
The Protection of Lactic Acid Bacteria Fermented-Mango Peel against Neuronal Damage Induced by Amyloid-Beta





Abstract
:1. Introduction
2. Results and Discussion
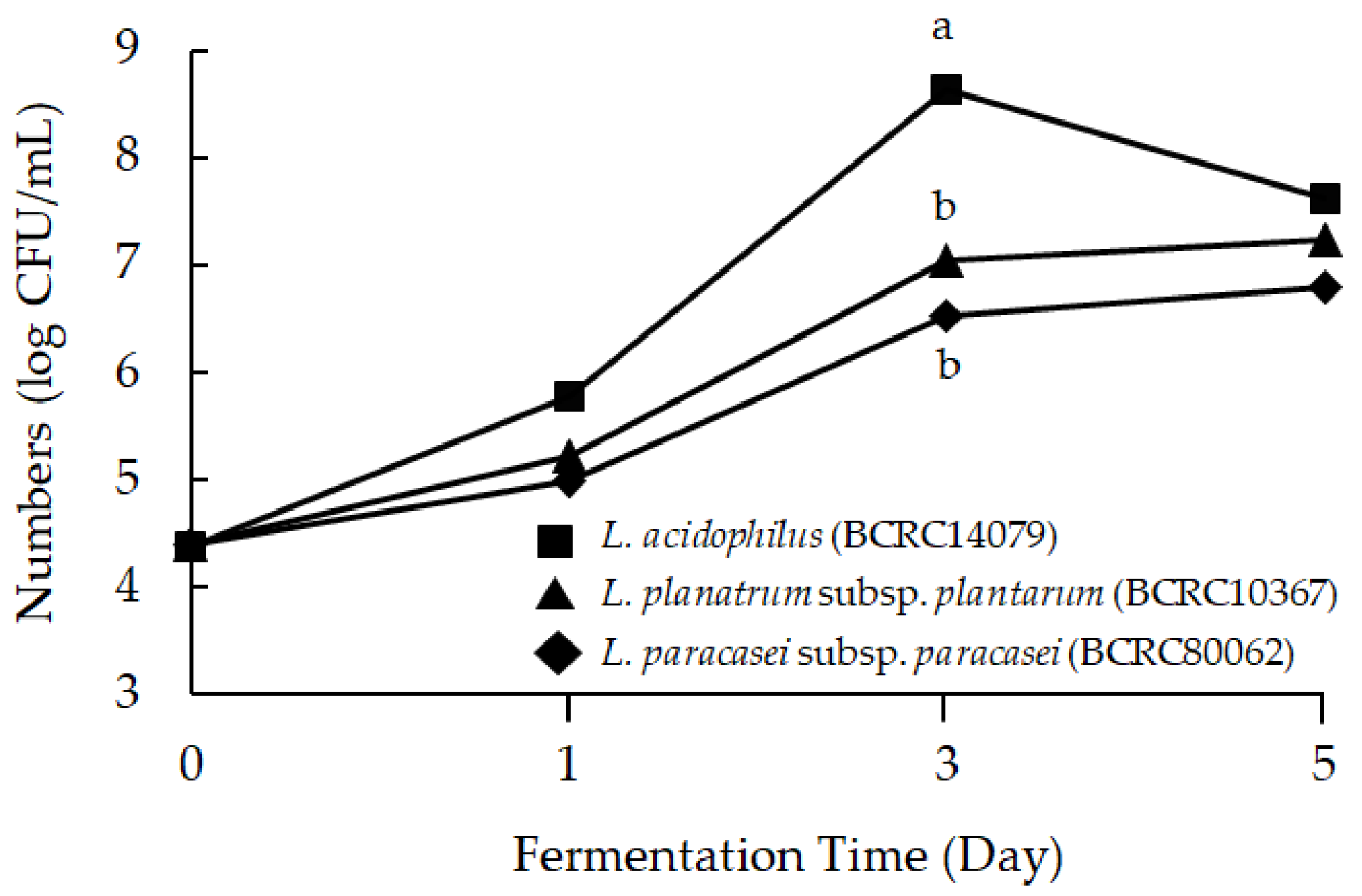
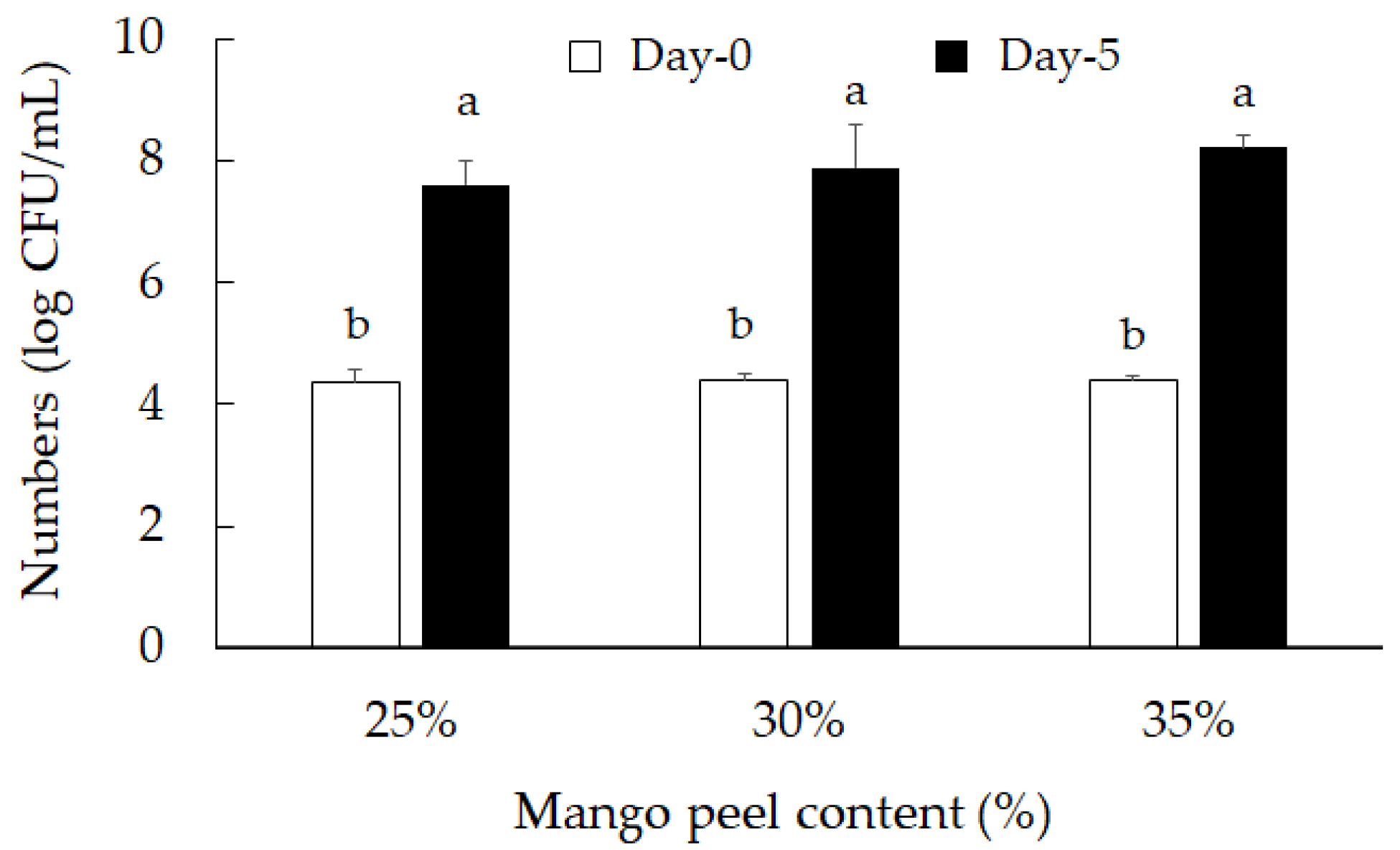
2.1. Mango Peel Fermentation by Lactic Acid Bacteria
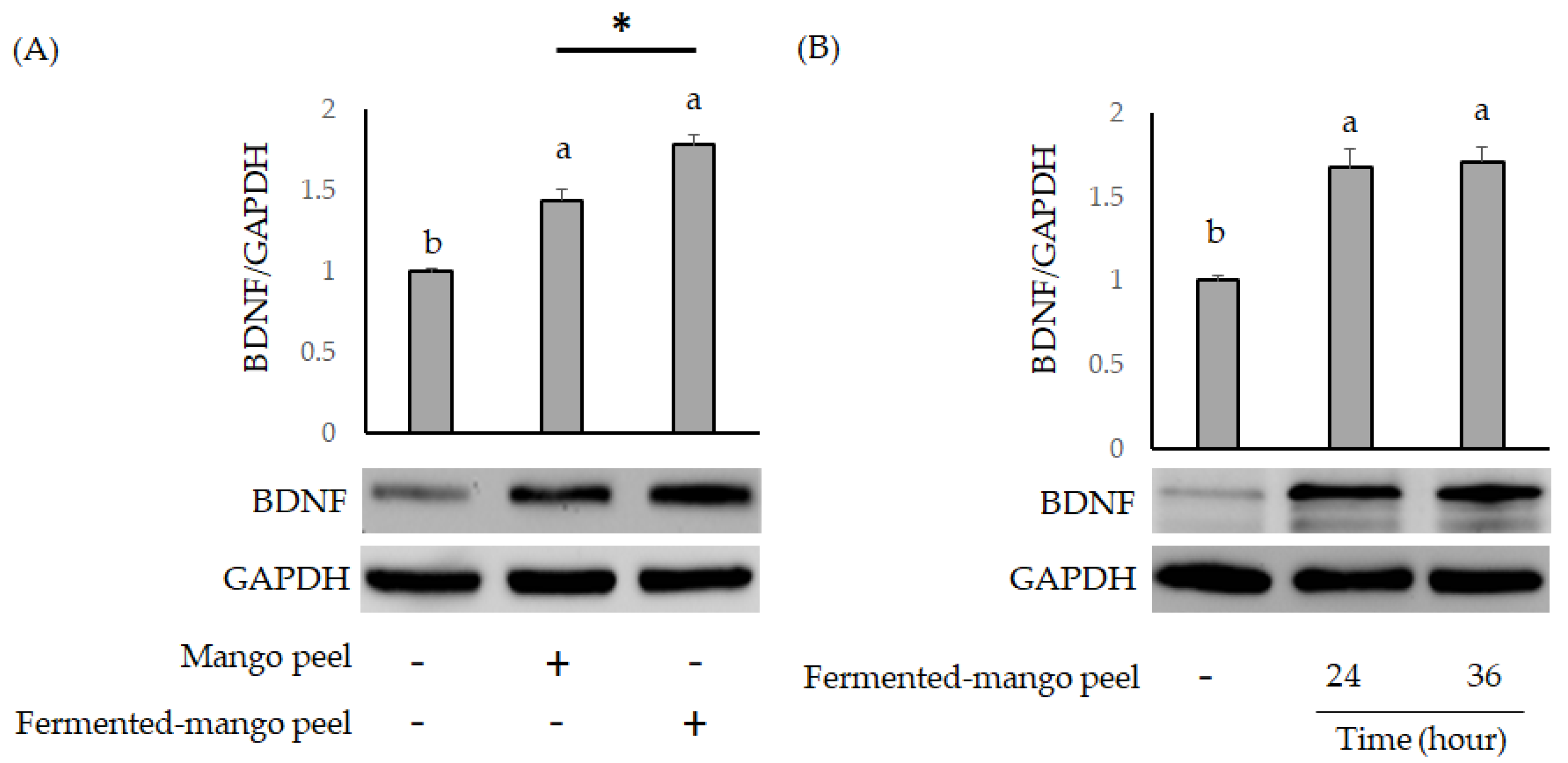
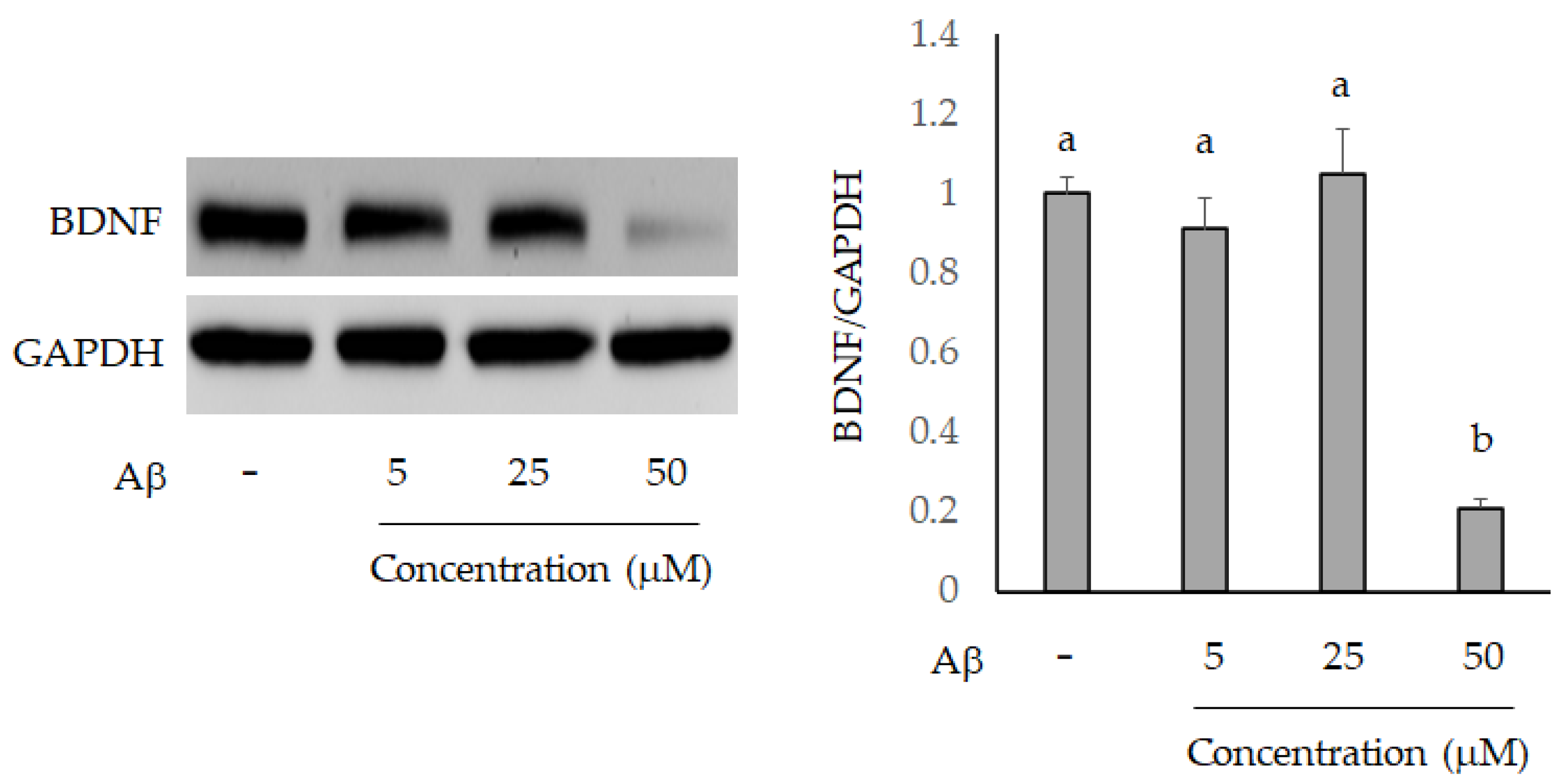
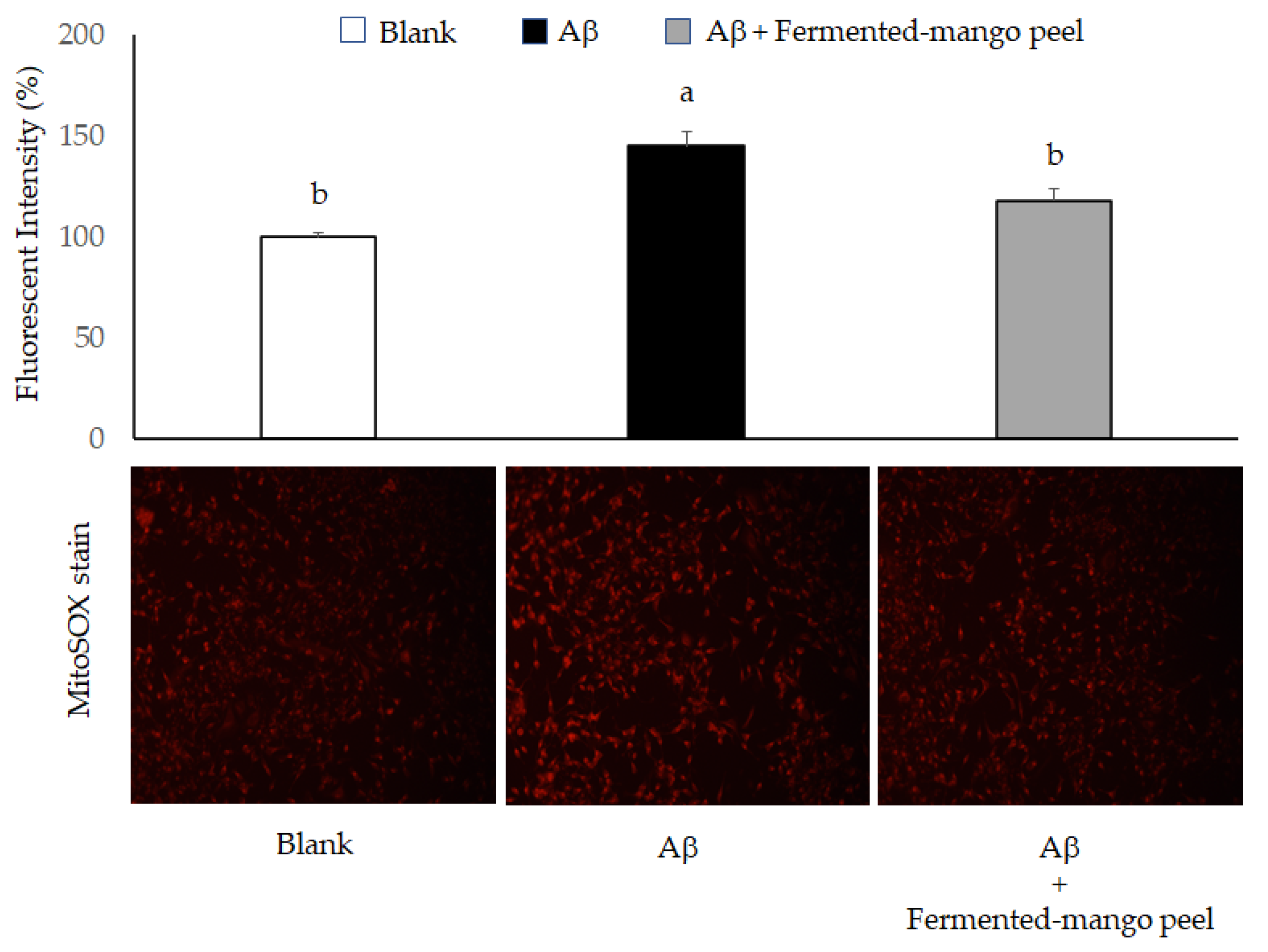
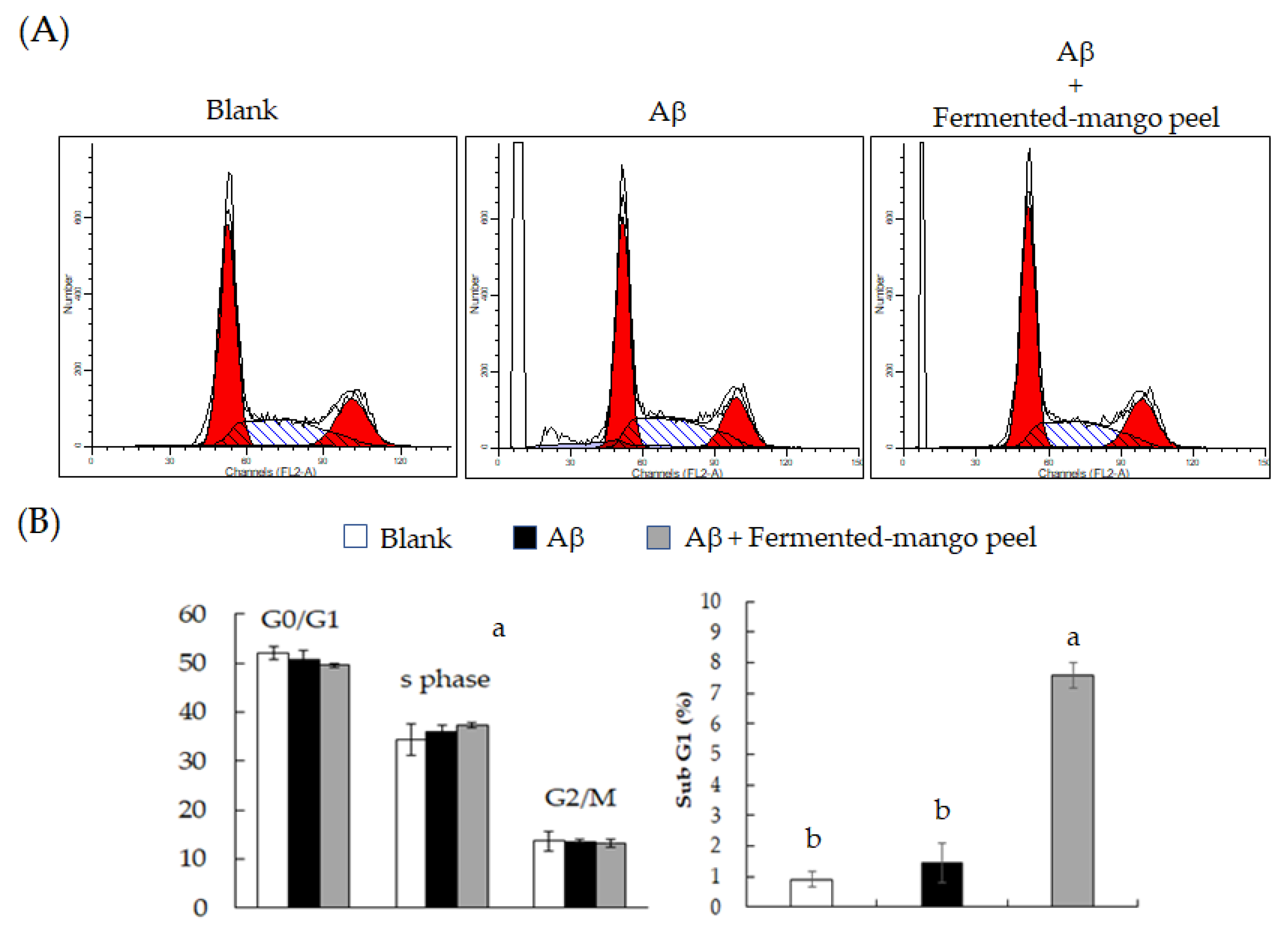
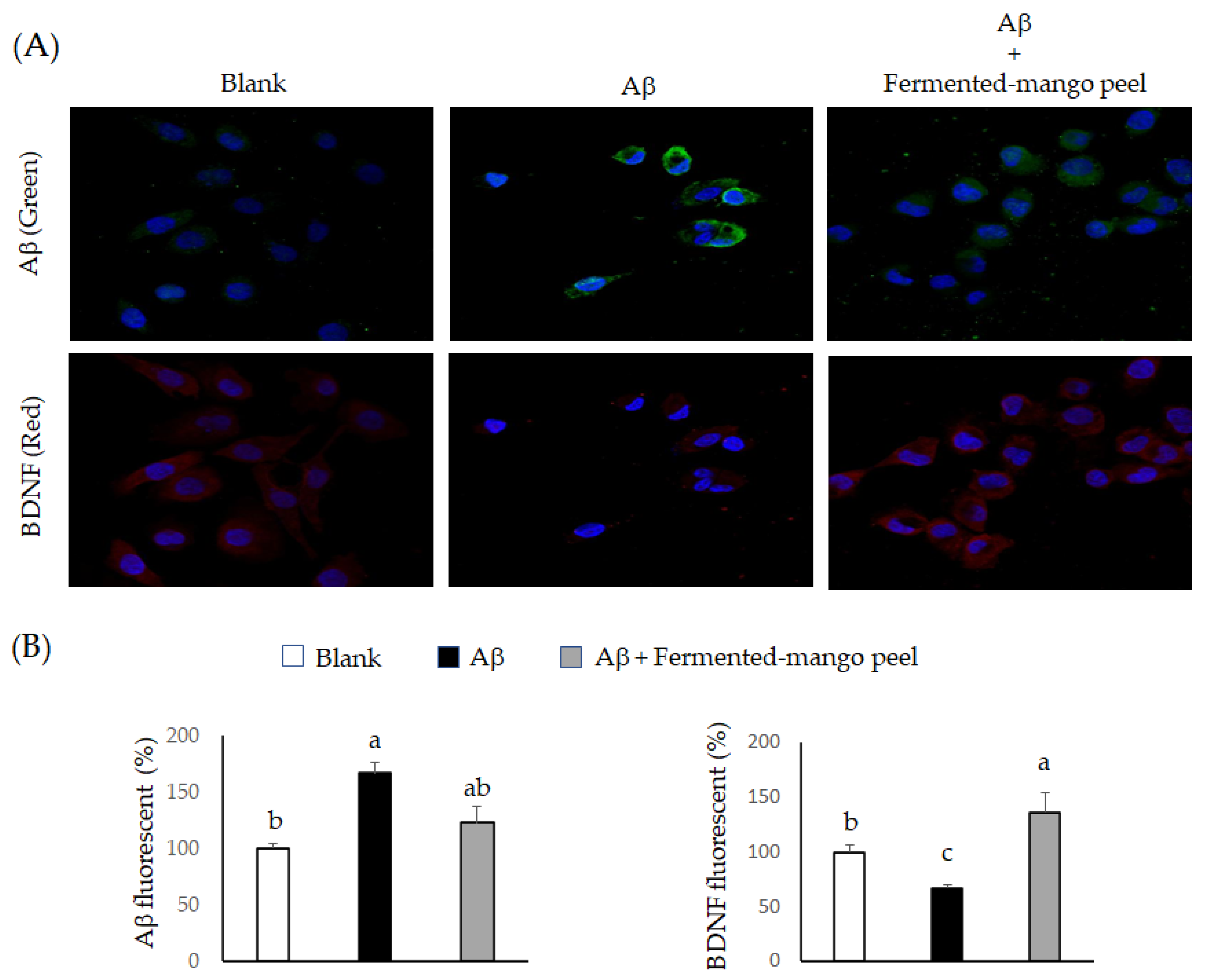
2.2. The Neuronal Protection of L. acidophilus (BCRC14079)-Fermented Mango Peel against Neuron-2A Cells Dysfunction Caused by Aβ Induction
3. Materials and Methods
3.1. Chemicals
3.2. Treatment and Fermentation of Mango Peel
3.3. Cell Culture
3.4. Western Blot
3.5. MitoSOX-Red Stain
3.6. Cell Cycle
3.7. Immunocytochemistry Stain
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
| Aβ | amyloid beta |
| APP | amyloid precursor protein |
| BDNF | brain-derived neurotrophic factor |
| CNS | central nervous system |
| DMEM | Dulbecco’s modified Eagle’s medium |
| DMSO | dimethyl sulfoxide |
| FBS | fetal bovine serum |
| ROS | reactive oxygen species |
| SDS | sodium dodecyl sulfate |
References
- Feng, S.T.; Wang, Z.Z.; Yuan, Y.H.; Sun, H.M.; Chen, N.H.; Zhang, Y. Mangiferin: A multipotent natural product preventing neurodegeneration in Alzheimer’s and Parkinson’s disease models. Pharmacol. Res. 2019, 146, 104336. [Google Scholar] [CrossRef]
- Kasbe, P.; Jangra, A.; Lahkar, M. Mangiferin ameliorates aluminium chloride-induced cognitive dysfunction via alleviation of hippocampal oxido-nitrosative stress, proinflammatory cytokines and acetylcholinesterase level. J. Trace Elem. Med. Biol. 2015, 31, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.Q.; Liu, L.; Gao, Q.W.; Wu, X.N.; Xiang, W.; Deng, W.T. Mangiferin prevents corticosterone-induced behavioural deficits via alleviation of oxido-nitrosative stress and down-regulation of indoleamine 2,3-dioxygenase (IDO) activity. Neurol. Res. 2017, 39, 709–718. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhao, L.; He, T.; Ou, Z.; Hu, Z.; Wang, K. Effects of mango peel powder on starch digestion and quality characteristics of bread. Int. J. Biol. Macromol. 2019, 140, 647–652. [Google Scholar] [CrossRef]
- Reddy, L.; Min, J.H.; Wee, Y.J. Production of probiotic mango juice by fermentation of lactic acid bacteria. Korean J. Microbiol. Biotechnol. 2015, 43, 120–125. [Google Scholar] [CrossRef] [Green Version]
- Liao, X.Y.; Guo, L.Q.; Ye, Z.W.; Qiu, L.Y.; Gu, F.W.; Lin, J.F. Use of autochthonous lactic acid bacteria starters to ferment mango juice for promoting its probiotic roles. Prep. Biochem. Biotechnol. 2016, 46, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Kamassah, A.K.Q.; Saalia, F.K.; Fosu, P.O.; Mensah-Brown, H.; Sinayobye, E.; Tano-Debrah, K. Fermentation capacity of yeasts using mango (Mangiferia indica Linn.) as substrate. Food Sci Qual. Manag. 2013, 22, 69–78. [Google Scholar]
- Reddy, L.V.; Reddy, O.V.S. Production of ethanol from mango (Mangifera indica L.) fruit juice fermentation. Res. J. Microbiol. 2007, 2, 763–769. [Google Scholar]
- Li, X.; Chan, L.J.; Yu, B.; Curran, P.; Liu, S.Q. Fermentation of three varieties of mango juices with a mixture of Saccharomyces cerevisiae and Williopsis saturnus var. mrakii. Int. J. Food Microbiol. 2012, 158, 28–35. [Google Scholar] [CrossRef]
