
Statistical Based Bioprocess Design for Improved Production of Amylase from Halophilic Bacillus sp. H7 Isolated from Marine Water


 , ,
, , 
 and
and 
Abstract
:
1. Introduction
2. Results
2.1. Screening and Quantitative Estimation of Amylase Production by Halophilic Isolates
2.2. Identification of Potent Isolate
2.2.1. Phenotypic Characterization
2.2.2. Genetic Identification of Isolate Using 16S rRNA Sequencing
2.3. Influence of Physico-Chemical Media Variables on Amylase Production
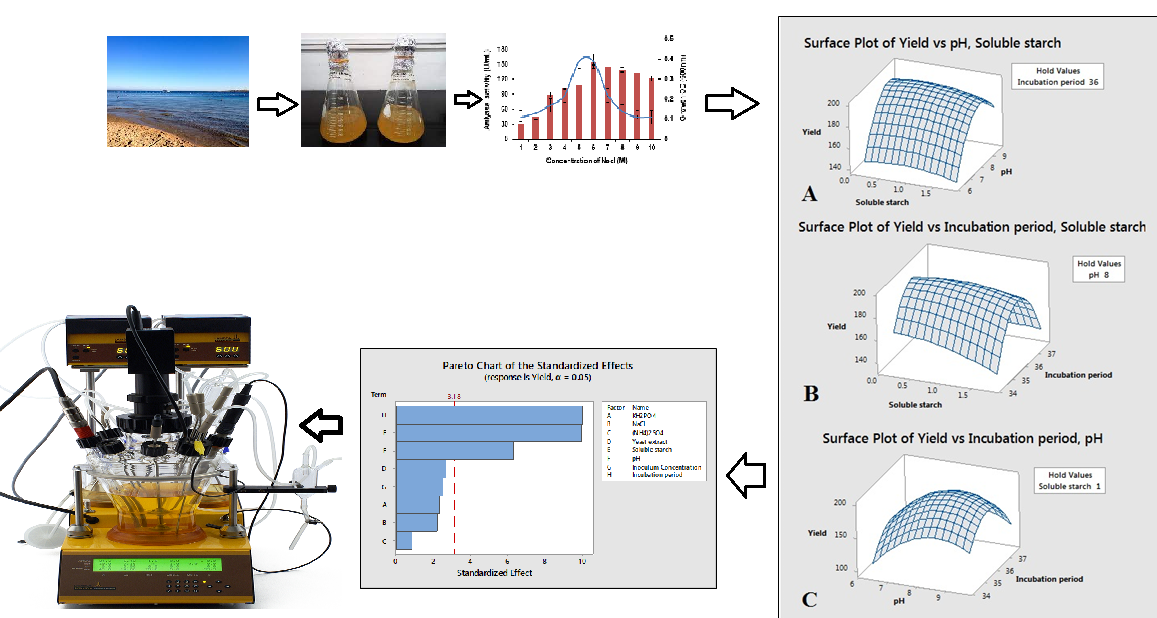
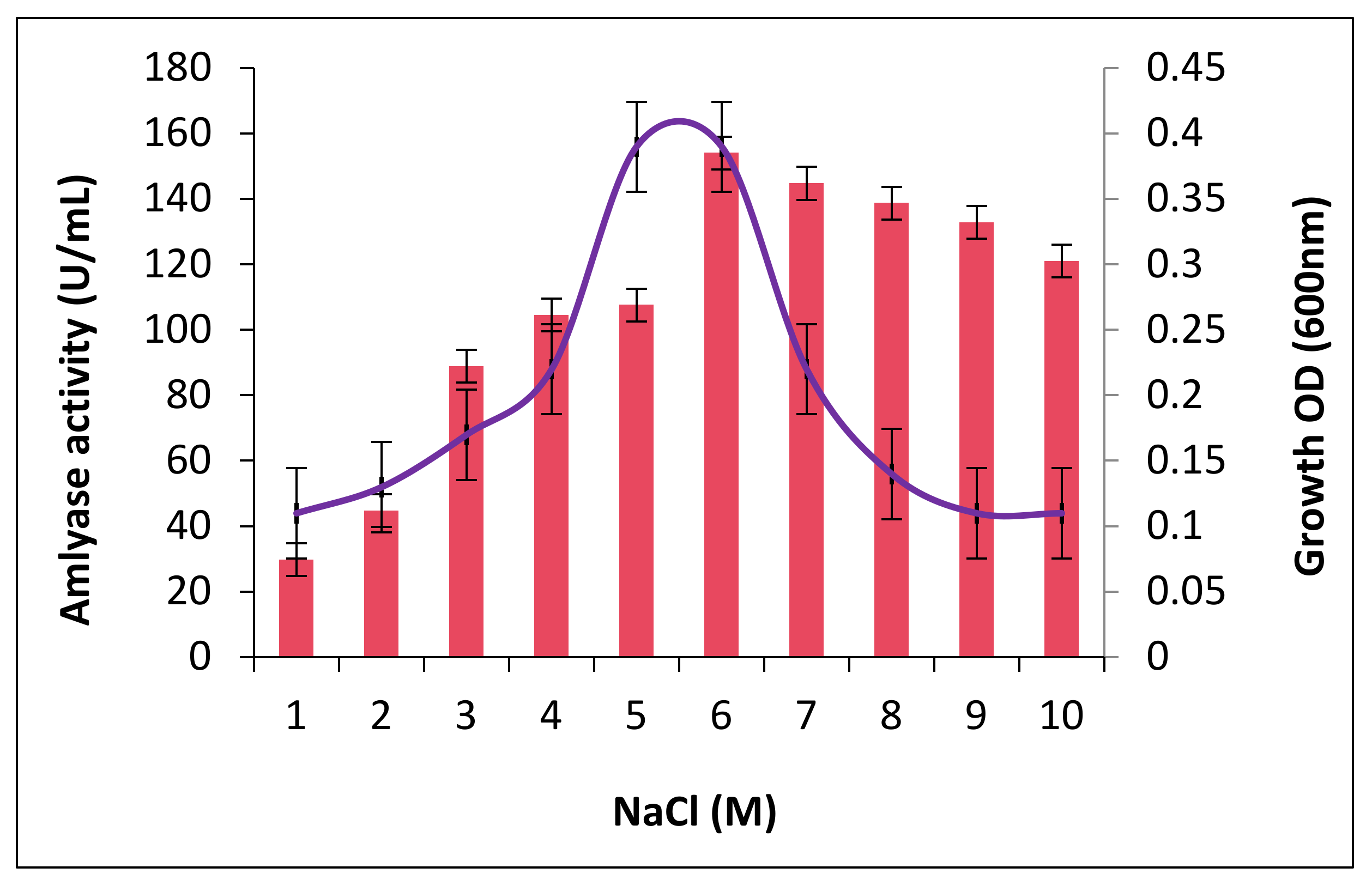
2.4. The Effect of Salt Concentrations on Growth and Amylase Production
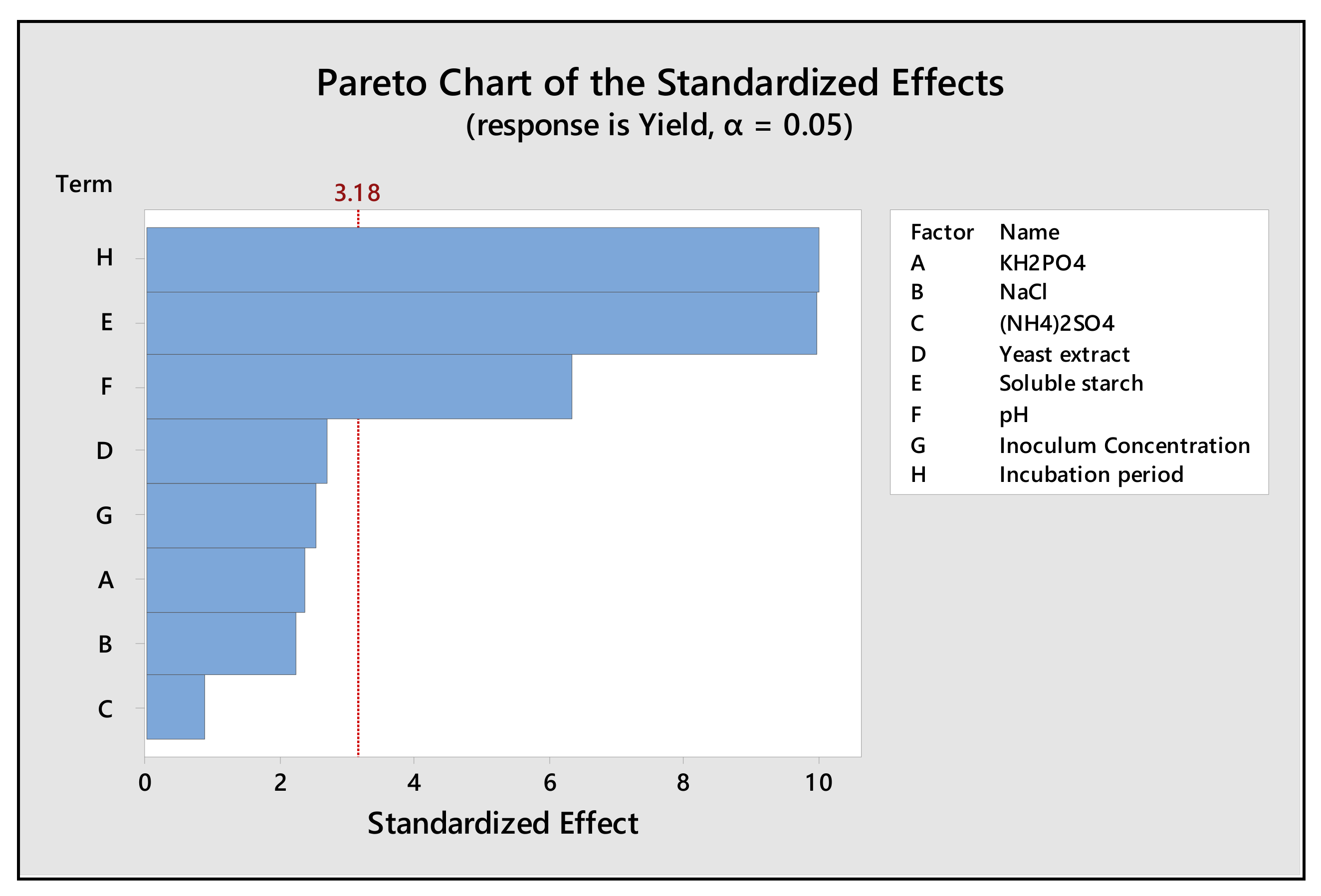
2.5. Evaluation of Significant Production Media Variable by Plackett-Burman Design (PBD)
2.6. Estimation of Optimization Concentration of Significant Variables Using Central Composite Design (CCD) of Response Surface Methodology (RSM)
+ 5.64 (E·H) − 0.729 (F * H)
2.7. Model Validation and Scale-Up Using a Laboratory-Scale (5L) Bioreactor
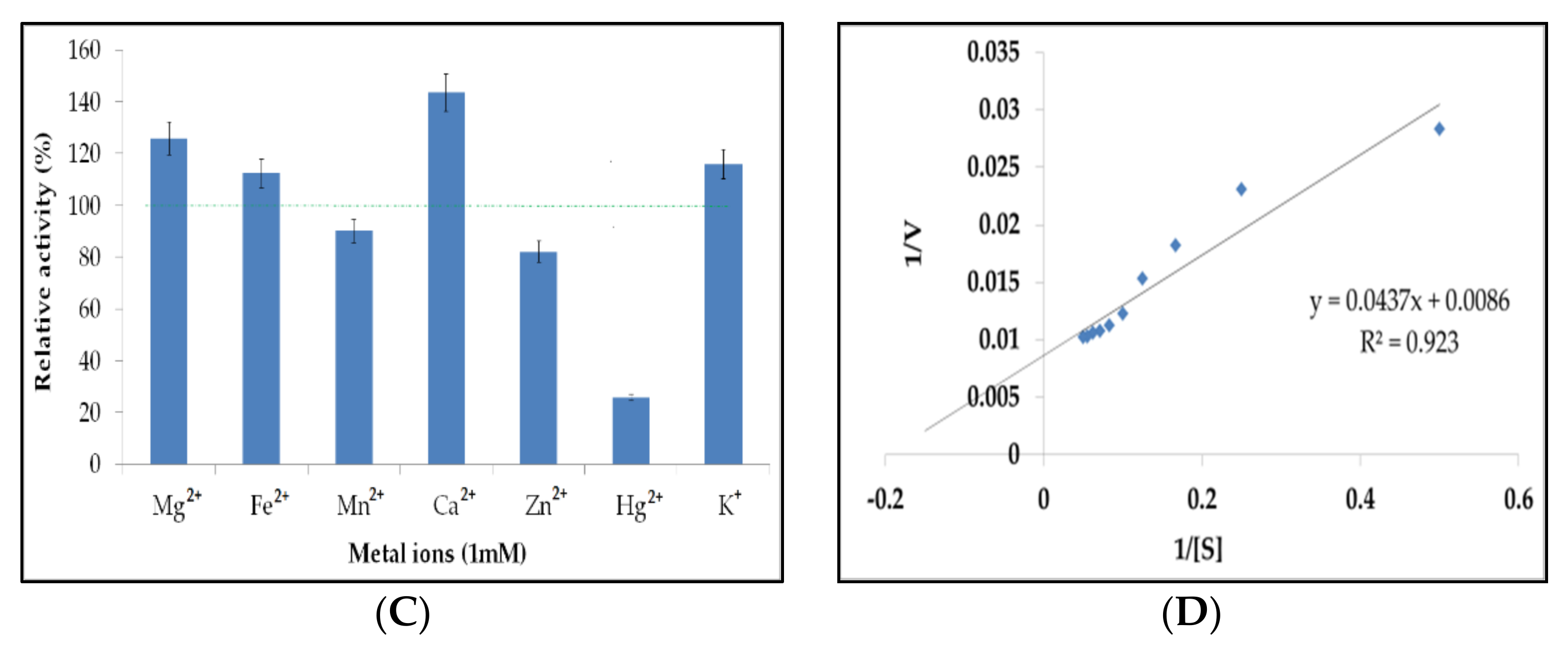
2.8. Characterization and Determination of Kinetic Parameters of Amylase from Bacillus sp. H7
3. Discussion
4. Materials and Methods
4.1. Sample Collection, Screening and Quantitative Estimation of Amylase Production by Halophilic Isolates
4.2. Characterization of the Potent Isolate
4.2.1. Phenotypic Characterization
4.2.2. Molecular Identification of Isolate Using 16S rRNA Sequencing
4.3. Influence of Physico-Chemical Media Variables of Amylase Production
4.4. Statistical Analyses
4.5. Effect of Salinity on Growth and Amylase Production
4.6. Evaluation of Significant Variable of Production Media by Plackett–Burman Design (PBD)
4.7. Estimation and Optimization Concentration of Significant Variables Using Central Composite Design (CCD) of Response Surface Methodology (RSM)
- Yi—predicted response, XiXj—input variables (influence the response variable Y),
- β0—constant, βi—ith linear coefficient,
- βii—quadratic coefficient, βij—ijth interaction coefficient.
4.8. Model Validation and Scale-Up Using a Laboratory-Scale (5L) Bioreactor
4.9. Software and Data Analysis
4.10. Characterization and Determination of Kinetic Parameters of Crude Amylase from Bacillus sp. H7
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Cherry, H.M.; Hossain, M.T.; Anwar, M.N. Extracellular glucoamylase from the isolate Aspergillus fumigatus. Pak. J. Biol. Sci. 2004, 7, 1988–1992. [Google Scholar] [CrossRef] [Green Version]
- Pandey, G.; Munguambe, D.; Tharmavaram, M.; Rawtani, D.; Agrawal, Y. Halloysite nanotube—An efficient ‘nano-support’ for the immobilization of α-amylase. Appl. Clay. Sci. 2017, 136, 184–191. [Google Scholar] [CrossRef]
- Asrat, B.; Girma, A. Isolation, production and characterization of amylase enzyme using the isolate Aspergillus niger FAB-211. Int. J. Biotechnol. Mol. Biol. Res. 2018, 9, 7–14. [Google Scholar]
