
The Concept of Folic Acid in Health and Disease

 , ,
, ,  ,
,
Abstract
:1. Introduction
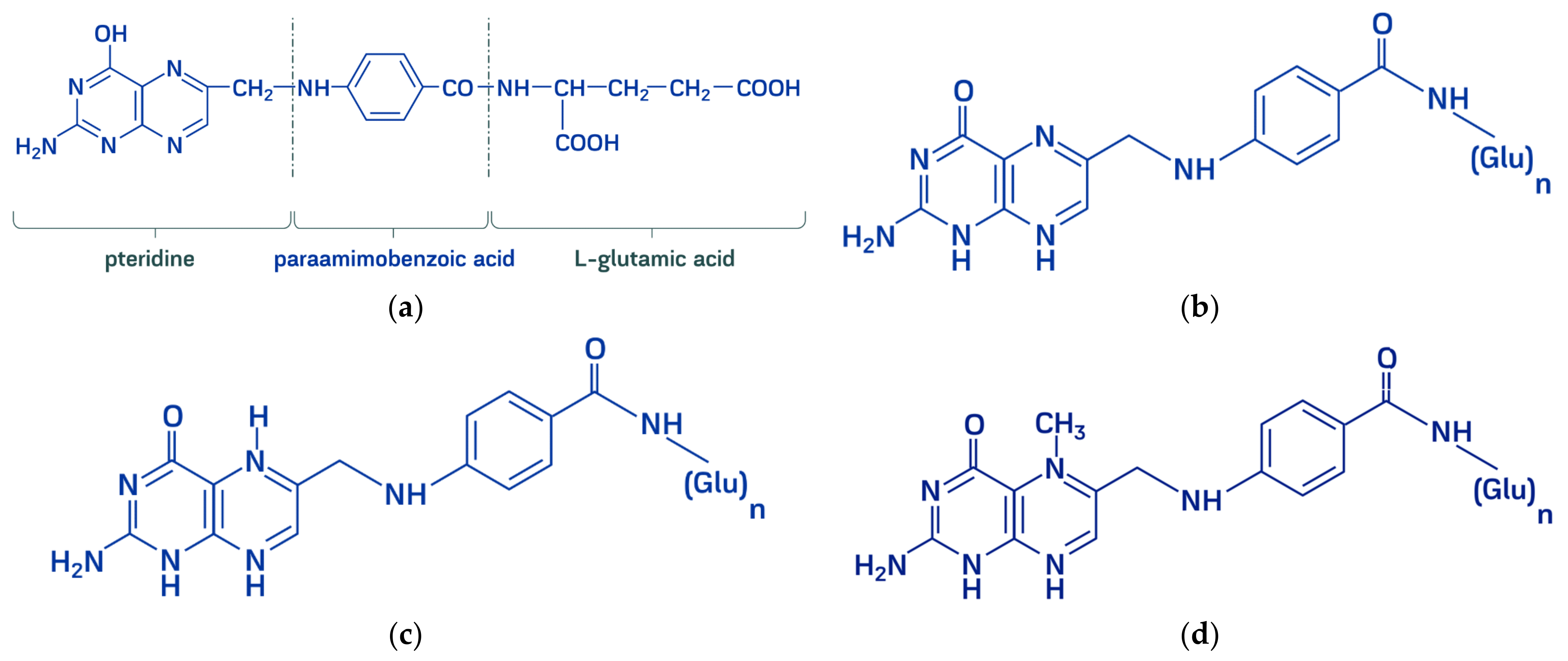
2. Natural Sources of Folates
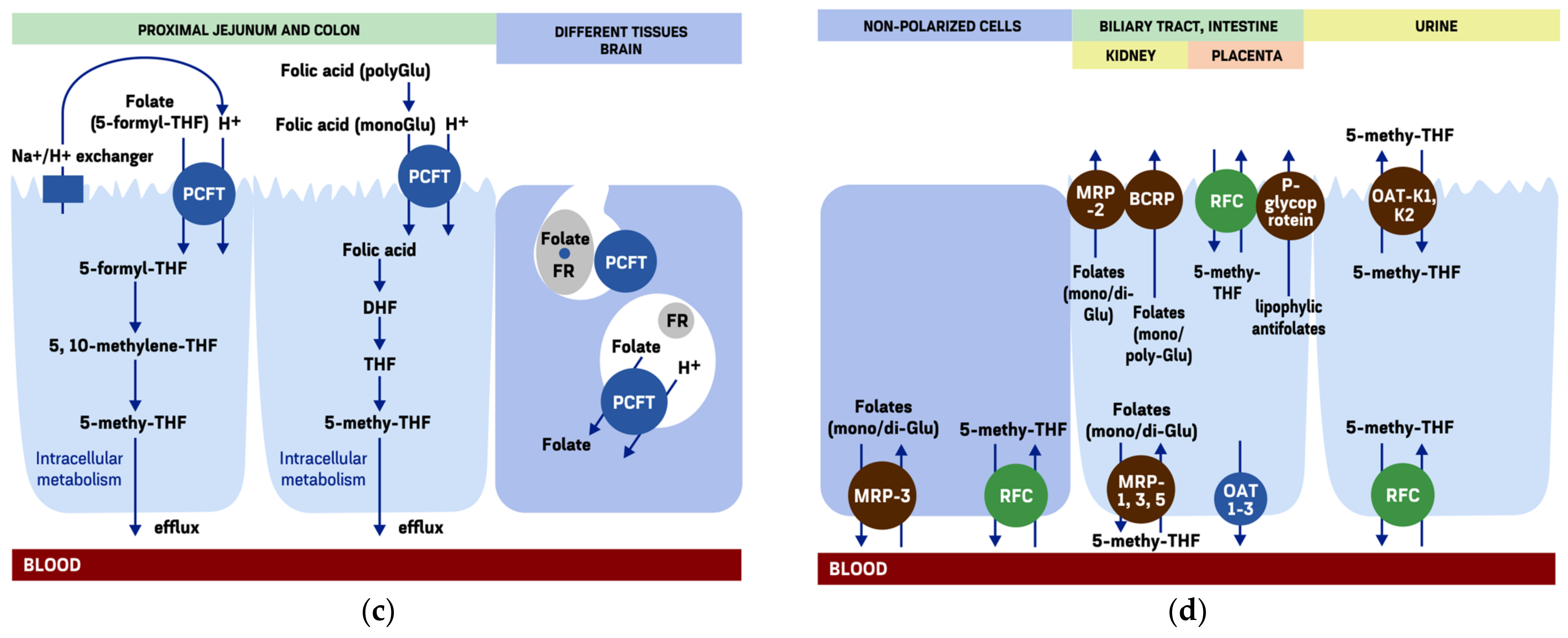
3. Intestinal Folate Absorption
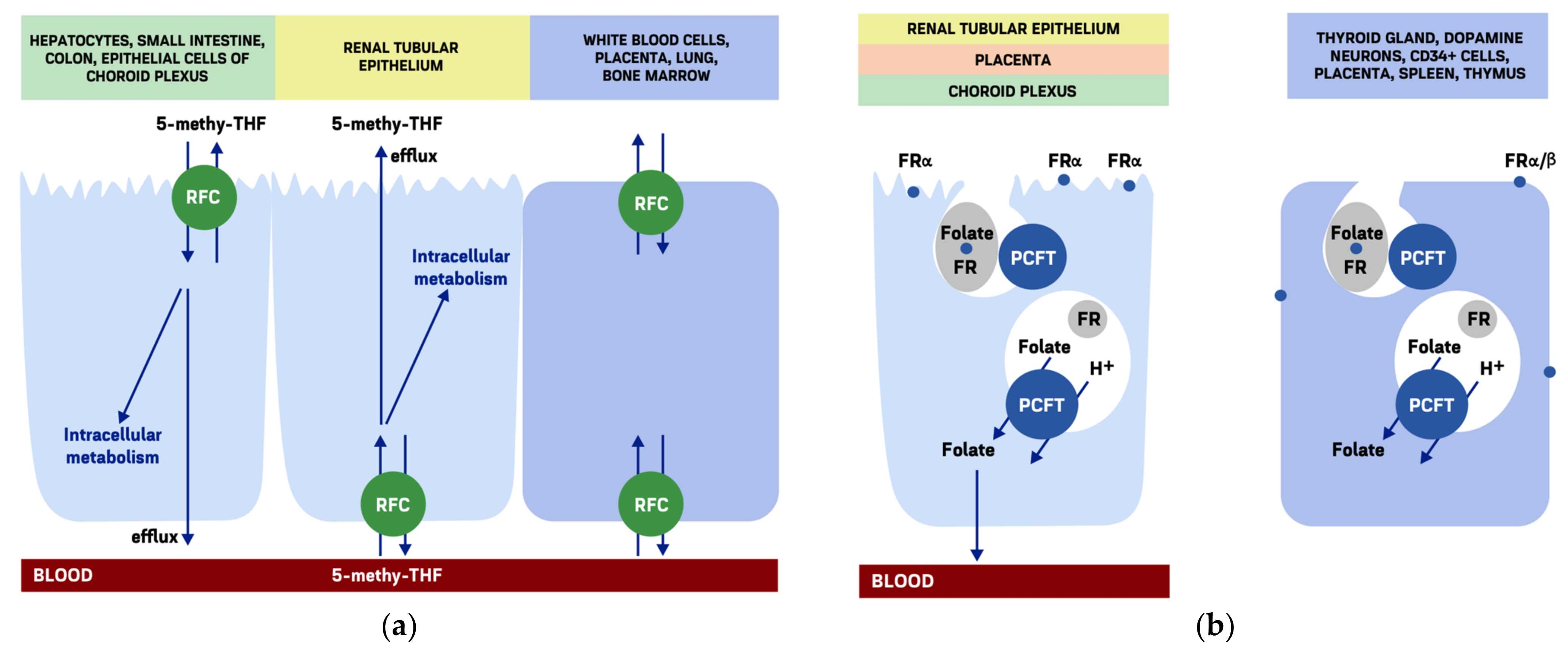
3.1. Folate Cellular Uptake
3.2. Reduced Folate Carrier
3.3. Folate Receptors
3.4. The Proton-Coupled Folate Transporter
4. Extracellular Efflux
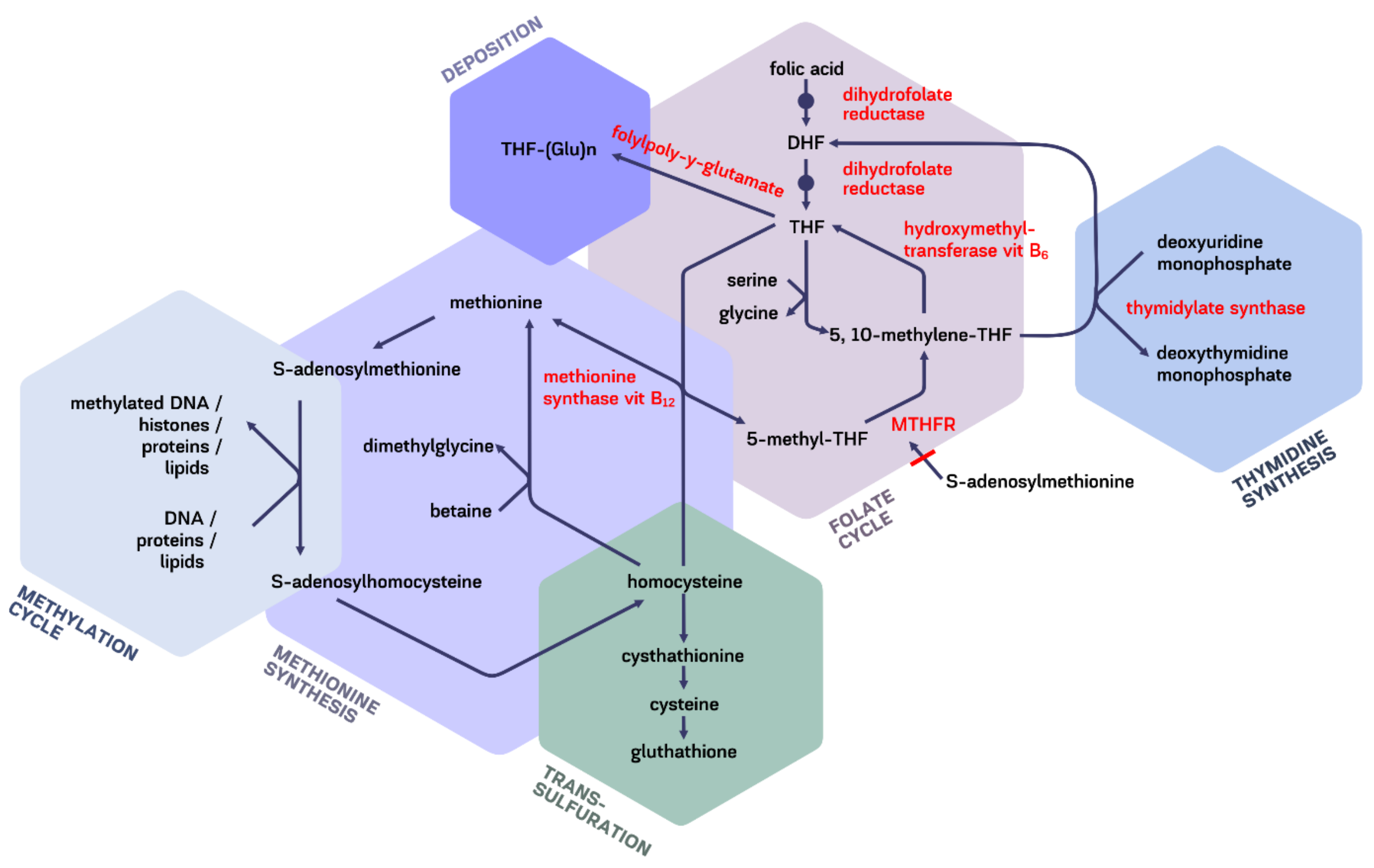
5. Intracellular Transformations
Folates in One-Carbon Pathway
6. Problem of Folate Deficiency and Oversupplementation
7. Laboratory Folate Metabolism Assessment
8. The Main Clinical Syndromes of Folate Deficiency
8.1. Hereditary Syndromes
8.2. MTHFR Deficit
8.3. Folylpoly-γ-Glutamate Synthetase Deficit
8.4. Acquired Folate Deficiency
9. Risks Associated with Folate Deficiency
9.1. Macrocytic Anemia, Mucositis, Infertility, Muscular Weakness
9.2. Cardiovascular Disease
9.3. Neurological Problems
9.4. Cancer
10. Antifolates
Resistance to Methotrexate and Other Antifolates and Their Toxicity
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Molloy, A.M. Folate bioavailability and health. Int. J. Vitam Nutr. Res. 2002, 72, 46–52. [Google Scholar] [CrossRef]
- Feirer, N.; Fuqua, C. Pterin function in bacteria. Pteridines 2017, 28. [Google Scholar] [CrossRef] [Green Version]
- Mahendran, R.; Thandeeswaran, M.; Kiran, G.; Arulkumar, M.; Ayub Nawaz, K.A.; Jabastin, J.; Janani, B.; Anto Thomas, T.; Angayarkanni, J. Evaluation of pterin, a promising drug candidate from cyanide degrading bacteria. Curr. Microbiol. 2018, 75, 684–693. [Google Scholar] [CrossRef]
- Thöny, B.; Auerbach, G.; Blau, N. Tetrahydrobiopterin biosynthesis, regeneration and functions. Biochem. J. 2000, 347, 1–16. [Google Scholar] [CrossRef]