- Ordonez-Diaz, J.L.; Moreno-Ortega, A.; Roldan-Guerra, F.J.; Ortiz-Somovilla, V.; Moreno-Rojas, J.M.; Pereira-Caro, G. In vitro gastrointestinal digestion and colonic catabolism of mango (Mangifera indica L.) pulp polyphenols. Foods 2020, 9, 1836. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Maldonado, L.M.; Blancas-Benitez, F.J.; Zamora-Gasga, V.M.; Cardenas-Castro, A.P.; Tovar, J.; Sayago-Ayerdi, S.G. In vitro gastrointestinal digestion and colonic fermentation of high dietary fiber and antioxidant-rich mango (Mangifera indica L.) “Ataulfo”-based fruit bars. Nutrients 2019, 11, 1564. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez-Sarmiento, W.; Sayago-Ayerdi, S.G.; Goni, I.; Gutierrez-Miceli, F.A.; Abud-Archila, M.; Rejon-Orantes, J.C.; Rincon-Rosales, R.; Pena-Ocana, B.A.; Ruiz-Valdiviezo, V.M. Changes in intestinal microbiota and predicted metabolic pathways during colonic fermentation of mango (Mangifera indica L.)-based bar indigestible fraction. Nutrients 2020, 12, 683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bostanciklioglu, M. The role of gut microbiota in pathogenesis of Alzheimer’s disease. J. Appl. Microbiol. 2019, 127, 954–967. [Google Scholar] [CrossRef] [PubMed]
- Tanida, M.; Yamano, T.; Maeda, K.; Okumura, N.; Fukushima, Y.; Nagai, K. Effects of intraduodenal injection of Lactobacillus johnsonii La1 on renal sympathetic nerve activity and blood pressure in urethane-anesthetized rats. Neurosci. Lett. 2005, 389, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Bravo, J.A.; Forsythe, P.; Chew, M.V.; Escaravage, E.; Savignac, H.M.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc. Natl. Acad. Sci. USA 2011, 108, 16050–16055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.C.; Su, Y.S.; Cheng, H.Y. Antioxidant properties of Lactobacillus-fermented and non-fermented Graptopetalum paraguayense E. Walther at different stages of maturity. Food Chem. 2011, 129, 804–809. [Google Scholar] [CrossRef]
