
Effect of Water and Ethanol Extracts from Hericium erinaceus Solid-State Fermented Wheat Product on the Protection and Repair of Brain Cells in Zebrafish Embryos


Abstract
:1. Introduction
2. Results
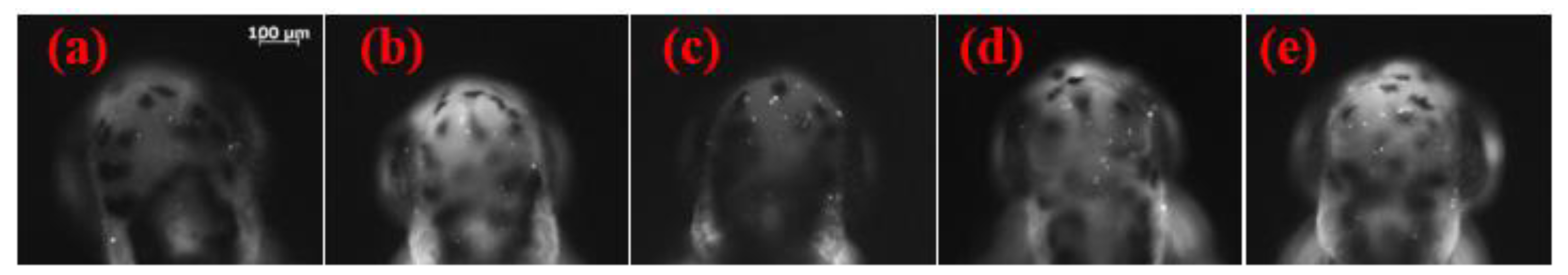
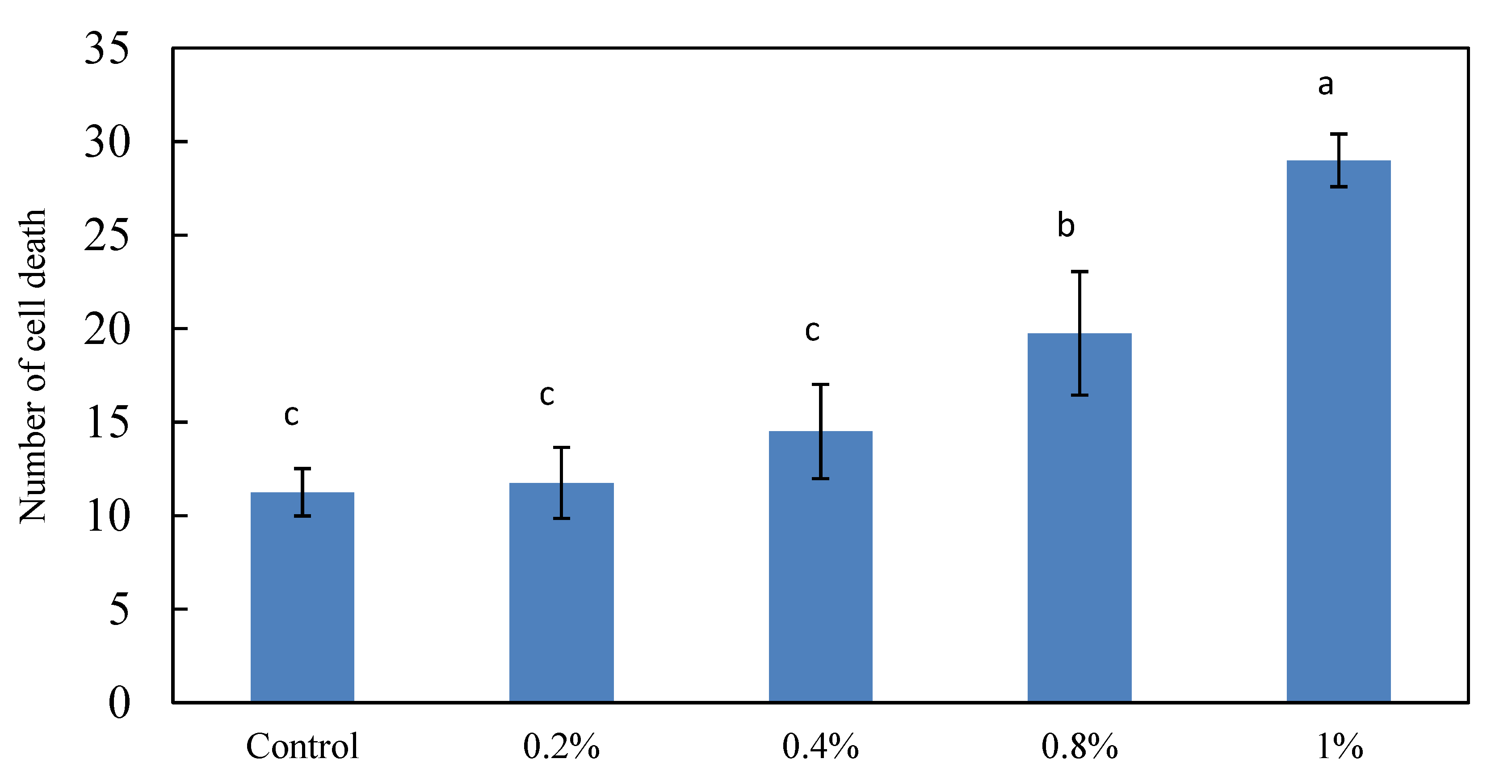
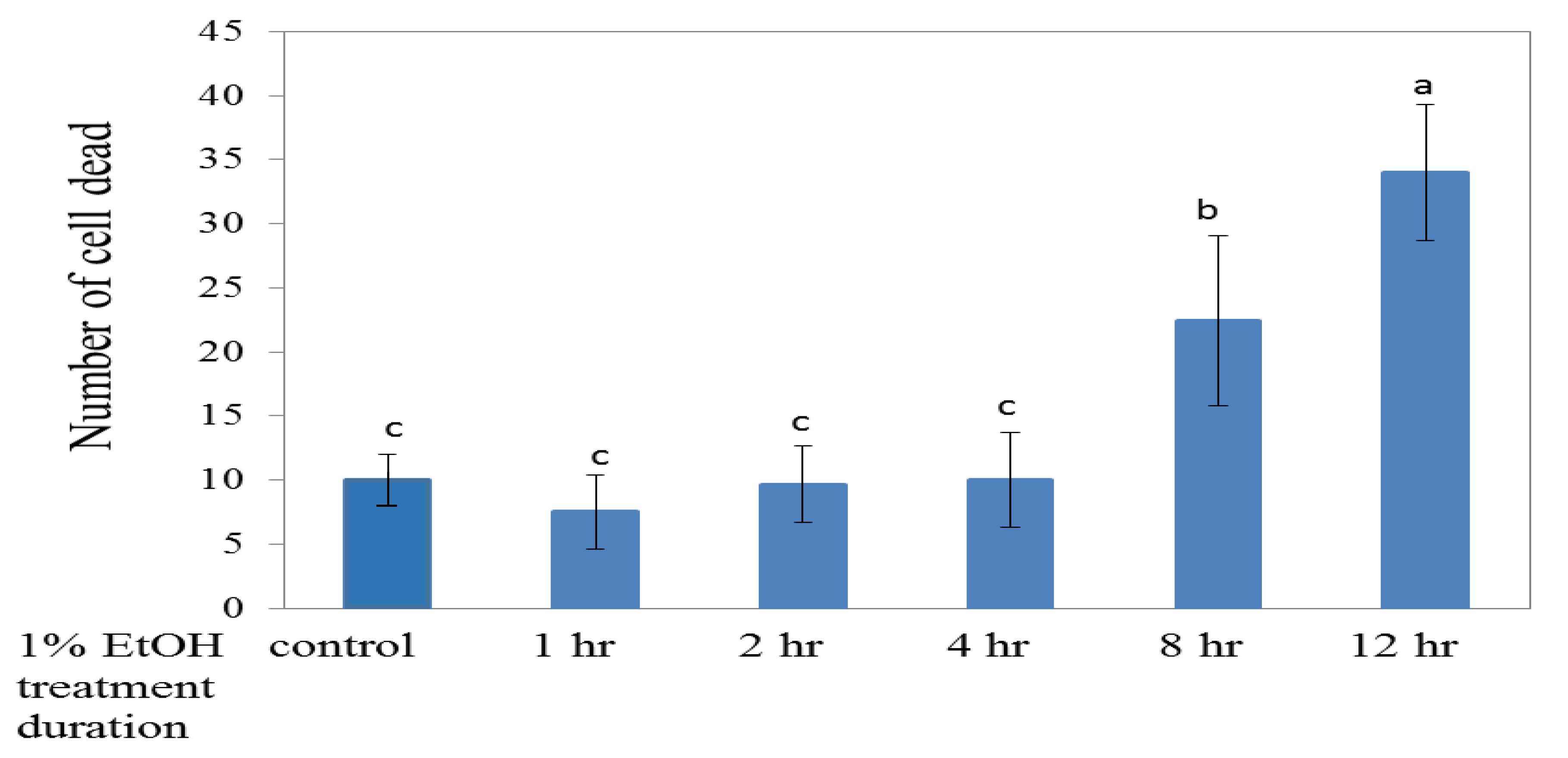
2.1. Effects of the Different Ethanol Concentrations on the Death of Brain Cells in Zebrafish Embryos
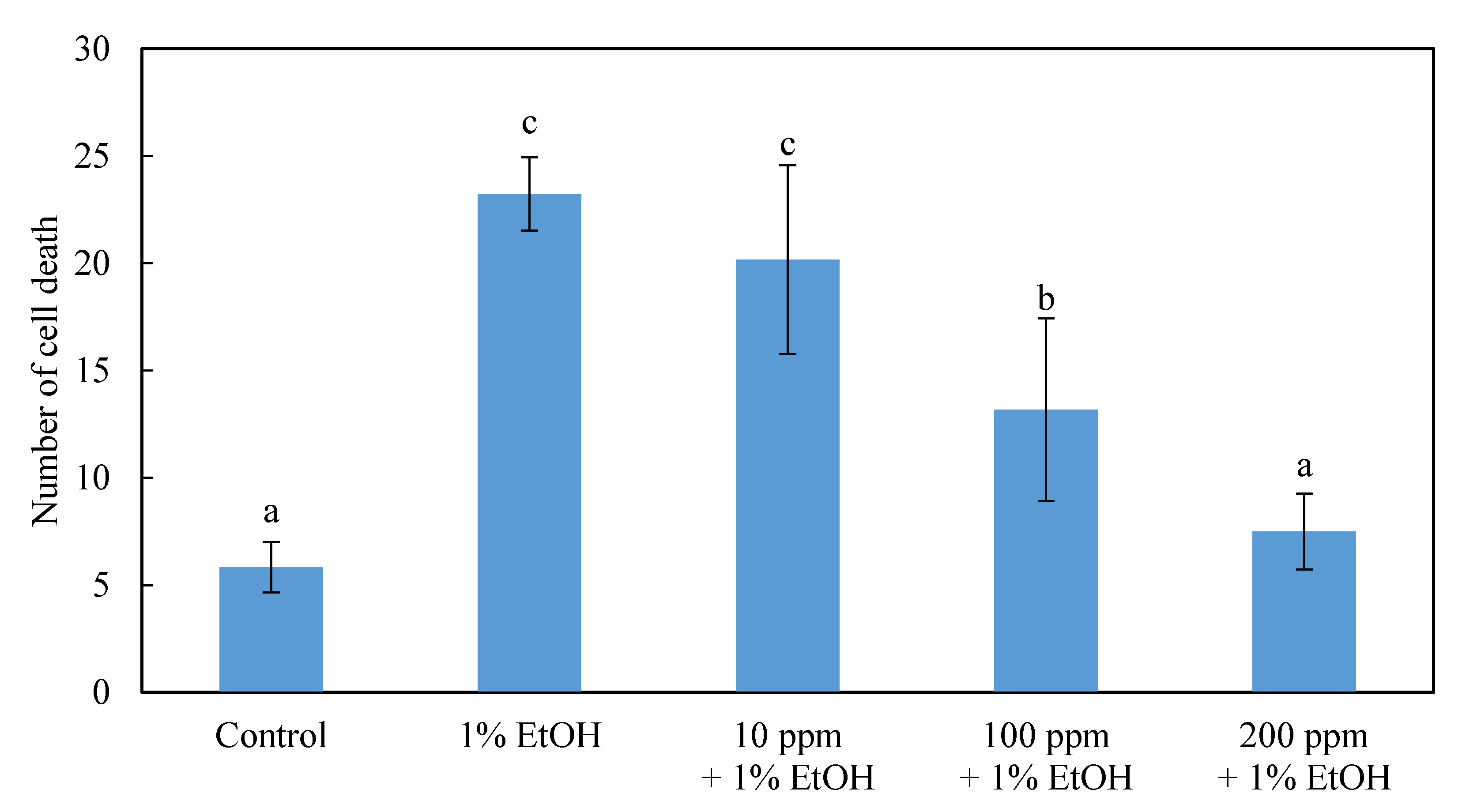
2.2. The Effects of Different Concentrations of HEEE and HEWE Pretreatments on the Neural Cells in the Brain of Zebrafish Embryos
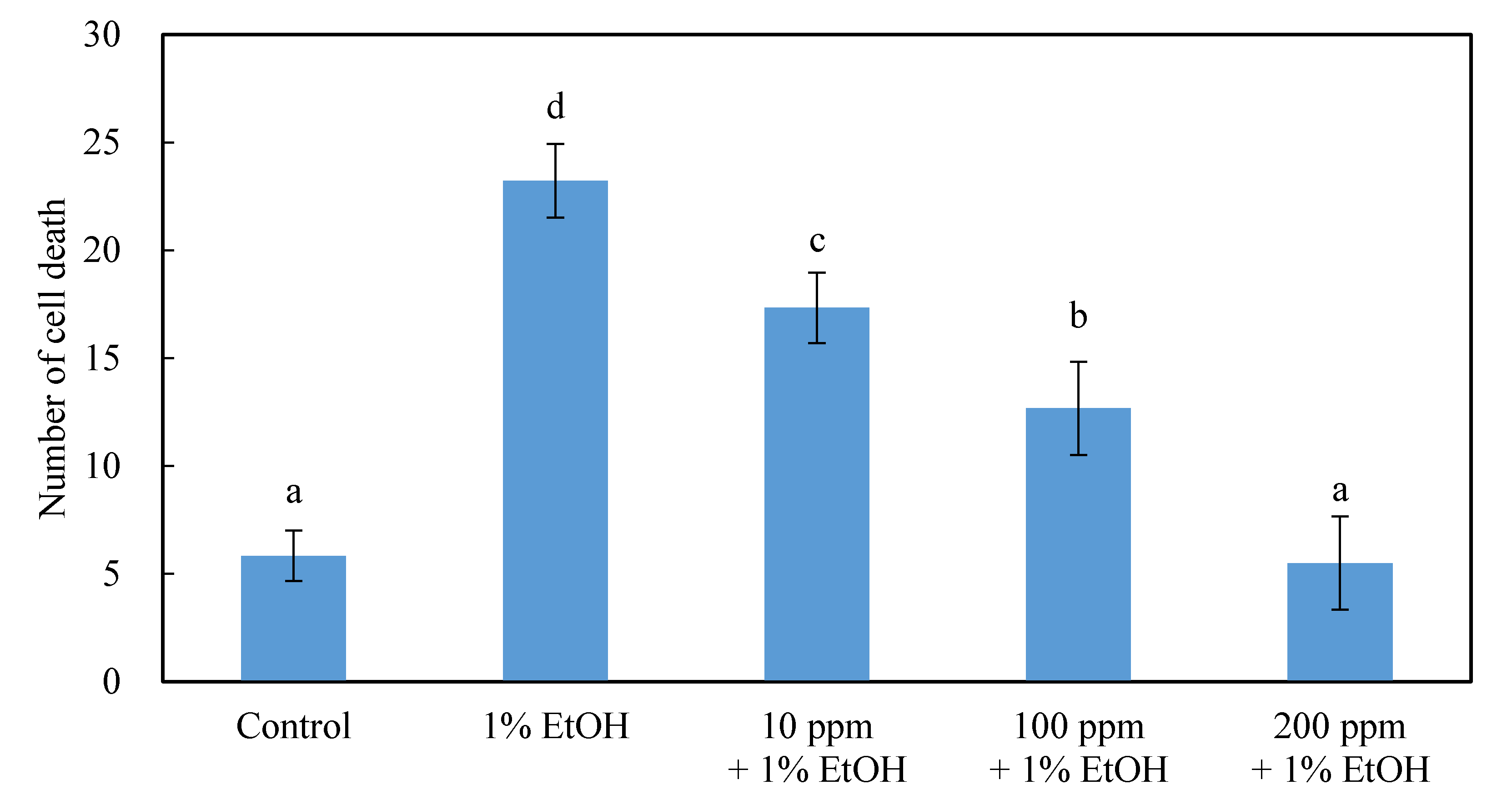
2.3. The Effect of 200 ppm of the HEEE Post-Dosing Treatment in Repairing the Neural Cells in the Brain of Zebrafish Embryos
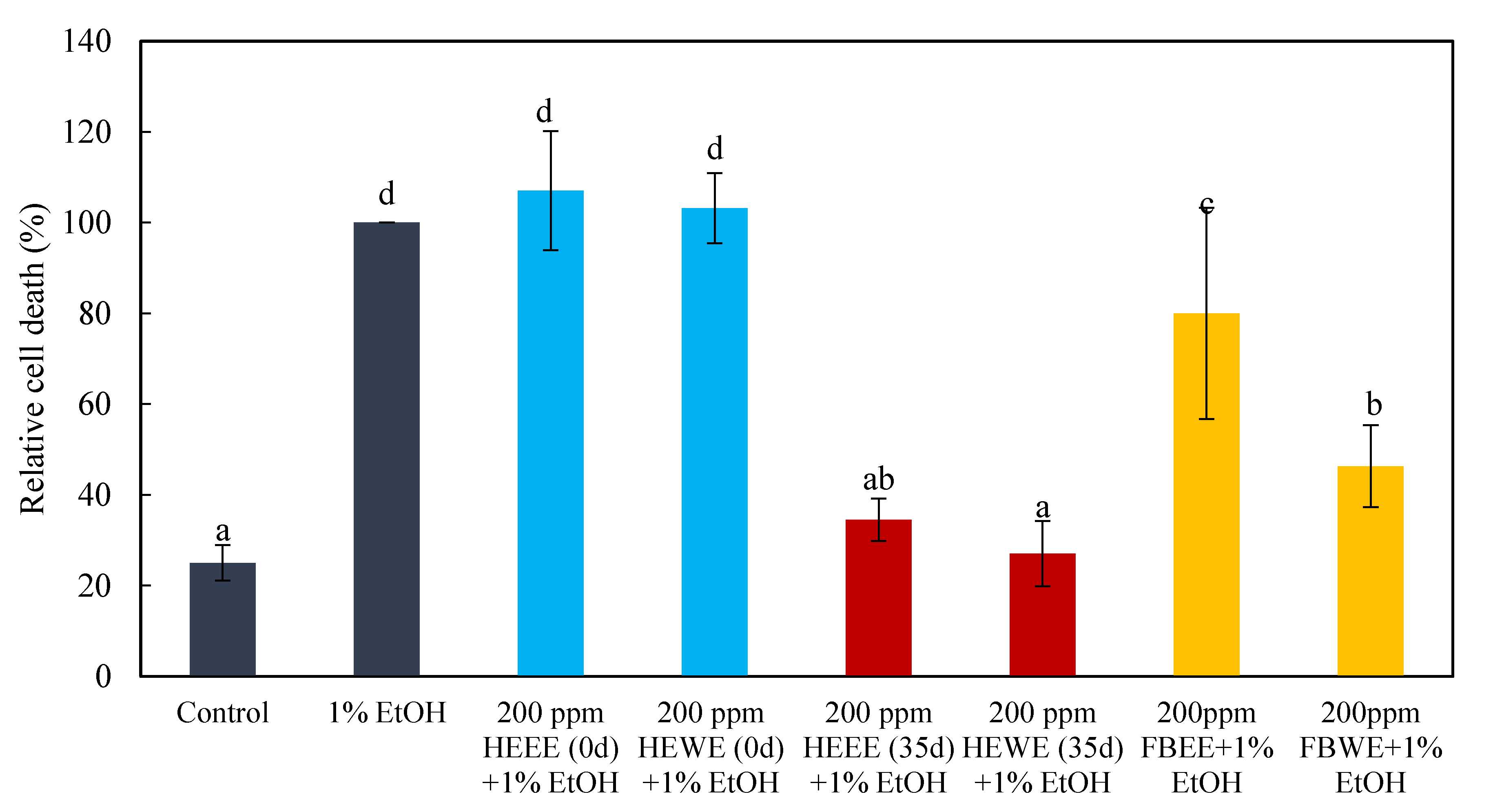
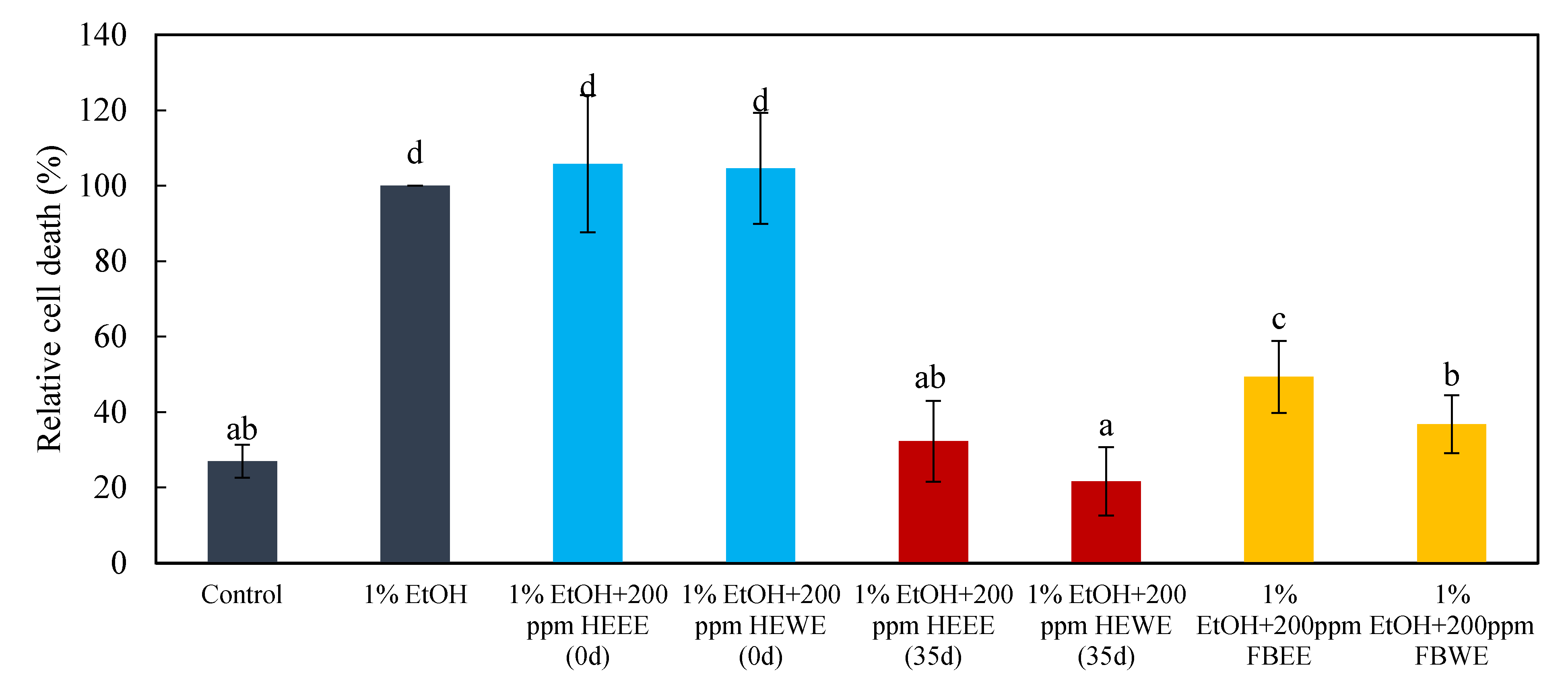
2.4. The Pre-Dosing and Post-Dosing Effects of Treatments with 200 ppm of 35-Day HEEE, 35-Day HWWE, 0-Day HEEE, 0-Day HEWE FBEE, and FBWE on the Neural Cells of Zebrafish Embryos
3. Discussion
3.1. Effects of Different Ethanol Concentrations on Brain Cell Deaths in Zebrafish Embryos
3.2. The Effects of Different Concentrations of HEEE and HEWE Pretreatments on the Neural Cells in the Brain of Zebrafish Embryos
3.3. The Effect of 200 ppm of the HEEE Post-Dosing Treatment in Repairing the Neural Cells in the Brain of Zebrafish Embryos
3.4. The Pre-Dosing and Post-Dosing Effect of Treatments with 200 ppm of 35-Day HEEE, 35-Day HWWE, 0-Day HEEE, 0-Day HEWE FBEE, and FBWE on the Neural cells of Zebrafish Embryos
4. Materials and Methods
4.1. Materials
4.2. Equipment
4.3. Cultivation of Hericium erinaceus
4.3.1. Activation of Hericium erinaceus
4.3.2. Cultivation of Hericium erinaceus Solid-State Fermented Wheat Product
4.4. Extraction of Hericium erinaceus Solid-State Fermented Wheat Product
4.5. The Breeding and Fertilized Zebrafish Egg Collection
4.6. Dosing Culture of Zebrafish Embryos
4.6.1. Pre-dosing Treatment
4.6.2. Post-Dosing Treatment
4.6.3. Fluorescent Staining of Zebrafish Embryos
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
References
- Kawagishi, H.; Ando, M.; Mizuno, T. Hericenone A and B as cytotoxic principles from the mushroom. Tetrahedron Lett. 1990, 31, 373–376. [Google Scholar] [CrossRef]