- Burton, M.; Abanobi, C.; Wang, K.; Ma, Y.; Rasche, M. Substrate specificity analysis of dihydrofolate/dihydromethanopterin reductase homologs in methylotrophic α-proteobacteria. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Colette Daubner, S.; Lanzas, R.O. Pteridines☆. In Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar] [CrossRef]
- Gamayurova, V.S.; Rzhechitskaya, L.E. Food Chemistry: A Textbook for University Students: A Manual, an Electronic Source for Network Distribution. Available online: https://bookonlime.ru/product/pishchevaya-himiya-uchebnik-dlya-studentov-vuzov (accessed on 7 May 2021).
- Hamed, E.; Attia, M.S.; Bassiouny, K. Synthesis, spectroscopic and thermal characterization of copper(II) and iron(III) complexes of folic acid and their absorption efficiency in the blood. Bioinorg. Chem. Appl. 2009, 2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engevik, M.A.; Morra, C.N.; Röth, D.; Engevik, K.; Spinler, J.K.; Devaraj, S.; Crawford, S.E.; Estes, M.K.; Kalkum, M.; Versalovic, J. Microbial metabolic capacity for intestinal folate production and modulation of host folate receptors. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.M.; Saulnier, D.M.A.; Spinler, J.K.; Hemarajata, P.; Gao, C.; Jones, S.E.; Grimm, A.; Balderas, M.A.; Burstein, M.D.; Morra, C.; et al. FolC2-mediated folate metabolism contributes to suppression of inflammation by probiotic lactobacillus reuteri. Microbiologyopen 2016, 5, 802–818. [Google Scholar] [CrossRef]
- National Institutes of Health. Folate.Fact Sheet for Health Professionals. Available online: https://ods.od.nih.gov/factsheets/Folate-HealthProfessional/ (accessed on 12 June 2021).