- Liu, C.F.; Tseng, K.C.; Chiang, S.S.; Lee, B.H.; Hsu, W.H.; Pan, T.M. Immunomodulatory and antioxidant potential of Lactobacillus exopolysaccharides. J. Sci. Food Agric. 2011, 91, 2284–2291. [Google Scholar] [CrossRef]
- Liu, C.F.; Hu, C.L.; Chiang, S.S.; Tseng, K.C.; Yu, R.C.; Pan, T.M. Beneficial preventive effects of gastric mucosal lesion for soy-skim milk fermented by lactic acid bacteria. J. Agric. Food Chem. 2009, 57, 4433–4438. [Google Scholar] [CrossRef]
- Chiu, C.H.; Lu, T.Y.; Tseng, Y.Y.; Pan, T.M. The effects of Lactobacillus-fermented milk on lipid metabolism in hamsters fed on high-cholesterol diet. Appl. Microbiol. Biotechnol. 2006, 71, 238–245. [Google Scholar] [CrossRef]
- Pan, T.M.; Chiu, C.H.; Guu, Y.K. Characterization of Lactobacillus isolates from pickled vegetables for use as dietary or pickle adjuncts. Foods Food Ingred. J. Jpn. 2002, 206, 45–51. [Google Scholar]
- Tsai, Y.Y.; Chu, L.H.; Lee, C.L.; Pan, T.M. Atherosclerosis-preventing activity of lactic acid bacteria-fermented milk-soymilk supplemented with Momordica charatia. J. Agric. Food Chem. 2009, 57, 2065–2071. [Google Scholar] [CrossRef]
- Lee, B.H.; Lo, Y.H.; Pan, T.M. Anti-obesity activity of Lactobacillus fermented soy milk products. J. Funct. Foods 2013, 5, 905–913. [Google Scholar] [CrossRef]
- Tung, Y.T.; Lee, B.H.; Liu, C.F.; Pan, T.M. Optimization of culture condition for ACEI and GABA production by lactic acid bacteria. J. Food Sci. 2011, 76, M585–M591. [Google Scholar] [CrossRef]