- Kawagishi, H.; Ando, M.; Sakamoto, H.; Yoshida, S.; Ojima, F.; Ishiguro, Y.; Ukai, N.; Furukawa, S. Hericenones C, D and E, stimulators of neuron growth factor (NGF)–synthesis, from the mushroom Hericium erinaceum. Tetrahedron Lett. 1991, 32, 4561–4564. [Google Scholar] [CrossRef]
- Kawagishi, H.; Ando, M.; Shinba, K.; Sakamoto, H.; Yoshida, S.; Ojima, F.; Ishiguro, Y.; Ukai, N.; Furukawa, S. Chromans, hericenones F, G and H from the mushroom Hericium erinaceum. Phytochemistry 1992, 32, 175–178. [Google Scholar] [CrossRef]
- Ueda, K.; Tsujimori, M.; Kodani, S.; Chiba, A.; Kubo, M.; Masuno, K.; Sekiya, A.; Nagai, K.; Kawagishi, H. An endoplasmic reticulum (ER) stress-suppressive compound and its analogues from the mushroom Hericium erinaceum. Bioorganic Med. Chem. 2008, 16, 9467–9470. [Google Scholar] [CrossRef] [PubMed]
- Kawagishi, H.; Shimada, A.; Shirai, R.; Okamoto, K.; Ojima, F.; Sakamoto, H.; Ishiguro, Y.; Furukawa, S. Erinacines A, B and C, strong stimulators of neuron growth factor (NGF) synthesis, from the mycelia of Hericium erinaceum. Tetrahedron Lett. 1994, 35, 1569–1572. [Google Scholar] [CrossRef]
- Kawagishi, H.; Shimada, A.; Hosokawa, S.; Mori, H.; Sakamoto, H.; Ishiguro, Y.; Sakemi, S.; Bordner, J.; Kojima, N.; Furukawa, S. Erinacines E, F, and G, stimulators of neuron growth factor (NGF) synthesis, from the mycelia of Hericium erinaceum. Tetrahedron Lett. 1996, 37, 7399–7402. [Google Scholar] [CrossRef]
- Kawagishi, H.; Masui, A.; Tokuyama, S.; Nakamura, T. Erinacines J and K from the mycelia of Hericium erinaceum. Tetrahedron 2006, 62, 8463–8466. [Google Scholar] [CrossRef]
- Lee, E.W.; Shizuki, K.; Hosokawa, S.; Suzuki, M.; Suganuma, H.; Inakuma, T.; Li, J.; Ohnishi-Kameyama, M.; Nagata, T.; Furukawa, S.; et al. Two novel diterpenoids, erinacines H and I from the mycelia of Hericium erinaceum. Biosci. Biotech. Biochem. 2000, 64, 2402–2405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenmoku, H.; Sassa, T.; Kato, N. Isolation of erinacine P, a new parental metabolite of cyathane-xylosides, from Hericium erinaceum and its biomimetic conversion into erinacines A and B. Tetrahedron Lett. 2000, 41, 4389–4393. [Google Scholar] [CrossRef]
- Kenmoku, H.; Shimai, T.; Toyomasu, T.; Kato, N.; Sassa, T. Erinacine Q, a New Erinacine from Hericium erinaceum, and its Biosynthetic Route to Erinacine C in the Basidiomycete. Biosci. Biotechnol. Biochem. 2002, 66, 571–575. [Google Scholar] [CrossRef] [Green Version]
