
Comparative Analysis of Milk Fat Globular Membrane (MFGM) Proteome between Saudi Arabia Camelus dromedary Safra and Wadha Breeds


 , , , and
, , , and
Abstract
:1. Introduction
2. Results
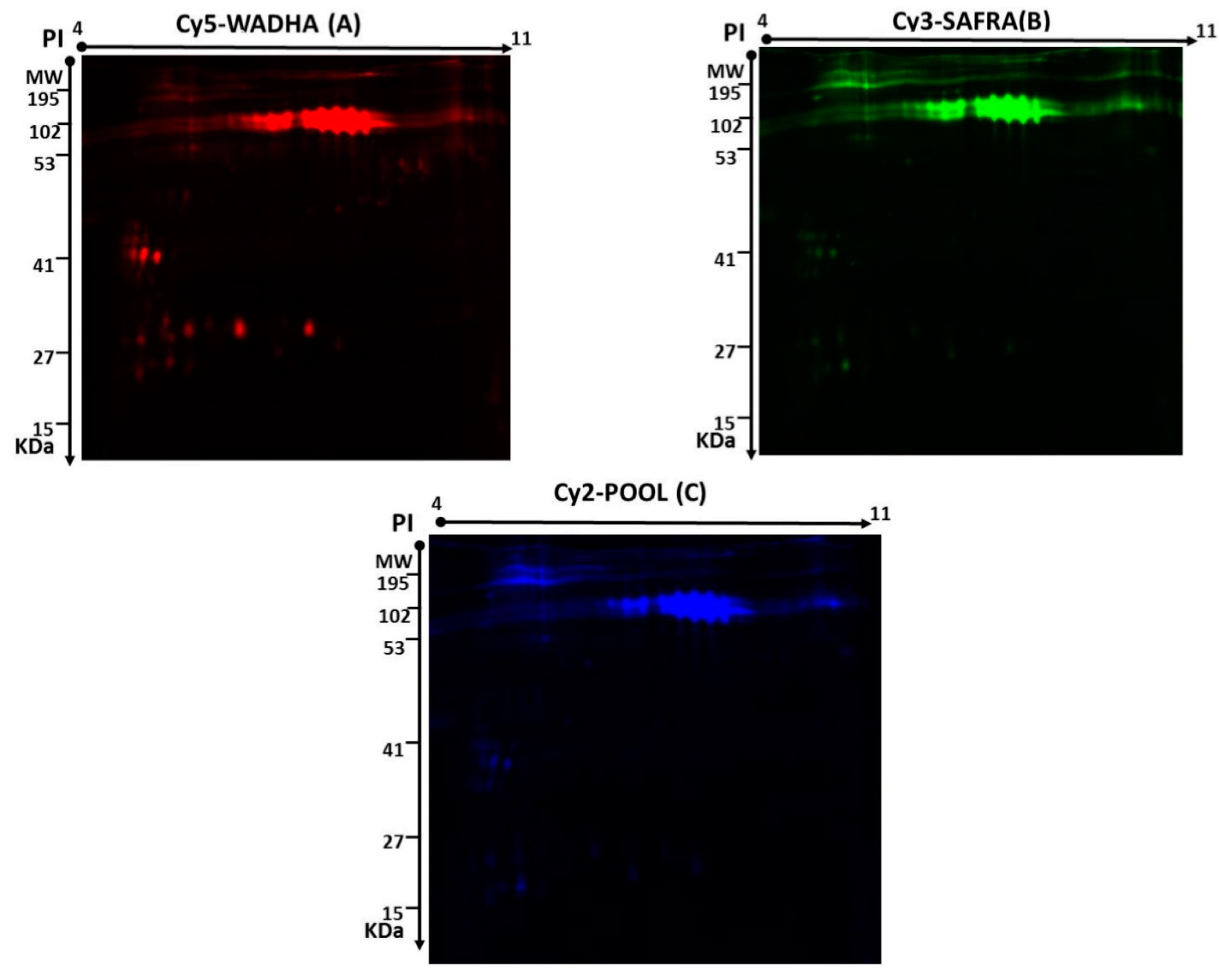
2.1. CyDye Fluorescence Labeling and Two-Dimensional Difference in Gel Electrophoresis (2D-DIGE)
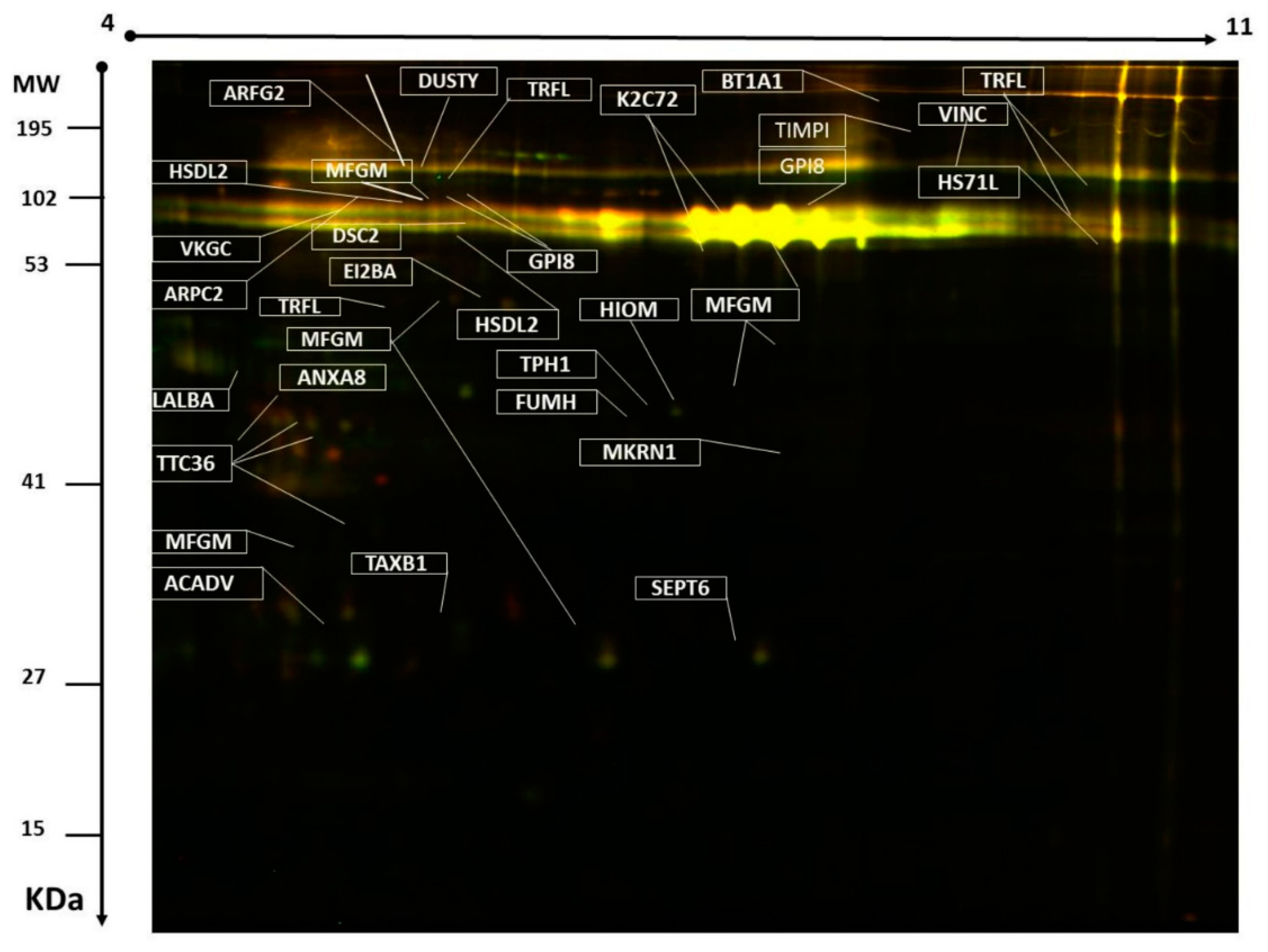
2.2. Mass Spectrometry and Identification of Proteins
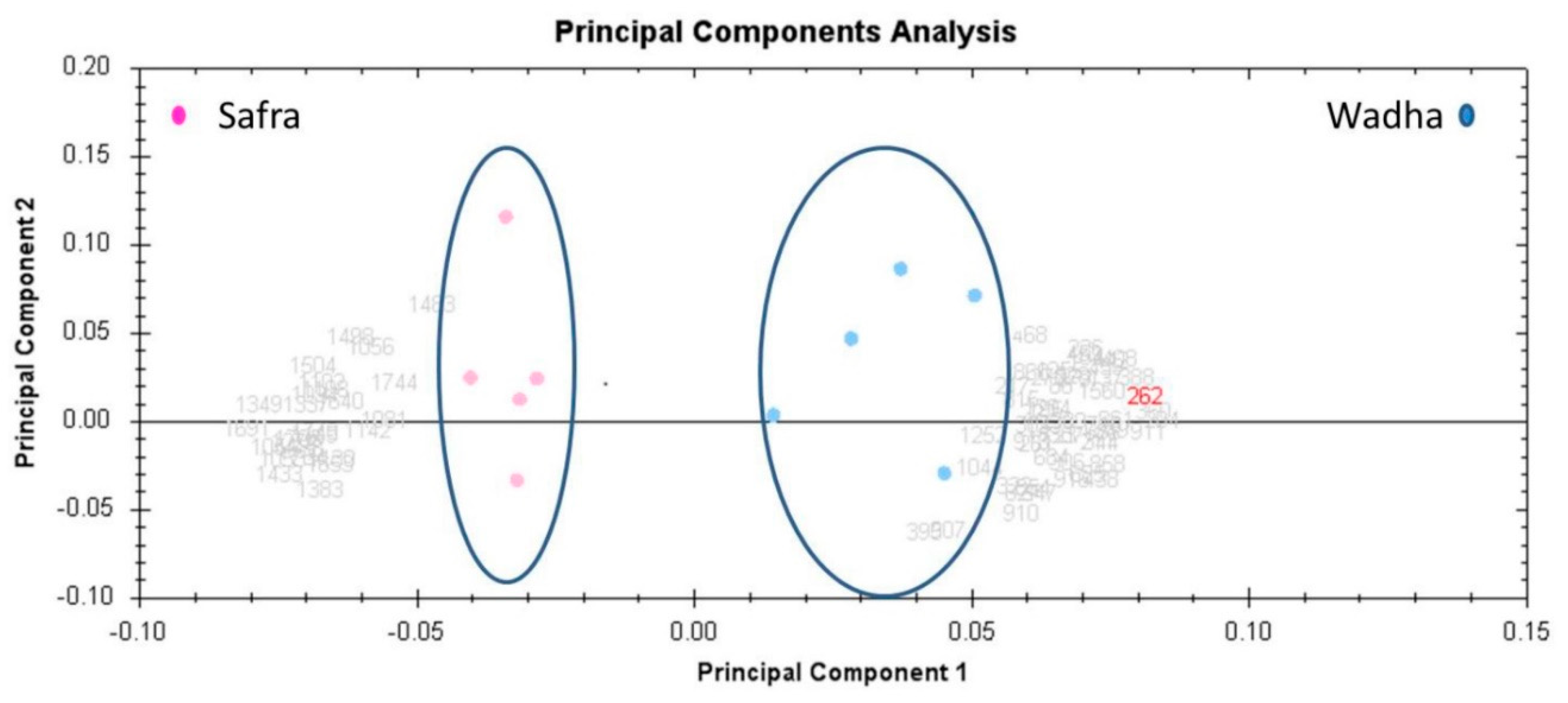
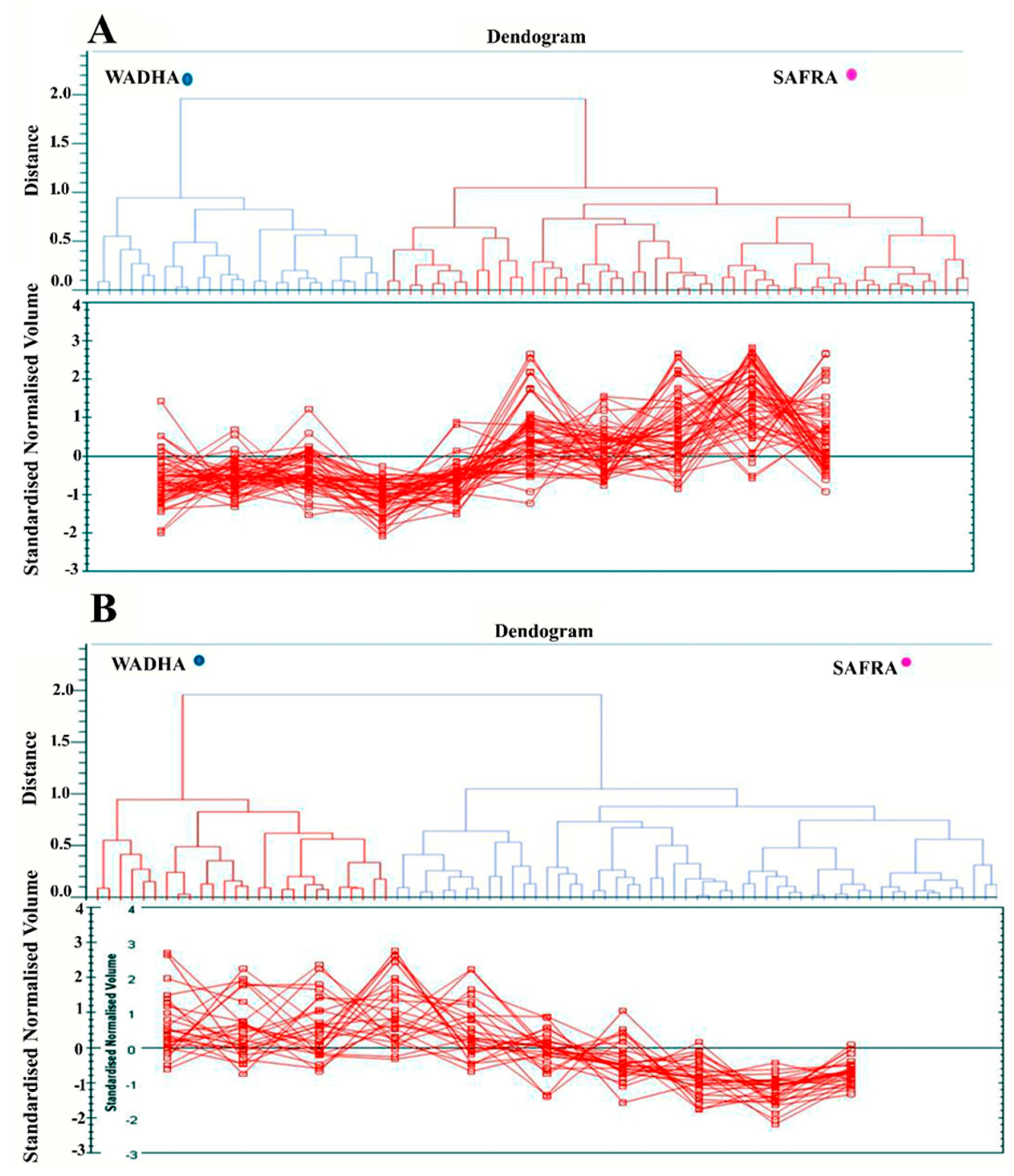
2.3. PCA and Cluster Analysis
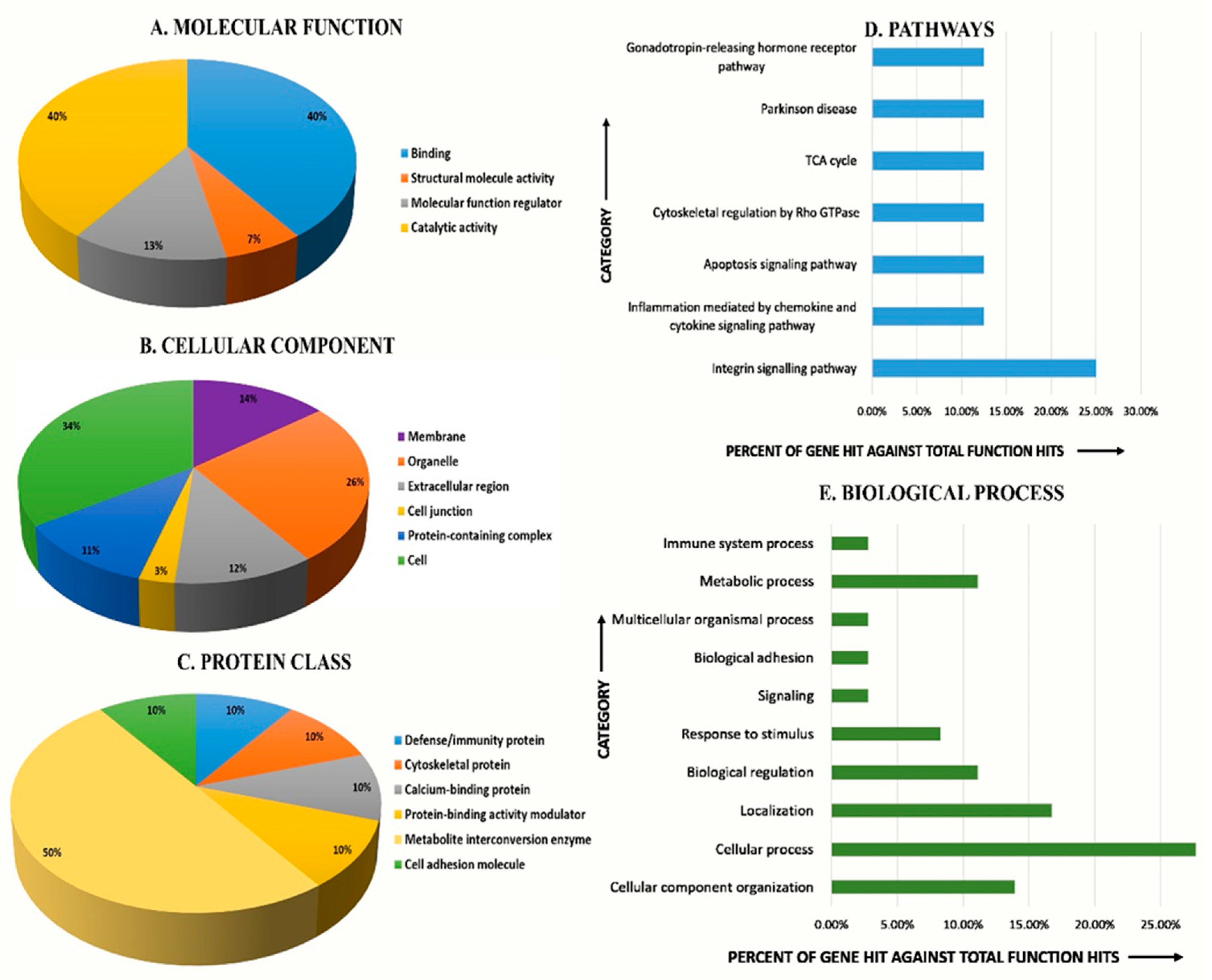
2.4. Classification of Proteins Based on Function, Cellular Component, Pathway, and Biological Process
2.5. Validation by ELISA Immunoassay
3. Discussion and Conclusions
4. Materials and Methods
4.1. Camel Milk Samples Collection
4.2. Preparation of MFGM Samples
4.3. Extraction of MFGM Proteins
4.4. D-DIGE and MALDI-TOF Analysis
4.4.1. Sample Labeling with Cyanine Dyes
4.4.2. One- and Two-Dimensional Difference Electrophoresis (2D-DIGE)
4.4.3. Statistical Analysis
4.4.4. Protein Identification by MALDI-TOF MS
4.5. Gene Ontology Enrichment Analysis
4.6. ELISA Immunoassay Validation for Selected MFGM Proteins
4.7. Hierarchical Clustering and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- El-Fakharany, E.M.; El-Baky, N.A.; Linjawi, M.H.; Aljaddawi, A.A.; Saleem, T.H.; Nassar, A.Y.; Osman, A.; Redwan, E.M. Influence of camel milk on the hepatitis C virus burden of infected patients. Exp. Ther. Med. 2017, 13, 1313–1320. [Google Scholar] [CrossRef] [PubMed]
- Cissé, H.; Muandze-Nzambe, J.U.; Somda, N.S.; Sawadogo, A.; Drabo, S.M.; Tapsoba, F.; Zongo, C.; Traoré, Y.; Savadogo, A. Assessment of safety and quality of fermented milk of camels, cows, and goats sold and consumed in five localities of Burkina Faso. Vet. World 2019, 12, 295. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, R.R.; Santos, R.; Cardoso, C.; Carvalho, M. Consumption of camel’s milk by patients intolerant to lactose. A preliminary study. Revista Alergia de Mexico 2010, 57, 26–32. [Google Scholar]
- El-Fakharany, E.M.; Tabll, A.; El-Wahab, A.A.; Haroun, B.M.; Redwan, E.R.M. Potential activity of camel milk-amylase and lactoferrin against hepatitis C virus infectivity in HepG2 and lymphocytes. Hepat. Mon. 2008, 8, 101–109. [Google Scholar]
- Faris, M.; Ghazzawi, H. Health-Improving and Disease-Preventing Potential of Camel Milk against Chronic Diseases and Autism: Camel Milk and Chronic Diseases; IGI Global: Hershey, PA, USA, 2020; pp. 155–184. [Google Scholar]
- Mirmiran, P.; Ejtahed, H.S.; Angoorani, P.; Eslami, F.; Azizi, F. Camel Milk Has Beneficial Effects on Diabetes Mellitus: A Systematic Review. Int. J. Endocrinol. Metab. 2017, 15, e42150. [Google Scholar] [CrossRef] [Green Version]
- Mihic, T.; Rainkie, D.; Wilby, K.J.; Pawluk, S.A. The Therapeutic Effects of Camel Milk: A Systematic Review of Animal and Human Trials. J. Evid.-Based Complement. Altern. Med. 2016, 21, Np110–Np126. [Google Scholar] [CrossRef]
- Albar, A.H.; Almehdar, H.A.; Uversky, V.N.; Redwan, E.M. Structural heterogeneity and multifunctionality of lactoferrin. Curr. Protein Pept. Sci. 2014, 15, 778–797. [Google Scholar] [CrossRef]
