± 1.0g | 1.5 ± 0.4g | 38 ± 4.8abc | 43.3 ± 4.8a | 1.8 | 1.5 |
| DH | 21.4 ± 4.5f | 46.5 ± 3.0c | 48.9 ± 8.4bc | 20.1 ± 5.7d | 39 ± 3.2abc | 47.1 ± 4.7a | −4.6 | −2 |
| HX1 | 82.7 ± 1.6ab | 88.3 ± 0.9ab | 15.2 ± 3.0e | 4.3 ± 0.1f | 32.2 ± 2.5bcd | 36.8 ± 3.1abc | 1.6 | 1.1 |
| VC | 3.6 ± 1.6g | 3.6 ± 1.6d | 9.1 ± 0.8f | 9.1 ± 0.8e | 19.1 ± 9.0d | 19.1 ± 9.0ef | ||
| Combination Name | Metabolite Name | Content | Fold Change (XJ4PM/DHPM; XJ4FM/DHFM) | VIP | Grouping of Specific Metabolites | |
|---|---|---|---|---|---|---|
| DH | XJ4 | |||||
| ANTHOCYANINS | ||||||
| DHPM vs. XJ4PM | Pelargonidin 3-O-β-d-glucoside (callistephin chloride) | 9.00E + 00 | 3.14E + 07 | 3.49E + 06 | 2.36 | XJ4PM |
| Cyanidin 3-O-malonylhexoside | 9.00E + 00 | 1.32E + 06 | 1.47E + 05 | 2.1 | XJ4PM | |
| Malvidin 3-acetyl-5-diglucoside | 9.00E + 00 | 2.56E + 05 | 2.84E + 04 | 1.93 | XJ4PM | |
| Cyanidin 3-O-rutinoside (Keracyanin) | 9.00E + 00 | 2.40E + 05 | 2.67E + 04 | 1.94 | XJ4PM | |
| Pelargonidin 3-O-malonylhexoside | 9.00E + 00 | 1.63E + 04 | 1.82E + 03 | 1.65 | XJ4PM | |
| DHFM vs. XJ4FM | Pelargonidin 3-O-β-d-glucoside | 9.00E + 00 | 6.78E + 07 | 7.54E + 06 | 2.16 | XJ4FM |
| Cyanidin 3-O-malonylhexoside | 9.00E + 00 | 1.56E + 06 | 1.74E + 05 | 1.89 | XJ4FM | |
| Malvidin 3-acetyl-5-diglucoside | 9.00E + 00 | 1.06E + 06 | 1.17E + 05 | 1.86 | XJ4FM | |
| Peonidin O-hexoside | 9.00E + 00 | 7.50E + 05 | 8.33E + 04 | 1.83 | XJ4FM | |
| Peonidin 3-O-glucoside chloride | 9.00E + 00 | 7.12E + 05 | 7.91E + 04 | 1.83 | XJ4FM | |
| Pelargonidin 3-O-malonylhexoside | 9.00E + 00 | 2.49E + 04 | 2.76E + 03 | 1.53 | XJ4FM | |
| Rosinidin O-hexoside | 7.00E + 04 | 9.00E + 00 | 1.29E-04 | 1.62 | DHFM | |
| FLAVONES | ||||||
| DHPM vs. XJ4PM | Selgin O-hexosyl-O-hexoside | 9.00E + 00 | 3.09E + 06 | 3.43E + 05 | 2.17 | XJ4PM |
| Tricin O-saccharic acid | 9.00E + 00 | 3.02E + 06 | 3.36E + 05 | 2.16 | XJ4PM | |
| 6-C-hexosyl-hesperetin O-hexoside | 9.00E + 00 | 2.41E + 06 | 2.68E + 05 | 2.14 | XJ4PM | |
| Luteolin 3’,7-di-O-glucoside | 9.00E + 00 | 1.26E + 06 | 1.39E + 05 | 2.09 | XJ4PM | |
| O-methylChrysoeriol 5-O-hexoside | 9.00E + 00 | 1.09E + 06 | 1.22E + 05 | 1.38 | XJ4PM | |
| O-methylChrysoeriol 7-O-hexoside | 9.00E + 00 | 1.09E + 06 | 1.21E + 05 | 1.37 | XJ4PM | |
| C-hexosyl-apigenin O-hexosyl-O-hexoside | 9.00E + 00 | 8.63E + 05 | 9.59E + 04 | 2.05 | XJ4PM | |
| Eriodictiol 6-C-hexoside 8-C-hexoside-O-hexoside | 9.00E + 00 | 5.88E + 05 | 6.54E + 04 | 2.02 | XJ4PM | |
| Hesperetin C-hexosyl-O-hexosyl-O-hexoside | 9.00E + 00 | 5.85E + 05 | 6.50E + 04 | 2.02 | XJ4PM | |
| C-hexosyl-isorhamnetin O-hexoside | 9.00E + 00 | 1.67E + 05 | 1.86E + 04 | 1.9 | XJ4PM | |
| Tricin 4′-O-(syringyl alcohol) ether 5-O-hexoside | 9.00E + 00 | 1.55E + 05 | 1.73E + 04 | 1.89 | XJ4PM | |
| Morin | 9.00E + 00 | 7.46E + 05 | 8.29E + 04 | 2.04 | XJ4PM | |