- El Enshasy, H.; Abdel Fattah, Y.; Othman, N.Z. Amylases. In Bioprocessing Technologes in Integrated Biorefinery from Production of Biofuels, Biochemicals, and Biopolymers from Biomass; Yang, S.-T., El Enshasy, H.A., Thongchul, N., Eds.; John Wiley & Sons: New Jersey, NJ, USA, 2013; pp. 111–130. ISBN 978-0-470-54195-1. [Google Scholar]
- Simair, A.A.; Qureshi, A.S.; Khushk, I.; Ali, C.H.; Lashari, S.; Bhutto, M.A.; Mangrio, G.S.; Lu, C. Production and partial characterization of α-amylase enzyme from Bacillus sp. BCC 01-50 and potential applications. BioMed Res. Int. 2017, 2017, 9173040. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Karan, R.; Kapoor, S.; Singh, S.P.; Khare, S.K. Screening and isolation of halophilic bacteria producing industrially important enzymes. Braz. J. Microbiol. 2012, 43, 1595–1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vijayaraghavan, P.; Kalaiyarasi, M.; Vincent, S.G.P. Cow dung is an ideal fermentation medium for amylase production in solid-state fermentation by Bacillus cereus. J. Gen. Eng. Biotechnol. 2015, 13, 111–117. [Google Scholar] [CrossRef] [Green Version]
- Sreena, C.P.; Sebastian, D. Augmented cellulase production by Bacillus subtilis strain MU S1 using different statistical experimental designs. J. Gen. Eng. Biotechnol. 2018, 16, 9–16. [Google Scholar] [CrossRef]
- Elsayed, E.A.; Omar, H.G.; Galil, S.A.; El-Enshasy, H.A. Optimization of fed-batch cultivation for extracellular α-amylase production by Bacillus amyloliquefaciens in submerged culture. J. Sci. Ind. Res. 2016, 75, 480–486. [Google Scholar]
- Nurkhasanah, S.; Nur Afifah, Z.; Al Husna, S.N.; Widowati, R.; El Enshasy, H.; Dailin, D.J. Metagenomic-based approach for the analysis of yeast diversity associated with amylase production in Lai (Durio kutejensis). J. Pure Appl. Microbiol. 2021, 15, 75–90. [Google Scholar] [CrossRef]
- Aullybux, A.; Puchooa, D. α-Amylase production on low-cost substrates by Naxibacter sp. isolated from Mauritian soils. Br. Microbiol. Res. J. 2013, 3, 478–491. [Google Scholar] [CrossRef]
- Mahapatra, P.D.; Mishra, B.B. Isolation and molecular identification of an extreme alkali-tolerant Pseudomonas mendocina 2E from brackish water of Chilica Lake. Cibte. J. Bio Proto 2014, 3, 11–18. [Google Scholar]
- Huma, T.; Maryam, A.; Rehman, S.; Tahir ul qamar, M.; Shaheen, T.; Haque, A.; Shaheen, B. Phylogenetic and comparative sequence analysis of thermostable alpha amylases of kingdom Archea, Prokaryotes and Eukaryotes. Bioinformation 2014, 10, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Sonawane, M.S.; Chaudhary, R.D.; Shouche, Y.S.; Sayyed, R.Z. Insect gut bacteria: A novel source for siderophore production. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2016, 88, 567–572. [Google Scholar] [CrossRef]
- Saha, S.P.; Mazumdar, D. Optimization of process parameter for alpha-amylase produced by Bacillus cereus amy3 using one factor at a time (OFAT) and central composite rotatable (CCRD) design based response surface methodology (RSM). Biocatal. Agric. Biotechnol. 2019, 19, 101168. [Google Scholar] [CrossRef]