- Rossi, M.; Amaretti, A.; Raimondi, S. Folate production by probiotic bacteria. Nutrients 2011, 3, 118–134. [Google Scholar] [CrossRef] [Green Version]
- Noiriel, A.; Naponelli, V.; Gregory, J.F.; Hanson, A.D. Pterin and folate salvage. Plants and escherichia coli lack capacity to reduce oxidized pterins. Plant. Physiol. 2007, 143, 1101–1109. [Google Scholar] [CrossRef] [Green Version]
- Maynard, C.; Cummins, I.; Green, J.; Weinkove, D. A bacterial route for folic acid supplementation. BMC Biol. 2018, 16, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aufreiter, S.; Gregory, J.F.; Pfeiffer, C.M.; Fazili, Z.; Kim, Y.-I.; Marcon, N.; Kamalaporn, P.; Pencharz, P.B.; O’Connor, D.L. Folate is absorbed across the colon of adults: Evidence from cecal infusion of (13)C-labeled [6S]-5-formyltetrahydrofolic acid. Am. J. Clin. Nutr. 2009, 90, 116–123. [Google Scholar] [CrossRef]
- Coppedè, F.; Lorenzoni, V.; Migliore, L. The reduced folate carrier (RFC-1) 80A>G polymorphism and maternal risk of having a child with down syndrome: A meta-analysis. Nutrients 2013, 5, 2551–2563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Z.; Matherly, L.H. Biology of the major facilitative folate transporters SLC19A1 and SLC46A1. Curr. Top. Membr. 2014, 73, 175–204. [Google Scholar] [CrossRef] [Green Version]
- Desmoulin, S.K.; Hou, Z.; Gangjee, A.; Matherly, L.H. The human proton-coupled folate transporter: Biology and therapeutic applications to cancer. Cancer Biol. Ther. 2012, 13, 1355–1373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, C.; Aufreiter, S.; Georgiou, C.J.; Hoque, M.T.; Finnell, R.H.; O’Connor, D.L.; Goldman, I.D.; Bendayan, R. Upregulation of reduced folate carrier by vitamin D enhances brain folate uptake in mice lacking folate receptor alpha. Proc. Natl. Acad. Sci. USA 2019, 116, 17531–17540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, Y.-S. Folate receptor-targeted diagnostics and therapeutics for inflammatory diseases. Immune Netw. 2016, 16, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Low, P.S.; Henne, W.A.; Doorneweerd, D.D. Discovery and development of folic-acid-based receptor targeting for imaging and therapy of cancer and inflammatory diseases. Acc. Chem. Res. 2008, 41, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Matherly, L.H.; Goldman, I.D. Membrane transporters and folate homeostasis: Intestinal absorption and transport into systemic compartments and tissues. Expert. Rev. Mol. Med. 2009, 11, e4. [Google Scholar] [CrossRef] [Green Version]
- Whetstine, J.R.; Flatley, R.M.; Matherly, L.H. The human reduced folate carrier gene is ubiquitously and differentially expressed in normal human tissues: Identification of seven non-coding exons and characterization of a novel promoter. Biochem. J. 2002, 367, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Rajgopal, A.; Edmondnson, A.; Goldman, I.D.; Zhao, R. SLC19A3 encodes a second thiamine transporter ThTr2. Biochim. Biophys. Acta 2001, 1537, 175–178. [Google Scholar] [CrossRef] [Green Version]
- Hou, Z.; Wu, J.; Ye, J.; Cherian, C.; Matherly, L.H. Substrate-specific binding and conformational changes involving Ser313 and transmembrane domain 8 of the human reduced folate carrier, as determined by site-directed mutagenesis and protein cross-linking. Biochem. J. 2010, 430, 265–274. [Google Scholar] [CrossRef] [Green Version]
- Abramson, J.; Smirnova, I.; Kasho, V.; Verner, G.; Kaback, H.R.; Iwata, S. Structure and mechanism of the lactose permease of escherichia coli. Science 2003, 301, 610–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Lemieux, M.J.; Song, J.; Auer, M.; Wang, D.-N. Structure and mechanism of the glycerol-3-phosphate transporter from escherichia coli. Science 2003, 301, 616–620. [Google Scholar] [CrossRef] [Green Version]
- Dixon, K.H.; Lanpher, B.C.; Chiu, J.; Kelley, K.; Cowan, K.H. A novel CDNA restores reduced folate carrier activity and methotrexate sensitivity to transport deficient cells. J. Biol. Chem. 1994, 269, 17–20. [Google Scholar] [CrossRef]
- Henderson, G.B.; Zevely, E.M. Structural requirements for anion substrates of the methotrexate transport system in L1210 cells. Arch. Biochem. Biophys. 1983, 221, 438–446. [Google Scholar] [CrossRef]
- Hou, Z.; Cherian, C.; Drews, J.; Wu, J.; Matherly, L.H. Identification of the minimal functional unit of the homo-oligomeric human reduced folate carrier. J. Biol. Chem. 2010, 285, 4732–4740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.B.; Huang, W.; Chancy, C.; Ganapathy, V. Regulation of the reduced-folate transporter by nitric oxide in cultured human retinal pigment epithelial cells. Biochem. Biophys. Res. Commun. 1999, 257, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Hou, Z.; Stapels, S.E.; Haska, C.L.; Matherly, L.H. Localization of a substrate binding domain of the human reduced folate carrier to transmembrane domain 11 by radioaffinity labeling and cysteine-substituted accessibility methods. J. Biol. Chem. 2005, 280, 36206–36213. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.; Gao, F.; Goldman, I.D. Reduced folate carrier transports thiamine monophosphate: An alternative route for thiamine delivery into mammalian cells. Am. J. Physiol. Cell Physiol. 2002, 282, C1512–C1517. [Google Scholar] [CrossRef] [Green Version]
- Ifergan, I.; Jansen, G.; Assaraf, Y.G. The reduced folate carrier (RFC) is cytotoxic to cells under conditions of severe folate deprivation. RFC as a double edged sword in folate homeostasis. J. Biol. Chem. 2008, 283, 20687–20695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matherly, L.H.; Hou, Z.; Deng, Y. Human reduced folate carrier: Translation of basic biology to cancer etiology and therapy. Cancer Metastasis Rev. 2007, 26, 111–128. [Google Scholar] [CrossRef]
- Zhao, R.; Gao, F.; Hanscom, M.; Goldman, I.D. A prominent low-PH methotrexate transport activity in human solid tumors: Contribution to the preservation of methotrexate pharmacologic activity in HeLa cells lacking the reduced folate carrier. Clin. Cancer Res. 2004, 10, 718–727. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhao, R.; Goldman, I.D. Characterization of a folate transporter in HeLa cells with a low PH optimum and high affinity for pemetrexed distinct from the reduced folate carrier. Clin. Cancer Res. 2004, 10, 6256–6264. [Google Scholar] [CrossRef] [Green Version]
- Fry, D.W.; White, J.C.; Goldman, I.D. Effects of 2,4-dinitrophenol and other metabolic inhibitors on the bidirectional carrier fluxes, net transport, and intracellular binding of methotrexate in ehrlich ascites tumor cells. Cancer Res. 1980, 40, 3669–3673. [Google Scholar] [PubMed]
- Dembo, M.; Sirotnak, F.M.; Moccio, D.M. Effects of metabolic deprivation on methotrexate transport in L1210 leukemia cells: Further evidence for separate influx and efflux systems with different energetic requirements. J. Membr. Biol. 1984, 78, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.C.; Zhang, L.; Proefke, S.A.; Matherly, L.H. Effects of the loss of capacity for N-glycosylation on the transport activity and cellular localization of the human reduced folate carrier. Biochim. Biophys. Acta 1998, 1375, 6–12. [Google Scholar] [CrossRef] [Green Version]