| Persicoside | 9.00E + 00 | 5.93E + 05 | 6.59E + 04 | 2.02 | XJ4PM | |
| Chrysoeriol 8-C-hexoside | 9.00E + 00 | 4.92E + 04 | 5.47E + 03 | 1.25 | XJ4PM | |
| Apigenin | 9.00E + 00 | 4.38E + 04 | 4.87E + 03 | 1.22 | XJ4PM | |
| Acacetin | 9.00E + 00 | 3.31E + 04 | 3.68E + 03 | 1.66 | XJ4PM | |
| Tricin 5-O-rutinoside | 9.00E + 00 | 1.14E + 04 | 1.26E + 03 | 1.15 | XJ4PM | |
| Baicalein-7-O-glucuronide (Baicalin) | 5.51E + 04 | 9.00E + 00 | 1.63E-04 | 1.79 | DHPM | |
| Diosmin | 2.21E + 06 | 9.00E + 00 | 4.08E-06 | 2.11 | DHPM | |
| Chrysoeriol 7-O-rutinoside | 9.30E + 05 | 9.00E + 00 | 9.67E-06 | 2.03 | DHPM | |
| DHFM vs. XJ4FM | 6-C-hexosyl-hesperetin O-hexoside | 9.00E + 00 | 7.26E + 06 | 8.06E + 05 | 2 | XJ4FM |
| Luteolin 3’,7-di-O-glucoside | 9.00E + 00 | 3.64E + 06 | 4.04E + 05 | 1.95 | XJ4FM | |
| Selgin O-hexosyl-O-hexoside | 9.00E + 00 | 1.98E + 06 | 2.20E + 05 | 1.91 | XJ4FM | |
| 8-C-hexosyl-hesperetin O-hexoside | 9.00E + 00 | 6.70E + 05 | 7.45E + 04 | 1.82 | XJ4FM | |
| Eriodictiol 6-C-hexoside 8-C-hexoside-O-hexoside | 9.00E + 00 | 6.06E + 05 | 6.74E + 04 | 1.81 | XJ4FM | |
| Hesperetin C-hexosyl-O-hexosyl-O-hexoside | 9.00E + 00 | 4.05E + 05 | 4.50E + 04 | 1.78 | XJ4FM | |
| Limocitrin O-hexoside | 9.00E + 00 | 3.43E + 05 | 3.81E + 04 | 1.77 | XJ4FM | |
| Chrysoeriol 8-C-hexoside | 9.00E + 00 | 2.49E + 05 | 2.76E + 04 | 1.74 | XJ4FM | |
| Chrysoeriol 6-C-hexoside 8-C-hexoside-O-hexoside | 9.00E + 00 | 2.28E + 05 | 2.53E + 04 | 1.73 | XJ4FM | |
| Butin | 9.00E + 00 | 1.82E + 05 | 2.02E + 04 | 1.71 | XJ4FM | |
| Eriocitrin | 9.00E + 00 | 7.77E + 04 | 8.63E + 03 | 1.64 | XJ4FM | |
| Tricin O-saccharic acid | 9.00E + 00 | 5.43E + 04 | 6.03E + 03 | 1.6 | XJ4FM | |
| O-methylChrysoeriol 7-O-hexoside | 9.00E + 00 | 4.73E + 04 | 5.26E + 03 | 1.59 | XJ4FM | |
| Tricin 5-O-rutinoside | 9.00E + 00 | 4.45E + 04 | 4.94E + 03 | 1.59 | XJ4FM | |
| O-methylChrysoeriol 5-O-hexoside | 9.00E + 00 | 3.99E + 04 | 4.43E + 03 | 1.58 | XJ4FM | |
| Apigenin | 9.00E + 00 | 1.39E + 04 | 1.55E + 03 | 1.03 | XJ4FM | |
| Chrysoeriol | 9.00E + 00 | 7.42E + 03 | 8.25E + 02 | 1.41 | XJ4FM | |
| Chrysoeriol 7-O-rutinoside | 1.68E + 04 | 9.00E + 00 | 5.37E-04 | 1.04 | DHFM | |
| C-hexosyl-luteolin O-sinapic acid | 2.41E + 04 | 9.00E + 00 | 3.74E-04 | 1.52 | DHFM | |
| Luteolin C-hexoside | 2.42E + 04 | 9.00E + 00 | 3.72E-04 | 1.53 | DHFM | |
| 6-C-hexosyl luteolin O-pentoside | 3.15E + 04 | 9.00E + 00 | 2.86E-04 | 1.09 | DHFM | |
| Diosmin | 3.20E + 04 | 9.00E + 00 | 2.81E-04 | 1.08 | DHFM | |
| Luteolin O-sinapoylhexoside | 4.34E + 04 | 9.00E + 00 | 2.07E-04 | 1.58 | DHFM | |
| 3-O-Acetylpinobanksin | 4.79E + 04 | 9.00E + 00 | 1.88E-04 | 1.53 | DHFM | |
| FLAVONOLS | ||||||
| DHPM vs. XJ4PM | Quercetin | 9.00E + 00 | 5.59E + 05 | 6.21E + 04 | 2.02 | XJ4PM |