- Shanmuganathan, B.; Sathya, S.; Balasubramaniam, B.; Balamurugan, K.; Devi, K.P. Amyloid-β induced neuropathological actions are suppressed by Padina gymnospora (Phaeophyceae) and its active constituent α-bisabolol in Neuro2a cells and transgenic Caenorhabditis elegans Alzheimer’s model. Nitric Oxide 2019, 91, 52–66. [Google Scholar] [CrossRef]
- Manczak, M.; Mao, P.; Calkins, M.; Cornea, A.; Arubala, R.P.; Murphy, M.P.; Szeto, H.H.; Park, B.; Reddy, P.H. Mitochondria-targeted antioxidants protect against Abeta toxicity in Alzheimer’s disease neurons. J. Alzheimers Dis. 2010, 20, S609–S631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Yuan, B.; Wu, Y.; Gong, Y.; Guo, W.; Fu, S.; Luan, Y.; Wang, W. Sodium butyrate protects N2a cells against Aβ toxicity in vitro. Media. Inflamm. 2020, 7605160. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Liu, P.C.; Liu, R.; Wu, X. Dual AO/EB staining to detect apoptosis in osteosarcoma cells compared with flow cytometry. Med. Sci. Mon. Basic Res. 2015, 21, 15–20. [Google Scholar]
- Wang, X.; Zhou, X.; Li, G.; Zhang, Y.; Wu, Y.; Song, W. Modifications and trafficking of APP in the pathogenesis of Alzheimer’s disease. Fr. Mol. Neurosci. 2017, 10, 294. [Google Scholar] [CrossRef] [PubMed]
- Seki, T.; Kanagawa, M.; Kobayashi, K.; Kowa, H.; Yahata, N.; Maruyama, K.; Iwata, N.; Inoue, H.; Toda, T. Galectin 3-binding protein suppresses amyloid-β production by modulating β-cleavage of amyloid precursor protein. J. Biol. Chem. 2020, 295, 3678–3691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esquerda-Canals, G.; Montoliu-Gaya, L.; Guell-Bosch, J.; Villegas, S. Mouse models of Alzheimer’s disease. J. Alzheimer Dis. 2017, 57, 1171–1183. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Wang, T.; Hu, X.; Luo, J.; Li, W.; Wu, X.; Duan, Y.; Fin, F. Administration of Lactobacillus helveticus NS8 improves behavioral, cognitive, and biochemical aberrations caused by chronic restraint stress. Neuroscience 2015, 310, 561–577. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Hu, X.; Liang, S.; Li, W.; Wu, X.; Wang, L.; Jin, F. Lactobacillus fermentum NS9 restores the antibiotic induced physiological and psychological abnormalities in rats. Benef. Microbes 2015, 6, 707–717. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.J.; Lee, K.E.; Kim, J.K.; Kim, D.H. Suppression of gut dysbiosis by Bifidobacterium longum alleviates cognitive decline in 5XFAD transgenic and aged mice. Sci. Rep. 2019, 9, 11814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azm, S.A.N.; Djazayery, A.; Safa, M.; Azami, K.; Ahmadvand, B.; Sabbaghziarani, F.; Sharifzadeh, M.; Vafa, M.R. Lactobacillus and Bifidobacterium ameliorate memory and learning deficits and oxidative stress in Aβ (1-42) injected rats. Appl. Physiol. Nutr. Metab. 2018, 43, 718–726. [Google Scholar]
- Choi, J.; Kim, Y.K.; Han, P.L. Extracellular vesicles derived from Lactobacillus plantarum increase BDNF expression in cultured hippocampal neurons and produce antidepressant-like effects in mice. Exp. Neurobiol. 2019, 28, 158–171. [Google Scholar] [CrossRef] [PubMed]
- Sayago-Ayerdi, S.G.; Zamora-Gasga, V.M.; Venema, K. Prebiotic effect of predigested mango peel on gut microbiota assessed in a dynamic in vitro model of the human colon (TIM-2). Food Res. Int. 2019, 118, 89–95. [Google Scholar] [CrossRef]
- Sayago-Ayerdi, S.G.; Venema, K.; Tabernero, M.; Sarria, B.; Bravo, L.L.; Mateos, R. Bioconversion by gut microbiota of predigested mango (Mangifera indica L)’Ataulfo’ peel polyphenols assessed in a dynamic (TIM-2) in vitro model of the human colon. Food Res. Int. 2021, 139, 109963. [Google Scholar] [CrossRef]