- Almehdar, H.A.; El-Baky, N.A.; Alhaider, A.A.; Almuhaideb, S.A.; Alhaider, A.A.; Albiheyri, R.S.; Uversky, V.N.; Redwan, E.M. Bacteriostatic and Bactericidal Activities of Camel Lactoferrins Against Salmonella enterica Serovar Typhi. Probiotics Antimicrob. Proteins 2019, 12, 18–31. [Google Scholar] [CrossRef]
- El-Fakharany, E.M.; Uversky, V.N.; Redwan, E.M. Comparative Analysis of the Antiviral Activity of Camel, Bovine, and Human Lactoperoxidases against Herpes Simplex Virus Type 1. Appl. Biochem. Biotechnol. 2017, 182, 294–310. [Google Scholar] [CrossRef]
- Redwan, E.M.; Uversky, V.N.; El-Fakharany, E.M.; Al-Mehdar, H. Potential lactoferrin activity against pathogenic viruses. Comptes Rendus Biol. 2014, 337, 581–595. [Google Scholar] [CrossRef]
- Redwan, E.M.; El-Baky, N.A.; Al-Hejin, A.M.; Baeshen, M.N.; Almehdar, H.A.; Elsaway, A.; Gomaa, A.B.; Al-Masaudi, S.B.; Al-Fassi, F.A.; AbuZeid, I.E.; et al. Significant antibacterial activity and synergistic effects of camel lactoferrin with antibiotics against methicillin-resistant Staphylococcus aureus (MRSA). Res. Microbiol. 2016, 167, 480–491. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Padhi, E.; Hasegawa, Y.; Larke, J.; Parenti, M.; Wang, A.; Hernell, O.; Lönnerdal, B.; Slupsky, C. Compositional dynamics of the milk fat globule and its role in infant development. Front. Pediatr. 2018, 6, 313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redwan, E.M.; Alkarim, S.A.; El-Hanafy, A.A.; Saad, Y.M.; Almehdar, H.A.; Uversky, V.N. Variability of Some Milk-Associated Genes and Proteins in Several Breeds of Saudi Arabian Camels. Protein J. 2018, 37, 333–352. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, T.A.; Lippolis, J.D. Bovine milk fat globule membrane proteome. J. Dairy Res. 2006, 73, 406–416. [Google Scholar] [CrossRef] [Green Version]
- McManaman, J.L.; Neville, M.C. Mammary physiology and milk secretion. Adv. Drug Deliv. Rev. 2003, 55, 629–641. [Google Scholar] [CrossRef]
- Keenan, T.W.; Patton, S. The structure of milk: Implications for sampling and storage: A. The milk lipid globule membrane. In Handbook of Milk Composition; Elsevier: Amsterdam, The Netherlands, 1995; pp. 5–50. [Google Scholar]
- Peterson, J.A.; Patton, S.; Hamosh, M. Glycoproteins of the human milk fat globule in the protection of the breast-fed infant against infections. Neonatology 1998, 74, 143–162. [Google Scholar] [CrossRef]
- Imam, A.; Drushella, M.M.; Taylor, C.R.; Tökés, Z.A. Preferential expression of a Mr 155,000 milk-fat-globule membrane glycoprotein on luminal epithelium of lobules in human breast. Cancer Res. 1986, 46 (12 Part 1), 6374–6379. [Google Scholar]
- Snow, D.R.; Jimenez-Flores, R.; Ward, R.E.; Cambell, J.; Young, M.J.; Nemere, I.; Hintze, K.J. Dietary milk fat globule membrane reduces the incidence of aberrant crypt foci in Fischer-344 rats. J. Agric. Food Chem. 2010, 58, 2157–2163. [Google Scholar] [CrossRef]
- Brink, L.R.; Herren, A.W.; McMillen, S.; Fraser, K.; Agnew, M.; Roy, N.; Lonnerdal, B. Omics analysis reveals variations among commercial sources of bovine milk fat globule membrane. J. Dairy Sci. 2020, 103, 3002–3016. [Google Scholar] [CrossRef] [Green Version]
- De Almagro Garcia, M.C.; Moreno Munoz, J.A.; Jimenez Lopez, J.; Rodriguez-Palmero Seuma, M. New ingredients in infant formula. Health Funct. Benefits Nutr. Hosp. 2017, 34 (Suppl. 4), 8–12. [Google Scholar]