- Brigle, K.E.; Spinella, M.J.; Sierra, E.E.; Goldman, I.D. Organization of the murine reduced folate carrier gene and identification of variant splice forms. Biochim. Biophys. Acta 1997, 1353, 191–198. [Google Scholar] [CrossRef]
- Zhang, L.; Wong, S.C.; Matherly, L.H. Transcript heterogeneity of the human reduced folate carrier results from the use of multiple promoters and variable splicing of alternative upstream exons. Biochem. J. 1998, 332, 773–780. [Google Scholar] [CrossRef] [Green Version]
- Payton, S.G.; Haska, C.L.; Flatley, R.M.; Ge, Y.; Matherly, L.H. Effects of 5’ untranslated region diversity on the posttranscriptional regulation of the human reduced folate carrier. Biochim. Biophys. Acta 2007, 1769, 131–138. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.; Russell, R.G.; Wang, Y.; Liu, L.; Gao, F.; Kneitz, B.; Edelmann, W.; Goldman, I.D. Rescue of embryonic lethality in reduced folate carrier-deficient mice by maternal folic acid supplementation reveals early neonatal failure of hematopoietic organs. J. Biol. Chem. 2001, 276, 10224–10228. [Google Scholar] [CrossRef] [Green Version]
- Bailey, L.B.; Rampersaud, G.C.; Kauwell, G.P.A. Folic acid supplements and fortification affect the risk for neural tube defects, vascular disease and cancer: Evolving science. J. Nutr. 2003, 133, 1961S–1968S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, R.; Diop-Bove, N.; Visentin, M.; Goldman, I.D. Mechanisms of membrane transport of folates into cells and across epithelia. Annu. Rev. Nutr. 2011, 31, 177–201. [Google Scholar] [CrossRef] [Green Version]
- Sirotnak, F.M.; Tolner, B. Carrier-mediated membrane transport of folates in mammalian cells. Annu. Rev. Nutr. 1999, 19, 91–122. [Google Scholar] [CrossRef]
- Brzezińska, A.; Wińska, P.; Balińska, M. Cellular aspects of folate and antifolate membrane transport. Acta Biochim. Pol. 2000, 47, 735–749. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Ge, Y.; Cabelof, D.C.; Aboukameel, A.; Heydari, A.R.; Mohammad, R.; Matherly, L.H. Structure and regulation of the murine reduced folate carrier gene: Identification of four noncoding exons and promoters and regulation by dietary folates. J. Biol. Chem. 2005, 280, 5588–5597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.H.; Pain, J.; Sirotnak, F.M. Alteration of folate analogue transport inward after induced maturation of HL-60 Leukemia cells. Molecular properties of the transporter in an overproducing variant and evidence for down-regulation of its synthesis in maturating cells. J. Biol. Chem. 1992, 267, 6628–6634. [Google Scholar] [CrossRef]
- Alam, C.; Hoque, M.T.; Finnell, R.H.; Goldman, I.D.; Bendayan, R. Regulation of reduced folate carrier (RFC) by vitamin D receptor at the blood-brain barrier. Mol. Pharm. 2017, 14, 3848–3858. [Google Scholar] [CrossRef] [PubMed]
- Hamid, A.; Wani, N.A.; Kaur, J. New perspectives on folate transport in relation to alcoholism-induced folate malabsorption--association with epigenome stability and cancer development. FEBS J. 2009, 276, 2175–2191. [Google Scholar] [CrossRef] [PubMed]
- Chango, A.; Emery-Fillon, N.; de Courcy, G.P.; Lambert, D.; Pfister, M.; Rosenblatt, D.S.; Nicolas, J.P. A polymorphism (80G->A) in the reduced folate carrier gene and its associations with folate status and homocysteinemia. Mol. Genet. Metab. 2000, 70, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Ranganathan, P.; McLeod, H.L. Methotrexate pharmacogenetics: The first step toward individualized therapy in rheumatoid arthritis. Arthritis Rheum. 2006, 54, 1366–1377. [Google Scholar] [CrossRef]
- Polymorphism G80A in the Reduced Folate Carrier Gene and Its Relationship to Methotrexate Plasma Levels and Outcome of Childhood Acute Lymphoblastic Leukemia. Available online: https://www.pubfacts.com/detail/12411325/Polymorphism-G80A-in-the-reduced-folate-carrier-gene-and-its-relationship-to-methotrexate-plasma-lev (accessed on 7 March 2021).
- Dervieux, T.; Kremer, J.; Lein, D.O.; Capps, R.; Barham, R.; Meyer, G.; Smith, K.; Caldwell, J.; Furst, D.E. Contribution of common polymorphisms in reduced folate carrier and gamma-glutamylhydrolase to methotrexate polyglutamate levels in patients with rheumatoid arthritis. Pharmacogenetics 2004, 14, 733–739. [Google Scholar] [CrossRef]
- Relton, C.L.; Wilding, C.S.; Jonas, P.A.; Lynch, S.A.; Tawn, E.J.; Burn, J. Genetic susceptibility to neural tube defect pregnancy varies with offspring phenotype. Clin. Genet. 2003, 64, 424–428. [Google Scholar] [CrossRef] [PubMed]
- De Marco, P.; Calevo, M.G.; Moroni, A.; Merello, E.; Raso, A.; Finnell, R.H.; Zhu, H.; Andreussi, L.; Cama, A.; Capra, V. Reduced folate carrier polymorphism (80A-->G) and neural tube defects. Eur. J. Hum. Genet. 2003, 11, 245–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, G.M.; Lammer, E.J.; Zhu, H.; Baker, M.W.; Neri, E.; Finnell, R.H. Maternal periconceptional vitamin use, genetic variation of infant reduced folate carrier (A80G), and risk of spina bifida. Am. J. Med. Genet. 2002, 108, 1–6. [Google Scholar] [CrossRef]
- Huang, X.; Gao, Y.; He, J.; Cai, J.; Ta, N.; Jiang, H.; Zhu, J.; Zheng, J. The association between RFC1 G80A polymorphism and cancer susceptibility: Evidence from 33 studies. J. Cancer 2016, 7, 144–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forat-Yazdi, M.; Hosseini-Biouki, F.; Salehi, J.; Neamatzadeh, H.; Masoumi Dehshiri, R.; Sadri, Z.; Ghanizadeh, F.; Sheikhpour, R.; Zare-Zardini, H. Association between RFC1 G80A polymorphism and acute lymphoblastic leukemia: A review and meta-analysis of 10 studies. Iran. J. Ped. Hematol. Oncol. 2016, 6, 52–63. [Google Scholar]
- Elnakat, H.; Ratnam, M. Distribution, functionality and gene regulation of folate receptor isoforms: Implications in targeted therapy. Adv. Drug Deliv. Rev. 2004, 56, 1067–1084. [Google Scholar] [CrossRef]
- Lu, Y.; Low, P.S. Folate-mediated delivery of macromolecular anticancer therapeutic agents. Adv. Drug Deliv. Rev. 2002, 54, 675–693. [Google Scholar] [CrossRef]
- Kamen, B.A.; Smith, A.K. A Review of folate receptor alpha cycling and 5-methyltetrahydrofolate accumulation with an emphasis on cell models in vitro. Adv. Drug Deliv. Rev. 2004, 56, 1085–1097. [Google Scholar] [CrossRef]
- Jansen, G. Receptor- and carrier-mediated transport systems for folates and antifolates. In Antifolate Drugs in Cancer Therapy; Jackman, A.L., Ed.; Cancer Drug Discovery and Development; Humana Press: Totowa, NJ, USA, 1999. [Google Scholar] [CrossRef]
- Brigle, K.E.; Spinella, M.J.; Westin, E.H.; Goldman, I.D. Increased expression and characterization of two distinct folate binding proteins in murine erythroleukemia cells. Biochem. Pharmacol. 1994, 47, 337–345. [Google Scholar] [CrossRef]
- Wang, X.; Shen, F.; Freisheim, J.H.; Gentry, L.E.; Ratnam, M. Differential stereospecificities and affinities of folate receptor isoforms for folate compounds and antifolates. Biochem. Pharmacol. 1992, 44, 1898–1901. [Google Scholar] [CrossRef] [PubMed]
- Spinella, M.J.; Brigle, K.E.; Sierra, E.E.; Goldman, I.D. Distinguishing between folate receptor-alpha-mediated transport and reduced folate carrier-mediated transport in L1210 leukemia cells. J. Biol. Chem. 1995, 270, 7842–7849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Unno, K.; Hyjek, E.; Liu, H.; Zimmerman, T.; Karmakar, S.; Putt, K.S.; Shen, J.; Low, P.S.; Wickrema, A. Expression of functional folate receptors in multiple myeloma. Leuk. Lymphoma 2018, 59, 2982–2989. [Google Scholar] [CrossRef]