- Khusro, A.; Barathikannan, K.; Aarti, C.; Agastian, P. Optimization of thermo-alkali stable amylase production and biomass yield from Bacillus sp. under submerged cultivation. Fermentation 2017, 3, 7. [Google Scholar] [CrossRef]
- Abdel Wahab, W.A.; Ahmed, S.A. Response surface methodology for production, characterization and application of solvent, salt and alkali-tolerant alkaline protease from isolated fungal strain Aspergillus niger WA 2017. Int. J. Biol. Macromol. 2018, 115, 447–458. [Google Scholar] [CrossRef]
- Elmansy, E.A.; Asker, M.; El-Kady, E.M.; Hassanein, S.M.; Fawkia, M. Production and optimization of α-amylase from thermo-halophilic bacteria isolated from different local marine environments. Bull. Natl. Res. Cent. 2018, 42, 31. [Google Scholar] [CrossRef] [Green Version]
- Balakrishnan, D.; Shilpa Shaji, V.S. Molecular Identification and Optimization of Amylase Producing Bacillus gingshengii SNB12 Using Response Surface Methodology. Int. J. Adv. Biotechnol. Res. 2020, 10, 1–16. [Google Scholar]
- Ojha, S.K.; Singh, P.K.; Mishra, S.; Pattnaik, R.; Dixit, S.; Verma, S.K. Response surface methodology based optimization and scale-up production of amylase from a novel bacterial strain, Bacillus aryabhattai KIIT BE-1. Biotechnol. Rep. 2020, 27, e00506. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.A.; Abdella, M.A.; El-Sherbiny, G.M.; Ibrahim, A.M.; El-Shamy, A.R.; Atalla, S.M. Application of one-factor-at-a-time and statistical designs to enhance α-amylase production by a newly isolate Bacillus subtilis strain-MK1. Biocatal. Agric. Biotechnol. 2019, 22, 101397. [Google Scholar] [CrossRef]
- Hashemi, M.; Razavi, S.H.; Shojaosadati, S.A.; Mousavi, S.M.; Khajeh, K.; Safari, M. Development of a solid-state fermentation process for production of an alpha amylase with potentially interesting properties. J. Biosci. Bioeng. 2010, 110, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Quan, S.; Liu, D.; Ma, H.; Li, F.; Zhou, F.; Chen, G. Purification and characterization of a novel α-amylase from a newly isolated Bacillus methylotrophicus strain P11-2. Proc. Biochem. 2014, 49, 47–53. [Google Scholar] [CrossRef]
- Kalpana, B.J.; Pandian, S.K. Halotolerant, acid-alkalistable, chelator resistant and raw starch digesting α-amylase from a marine bacterium Bacillus subtilis S8–18. J. Basic Microbiol. 2014, 54, 802–811. [Google Scholar] [CrossRef]
- Carvalho, R.V.D.; Côrrea, T.L.R.; Silva, J.C.M.D.; Mansur, L.R.C.D.O.; Martins, M.L.L. Properties of an amylase from thermophilic Bacillus sp. Braz. J. Microbiol. 2008, 39, 102–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, L.; Irorere, V.U.; Marchant, R.; Banat, I.M. Marine derived biosurfactants: A vast potential future resource. Biotechnol. Lett. 2018, 40, 1441–1457. [Google Scholar] [CrossRef] [Green Version]
- Nayak, N.S.; Thacker, S.C.; Tipre, D.R.; Dave, S.R. Bacillus pumilus—A marine bacteria: Unexplored source for potential biosurfactant production. Biosci. Biotechnol. Res. Comm. 2020, 13, 180–187. [Google Scholar]