- Demmelmair, H.; Prell, C.; Timby, N.; Lonnerdal, B. Benefits of Lactoferrin, Osteopontin and Milk Fat Globule Membranes for Infants. Nutrients 2017, 9, 817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fil, J.E.; Fleming, S.A.; Chichlowski, M.; Gross, G.; Berg, B.M.; Dilger, R.N. Evaluation of Dietary Bovine Milk Fat Globule Membrane Supplementation on Growth, Serum Cholesterol and Lipoproteins, and Neurodevelopment in the Young Pig. Front. Pediatr. 2019, 7, 417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernell, O.; Timby, N.; Domellof, M.; Lonnerdal, B. Clinical Benefits of Milk Fat Globule Membranes for Infants and Children. J. Pediatr. 2016, 173, S60–S65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernell, O.; Domellof, M.; Grip, T.; Lonnerdal, B.; Timby, N. Physiological Effects of Feeding Infants and Young Children Formula Supplemented with Milk Fat Globule Membranes. In Human Milk: Composition, Clinical Benefits and Future Opportunities; Nestle Nutrition Institute workshop series; Karger: Basel, Switzerland, 2019; Volume 90, pp. 35–42. [Google Scholar]
- Huang, S.; Wu, Z.; Liu, C.; Han, D.; Feng, C.; Wang, S.; Wang, J. Milk Fat Globule Membrane Supplementation Promotes Neonatal Growth and Alleviates Inflammation in Low-Birth-Weight Mice Treated with Lipopolysaccharide. Biomed Res. Int. 2019, 2019, 4876078. [Google Scholar] [CrossRef] [Green Version]
- Juvarajah, T.; Wan-Ibrahim, W.I.; Ashrafzadeh, A.; Othman, S.; Hashim, O.H.; Fung, S.Y.; Abdul-Rahman, P.S. Human Milk Fat Globule Membrane Contains Hundreds of Abundantly Expressed and Nutritionally Beneficial Proteins That Are Generally Lacking in Caprine Milk. Breastfeed. Med. 2018, 13, 631–637. [Google Scholar] [CrossRef]
- Lucey, J.A.; Otter, D.; Horne, D.S. A 100-Year Review: Progress on the chemistry of milk and its components. J. Dairy Sci. 2017, 100, 9916–9932. [Google Scholar] [CrossRef] [Green Version]
- Norris, T.; Souza, R.; Xia, Y.; Zhang, T.; Rowan, A.; Gallier, S.; Zhang, H.; Qi, H.; Baker, P. Effect of supplementation of complex milk lipids in pregnancy on fetal growth: Results from the Complex Lipids in Mothers and Babies (CLIMB) randomized controlled trial. J. Matern.-Fetal Neonatal Med. 2019, 1–10. [Google Scholar] [CrossRef]
- Davies, C.R.; Morris, J.S.; Griffiths, M.R.; Page, M.J.; Pitt, A.; Stein, T.; Gusterson, B.A. Proteomic analysis of the mouse mammary gland is a powerful tool to identify novel proteins that are differentially expressed during mammary development. Proteomics 2006, 6, 5694–5704. [Google Scholar] [CrossRef]
- Yamazaki, T.; Goya, I.; Graf, D.; Craig, S.; Martin-Orozco, N.; Dong, C. A butyrophilin family member critically inhibits T cell activation. J. Immunol. 2010, 185, 5907–5914. [Google Scholar] [CrossRef] [Green Version]
- Ji, X.; Li, X.; Ma, Y.; Li, D. Differences in proteomic profiles of milk fat globule membrane in yak and cow milk. Food Chem. 2017, 221, 1822–1827. [Google Scholar] [CrossRef]