- Low, P.S.; Antony, A.C. Folate receptor-targeted drugs for cancer and inflammatory diseases. Adv. Drug Deliv. Rev. 2004, 56, 1055–1058. [Google Scholar] [CrossRef] [PubMed]
- Gennet, N.; Tamburini, C.; Nan, X.; Li, M. FolR1: A novel cell surface marker for isolating midbrain dopamine neural progenitors and nascent dopamine neurons. Sci. Rep. 2016, 6, 32488. [Google Scholar] [CrossRef]
- Smith, S.B.; Kekuda, R.; Gu, X.; Chancy, C.; Conway, S.J.; Ganapathy, V. Expression of folate receptor alpha in the mammalian retinol pigmented epithelium and retina. Invest. Ophthalmol. Vis. Sci. 1999, 40, 840–848. [Google Scholar] [PubMed]
- Chancy, C.D.; Kekuda, R.; Huang, W.; Prasad, P.D.; Kuhnel, J.M.; Sirotnak, F.M.; Roon, P.; Ganapathy, V.; Smith, S.B. Expression and differential polarization of the reduced-folate transporter-1 and the folate receptor alpha in mammalian retinal pigment epithelium. J. Biol. Chem. 2000, 275, 20676–20684. [Google Scholar] [CrossRef] [Green Version]
- Parker, N.; Turk, M.J.; Westrick, E.; Lewis, J.D.; Low, P.S.; Leamon, C.P. Folate receptor expression in carcinomas and normal tissues determined by a quantitative radioligand binding assay. Anal. Biochem. 2005, 338, 284–293. [Google Scholar] [CrossRef]
- Birn, H.; Nielsen, S.; Christensen, E.I. Internalization and apical-to-basolateral transport of folate in rat kidney proximal tubule. Am. J. Physiol. 1997, 272, F70–F78. [Google Scholar] [CrossRef]
- Ratnam, M.; Marquardt, H.; Duhring, J.L.; Freisheim, J.H. Homologous membrane folate binding proteins in human placenta: Cloning and sequence of a CDNA. Biochemistry 1989, 28, 8249–8254. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zheng, X.; Behm, F.G.; Ratnam, M. Differentiation-independent retinoid induction of folate receptor type beta, a potential tumor target in myeloid leukemia. Blood 2000, 96, 3529–3536. [Google Scholar] [CrossRef]
- Reddy, J.A.; Haneline, L.S.; Srour, E.F.; Antony, A.C.; Clapp, D.W.; Low, P.S. Expression and functional characterization of the beta-isoform of the folate receptor on CD34(+) cells. Blood 1999, 93, 3940–3948. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.F.; Wang, H.; Behm, F.G.; Mathew, P.; Wu, M.; Booth, R.; Ratnam, M. Folate receptor type beta is a neutrophilic lineage marker and is differentially expressed in myeloid leukemia. Cancer 1999, 85, 348–357. [Google Scholar] [CrossRef]
- Shen, F.; Wu, M.; Ross, J.F.; Miller, D.; Ratnam, M. Folate receptor type gamma is primarily a secretory protein due to lack of an efficient signal for glycosylphosphatidylinositol modification: Protein characterization and cell type specificity. Biochemistry 1995, 34, 5660–5665. [Google Scholar] [CrossRef]
- Mayor, S.; Rothberg, K.G.; Maxfield, F.R. Sequestration of GPI-anchored proteins in caveolae triggered by cross-linking. Science 1994, 264, 1948–1951. [Google Scholar] [CrossRef] [PubMed]
- Kelemen, L.E. The role of folate receptor alpha in cancer development, progression and treatment: Cause, consequence or innocent bystander? Int. J. Cancer 2006, 119, 243–250. [Google Scholar] [CrossRef]
- Kamen, B.A.; Wang, M.T.; Streckfuss, A.J.; Peryea, X.; Anderson, R.G. Delivery of folates to the cytoplasm of MA104 cells is mediated by a surface membrane receptor that recycles. J. Biol. Chem. 1988, 263, 13602–13609. [Google Scholar] [CrossRef]
- Rothberg, K.G.; Ying, Y.S.; Kolhouse, J.F.; Kamen, B.A.; Anderson, R.G. The glycophospholipid-linked folate receptor internalizes folate without entering the clathrin-coated pit endocytic pathway. J. Cell Biol. 1990, 110, 637–649. [Google Scholar] [CrossRef] [Green Version]
- Anderson, R.G.; Kamen, B.A.; Rothberg, K.G.; Lacey, S.W. Potocytosis: Sequestration and transport of small molecules by caveolae. Science 1992, 255, 410–411. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Chen, H.; Vlahov, I.R.; Cheng, J.-X.; Low, P.S. Characterization of the PH of folate receptor-containing endosomes and the rate of hydrolysis of internalized acid-labile folate-drug conjugates. J. Pharmacol. Exp. Ther. 2007, 321, 462–468. [Google Scholar] [CrossRef] [PubMed]
- Prasad, P.D.; Mahesh, V.B.; Leibach, F.H.; Ganapathy, V. Functional coupling between a bafilomycin a1-sensitive proton pump and a probenecid-sensitive folate transporter in human placental choriocarcinoma cells. Biochim. Biophys. Acta 1994, 1222, 309–314. [Google Scholar] [CrossRef]
- Salazar, M.D.; Ratnam, M. The folate receptor: What does it promise in tissue-targeted therapeutics? Cancer Metastasis Rev. 2007, 26, 141–152. [Google Scholar] [CrossRef]
- Monick, S.; Mohanty, V.; Khan, M.; Yerneni, G.; Kumar, R.; Cantu, J.; Ichi, S.; Xi, G.; Singh, B.R.; Tomita, T.; et al. A phenotypic switch of differentiated glial cells to dedifferentiated cells is regulated by folate receptor α. Stem Cells 2019, 37, 1441–1454. [Google Scholar] [CrossRef] [Green Version]
- Kamen, B.A.; Smith, A.K.; Anderson, R.G. The folate receptor works in tandem with a probenecid-sensitive carrier in MA104 cells in vitro. J. Clin. Investig. 1991, 87, 1442–1449. [Google Scholar] [CrossRef] [PubMed]
- Low, P.S.; Singhal, S.; Srinivasarao, M. Fluorescence-guided surgery of cancer: Applications, tools and perspectives. Curr. Opin. Chem. Biol. 2018, 45, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Toffoli, G.; Cernigoi, C.; Russo, A.; Gallo, A.; Bagnoli, M.; Boiocchi, M. Overexpression of folate binding protein in ovarian cancers. Int. J. Cancer 1997, 74, 193–198. [Google Scholar] [CrossRef]
- Lee, J.Y.K.; Cho, S.S.; Zeh, R.; Pierce, J.T.; Martinez-Lage, M.; Adappa, N.D.; Palmer, J.N.; Newman, J.G.; Learned, K.O.; White, C.; et al. Folate receptor overexpression can be visualized in real time during pituitary adenoma endoscopic transsphenoidal surgery with near-infrared imaging. J. Neurosurg. 2018, 129, 390–403. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Ho, W.; Zhang, X.; Bertrand, N.; Farokhzad, O. Cancer nanomedicine: From targeted delivery to combination therapy. Trends Mol. Med. 2015, 21, 223–232. [Google Scholar] [CrossRef] [Green Version]
- Cheung, A.; Bax, H.J.; Josephs, D.H.; Ilieva, K.M.; Pellizzari, G.; Opzoomer, J.; Bloomfield, J.; Fittall, M.; Grigoriadis, A.; Figini, M.; et al. Targeting folate receptor alpha for cancer treatment. Oncotarget 2016, 7, 52553–52574. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.; Guo, J.; Li, C.; Wang, Z. A current review of folate receptor alpha as a potential tumor target in non-small-cell lung cancer. Drug Des. Devel. Ther. 2015, 9, 4989–4996. [Google Scholar] [CrossRef] [Green Version]
- Song, S.; Li, M.; Gong, X.; Han, H.; Zhou, Y.; Wang, L.; Shuang, S.; Dong, C. Controlled release of curcumin via folic acid conjugated magnetic drug delivery system. Chem. Res. Chin. Univ. 2018, 34, 203–211. [Google Scholar] [CrossRef]
- Moret, F.; Scheglmann, D.; Reddi, E. Folate-targeted PEGylated liposomes improve the selectivity of PDT with meta-tetra(hydroxyphenyl)chlorin (m-THPC). Photochem. PhotoBiol. Sci. 2013, 12, 823–834. [Google Scholar] [CrossRef] [PubMed]