- Rehman, A.; Saeed, A.; Asad, W.; Kiran, T.; Baloch, M.N.; Eijaz, S. Optimization of physicochemical parameters for maximum amylase production by indigenously isolated Bacillus cereus AS2 strain. Pak. J. Pharm. Sci. 2019, 32, 889–894. [Google Scholar] [PubMed]
- Aiyer, P.V. Amylases and their applications. Afr. J. Biotechnol. 2005, 4, 1525–1529. [Google Scholar]
- Abdel-Fattah, Y.R.; Soliman, N.A.; El-Toukhy, N.M.; El-Gendi, H.; Ahmed, R.S. Production, purification, and characterization of thermostable α-amylase produced by Bacillus licheniformis isolate AI20. J. Chem. 2013, 2013, 673173. [Google Scholar] [CrossRef] [Green Version]
- Ghorbel, R.E.; Maktouf, S.; Massoud, E.B.; Bejar, S.; Chaabouni, S.E. New thermostable amylase from Bacillus cohnii US147 with a broad pH applicability. Appl. Biochem. Biotechnol. 2009, 157, 50. [Google Scholar] [CrossRef]
- Balkan, B.; Ertan, F. Production of α-Amylase from Penicillium chrysogenum under solid-state fermentation by using some agricultural by-products. Food Technol. Biotechnol. 2007, 45, 439–442. [Google Scholar]
- Mishra, T.; Ahluwalia, S.; Joshi, M. Isolation and medium optimization for amylase producing bacterial strain isolated from potato field of Bhatinda, Punjab, India. Eur. J. Exp. Biol. 2014, 4, 588–594. [Google Scholar]
- Nithya, K.; Muthukumar, C.; Dhanasekaran, D.; Kadaikunnan, S.; Alharbi, N.S.; Khaled, J.M.; Thajuddin, N. Production, optimization and partial characterization of thermostable and alkaline amylase from Bacillus licheniformis KSU-6. Int. J. Agric. Biol. 2016, 18, 1188–1194. [Google Scholar] [CrossRef]
- Jadhav, H.P.; Sonawane, M.S.; Khairnar, M.H.; Sayyed, R.Z. Production of alkaline protease by rhizospheric Bacillus cereus HP_RZ17 and Paenibacillus xylanilyticus HP_RZ19. Environ. Sustain. 2020, 3, 5–13. [Google Scholar] [CrossRef]
- Saxena, R.; Dutt, K.; Agarwal, L.; Nayyar, P. A highly thermostable and alkaline amylase from a Bacillus sp. Bioresour. Technol. 2007, 98, 260–265. [Google Scholar] [CrossRef]
- Božić, N.; Ruiz, J.; Lόoez-Santin, J.; Vujčić, Z. Optimization of the growth and alpha-amylase production of Bacillus subtilis IP 5832 in shake flask and laboratory fermenter batch cultures. J. Serb. Chem. Soc. 2011, 76, 965–972. [Google Scholar] [CrossRef]
- Al-Johani, N.B.; Al-seeni, M.N.; Ahmed, Y.M. Optimization of alkaline α-amylase production by thermophilic Bacillus subtilis. Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 288–301. [Google Scholar] [CrossRef] [Green Version]
- Hillier, P.; Wase, D.A.J.; Emery, A.N. Production of α-amylase (E.C.3.2.1.1.) by Bacillus amyloliquefaciens in batch and continuous culture using a defined synthetic medium. Biotechnol. Lett. 1996, 18, 795–800. [Google Scholar] [CrossRef]
- Habicher, T.; John, A.; Scholl, N.; Daub, A.; Klein, T.; Philip, P.; Büchs, J. Introducing substrate limitations to overcome catabolite repression in a protease producing Bacillus licheniformis strain using membrane-based fed-batch shake flasks. Biotechnol. Bioeng. 2019, 116, 1326–1340. [Google Scholar] [CrossRef]