- Sui, S.; Zhao, J.; Wang, J.; Zhang, R.; Guo, C.; Yu, T.; Li, N. Comparative proteomics of milk fat globule membrane proteins from transgenic cloned cattle. PLoS ONE 2014, 9, e105378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cebo, C.; Caillat, H.; Bouvier, F.; Martin, P. Major proteins of the goat milk fat globule membrane. J. Dairy Sci. 2010, 93, 868–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cebo, C.; Rebours, E.; Henry, C.; Makhzami, S.; Cosette, P.; Martin, P. Identification of major milk fat globule membrane proteins from pony mare milk highlights the molecular diversity of lactadherin across species. J. Dairy Sci. 2012, 95, 1085–1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murgiano, L.; Timperio, A.M.; Zolla, L.; Bongiorni, S.; Valentini, A.; Pariset, L. Comparison of milk fat globule membrane (MFGM) proteins of Chianina and Holstein cattle breed milk samples through proteomics methods. Nutrients 2009, 1, 302–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Costa, W.K.A.; de Souza, E.L.; Beltrao-Filho, E.M.; Vasconcelos, G.K.V.; Santi-Gadelha, T.; de Almeida Gadelha, C.A.; Franco, O.L.; do Egypto, R.d.C.R.; Magnani, M. Comparative protein composition analysis of goat milk produced by the Alpine and Saanen breeds in northeastern Brazil and related antibacterial activities. PLoS ONE 2014, 9, e93361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saadaoui, B.; Henry, C.; Khorchani, T.; Mars, M.; Martin, P.; Cebo, C. Proteomics of the milk fat globule membrane from C amelus dromedarius. Proteomics 2013, 13, 1180–1184. [Google Scholar] [CrossRef]
- Nilsson, Å. Role of sphingolipids in infant gut health and immunity. J. Pediatr. 2016, 173, S53–S59. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Zavaleta, N.; Chen, S.-Y.; Lönnerdal, B.; Slupsky, C. Effect of bovine milk fat globule membranes as a complementary food on the serum metabolome and immune markers of 6-11-month-old Peruvian infants. NPJ Sci. Food 2018, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Alvarado, R.; Phinney, B.; Lönnerdal, B. Proteomic characterization of human milk fat globule membrane proteins during a 12 month lactation period. J. Proteome Res. 2011, 10, 3530–3541. [Google Scholar] [CrossRef]
- Pacheco, A.R.; Barile, D.; Underwood, M.A.; Mills, D.A. The impact of the milk glycobiome on the neonate gut microbiota. Annu. Rev. Anim. Biosci. 2015, 3, 419–445. [Google Scholar] [CrossRef] [Green Version]
- Georgi, G.; Bartke, N.; Wiens, F.; Stahl, B. Functional glycans and glycoconjugates in human milk. Am. J. Clin. Nutr. 2013, 98, 578S–585S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, X.; Xu, W.; Cui, J.; Ma, Y.; Zhou, S. Goat and buffalo milk fat globule membranes exhibit better effects at inducing apoptosis and reduction the viability of HT-29 cells. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanabria, R.; Tellez, A.M.; Griffiths, M.; Corredig, M. Milk fat globule membrane isolate induces apoptosis in HT-29 human colon cancer cells. Food Funct. 2013, 4, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Redwan, E.M.; Al-Hejin, A.M.; Almehdar, H.A.; Elsaway, A.M.; Uversky, V.N. Prediction of Disordered Regions and Their Roles in the Anti-Pathogenic and Immunomodulatory Functions of Butyrophilins. Molecules 2018, 23, 328. [Google Scholar] [CrossRef] [Green Version]
- Guggenmos, J.; Schubart, A.S.; Ogg, S.; Andersson, M.; Olsson, T.; Mather, I.H.; Linington, C. Antibody Cross-Reactivity between Myelin Oligodendrocyte Glycoprotein and the Milk Protein Butyrophilin in Multiple Sclerosis. J. Immunol. 2004, 172, 661–668. [Google Scholar] [CrossRef] [Green Version]