- De Boer, E.; Crane, L.M.A.; van Oosten, M.; van der Vegt, B.; van der Sluis, T.; Kooijman, P.; Low, P.S.; van der Zee, A.G.J.; Arts, H.J.G.; van Dam, G.M.; et al. Folate receptor-beta has limited value for fluorescent imaging in ovarian, breast and colorectal cancer. PLoS ONE 2015, 10, e0135012. [Google Scholar] [CrossRef] [PubMed]
- Saikawa, Y.; Price, K.; Hance, K.W.; Chen, T.Y.; Elwood, P.C. Structural and functional analysis of the human KB cell folate receptor gene P4 promoter: Cooperation of three clustered Sp1-binding sites with initiator region for basal promoter activity. Biochemistry 1995, 34, 9951–9961. [Google Scholar] [CrossRef] [PubMed]
- Antony, A.; Tang, Y.-S.; Khan, R.A.; Biju, M.P.; Xiao, X.; Li, Q.-J.; Sun, X.-L.; Jayaram, H.N.; Stabler, S.P. Translational upregulation of folate receptors is mediated by homocysteine via RNA-heterogeneous nuclear ribonucleoprotein E1 interactions. J. Clin. Investig. 2004, 113, 285–301. [Google Scholar] [CrossRef] [Green Version]
- Elwood, P.C.; Nachmanoff, K.; Saikawa, Y.; Page, S.T.; Pacheco, P.; Roberts, S.; Chung, K.N. The divergent 5’ termini of the alpha human folate receptor (HFR) MRNAs originate from two tissue-specific promoters and alternative splicing: Characterization of the alpha HFR gene structure. Biochemistry 1997, 36, 1467–1478. [Google Scholar] [CrossRef]
- Hsueh, C.T.; Dolnick, B.J. Regulation of folate-binding protein gene expression by DNA methylation in methotrexate-resistant KB cells. Biochem. Pharmacol. 1994, 47, 1019–1027. [Google Scholar] [CrossRef]
- Kelley, K.M.M.; Rowan, B.G.; Ratnam, M. Modulation of the folate receptor alpha gene by the estrogen receptor: Mechanism and implications in tumor targeting. Cancer Res. 2003, 63, 2820–2828. [Google Scholar]
- Tran, T.; Shatnawi, A.; Zheng, X.; Kelley, K.M.M.; Ratnam, M. Enhancement of folate receptor alpha expression in tumor cells through the glucocorticoid receptor: A promising means to improved tumor detection and targeting. Cancer Res. 2005, 65, 4431–4441. [Google Scholar] [CrossRef] [Green Version]
- Piedrahita, J.A.; Oetama, B.; Bennett, G.D.; van Waes, J.; Kamen, B.A.; Richardson, J.; Lacey, S.W.; Anderson, R.G.; Finnell, R.H. Mice lacking the folic acid-binding protein folbp1 are defective in early embryonic development. Nat. Genet. 1999, 23, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Qiu, A.; Jansen, M.; Sakaris, A.; Min, S.H.; Chattopadhyay, S.; Tsai, E.; Sandoval, C.; Zhao, R.; Akabas, M.H.; Goldman, I.D. Identification of an intestinal folate transporter and the molecular basis for hereditary folate malabsorption. Cell 2006, 127, 917–928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakai, Y.; Inoue, K.; Abe, N.; Hatakeyama, M.; Ohta, K.; Otagiri, M.; Hayashi, Y.; Yuasa, H. Functional characterization of human proton-coupled folate transporter/heme carrier protein 1 heterologously expressed in mammalian cells as a folate transporter. J. Pharmacol. Exp. Ther. 2007, 322, 469–476. [Google Scholar] [CrossRef] [Green Version]
- Sirotnak, F.M.; Chello, P.L.; Moccio, D.M.; Kisliuk, R.L.; Combepine, G.; Gaumont, Y.; Montgomery, J.A. Stereospecificity at carbon 6 of fomyltetrahydrofolate as a competitive inhibitor of transport and cytotoxicity of methotrexate in vitro. Biochem. Pharmacol. 1979, 28, 2993–2997. [Google Scholar] [CrossRef]
- Qiu, A.; Min, S.H.; Jansen, M.; Malhotra, U.; Tsai, E.; Cabelof, D.C.; Matherly, L.H.; Zhao, R.; Akabas, M.H.; Goldman, I.D. Rodent intestinal folate transporters (SLC46A1): Secondary structure, functional properties, and response to dietary folate restriction. Am. J. Physiol. Cell Physiol. 2007, 293, C1669–C1678. [Google Scholar] [CrossRef] [Green Version]
- Inoue, K.; Nakai, Y.; Ueda, S.; Kamigaso, S.; Ohta, K.; Hatakeyama, M.; Hayashi, Y.; Otagiri, M.; Yuasa, H. Functional characterization of PCFT/HCP1 as the Molecular entity of the carrier-mediated intestinal folate transport system in the rat model. Am. J. Physiol. Gastrointest Liver Physiol. 2008, 294, G660–G668. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhao, R.; Russell, R.G.; Goldman, I.D. Localization of the murine reduced folate carrier as assessed by immunohistochemical analysis. Biochim. Biophys. Acta 2001, 1513, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, V.S.; Marchant, J.S.; Said, H.M. Apical membrane targeting and trafficking of the human proton-coupled transporter in polarized epithelia. Am. J. Physiol. Cell Physiol. 2008, 294, C233–C240. [Google Scholar] [CrossRef] [PubMed]
- Hiraoka, M.; Kagawa, Y. Genetic Polymorphisms and folate status. Congenit. Anom. 2017, 57, 142–149. [Google Scholar] [CrossRef] [Green Version]
- Unal, E.S.; Zhao, R.; Qiu, A.; Goldman, I.D. N-linked glycosylation and its impact on the electrophoretic mobility and function of the human proton-coupled folate transporter (HsPCFT). Biochim. Biophys. Acta 2008, 1778, 1407–1414. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.; Goldman, I.D. Folate and thiamine transporters mediated by facilitative carriers (SLC19A1-3 and SLC46A1) and folate receptors. Mol. Asp. Med. 2013, 34, 373–385. [Google Scholar] [CrossRef] [Green Version]
- Kronn, D.; Goldman, I.D. Hereditary folate malabsorption. In GeneReviews®; Adam, M.P., Ardinger, H.H., Pagon, R.A., Wallace, S.E., Bean, L.J., Mirzaa, G., Amemiya, A., Eds.; University of Washington: Seattle, WA, USA, 1993. [Google Scholar]
- Goldman, I.D.; Matherly, L.H. The cellular pharmacology of methotrexate. Pharmacol. Ther. 1985, 28, 77–102. [Google Scholar] [CrossRef]
- Assaraf, Y.G. The role of multidrug resistance efflux transporters in antifolate resistance and folate homeostasis. Drug Resist. Updat. 2006, 9, 227–246. [Google Scholar] [CrossRef] [PubMed]
- Wielinga, P.; Hooijberg, J.H.; Gunnarsdottir, S.; Kathmann, I.; Reid, G.; Zelcer, N.; van der Born, K.; de Haas, M.; van der Heijden, I.; Kaspers, G.; et al. The human multidrug resistance protein MRP5 transports folates and can mediate cellular resistance against antifolates. Cancer Res. 2005, 65, 4425–4430. [Google Scholar] [CrossRef] [Green Version]
- Kruh, G.D.; Belinsky, M.G. The MRP family of drug efflux pumps. Oncogene 2003, 22, 7537–7552. [Google Scholar] [CrossRef] [PubMed]
- Matherly, L.H. Molecular and cellular biology of the human reduced folate carrier. Prog. Nucleic Acid Res. Mol. Biol. 2001, 67, 131–162. [Google Scholar] [CrossRef] [PubMed]
- Keppler, D. Multidrug resistance proteins (MRPs, ABCCs): Importance for pathophysiology and drug therapy. Handb. Exp. Pharmacol. 2011, No. 201, 299–323. [Google Scholar] [CrossRef]
- Sodani, K.; Patel, A.; Kathawala, R.J.; Chen, Z.-S. Multidrug resistance associated proteins in multidrug resistance. Chin. J. Cancer 2012, 31, 58–72. [Google Scholar] [CrossRef] [Green Version]
- McGuire, J.J.; Russell, C.A.; Balinska, M. Human cytosolic and mitochondrial folylpolyglutamate synthetase are electrophoretically distinct. expression in antifolate-sensitive and -resistant human cell lines. J. Biol. Chem. 2000, 275, 13012–13016. [Google Scholar] [CrossRef] [Green Version]