- Ma, B.-J.; Zhou, Y.; Li, L.-Z.; Li, H.-M.; Gao, Z.-M.; Ruan, Y. A New Cyathane-xyloside from the Mycelia of Hericium erinaceum. Zeitschrift Naturforschung B 2008, 63, 1241–1242. [Google Scholar] [CrossRef]
- Ma, B.-J.; Shen, J.-W.; Yu, H.-Y.; Ruan, Y.; Wu, T.-T.; Zhao, X. Hericenones and erinacines: Stimulators of nerve growth factor (NGF) biosynthesis in Hericium erinaceus. Mycology 2010, 1, 92–98. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.H.; Naidu, M.; David, R.P.; Abdulla, M.A.; Abdullah, N.; Kuppusamy, U.R.; Sabaratnam, V. Functional recovery enhancement following injury to rodent peroneal nerve by lion’s mane mushroom, Hericium erinaceus (Bull.: Fr.) Pers. (Aphyllophoromycetideae). Int. J. Med. Mushrooms 2009, 11, 225–236. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.-H.; Kanagasabapathy, G.; Bakar, R.; Phan, C.-W.; Sabaratnam, V. Restoration of sensory dysfunction following peripheral nerve injury by the polysaccharide from culinary and medicinal mushroom, Hericium erinaceus (Bull.: Fr.) Pers. through its neuroregenerative action. Food Sci. Technol. 2015, 35, 712–721. [Google Scholar] [CrossRef] [Green Version]
- Yang, P.-P.; Chueh, S.-H.; Shie, H.-L.; Chen, C.-C.; Lee, L.-Y.; Chen, W.-P.; Chen, Y.-W.; Shiu, L.-Y.; Liu, P.-S. Effects of Hericium erinaceus Mycelium Extracts on the Functional Activity of Purinoceptors and Neuropathic Pain in Mice with L5 Spinal Nerve Ligation. Evid. Based Complement. Altern. Med. 2020, 2020, 1–12. [Google Scholar] [CrossRef]
- Hazekawa, M.; Kataoka, A.; Hayakawa, K.; Uchimasu, T.; Furuta, R.; Irie, K.; Akitake, Y.; Yoshida, M.; Fujioka, T.; Egashira, N.; et al. Neuroprotective Effect of Repeated Treatment with Hericium erinaceum in Mice Subjected to Middle Cerebral Artery Occlusion. J. Health Sci. 2010, 56, 296–303. [Google Scholar] [CrossRef] [Green Version]
- Mori, K.; Obara, Y.; Moriya, T.; Inatomi, S.; Nakahata, N. Effects of Hericium erinaceus on amyloid β(25–35) peptide-induced learning and memory deficits in mice. Biomed. Res. 2011, 32, 67–72. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.-H.; Naidu, M.; David, P.; Abdulla, M.A.; Abdullah, N.; Kuppusamy, U.R.; Sabaratnam, V. Corrigendum to “Peripheral Nerve Regeneration Following Crush Injury to Rat Peroneal Nerve by Aqueous Extract of Medicinal Mushroom Hericium erinaceus (Bull.: Fr) Pers. (Aphyllophoromycetideae)”. Evid. Based Complement. Altern. Med. 2018, 2018, 9820769. [Google Scholar] [CrossRef] [Green Version]