- Malinowska, M.; Tokarz-Deptuła, B.; Deptuła, W. Butyrophilins: An important new element of resistance. Cent.-Eur. J. Immunol. 2017, 42, 399. [Google Scholar] [CrossRef]
- Stefferl, A.; Schubart, A.; Storch, M.; Amini, A.; Mather, I.; Lassmann, H.; Linington, C. Butyrophilin, a milk protein, modulates the encephalitogenic T cell response to myelin oligodendrocyte glycoprotein in experimental autoimmune encephalomyelitis. J. Immunol. 2000, 165, 2859–2865. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Wang, X.; Zhang, W.; Liu, L.; Pang, X.; Zhang, S.; Lv, J. Comparative proteomics of milk fat globule membrane in different species reveals variations in lactation and nutrition. Food Chem. 2016, 196, 665–672. [Google Scholar] [CrossRef]
- Redington, J.M.; Breydo, L.; Almehdar, H.A.; Redwan, E.M.; Uversky, V.N. α-lactalbumin: Of Camels and Cows. Protein Pept. Lett. 2016, 23, 1072–1080. [Google Scholar] [CrossRef]
- Lönnerdal, B. Bioactive proteins in human milk: Health, nutrition, and implications for infant formulas. J. Pediatr. 2016, 173, S4–S9. [Google Scholar] [CrossRef] [Green Version]
- Håkansson, A.; Zhivotovsky, B.; Orrenius, S.; Sabharwal, H.; Svanborg, C. Apoptosis induced by a human milk protein. Proc. Natl. Acad. Sci. USA 1995, 92, 8064–8068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svensson, M.; Düringer, C.; Hallgren, O.; Mossberg, A.-K.; Håkansson, A.; Linse, S.; Svanborg, C. Hamlet—A complex from human milk that induces apoptosis in tumor cells but spares healthy cells. In Integrating Population Outcomes, Biological Mechanisms and Research Methods in the Study of Human Milk and Lactation; Springer: Berlin/Heidelberg, Germany, 2002; pp. 125–132. [Google Scholar]
- Svensson, M.; Håkansson, A.; Mossberg, A.-K.; Linse, S.; Svanborg, C. Conversion of α-lactalbumin to a protein inducing apoptosis. Proc. Natl. Acad. Sci. USA 2000, 97, 4221–4226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Fakharany, E.M.; Redwan, E.M. Protein–lipid complexes: Molecular structure, current scenarios and mechanisms of cytotoxicity. RSC Adv. 2019, 9, 36890–36906. [Google Scholar] [CrossRef] [Green Version]
- Uversky, V.N.; El-Fakharany, E.M.; Abu-Serie, M.M.; Almehdar, H.A.; Redwan, E.M. Divergent Anticancer Activity of Free and Formulated Camel Milk α-lactalbumin. Cancer Investig. 2017, 35, 610–623. [Google Scholar] [CrossRef]
- Breydo, L.; Almehdar, H.A.; El-Fakharany, E.M.; Redwan, E.M.; Uversky, V.N. Not all AMLETs are made equal: Complexes of cow and camel α-lactalbumin with oleic acid show different structure and stability. J. Biomol. Struct. Dyn. 2018, 36, 4405–4412. [Google Scholar] [CrossRef]
- Jiang, L.; Kwong, D.L.-W.; Li, Y.; Liu, M.; Yuan, Y.-F.; Li, Y.; Fu, L.; Guan, X.-Y. HBP21, a chaperone of heat shock protein 70, functions as a tumor suppressor in hepatocellular carcinoma. Carcinogenesis 2015, 36, 1111–1120. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Gao, J.; Chen, X.; Chen, Y.; Chen, J.; Wang, S.; Liu, J.; Liu, X.; Li, J. HBP21: A novel member of TPR motif family, as a potential chaperone of heat shock protein 70 in proliferative vitreoretinopathy (PVR) and breast cancer. Mol. Biotechnol. 2008, 40, 231–240. [Google Scholar] [CrossRef]