- Teodoro, C.E.; Martins, M.L. Culture conditions for the production of thermostable amylase by Bacillus sp. Braz. J. Microbiol. 2000, 31, 298–302. [Google Scholar] [CrossRef]
- Blanco, A.S.; Durive, O.P.; Pérez, S.B.; Montes, Z.D.; Guerra, N.P. Simultaneous production of amylases and proteases by Bacillus subtilis in brewery wastes. Braz. J. Microbiol. 2016, 47, 665–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poddar, A.; Ghara, T.K.; Jana, S.C. Critical process parameters optimization for hyperthermostable βamylase production by Bacillus subtilis DJ5 using response surface methodology. Acta Sci. Biol. Sci. 2014, 36, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Shofiyah, S.S.; Yuliani, D.; Widya, N.; Sarian, F.D.; Puspasari, F.; Radjasa, O.K.; Natalia, D. Isolation, expression, and characterization of raw starch degrading α-amylase from a marine lake Bacillus megaterium NL3. Heliyon 2020, 6, e05796. [Google Scholar] [CrossRef] [PubMed]
- Alonazi, M.; Karray, A.; Badjah-Hadj-Ahmed, A.Y.; Ben Bacha, A. Alpha Amylase from Bacillus pacificus Associated with Brown Algae Turbinaria ornata: Cultural Conditions, Purification, and Biochemical Characterization. Processes 2021, 9, 16. [Google Scholar] [CrossRef]
- Ghollasi, M.; Khajeh, K.; Naderi-Manesh, H.; Ghasemi, A. Engineering of a Bacillus alpha-amylase with improved thermostability and calcium independency. Appl. Biochem. Biotechnol. 2010, 162, 444–459. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, S.O.; Srivastava, P.K.; Rathore, P.R.; Pandey, A.K. Amylases: A prospective enzyme in the field of biotechnology. J. Appl. Biosci. 2015, 41, 1–8. [Google Scholar]
- Thebti, W.; Riahi, Y.; Gharsalli, R.; Belhadj, O. Screening and characterization of thermo-active enzymes of biotechnological interest produced by thermophilic Bacillus isolated from hot springs in Tunisia. Acta Biochim. Pol. 2016, 63, 581–587. [Google Scholar] [CrossRef]
- Kunamneni, A.; Kumar, K.S.; Singh, S. Response surface methodological approach to optimize the nutritional parameters for enhanced production of-amylase in solid state fermentation by Thermomyces lanuginosus. Afr. J. Biotechnol. 2005, 4, 708–716. [Google Scholar]
- Sambrook, J.; Russell, D.W. Molecular Cloning-Sambrook and Russel, 3rd ed.; Cold Springs Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Parker, R.E. Continuous distribution: Tests of significance. In Introductory Statistics for Biology, 2nd ed.; Parker, R.E., Ed.; Cambridge University Press: London, UK, 1979; pp. 18–42. [Google Scholar]
- Deepa, T.; Gangwane, A.K.; Sayyed, R.Z.; Jadhav, H.P. Optimization and scale-up of laccase production by Bacillus sp. BAB-4151 isolated from the waste of the soap industry. Environ. Sustain. 2020, 3, 471–479. [Google Scholar] [CrossRef]
- Jadhav, H.P.; Sayyed, R.Z.; Shaikh, S.S.; Bhamre, H.M.; Sunita, K.; El Enshasy, H.A. Statistically designed bioprocess for enhanced production of alkaline protease in Bacillus cereus HP_RZ17. J. Sci. Ind. Res. 2020, 79, 491–498. [Google Scholar]