- Mori, K.; Inatomi, S.; Ouchi, K.; Azumi, Y.; Tuchida, T. Improving effects of the mushroom Yamabushitake (Hericium erinaceus) on mild cognitive impairment: A double-blind placebo-controlled clinical trial. Phytother. Res. 2009, 23, 367–372. [Google Scholar] [CrossRef]
- Li, I.-C.; Chang, H.-H.; Lin, C.-H.; Chen, W.-P.; Lu, T.-H.; Lee, L.-Y.; Chen, Y.-W.; Chen, Y.-P.; Chen, C.-C.; Lin, D.P.-C. Prevention of Early Alzheimer’s Disease by Erinacine A-Enriched Hericium erinaceus Mycelia Pilot Double-Blind Placebo-Controlled Study. Front. Aging Neurosci. 2020, 12, 155. [Google Scholar] [CrossRef]
- Andreasen, E.A.; Hahn, M.E.; Heideman, W.; Peterson, R.E.; Tanguay, R.L. The zebrafish (Danio rerio) aryl hydrocarbon receptor type 1 (zfAHR1) is a novel vertebrate receptor. Mol. Pharmacol. 2002, 62, 234–249. [Google Scholar] [CrossRef] [Green Version]
- Gerhard, G.S.; Kauffman, E.J.; Wang, X.; Stewart, R.; Moore, J.L.; Kasales, C.J.; Demidenko, E.; Cheng, K.C. Life spans and senescent phenotypes in two strains of Zebrafish (Danio rerio). Exp. Gerontol. 2002, 37, 1055–1068. [Google Scholar] [CrossRef]
- Tanguay, R.L.; Abnet, C.C.; Heideman, W.; Peterson, R.E. Cloning and characterization of the zebrafish (Danio rerio) aryl hydrocarbon receptor. Biochim. Biophys. Acta Gene Struct. Expr. 1999, 1444, 35–48. [Google Scholar] [CrossRef]
- Zodrow, J.M.; Stegeman, J.J.; Tanguay, R.L. Histological analysis of acute toxicity of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) in zebrafish. Aquat. Toxicol. 2004, 66, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Reimers, M.J.; La Du, J.K.; Periera, C.B.; Giovanini, J.; Tanguay, R.L. Ethanol-dependent toxicity in zebrafish is partially attenuated by antioxidants. Neurotoxicol. Teratol. 2006, 28, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Arendt, T.; Allen, Y.; Sinden, J.; Schugens, M.M.; Marchbanks, R.M.; Lantos, P.L.; Gray, J.A. Cholinergic-rich brain transplants reverse alcohol-induced memory deficits. Nat. Cell Biol. 1988, 332, 448–450. [Google Scholar] [CrossRef]
- Charness, M.E. Brain Lesions in Alcoholics. Alcohol. Clin. Exp. Res. 1993, 17, 2–11. [Google Scholar] [CrossRef]