| Laricitrin | 9.00E + 00 | 6.04E + 04 | 6.71E + 03 | 1.8 | XJ4PM | |
| Isorhamnetin | 9.00E + 00 | 2.26E + 04 | 2.51E + 03 | 1.7 | XJ4PM | |
| Rhamnetin (7-O-methxyl quercetin) | 9.00E + 00 | 3.26E + 03 | 3.62E + 02 | 1.47 | XJ4PM | |
| Kaempferol 3-O-rutinoside (Nicotiflorin) | 1.16E + 06 | 9.00E + 00 | 7.77E-06 | 2.08 | DHPM | |
| DHFM vs. XJ4FM | Syringetin 3-O-hexoside | 9.00E + 00 | 4.68E + 05 | 5.20E + 04 | 1.79 | XJ4FM |
| Rhamnetin | 9.00E + 00 | 5.00E + 03 | 5.56E + 02 | 1.36 | XJ4FM | |
| Isorhamnetin 3-O-neohesperidoside | 1.08E + 04 | 9.00E + 00 | 8.33E-04 | 1.01 | DHFM | |
| Kaempferol 3-O-robinobioside (Biorobin) | 3.84E + 04 | 9.00E + 00 | 2.34E-04 | 1.1 | DHFM | |
| Syringetin | 6.33E + 04 | 9.00E + 00 | 1.42E-04 | 1.6 | DHFM | |
| FLAVANONES | ||||||
| DHPM vs. XJ4PM | Hesperetin O-malonylhexoside | 9.00E + 00 | 1.22E + 05 | 1.35E + 04 | 1.87 | XJ4PM |
| DHFM vs. XJ4FM | Naringenin | 9.00E + 00 | 1.92E + 05 | 2.13E + 04 | 1.72 | XJ4FM |
| Hesperetin O-malonylhexoside | 9.00E + 00 | 1.55E + 05 | 1.72E + 04 | 1.7 | XJ4FM | |
| Naringenin 7-O-neohesperidoside (Naringin) | 9.00E + 00 | 9.60E + 04 | 1.07E + 04 | 1.66 | XJ4FM | |
| Hesperetin O-Glucuronic acid | 9.00E + 00 | 4.25E + 04 | 4.72E + 03 | 1.58 | XJ4FM | |
| ISOFLAVONES | ||||||
| DHPM vs. XJ4PM | Genistein 7-O-Glucoside (Genistin) | 9.00E + 00 | 1.87E + 05 | 2.08E + 04 | 1.92 | XJ4PM |
| DHFM vs. XJ4FM | Rotenone | 9.00E + 00 | 4.27E + 04 | 4.74E + 03 | 1.58 | XJ4FM |
| Genistein 7-O-Glucoside | 9.00E + 00 | 2.79E + 04 | 3.10E + 03 | 1.08 | XJ4FM | |
| Glycitein | 9.00E + 00 | 9.10E + 03 | 1.01E + 03 | 1.43 | XJ4FM | |
| Calycosin | 9.00E + 00 | 6.75E + 03 | 7.50E + 02 | 1.4 | XJ4FM | |
| POLYPHENOLS | ||||||
| DHFM vs. XJ4FM | Gallocatechin-gallocatechin | 9.00E + 00 | 2.90E + 07 | 3.23E + 06 | 2.11 | XJ4FM |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Xu, J.; Xu, Z.; Sun, X.; Zhu, J.; Zhang, Y. Analysis of Antioxidant Activity and Flavonoids Metabolites in Peel and Flesh of Red-Fleshed Apple Varieties. Molecules 2020, 25, 1968. https://doi.org/10.3390/molecules25081968
Zhang X, Xu J, Xu Z, Sun X, Zhu J, Zhang Y. Analysis of Antioxidant Activity and Flavonoids Metabolites in Peel and Flesh of Red-Fleshed Apple Varieties. Molecules. 2020; 25(8):1968. https://doi.org/10.3390/molecules25081968
Chicago/Turabian StyleZhang, Xiang, Jihua Xu, Zhaobo Xu, Xiaohong Sun, Jun Zhu, and Yugang Zhang. 2020. "Analysis of Antioxidant Activity and Flavonoids Metabolites in Peel and Flesh of Red-Fleshed Apple Varieties" Molecules 25, no. 8: 1968. https://doi.org/10.3390/molecules25081968
APA StyleZhang, X., Xu, J., Xu, Z., Sun, X., Zhu, J., & Zhang, Y. (2020). Analysis of Antioxidant Activity and Flavonoids Metabolites in Peel and Flesh of Red-Fleshed Apple Varieties. Molecules, 25(8), 1968. https://doi.org/10.3390/molecules25081968