- Sun, J.; Xu, J.; Yang, B.; Chen, K.; Kong, Y.; Fang, N.; Gong, T.; Wang, F.; Ling, Z.; Liu, J. Effect of Clostridium butyricum against mnicroglia-mediated neuroinflammation in Alzheimer’s disease via regulating gut microbiota and metabolites butyrate. Mol. Nutr. Food Res. 2020, 64, e1900636. [Google Scholar] [CrossRef]
- Lee, C.C.; Lee, B.H.; Wu, S.C. Actinidia callosa peel (kiwi fruit) ethanol extracts protected neural cells apoptosis induced by methylglyoxal through Nrf2 activation. Pharm. Biol. 2014, 52, 628–636. [Google Scholar] [CrossRef]
- Domert, J.; Rao, S.B.; Agholme, L.; Brorsson, A.C.; Marcusson, J.; Hallbeck, M.; Nath, S. Spreading of amyloid-β peptides via neuritic cell-to cell transfer is dependent on insufficient cellular clearance. Neurobiol. Dis. 2014, 65, 82–92. [Google Scholar] [CrossRef] [Green Version]
- Hsu, W.H.; Lin, Y.C.; Chen, B.R.; Wu, S.C.; Lee, B.H. The neuronal protection of a zinc-binding protein isolated from oyster. Food Chem. Toxicol. 2018, 114, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Cui, Z.; Li, Y.H.; Hsu, W.H.; Lee, B.H. In vitro and in vivo anticancer activity of pardaxin against proliferation and growth of oral squamous cell carcinoma. Mar. Drugs 2016, 14, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, W.H.; Lee, B.H.; Pan, T.M. Leptin-induced mitochondrial fusion mediates hepatic lipid accumulation. Int. J. Obesity 2015, 39, 1750–1756. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fermentation Time (Day) | L. acidophilus (BCRC14079) | L. planatrum Subsp. plantarum (BCRC10367) | L. paracasei Subsp. paracasei (BCRC80062) |