- Giansanti, F.; Panella, G.; Leboffe, L.; Antonini, G. Lactoferrin from milk: Nutraceutical and pharmacological properties. Pharmaceuticals 2016, 9, 61. [Google Scholar] [CrossRef] [Green Version]
- Somiari, R.I.; Sullivan, A.; Russell, S.; Somiari, S.; Hu, H.; Jordan, R.; George, A.; Katenhusen, R.; Buchowiecka, A.; Arciero, C. High-throughput proteomic analysis of human infiltrating ductal carcinoma of the breast. Proteomics 2003, 3, 1863–1873. [Google Scholar] [CrossRef]
- Kawahara, E.; Tokuda, R.; Nakanishi, I. Migratory phenotypes of HSC-3 squamous carcinoma cell line induced by EGF and PMA: Relevance to migration of loosening of adhesion and vinculin-associated focal contacts with prominent filopodia. Cell Biol. Int. 1999, 23, 163–174. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Guo, H.; Song, Y.; Zhao, X.; Shi, Y.; Lu, Y.; Hu, S.; Nie, Y.; Fan, D.; Wu, K. Loss of vinculin and membrane-bound β-catenin promotes metastasis and predicts poor prognosis in colorectal cancer. Mol. Cancer 2014, 13, 263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zha, J.; Zhou, Q.; Xu, L.-G.; Chen, D.; Li, L.; Zhai, Z.; Shu, H.-B. RIP5 is a RIP-homologous inducer of cell death. Biochem. Biophys. Res. Commun. 2004, 319, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Han, Y.; Yoon, Y.; Yun, C.; Yun, S.; Kim, S.; Kwon, H.; Jeong, D.; Baek, M.; Lee, H. Role of HSPA1L as a cellular prion protein stabilizer in tumor progression via HIF-1α/GP78 axis. Oncogene 2017, 36, 6555–6567. [Google Scholar] [CrossRef] [PubMed]
- Rammer, P.; Groth-Pedersen, L.; Kirkegaard, T.; Daugaard, M.; Rytter, A.; Szyniarowski, P.; Høyer-Hansen, M.; Povlsen, L.K.; Nylandsted, J.; Larsen, J.E. BAMLET activates a lysosomal cell death program in cancer cells. Mol. Cancer Ther. 2010, 9, 24–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mossberg, A.K.; Wullt, B.; Gustafsson, L.; Månsson, W.; Ljunggren, E.; Svanborg, C. Bladder cancers respond to intravesical instillation of (HAMLET human α-lactalbumin made lethal to tumor cells). Int. J. Cancer 2007, 121, 1352–1359. [Google Scholar] [CrossRef] [PubMed]
- Fischer, W.; Gustafsson, L.; Mossberg, A.-K.; Gronli, J.; Mork, S.; Bjerkvig, R.; Svanborg, C. Human α-lactalbumin made lethal to tumor cells (HAMLET) kills human glioblastoma cells in brain xenografts by an apoptosis-like mechanism and prolongs survival. Cancer Res. 2004, 64, 2105–2112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Zhang, J.; Gao, H.; Vieth, E.; Bae, K.-H.; Zhang, Y.-P.; Lee, S.-J.; Raikwar, S.; Gardner, T.A.; Hutchins, G.D. Transcriptional targeting modalities in breast cancer gene therapy using adenovirus vectors controlled by α-lactalbumin promoter. Mol. Cancer Ther. 2005, 4, 1850–1859. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Coope, H.; Grant, S.; Ma, A.; Ley, S.C.; Harhaj, E.W. ABIN1 protein cooperates with TAX1BP1 and A20 proteins to inhibit antiviral signaling. J. Biol. Chem. 2011, 286, 36592–36602. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.B.; Shembade, N.; Parvatiyar, K.; Balachandran, S.; Harhaj, E.W. TAX1BP1 restrains virus-induced apoptosis by facilitating itch-mediated degradation of the mitochondrial adaptor MAVS. Mol. Cell. Biol. 2017, 37, e00422-16. [Google Scholar] [CrossRef] [Green Version]