- Bhamare, H.M.; Jadhav, H.P.; Sayyed, R.Z. Statistical optimization for enhanced production of extracellular laccase from Aspergillus sp. HB_RZ4 isolated from bark scrapping. Environ. Sustain. 2018, 1, 159–166. [Google Scholar] [CrossRef]
- Plackett, R.L.; Burman, J.P. The design of optimum multifactorial experiments. Biometrika 1946, 33, 305–325. [Google Scholar] [CrossRef]
- Box, G.E.; Wilson, K.B. On the experimental attainment of optimum conditions. J. R. Stat. Soc. Ser. B 1951, 13, 1–38. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| INC (h) | Amylase (U/mL) | pH | Amylase (U/mL) | Temp (°C) | Amylase (U/mL) | CS (1%) | Amylase (U/mL) | NS | Amylase (U/mL) | INO (1%) | Amylase (U/mL) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 0.00 | 4 | 56.89 ± 5.3 | 20 | 90.1 ± 5 | Glucose | 144.64 ± 2.1 | Peptone | 132.6 ± 3.2 | 0.50% | 126.53 ± 0.5 |
| 6 | 59.81 ± 7.5 | 5 | 87.53 ± 6.7 | 25 | 109.8 ± 4.3 | Starch | 149.1 ± 2.1 | Casein | 121.24 ± 5.0 | 1.00% | 133.17 ± 0.6 |
| 12 | 87.24 ± 6.6 | 6 | 113.61 ± 0.6 | 30 | 121.24 ± 2.1 | Fructose | 129.81 ± 2.1 | Tryptone | 111.6 ± 9.0 | 1.50% | 140.03 ± 0.7 |
| 18 | 102.89 ± 1.0 | 7 | 128.89 ± 1.0 | 35 | 127.24 ± 2.1 | Sucrose | 121.24 ± 7.5 | Yeast ext. | 152.96 ± 3.2 | 2.00% | 132.6 ± 3.2 |
| 24 | 117.24 ± 2.8 | 8 | 133.53 ± 1.2 | 40 | 122.81 ± 1.7 | Lactose | 109.17 ± 0.6 | Urea | 88.24 ± 7.0 | ||
| 30 | 125.89 ± 1.0 | 9 | 131.53 ± 1.5 | 45 | 112.8 ± 5.5 | Maltose | 127.81 ± 4.0 | NH4NO3 | 75.89 ± 7.5 | ||
| 36 | 130.53 ± 2.0 | 10 | 125.17 ± 1.5 | 50 | 101.24 ± 4.9 | Dextrose | 97.1 ± 4.3 | NH4CL2 | 60.96 ± 4.4 | ||
| 42 | 128.89 ± 2.7 | 11 | 119.23 ± 4.7 | ||||||||
| 48 | 125.53 ± 1.5 | 12 | 111.17 ± 4.0 | ||||||||
| 54 | 120.53 ± 1.5 | 13 | 98.24 ± 5.0 | ||||||||
| 60 | 113.89 ± 4.0 | 14 | 93.83 ± 7.2 | ||||||||
| 66 | 107.53 ± 4.7 | ||||||||||
| 72 | 100.53 ± 8.4 | ||||||||||
| 78 | 98.89 ± 7.0 | ||||||||||
| 84 | 95.17 ± 7.5 | ||||||||||
| 90 | 92.89 ± 6.2 | ||||||||||
| 96 | 90.24 ± 7.5 |
| Run | A | B | C | D | E | F | G | H | Amylase Activity (U/mL) | |
|---|---|---|---|---|---|---|---|---|---|---|
| Predicted | Experimental | |||||||||
| 1 | +1 | −1 | +1 | −1 | −1 | −1 | +1 | +1 | 135.60 | 133.49 ± 1.22 |
| 2 | +1 | +1 | −1 | +1 | −1 | −1 | −1 | +1 | 122.86 | 121.89 ± 0.48 |
| 3 | −1 | +1 | +1 | −1 | +1 | −1 | −1 | −1 | 130.90 | 128.60 ± 0.68 |
| 4 | +1 | −1 | +1 | +1 | −1 | +1 | −1 | −1 | 118.38 | 119.17 ± 0.76 |
| 5 | +1 | +1 | −1 | +1 | +1 | −1 | +1 | −1 | 127.68 | 128.65 ± 0.79 |
| 6 | +1 | +1 | +1 | −1 | +1 | +1 | −1 | +1 | 158.06 | 160.36 ± 1.18 |
| 7 | −1 | +1 | +1 | +1 | −1 | +1 | +1 | −1 | 123.50 | 122.71 ± 0.34 |
| 8 | −1 | −1 | +1 | +1 | +1 | −1 | +1 | +1 | 154.32 | 156.43 ± 0.77 |
| 9 | −1 | −1 | −1 | +1 | +1 | +1 | −1 | +1 | 163.41 | 161.30 ± 0.68 |
| 10 | +1 | −1 | −1 | −1 | +1 | +1 | +1 | −1 | 149.51 | 148.54 ± 1.04 |
| 11 | −1 | +1 | −1 | −1 | −1 | +1 | +1 | +1 | 149.81 | 150.60 ± 1.53 |
| 12 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | 117.53 | 119.82 ± 0.82 |
| Term | Effect | Coef | SE Coef | t-Value | p-Value |