| pH | |||
| 0 | 6.55 ± 0.03 a | 6.55 ± 0.03 a | 6.55 ± 0.03 a |
| 1 | 5.13 ± 0.07 b | 5.74 ± 0.04 a,b | 5.48 ± 0.05 b |
| 3 | 4.45 ± 0.11 b | 5.06 ± 0.06 b | 4.89 ± 0.03 c |
| 5 | 4.03 ± 0.06 c | 4.71 ± 0.05 c | 4.33 ± 0.03 c |
| Fermentation Time (Day) | Titratable Acid (%) | ||
| 0 | 0.07 ± 0.01 c | 0.07 ± 0.01 c | 0.07 ± 0.01 c |
| 1 | 0.13 ± 0.01 c | 0.09 ± 0.01 c | 0.11 ± 0.02 c |
| 3 | 0.43 ± 0.02 b | 0.23 ± 0.01 b | 0.27 ± 0.02 b |
| 5 | 0.56 ± 0.03 a | 0.37 ± 0.01 a | 0.42 ± 0.01 a |
| Mango Peel | pH | Titratable Acid (%) |
|---|---|---|
| 25% | 4.53 ± 0.05 a | 0.41 ± 0.02 c |
| 30% | 4.11 ± 0.03 a,b | 0.68 ± 0.03 b |
| 35% | 3.89 ± 0.02 b | 0.85 ± 0.09 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, B.-H.; Hsu, W.-H.; Hou, C.-Y.; Chien, H.-Y.; Wu, S.-C. The Protection of Lactic Acid Bacteria Fermented-Mango Peel against Neuronal Damage Induced by Amyloid-Beta. Molecules 2021, 26, 3503. https://doi.org/10.3390/molecules26123503
Lee B-H, Hsu W-H, Hou C-Y, Chien H-Y, Wu S-C. The Protection of Lactic Acid Bacteria Fermented-Mango Peel against Neuronal Damage Induced by Amyloid-Beta. Molecules. 2021; 26(12):3503. https://doi.org/10.3390/molecules26123503
Chicago/Turabian StyleLee, Bao-Hong, Wei-Hsuan Hsu, Chih-Yao Hou, Hao-Yuan Chien, and She-Ching Wu. 2021. "The Protection of Lactic Acid Bacteria Fermented-Mango Peel against Neuronal Damage Induced by Amyloid-Beta" Molecules 26, no. 12: 3503. https://doi.org/10.3390/molecules26123503
APA StyleLee, B. -H., Hsu, W. -H., Hou, C. -Y., Chien, H. -Y., & Wu, S. -C. (2021). The Protection of Lactic Acid Bacteria Fermented-Mango Peel against Neuronal Damage Induced by Amyloid-Beta. Molecules, 26(12), 3503. https://doi.org/10.3390/molecules26123503