- Santín, L.; Rubio, S.; Begega, A.; Arias, J. Effects of chronic alcohol consumption on spatial reference and working memory tasks. Alcohol 2000, 20, 149–159. [Google Scholar] [CrossRef]
- Walker, D.W.; Freund, G. Impairment of shuttle box avoidance learning following prolonged alcohol consumption in rats. Physiol. Behav. 1971, 7, 773–778. [Google Scholar] [CrossRef]
- Neuhauss, S.C.; Solnica-Krezel, L.; Schier, A.F.; Zwartkruis, F.; Stemple, D.L.; Malicki, J.; Abdelilah, S.; Stainier, D.; Driever, W. Mutations affecting craniofacial development in zebrafish. Development 1996, 123, 357–367. [Google Scholar] [CrossRef]
- Piotrowski, T.; Schilling, T.F.; Brand, M.; Jiang, Y.J.; Heisenberg, C.P.; Beuchle, D.; Grandel, H.; van Eeden, F.J.; Furutani-Seiki, M.; Granato, M.; et al. Jaw and branchial arch mutants in zebrafish: II. Anterior arches and cartilage differentiation. Development 1996, 123, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Reimers, M.J.; Flockton, A.R.; Tanguay, R.L. Ethanol- and acetaldehyde-mediated developmental toxicity in zebrafish. Neurotoxicol. Teratol. 2004, 26, 769–781. [Google Scholar] [CrossRef]
- Schilling, T.F.; Piotrowski, T.; Grandel, H.; Brand, M.; Heisenberg, C.P.; Jiang, Y.J.; Beuchle, D.; Hammerschmidt, M.; Kane, D.A.; Mullins, M.C.; et al. Jaw and branchial arch mutants in zebrafish: I. Branchial arches. Development 1996, 123, 329–344. [Google Scholar] [CrossRef]
- Carvan, M.J.; Loucks, E.; Weber, D.N.; Williams, F.E. Ethanol effects on the developing zebrafish: Neurobehavior and skeletal morphogenesis. Neurotoxicol. Teratol. 2004, 26, 757–768. [Google Scholar] [CrossRef] [PubMed]
- Parng, C.; Roy, N.M.; Ton, C.; Lin, Y.; McGrath, P. Neurotoxicity assessment using zebrafish. J. Pharmacol. Toxicol. Methods 2007, 55, 103–112. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Yen, W.-Y.; Sun, S.-K.; Chen, S.-D. Effects of extracts from Hericium erinaceus solid-state fermented products on protection and repair of brain cells in zebrafish embryos. Taiwan. J. Agric. Chem. Food Sci. 2016, 54, 98–106. [Google Scholar]