|---|---|---|---|---|---|
| Constant | 137.630 | 0.969 | 141.97 | 0.000 | |
| KH2PO4 | −4.563 | −2.281 | 0.969 | −2.35 | 0.100 |
| NaCl | −4.325 | −2.162 | 0.969 | −2.23 | 0.112 |
| (NH4)2SO4 | −1.673 | −0.836 | 0.969 | −0.86 | 0.452 |
| Yeast extract | −5.211 | −2.605 | 0.969 | −2.69 | 0.075 |
| Soluble starch | 19.367 | 9.684 | 0.969 | 9.99 | 0.002 * |
| pH | 12.301 | 6.150 | 0.969 | 6.34 | 0.008 * |
| Inoculum concentration | 4.879 | 2.440 | 0.969 | 2.52 | 0.086 |
| Incubation period | 19.429 | 9.715 | 0.969 | 10.02 | 0.002 * |
| Run Order | Type | Soluble Starch | pH | Incubation Period | Amylase Activity (U/mL) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Coded | Experimental | Coded | Experimental | Coded | Experimental | Predicted | Experimental | ||
| 1 | Factorial | −1 | 0.50 | −1 | 7.00 | −1 | 35.00 | 165.29 | 165.70 ± 0.64 |
| 2 | Factorial | 1 | 1.50 | −1 | 7.00 | −1 | 35.00 | 156.06 | 158.86 ± 0.29 |
| 3 | Factorial | −1 | 0.50 | 1 | 9.00 | −1 | 35.00 | 188.04 | 188.61 ± 0.97 |
| 4 | Factorial | 1 | 1.50 | 1 | 9.00 | −1 | 35.00 | 177.95 | 178.85 ± 1.05 |
| 5 | Factorial | −1 | 0.50 | −1 | 7.00 | 1 | 37.00 | 152.51 | 153.79 ± 0.95 |
| 6 | Factorial | 1 | 1.50 | −1 | 7.00 | 1 | 37.00 | 154.57 | 156.19 ± 0.64 |
| 7 | Factorial | −1 | 0.50 | 1 | 9.00 | 1 | 37.00 | 172.34 | 171.74 ± 1.62 |
| 8 | Factorial | 1 | 1.50 | 1 | 9.00 | 1 | 37.00 | 173.54 | 175.32 ± 0.75 |
| 9 | Axial | −1.68 | 0.16 | 0 | 8.00 | 0 | 36.00 | 194.70 | 194.78 ± 0.88 |
| 10 | Axial | 1.68 | 1.84 | 0 | 8.00 | 0 | 36.00 | 187.95 | 184.79 ± 0.20 |
| 11 | Axial | 0 | 1.00 | −1.68 | 6.32 | 0 | 36.00 | 147.45 | 144.88 ± 0.34 |
| 12 | Axial | 0 | 1.00 | 1.68 | 9.68 | 0 | 36.00 | 182.54 | 182.02 ± 1.51 |
| 13 | Axial | 0 | 1.00 | 0.0 | 8.00 | −1.68 | 34.32 | 158.41 | 156.69 ± 0.69 |
| 14 | Axial | 0 | 1.00 | 0.0 | 8.00 | 1.68 | 37.68 | 143.96 | 142.59 ± 0.99 |
| 15 | Central | 0 | 1.00 | 0.0 | 8.00 | 0.0 | 36.00 | 196.06 | 196.25 ± 1.22 |
| 16 | Central | 0 | 1.00 | 0.0 | 8.00 | 0.0 | 36.00 | 196.06 | 196.62 ± 0.95 |
| 17 | Central | 0 | 1.00 | 0.0 | 8.00 | 0.0 | 36.00 | 196.06 | 195.68 ± 1.02 |
| 18 | Central | 0 | 1.00 | 0.0 | 8.00 | 0.0 | 36.00 | 196.06 | 195.15 ± 0.35 |
| 19 | Central | 0 | 1.00 | 0.0 | 8.00 | 0.0 | 36.00 | 196.06 | 196.53 ± 0.79 |
| 20 | Central | 0 | 1.00 | 0.0 | 8.00 | 0.0 | 36.00 | 196.06 | 196.66 ± 1.09 |
| Source | DF | Coef | SE Coef | Adj SS | Adj MS | t-Value | f-Value | p-Value |
|---|---|---|---|---|---|---|---|---|
| Constant | 196.060 | 0.821 | 238.89 | 0.000 | ||||
| Model | 9 | 6785.68 | 753.96 | 186.20 | 0.000 | |||
| Soluble starch | 1 | −2.007 | 0.545 | 55.01 | 55.01 | −3.69 | 13.59 | 0.004 |
| pH | 1 | 10.431 | 0.545 | 1485.93 | 1485.93 | 19.16 | 366.97 | 0.000 |
| Incubation period | 1 | −4.298 | 0.545 | 252.30 | 252.30 | −7.89 | 62.31 | 0.000 |
| Soluble starch * soluble starch | 1 | −1.673 | 0.530 | 40.31 | 40.31 | −3.16 | 9.96 | 0.010 |
| pH * pH | 1 | −10.983 | 0.530 | 1738.31 | 1738.31 | −20.72 | 429.29 | 0.000 |