- Gu, X.; Manautou, J.E. Regulation of hepatic ABCC transporters by xenobiotics and in disease states. Drug Metab. Rev. 2010, 42, 482–538. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, H.M.; Al-Abd, A.M.; El-Dine, R.S.; El-Halawany, A.M. P-glycoprotein inhibitors of natural origin as potential tumor chemo-sensitizers: A review. J. Adv. Res. 2015, 6, 45–62. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.W.; Mason, J.B. Folate and carcinogenesis: An integrated scheme. J. Nutr. 2000, 130, 129–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ifergan, I.; Shafran, A.; Jansen, G.; Hooijberg, J.H.; Scheffer, G.L.; Assaraf, Y.G. Folate deprivation results in the loss of breast cancer resistance protein (BCRP/ABCG2) expression. A role for BCRP in cellular folate homeostasis. J. Biol. Chem. 2004, 279, 25527–25534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nigam, S.K.; Bush, K.T.; Martovetsky, G.; Ahn, S.-Y.; Liu, H.C.; Richard, E.; Bhatnagar, V.; Wu, W. The organic anion transporter (OAT) family: A systems biology perspective. Physiol. Rev. 2015, 95, 83–123. [Google Scholar] [CrossRef]
- Masuda, S. Functional characteristics and pharmacokinetic significance of kidney-specific organic anion transporters, OAT-K1 and OAT-K2, in the urinary excretion of anionic drugs. Drug Metab. Pharmacokinet. 2003, 18, 91–103. [Google Scholar] [CrossRef] [Green Version]
- Rizwan, A.N.; Burckhardt, G. Organic Anion transporters of the SLC22 family: Biopharmaceutical, Physiological, and pathological roles. Pharm. Res. 2007, 24, 450–470. [Google Scholar] [CrossRef]
- Raz, S.; Stark, M.; Assaraf, Y.G. Folylpoly-γ-glutamate synthetase: A key determinant of folate homeostasis and antifolate resistance in cancer. Drug Resist. Updat. 2016, 28, 43–64. [Google Scholar] [CrossRef]
- Balinska, M.; Nimec, Z.; Galivan, J. Characteristics of methotrexate polyglutamate formation in cultured hepatic cells. Arch. Biochem. Biophys. 1982, 216, 466–476. [Google Scholar] [CrossRef]
- Cannella, A.C.; O’Dell, J.R. Kelley and Firestein’s Textbook of Rheumatology, 10th ed.; Elsevier Health Sciences: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Serot, J.M.; Christmann, D.; Dubost, T.; Béne, M.C.; Faure, G.C. CSF-folate levels are decreased in late-onset AD patients. J. Neural Transm. 2001, 108, 93–99. [Google Scholar] [CrossRef]
- Glynn, S.A.; Albanes, D. Folate and cancer: A review of the literature. Nutr. Cancer 1994, 22, 101–119. [Google Scholar] [CrossRef]
- Spinella, M.J.; Brigle, K.E.; Freemantle, S.J.; Sierra, E.E.; Goldman, I.D. Comparison of methotrexate polyglutamylation in L1210 leukemia cells when influx is mediated by the reduced folate carrier or the folate receptor. Lack of evidence for influx route-specific effects. Biochem. Pharmacol. 1996, 52, 703–712. [Google Scholar] [CrossRef]
- Locasale, J.W. Serine, glycine and the one-carbon cycle: Cancer metabolism in full circle. Nat. Rev. Cancer 2013, 13, 572–583. [Google Scholar] [CrossRef] [Green Version]
- Relton, C.L.; Wilding, C.S.; Laffling, A.J.; Jonas, P.A.; Burgess, T.; Binks, K.; Tawn, E.J.; Burn, J. Low erythrocyte folate status and polymorphic variation in folate-related genes are associated with risk of neural tube defect pregnancy. Mol. Genet. Metab. 2004, 81, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Stover, P.J. One-carbon metabolism-genome interactions in folate-associated pathologies. J. Nutr. 2009, 139, 2402–2405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swanson, J.M.; Entringer, S.; Buss, C.; Wadhwa, P.D. Developmental origins of health and disease: Environmental exposures. Semin. Reprod. Med. 2009, 27, 391–402. [Google Scholar] [CrossRef] [Green Version]
- Crider, K.S.; Yang, T.P.; Berry, R.J.; Bailey, L.B. Folate and DNA methylation: A review of molecular mechanisms and the evidence for folate’s role. Adv. Nutr. 2012, 3, 21–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matherly, L.H.; Hou, Z.; Gangjee, A. The promise and challenges of exploiting the proton-coupled folate transporter for selective therapeutic targeting of cancer. Cancer Chemother. Pharmacol. 2018, 81, 1–15. [Google Scholar] [CrossRef]
- Metabolism Lecture Notes. Available online: http://watcut.uwaterloo.ca/webnotes/Metabolism/index.html%20 (accessed on 8 March 2021).
- World Health Organization. Serum and Red Blood Cell Folate Concentrations for Assessing Folate Status in Populations; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- Ferrari, A.; Torrezan, G.T.; Carraro, D.M.; Aguiar Junior, S. Association of folate and vitamins involved in the 1-carbon cycle with polymorphisms in the methylenetetrahydrofolate reductase gene (MTHFR) and global DNA methylation in patients with colorectal cancer. Nutrients 2019, 11, 1368. [Google Scholar] [CrossRef] [Green Version]
- Dwyer, J.T.; Wiemer, K.L.; Dary, O.; Keen, C.L.; King, J.C.; Miller, K.B.; Philbert, M.A.; Tarasuk, V.; Taylor, C.L.; Gaine, P.C.; et al. Fortification and health: Challenges and opportunities. Adv. Nutr. 2015, 6, 124–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Nutrition for Health and Development: A Global Agenda for Combating Malnutrition; World Health Organization: Geneva, Switzerland, 2000. [Google Scholar]
- Anaemias, W.S.G.; On, N.; World Health Organization. Nutritional Anaemias: Report of a WHO Scientific Group [Meeting Held in Geneva from 13 to 17 March 1967]; World Health Organization: Geneva, Switzerland, 1968. [Google Scholar]
- Mursleen, M.T.; Riaz, S. Implication of homocysteine in diabetes and impact of folate and vitamin B12 in diabetic population. Diabetes Metab. Syndr. 2017, 11 (Suppl. 1) (Suppl. 1), S141–S146. [Google Scholar] [CrossRef]
- Iqbal, M.P.; Ishaq, M.; Kazmi, K.A.; Yousuf, F.A.; Mehboobali, N.; Ali, S.A.; Khan, A.H.; Waqar, M.A. Role of vitamins B6, B12 and folic acid on hyperhomocysteinemia in a Pakistani population of patients with acute myocardial infarction. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 100–108. [Google Scholar] [CrossRef]
- Rogers, L.M.; Cordero, A.M.; Pfeiffer, C.M.; Hausman, D.B.; Tsang, B.L.; De-Regil, L.M.; Rosenthal, J.; Razzaghi, H.; Wong, E.C.; Weakland, A.P.; et al. Global folate status in women of reproductive age: A systematic review with emphasis on methodological issues. Ann. N. Y. Acad. Sci. 2018, 1431, 35–57. [Google Scholar] [CrossRef]
- Obeid, R.; Oexle, K.; Rißmann, A.; Pietrzik, K.; Koletzko, B. Folate status and health: Challenges and opportunities. J. Perinat. Med. 2016, 44, 261–268. [Google Scholar] [CrossRef]
- Yila, T.A.; Araki, A.; Sasaki, S.; Miyashita, C.; Itoh, K.; Ikeno, T.; Yoshioka, E.; Kobayashi, S.; Goudarzi, H.; Baba, T.; et al. Predictors of folate status among pregnant japanese women: The hokkaido study on environment and children’s health, 2002–2012. Br. J. Nutr. 2016, 115, 2227–2235. [Google Scholar] [CrossRef] [Green Version]
- Teng, Y.; Hu, J.; Dong, S.; Zhang, L.; Lu, X.; Yang, H. Concentration of red blood cell folate and the analysis of folic acid supplement optimum time and dose in first trimester. J. Hyg. Res. 2017, 46, 569–578. [Google Scholar]
- Murray, L.K.; Smith, M.J.; Jadavji, N.M. Maternal oversupplementation with folic acid and its impact on neurodevelopment of offspring. Nutr. Rev. 2018, 76, 708–721. [Google Scholar] [CrossRef] [PubMed]