- Xia, G.; Han, X.; Qi, J.; Liu, W.; Song, J.; Qin, J.; Liu, L. The effects of astragalus polysaccharide on zebrafish cell apoptosis and senescence. Am. J. Mol. Biol. 2012, 2, 103–109. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zhang, Z.; Cheang, L.C.; Lin, Z.; Lee, S.M. Eriocaulon buergerianum extract protects PC12 cells and neurons in zebrafish against 6-hydroxydopamine-induced damage. Chin. Med. 2011, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.-J.; Cheang, L.C.V.; Wang, M.-W.; Li, G.-H.; Chu, I.K.; Lin, Z.-X.; Lee, S.M.Y. Ethanolic Extract of Fructus Alpinia oxyphylla Protects Against 6-Hydroxydopamine-Induced Damage of PC12 Cells In Vitro and Dopaminergic Neurons in Zebrafish. Cell. Mol. Neurobiol. 2011, 32, 27–40. [Google Scholar] [CrossRef]
- Rosemberg, D.B.; da-Rocha, R.F.; Rico, E.P.; Zanotto–Filho, A.; Dias, R.D.; Bogo, M.R.; Bonan, C.D.; Moreira, J.C.; Klamt, F.; Souza, D.O. Taurine prevents enhancement of acetylcholinesterase activity induced by acute ethanol exposure and decreases the level of markers of oxidative stress in zebrafish brain. Neuroscience 2010, 171, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, V.; Kuhad, A.; Chopra, K. Suppression of neuro-inflammatory signaling cascade by tocotrienol can prevent chronic alcohol-induced cognitive dysfunction in rats. Behav. Brain Res. 2009, 203, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.-C.; Kim, K.-N.; Wijesinghe, W.; Yang, X.; Ahn, G.; Jeon, Y.-J. Protective effect of polyphenol extracted from Ecklonia cava against ethanol induced oxidative damage in vitro and in zebrafish model. J. Funct. Foods 2014, 6, 339–347. [Google Scholar] [CrossRef]
- Tiwari, V.; Chopra, K. Resveratrol abrogates alcohol-induced cognitive deficits by attenuating oxidative–nitrosative stress and inflammatory cascade in the adult rat brain. Neurochem. Int. 2013, 62, 861–869. [Google Scholar] [CrossRef]
- Lin, Y.T.; Lin, S.L.; Chen, S.D.; Cheng, Y.H. Studies on the Hericium erinaceus liquid fermentation on conditions with different grains for their antioxidant activities. Ilan Univ. J. Bioresour. 2015, 11, 37–53. [Google Scholar]
- Mitchell, J.; Paiva, M.; Moore, D.B.; Walker, D.W.; Heaton, M.B. A comparative study of ethanol, hypoglycemia, hypoxia and neurotrophic factor interactions with fetal rat hippocampal neurons: A multi-factor in vitro model for developmental ethanol effects. Dev. Brain Res. 1998, 105, 241–250. [Google Scholar] [CrossRef]
- Shimbo, M.; Kawagishi, H.; Yokogoshi, H. Erinacine A increases catecholamine and neuron growth factor content in the central nervous system of rats. Nutr. Res. 2005, 25, 617–623. [Google Scholar] [CrossRef]