| Incubation period * incubation period | 1 | −15.866 | 0.530 | 3627.72 | 3627.72 | −29.93 | 895.90 | 0.000 |
| Soluble starch * pH | 1 | −0.217 | 0.711 | 0.38 | 0.38 | −0.30 | 0.09 | 0.767 |
| Soluble starch * incubation period | 1 | 2.822 | 0.711 | 63.69 | 63.69 | 3.97 | 15.73 | 0.003 |
| pH * incubation period | 1 | −0.729 | 0.711 | 4.25 | 4.25 | −1.02 | 1.05 | 0.330 |
| Lack-of-Fit | 5 | 38.64 | 7.73 | 20.88 | 0.002 | |||
| Pure error | 5 | 1.85 | 0.37 |
| Properties | Bacillus sp. H7 | Bacillus Methylotrophicus [23] | Bacillus sp. BCC 01-50 [5] | Bacillus Subtilis S8-18 [24] | Bacillus sp. SMIA-2 [25] |
|---|---|---|---|---|---|
| Production time (h) | 24 | 42 | 60 | 24 | 32 |
| Enzyme yield (U/mL) | 130.53 | 33.50 | 90 | 61 | 37 |
| Salt (NaCl) tolerance threshold (M) | 6.0 | 00 | 00 | 2.8 | 2.0 |
| pH range | 7.00–10.00 | 7.0–9.00 | 7.00–9.00 | 4.00–12.00 | 7.00–8.50 |
| pH optima | 8.0 | 7.0 | 9.0 | 9.00 | 8.0 |
| Temperature range (°C) | 30–70 | 40–70 | 60–70 | 30–40 | 40–50 |
| Temperature optima (°C) | 35 | 70 | 60 | 30 | 40 |
| Yield after statistical optimization (U/mL) (fold increase) | 199.90 (1.29) | ND | ND | ND | ND |
| Activating metal ions | Mg2+, Ca2+, Fe2+ and K+ | Mg2+, Ba2+, and Al3+ | Mg2+, Ca2+ | Mn2+, Mg2+, Ca2+ | Ca2+ |
| Inhibitory metal ions | Mn2+, Zn2+ and Hg2+ | ND | ND | ND | Mg, Na+ and K+ |
| Km (mg/mL) | 5.1 | 3.9 | 4.3 | 4.5 | ND |
| Vmax (μM/min/mL) | 116.28 | 96.22 | 98.11 | 103.02 | ND |
| Kcat (per second) | 69.77 | 57.62 | 63.31 | 68.33 | ND |
| Independent Variable | Code | Coded Level | ||||
|---|---|---|---|---|---|---|
| −α | −1 | 0 | +1 | +α | ||
| Soluble starch (%) | E | 0.16 | 0.5 | 1 | 1.5 | 1.84 |
| Media pH | F | 6.32 | 7.00 | 8.00 | 9.00 | 9.68 |
| Incubation period (h) | H | 34.32 | 35.0 | 36 | 37 | 37.68 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bandal, J.N.; Tile, V.A.; Sayyed, R.Z.; Jadhav, H.P.; Azelee, N.I.W.; Danish, S.; Datta, R. Statistical Based Bioprocess Design for Improved Production of Amylase from Halophilic Bacillus sp. H7 Isolated from Marine Water. Molecules 2021, 26, 2833. https://doi.org/10.3390/molecules26102833
Bandal JN, Tile VA, Sayyed RZ, Jadhav HP, Azelee NIW, Danish S, Datta R. Statistical Based Bioprocess Design for Improved Production of Amylase from Halophilic Bacillus sp. H7 Isolated from Marine Water. Molecules. 2021; 26(10):2833. https://doi.org/10.3390/molecules26102833
Chicago/Turabian StyleBandal, J. N., V. A. Tile, R. Z. Sayyed, H. P. Jadhav, N. I. Wan Azelee, Subhan Danish, and Rahul Datta. 2021. "Statistical Based Bioprocess Design for Improved Production of Amylase from Halophilic Bacillus sp. H7 Isolated from Marine Water" Molecules 26, no. 10: 2833. https://doi.org/10.3390/molecules26102833
APA StyleBandal, J. N., Tile, V. A., Sayyed, R. Z., Jadhav, H. P., Azelee, N. I. W., Danish, S., & Datta, R. (2021). Statistical Based Bioprocess Design for Improved Production of Amylase from Halophilic Bacillus sp. H7 Isolated from Marine Water. Molecules, 26(10), 2833. https://doi.org/10.3390/molecules26102833