- Pisanu, S.; Ghisaura, S.; Pagnozzi, D.; Falchi, G.; Biosa, G.; Tanca, A.; Roggio, T.; Uzzau, S.; Addis, M.F. Characterization of sheep milk fat globule proteins by two-dimensional polyacrylamide gel electrophoresis/mass spectrometry and generation of a reference map. Int. Dairy J. 2012, 24, 78–86. [Google Scholar] [CrossRef]
- Alfadda, A.A.; Benabdelkamel, H.; Masood, A.; Moustafa, A.; Sallam, R.; Bassas, A.; Duncan, M. Proteomic analysis of mature adipo cytes from obese patients in relation to aging. Exp. Gerontol. 2013, 48, 1196–1203. [Google Scholar] [CrossRef] [PubMed]
- Benabdelkamel, H.; Masood, A.; Alanazi, I.O.; Alzahrani, D.A.; Alrabiah, D.K.; AlYahya, S.A.; Alfadda, A.A. Proteomic profiling comparing the effects of different heat treatments on camel (Camelus dromedarius) milk whey proteins. Int. J. Mol. Sci. 2017, 18, 721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfadda, A.A.; Benabdelkamel, H.; Masood, A.; Jammah, A.A.; Ekhzaimy, A.A. Differences in the plasma proteome of patients with hypothyroidism before and after thyroid hormone replacement: A proteomic analysis. Int. J. Mol. Sci. 2018, 19, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass spectrometric sequencing of proteins from silver-stained polyacrylamide gels. Anal. Chem. 1996, 68, 850–858. [Google Scholar] [CrossRef]
- Benabdelkamel, H.; Masood, A.; Almidani, G.M.; Alsadhan, A.A.; Bassas, A.F.; Duncan, M.W.; Alfadda, A.A. Mature adipocyte proteome reveals differentially altered protein abundances between lean, overweight and morbidly obese human subjects. Mol. Cell. Endocrinol. 2015, 401, 142–154. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot Number | Accession Number | Mascot ID | Protein Name | Function | Fold Change S/W |
|---|---|---|---|---|---|
| 1346 | P00710 | LALBA_CAMDR | A-lactalbumin | - | 2.8 |
| 1583 | Q3SZV0 | TTC36_BOVIN | Tetratricopeptide repeat protein 36 | - | 2.6 |
| 1662 | Q2KJE0 | TAXB1_BOVIN | Tax1-binding protein 1 homolog | Binding Protein | 2.5 |
| 1613 | P79385 | MFGE8_PIG | Lactadherin | Cell adhesion | 2.3 |
| 1004 | A4FUZ6 | HSDL2_BOVIN | Hydroxysteroid dehydrogenase-like protein 2 | Enzyme | 2 |
| 1001 | Q95114 | MFGE8_BOVIN | Lactadherin | Cell adhesion | 2 |
| 1648 | P48818 | ACADV_BOVIN | Very long-chain specific acyl-CoA dehydrogenase, mitochondrial | Enzyme | 1.6 |
| 1407 | Q95L54 | ANXA8_BOVIN | Annexin A8 | Cell membrane | 1.6 |
| 1155 | A4FUZ6 | HSDL2_BOVIN | Hydroxysteroid dehydrogenase-like protein 2 | Enzyme | 1.6 |
| 1078 | P79385 | MFGE8_PIG | Lactadherin | Cell adhesion | 1.6 |
| 1676 | Q3SZN0 | SEPT6_BOVIN | Septin-6 | Cell membrane | 1.5 |
| 1460 | Q9TT91 | MKRN1_MACEU | E3 ubiquitin-protein ligase makorin-1 | Enzyme | 1.5 |
| 1429 | P10173 | FUMH_PIG | Fumarate hydratase, mitochondrial | Enzyme | 1.5 |
| 1416 | Q3SZV0 | TTC36_BOVIN | Tetratricopeptide repeat protein 36 | - | 1.5 |
| 1384 | P79385 | MFGM_PIG | Lactadherin | Cell adhesion | 1.5 |
| 1373 | P17290 | TPH1_RABIT | Tryptophan 5-hydroxylase 1 | Enzyme | 1.5 |
| 1235 | P79385 | MFGE8_PIG | Lactadherin | Cell adhesion | 1.5 |
| 1181 | P79385 | MFGE8_PIG | Lactadherin | Cell adhesion | 1.5 |
| 1072 | P33545 | DSC2_BOVIN | Desmocollin-2 | Cell membrane | 1.5 |