- He, C.Y.; Chen, S.D.; Lu, W.J.; Wang, H.C.; Chang, Y.C.; Cheng, Y.H.; Lai, Y.S. Effect of ethanol extracts from Hericium erinaceum solid-state fermented wheat on nerve cells growth. Hans J. Food Nutr. Sci. 2015, 4, 1–10. [Google Scholar] [CrossRef]
- Tapia-Arancibia, L.; Rage, F.; Givalois, L.; Dingeon, P.; Arancibia, S. Effects of alcohol on brain-derived neurotrophic factor mRNA expression in discrete regions of the rat hippocampus and hypothalamus. J. Neurosci. Res. 2001, 63, 200–208. [Google Scholar] [CrossRef]
- Bhave, S.V.; Ghoda, L.; Hoffman, P.L. Brain-derived neurotrophic factor mediates the anti-apoptotic effect of NMDA in cerebellar granule neuron: Signal transduction cascades and site of alcohol action. J. Neurosci. 1999, 19, 3277–3286. [Google Scholar] [CrossRef]
- Alderson, R.F.; Alterman, A.L.; Barde, Y.-A.; Lindsay, R.M. Brain-derived neurotrophic factor increases survival and differentiated functions of rat septal cholinergic neurons in culture. Neuron 1990, 5, 297–306. [Google Scholar] [CrossRef]
- Cohen-Cory, S.; Kidane, A.H.; Shirkey, N.J.; Marshak, S. Brain-derived neurotrophic factor and the development of structural neuronal connectivity. Dev. Neurobiol. 2010, 70, 271–288. [Google Scholar] [CrossRef] [Green Version]
- Madeira, M.D.; Sousa, N.; Lieberman, A.; Paula-Barbosa, M. Effects of chronic alcohol consumption and of dehydration on the supraoptic nucleus of adult male and female rats. Neuroscience 1993, 56, 657–672. [Google Scholar] [CrossRef]
- Lindvall, O.; Kokaia, Z.; Bengzon, J.; Elme, E.; Kokaia, M. Neurotrophins and brain insults. Trends Neurosci. 1994, 17, 490–496. [Google Scholar] [CrossRef]
- Wilson, S.; Ross, L.; Parrett, T.; Easter, S. The development of a simple scaffold of axon tracts in the brain of the embryonic zebrafish, Brachydanio rerio. Development 1990, 108, 121–145. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Heinrich, G. Brain-derived neurotrophic factor gene expression in the developing zebrafish. Int. J. Dev. Neurosci. 1997, 15, 983–997. [Google Scholar] [CrossRef]
- Weis, J.S. Analysis of the development of the nervous system of the zebrafish, Brachydanio rerio. II. The effect of nerve growth factor and its antiserum on the nervous system of the zebrafish. J. Embryol. Exp. Morphol. 1968, 19, 121–135. [Google Scholar]
- Mori, K.; Obara, Y.; Hirota, M.; Azumi, Y.; Kinugasa, S. Neuron growth factor-inducing activity of Hericium erinaceus in 1321N1 human astrocytoma cells. Biol. Pharm. Bull. 2008, 31, 1727–1732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westerfield, M. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish (Danio rerio), 4th ed.; University of Oregon Press: Eugene, OR, USA, 2000. [Google Scholar]
- Parng, C.; Ton, C.; Lin, Y.-X.; Roy, N.M.; McGrath, P. A zebrafish assay for identifying neuroprotectants in vivo. Neurotoxicol. Teratol. 2006, 28, 509–516. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (day) | Mycelium (%) | Crude Polysaccharide (%) | Crude Triterpenoid (%) | Erinacine A (%) |
|---|---|---|---|---|
| 0 | - | 10.402 ± 0.392 d | 0.054 ± 0.025 d | 0.003 ± 0.002 c |
| 21 | 2.118 ± 0.015 d | 11.749 ± 0.321 d | 0.071 ± 0.027 c | 0.029 ± 0.002 b |
| 28 | 2.978 ± 0.065 d | 13.412 ± 0.691 b | 0.108 ± 0.027 b | 0.031 ± 0.002 b |
| 35 | 3.500 ± 0.044 c | 15.649 ± 0.931 a | 0.153 ± 0.029 a | 0.042 ± 0.002 b |
| 42 | 5.846 ± 0.200 b | 10.764 ± 0.315 d | 0.077 ± 0.029 c | 0.051 ± 0.002 ab |
| 56 | 7.415 ± 0.251 a | 10.323 ± 0.464 d | 0.071 ± 0.029 c | 0.062 ± 0.004 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, S.-K.; Ho, C.-Y.; Yen, W.-Y.; Chen, S.-D. Effect of Water and Ethanol Extracts from Hericium erinaceus Solid-State Fermented Wheat Product on the Protection and Repair of Brain Cells in Zebrafish Embryos. Molecules 2021, 26, 3297. https://doi.org/10.3390/molecules26113297
Sun S-K, Ho C-Y, Yen W-Y, Chen S-D. Effect of Water and Ethanol Extracts from Hericium erinaceus Solid-State Fermented Wheat Product on the Protection and Repair of Brain Cells in Zebrafish Embryos. Molecules. 2021; 26(11):3297. https://doi.org/10.3390/molecules26113297
Chicago/Turabian StyleSun, Shun-Kuo, Chun-Yi Ho, Wei-Yang Yen, and Su-Der Chen. 2021. "Effect of Water and Ethanol Extracts from Hericium erinaceus Solid-State Fermented Wheat Product on the Protection and Repair of Brain Cells in Zebrafish Embryos" Molecules 26, no. 11: 3297. https://doi.org/10.3390/molecules26113297
APA StyleSun, S. -K., Ho, C. -Y., Yen, W. -Y., & Chen, S. -D. (2021). Effect of Water and Ethanol Extracts from Hericium erinaceus Solid-State Fermented Wheat Product on the Protection and Repair of Brain Cells in Zebrafish Embryos. Molecules, 26(11), 3297. https://doi.org/10.3390/molecules26113297