| 1066 | Q3MHZ7 | GPI8_BOVIN | GPI-anchor transamidase | Enzyme | 1.5 |
| 1063 | P79385 | MFGE8_PIG | Lactadherin | Cell membrane | 1.5 |
| 1056 | Q148H8 | K2C72_BOVIN | Keratin, type II cytoskeletal 72 | Cell membrane | 1.5 |
| 1054 | P26234 | VINC_PIG | Vinculin | Cell membrane | 1.5 |
| 1040 | Q3MHR7 | ARPC2_BOVIN | Actin-related protein 2/3 complex subunit 2 | Cell membrane | 1.5 |
| 1039 | Q9MYY3 | VKGC_DELLE | Vitamin K-dependent gamma-carboxylase | Enzyme | 1.5 |
| 1026 | Q148H8 | K2C72_BOVIN | Keratin, type II cytoskeletal 72 | Cell membrane | 1.5 |
| 1011 | Q3MHZ7 | GPI8_BOVIN | GPI-anchor transamidase | Enzyme | 1.5 |
| 1009 | Q3MHZ7 | GPI8_BOVIN | GPI-anchor transamidase | Enzyme | 1.5 |
| 1005 | P79385 | MFGE8_PIG | Lactadherin | Cell adhesion | 1.5 |
| 396 | P79385 | MFGE8_PIG | Lactadherin | Cell adhesion | 1.5 |
| 1639 | P79385 | MFGE8_PIG | Lactadherin | Cell adhesion | 1.4 |
| 898 | P20414 | TIMPI_BOVIN | Metalloproteinase inhibitor 1 | Enzyme | −1.4 |
| 1423 | Q3SZV0 | TTC36_BOVIN | Tetratricopeptide repeat protein 36 | Binding Protein | −1.5 |
| 1210 | Q0IIF2 | EI2BA_BOVIN | Translation initiation factor eIF-2B subunit alpha | Binding Protein | −1.5 |
| 1225 | Q9TUM0 | TRFL_CAMDR | Lactotransferrin | Enzyme | −1.6 |
| 699 | A1L520 | ARFG2_BOVIN | ADP-ribosylation factor GTPase-activating protein 2 | Binding Protein | −1.9 |
| 358 | Q9TUM0 | TRFL_CAMDR | Lactotransferrin | Enzyme | −1.9 |
| 858 | P10950 | HIOM_BOVIN | Acetyls erotonin O-methyl transferase | Enzyme | −2.2 |
| 1103 | P0CB32 | HS71L_BOVIN | Heat shock 70 kDa protein 1-like | Immune system | −2.4 |
| 871 | P18892 | BT1A1_BOVIN | Butyrophilin subfamily 1 member A1 | Enzyme | −2.5 |
| 821 | Q4TVR5 | DUSTY_BOVIN | Dual serine/threonine and tyrosine protein kinase | Enzyme | −2.5 |
| 916 | P26234 | VINC_PIG | Vinculin | Cell membrane | −4.8 |
| 917 | Q9TUM0 | TRFL_CAMDR | Lactotransferrin | Enzyme | −5.8 |
| 864 | Q9TUM0 | TRFL_CAMDR | Lactotransferrin | Enzyme | −15.8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sabha, B.H.; Masood, A.; Alanazi, I.O.; Alfadda, A.A.; Almehdar, H.A.; Benabdelkamel, H.; Redwan, E.M. Comparative Analysis of Milk Fat Globular Membrane (MFGM) Proteome between Saudi Arabia Camelus dromedary Safra and Wadha Breeds. Molecules 2020, 25, 2146. https://doi.org/10.3390/molecules25092146
Sabha BH, Masood A, Alanazi IO, Alfadda AA, Almehdar HA, Benabdelkamel H, Redwan EM. Comparative Analysis of Milk Fat Globular Membrane (MFGM) Proteome between Saudi Arabia Camelus dromedary Safra and Wadha Breeds. Molecules. 2020; 25(9):2146. https://doi.org/10.3390/molecules25092146
Chicago/Turabian StyleSabha, Bassam H., Afshan Masood, Ibrahim O. Alanazi, Assim A. Alfadda, Hussein A. Almehdar, Hicham Benabdelkamel, and Elrashdy M. Redwan. 2020. "Comparative Analysis of Milk Fat Globular Membrane (MFGM) Proteome between Saudi Arabia Camelus dromedary Safra and Wadha Breeds" Molecules 25, no. 9: 2146. https://doi.org/10.3390/molecules25092146
APA StyleSabha, B. H., Masood, A., Alanazi, I. O., Alfadda, A. A., Almehdar, H. A., Benabdelkamel, H., & Redwan, E. M. (2020). Comparative Analysis of Milk Fat Globular Membrane (MFGM) Proteome between Saudi Arabia Camelus dromedary Safra and Wadha Breeds. Molecules, 25(9), 2146. https://doi.org/10.3390/molecules25092146