- Crider, K.S.; Qi, Y.P.; Devine, O.; Tinker, S.C.; Berry, R.J. Modeling the impact of folic acid fortification and supplementation on red blood cell folate concentrations and predicted neural tube defect risk in the United States: Have we reached optimal prevention? Am. J. Clin. Nutr. 2018, 107, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Czeizel, A.E.; Dudás, I. Prevention of the first occurrence of neural-tube defects by periconceptional vitamin supplementation. N. Engl. J. Med. 1992, 327, 1832–1835. [Google Scholar] [CrossRef] [PubMed]
- MRC Vitamin Study Research Group. Prevention of neural tube defects: Results of the medical research council vitamin study. Lancet 1991, 338, 131–137. [Google Scholar] [CrossRef]
- Giovannucci, E.; Stampfer, M.J.; Colditz, G.A.; Hunter, D.J.; Fuchs, C.; Rosner, B.A.; Speizer, F.E.; Willett, W.C. Multivitamin use, folate, and colon cancer in women in the nurses’ health study. Ann. Intern. Med. 1998, 129, 517–524. [Google Scholar] [CrossRef]
- Stanisławska-Sachadyn, A.; Borzyszkowska, J.; Krzemiński, M.; Janowicz, A.; Dziadziuszko, R.; Jassem, J.; Rzyman, W.; Limon, J. Folate/homocysteine metabolism and lung cancer risk among smokers. PLoS ONE 2019, 14, e0214462. [Google Scholar] [CrossRef]
- Savini, C.; Yang, R.; Savelyeva, L.; Göckel-Krzikalla, E.; Hotz-Wagenblatt, A.; Westermann, F.; Rösl, F. Folate repletion after deficiency induces irreversible genomic and transcriptional changes in human papillomavirus type 16 (HPV16)-immortalized human keratinocytes. Int. J. Mol. Sci. 2019, 20, 1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, S.; Tang, Y.-S.; Kusumanchi, P.; Stabler, S.P.; Zhang, Y.; Antony, A.C. Folate deficiency facilitates genomic integration of human papillomavirus type 16 DNA in vivo in a novel mouse model for rapid oncogenic transformation of human keratinocytes. J. Nutr. 2018, 148, 389–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanley, M.P.; Aladelokun, O.; Kadaveru, K.; Rosenberg, D.W. Methyl donor deficiency blocks colorectal cancer development by affecting key metabolic pathways. Cancer Prev. Res. 2020, 13, 1–14. [Google Scholar] [CrossRef]
- Niemann, B.; Nemitz, A.; Werner, J.; Mai, H.D.; Steinberg, P.; Lampen, A.; Ehlers, A. Folic acid modulates cancer-associated micro RNAs and inflammatory mediators in neoplastic and non-neoplastic colonic cells in a different way. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef]
- Cravo, M.L.; Pinto, A.G.; Chaves, P.; Cruz, J.A.; Lage, P.; Nobre Leitão, C.; Costa Mira, F. Effect of folate supplementation on DNA methylation of rectal mucosa in patients with colonic adenomas: Correlation with nutrient intake. Clin. Nutr. 1998, 17, 45–49. [Google Scholar] [CrossRef]
- Cravo, M.; Pinto, R.; Fidalgo, P.; Chaves, P.; Glória, L.; Nobre-Leitão, C.; Costa Mira, F. Global DNA hypomethylation occurs in the early stages of intestinal type gastric carcinoma. Gut 1996, 39, 434–438. [Google Scholar] [CrossRef] [Green Version]
- Wallace, K.; Grau, M.V.; Levine, A.J.; Shen, L.; Hamdan, R.; Chen, X.; Gui, J.; Haile, R.W.; Barry, E.L.; Ahnen, D.; et al. Association between folate levels and CpG island hypermethylation in normal colorectal mucosa. Cancer Prev. Res. 2010, 3, 1552–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beard, C.M.; Panser, L.A.; Katusic, S.K. Is excess folic acid supplementation a risk factor for autism? Med. Hypotheses 2011, 77, 15–17. [Google Scholar] [CrossRef] [PubMed]
- Egorova, O.; Myte, R.; Schneede, J.; Hägglöf, B.; Bölte, S.; Domellöf, E.; Ivars A’roch, B.; Elgh, F.; Ueland, P.M.; Silfverdal, S.-A. Maternal blood folate status during early pregnancy and occurrence of autism. Spectrum disorder in offspring: A study of 62 serum biomarkers. Mol. Autism. 2020, 11, 7. [Google Scholar] [CrossRef] [Green Version]
- Deshmukh, U.; Katre, P.; Yajnik, C.S. Influence of maternal vitamin B12 and folate on growth and insulin resistance in the offspring. Nestle Nutr. Inst. Workshop. Ser. 2013, 74, 145–154. [Google Scholar] [CrossRef]
- Metz, J. Appropriate use of tests for folate and vitamin B12 deficiency. Aust Prescr 1999, 22, 16–18. [Google Scholar] [CrossRef]
- Socha, D.S.; DeSouza, S.I.; Flagg, A.; Sekeres, M.; Rogers, H.J. Severe megaloblastic anemia: Vitamin deficiency and other causes. Cleve Clin. J. Med. 2020, 87, 153–164. [Google Scholar] [CrossRef] [Green Version]
- Bailey, L.B.; Stover, P.J.; McNulty, H.; Fenech, M.F.; Gregory, J.F.; Mills, J.L.; Pfeiffer, C.M.; Fazili, Z.; Zhang, M.; Ueland, P.M.; et al. Biomarkers of nutrition for development-folate review. J. Nutr. 2015, 145, 1636S–1680S. [Google Scholar] [CrossRef] [Green Version]
- Quadros, E.V.; Sequeira, J.M.; Brown, W.T.; Mevs, C.; Marchi, E.; Flory, M.; Jenkins, E.C.; Velinov, M.T.; Cohen, I.L. Folate receptor autoantibodies are prevalent in children diagnosed with autism. spectrum disorder, their normal siblings and parents. Autism. Res. 2018, 11, 707–712. [Google Scholar] [CrossRef]
- Ramaekers, V.T.; Thöny, B.; Sequeira, J.M.; Ansseau, M.; Philippe, P.; Boemer, F.; Bours, V.; Quadros, E.V. Folinic acid treatment for schizophrenia associated with folate receptor autoantibodies. Mol. Genet. Metab. 2014, 113, 307–314. [Google Scholar] [CrossRef]
- Colson, N.J.; Naug, H.L.; Nikbakht, E.; Zhang, P.; McCormack, J. The impact of MTHFR 677 C/T genotypes on folate status markers: A meta-analysis of folic acid intervention studies. Eur. J. Nutr. 2017, 56, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Nasri, K.; Midani, F.; Kallel, A.; Ben Jemaa, N.; Aloui, M.; Boulares, M.; Lassoued, M.; Ben Halima, M.; Ben Wafi, S.; Soussi, M.; et al. Association of MTHFR C677T, MTHFR A1298C, and MTRR A66G polymorphisms with neural tube defects in tunisian parents. Pathobiology 2019, 86, 190–200. [Google Scholar] [CrossRef]
- Wani, N.A.; Kaur, J. Reduced levels of folate transporters (PCFT and RFC) in membrane lipid rafts result in colonic folate malabsorption in chronic alcoholism. J. Cell Physiol. 2011, 226, 579–587. [Google Scholar] [CrossRef]
- Krupenko, S.A.; Krupenko, N.I. ALDH1L1 and ALDH1L2 folate regulatory enzymes in cancer. Adv. Exp. Med. Biol. 2018, 1032, 127–143. [Google Scholar] [CrossRef] [PubMed]
- Mahabir, S.; Ettinger, S.; Johnson, L.; Baer, D.J.; Clevidence, B.A.; Hartman, T.J.; Taylor, P.R. Measures of adiposity and body fat distribution in relation to serum folate levels in postmenopausal women in a feeding study. Eur. J. Clin. Nutr. 2008, 62, 644–650. [Google Scholar] [CrossRef] [Green Version]
- Geiker, N.R.W.; Veller, M.; Kjoelbaek, L.; Jakobsen, J.; Ritz, C.; Raben, A.; Astrup, A.; Lorenzen, J.K.; Larsen, L.H.; Bügel, S. Effect of low energy diet for eight weeks to adults with overweight or obesity on folate, retinol, vitamin B12, D and E status and the degree of inflammation: A post hoc analysis of a randomized intervention trial. Nutr. Metab. 2018, 15, 24. [Google Scholar] [CrossRef] [Green Version]
- Frame-Peterson, L.A.; Megill, R.D.; Carobrese, S.; Schweitzer, M. Nutrient deficiencies are common prior to bariatric surgery. Nutr. Clin. Pract. 2017, 32, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Charatcharoenwitthaya, P.; Levy, C.; Angulo, P.; Keach, J.; Jorgensen, R.; Lindor, K.D. Open-label pilot study of folic acid in patients with nonalcoholic steatohepatitis. Liver Int. 2007, 27, 220–226. [Google Scholar] [CrossRef]
- Sid, V.; Siow, Y.L.; O, K. Role of folate in nonalcoholic fatty liver disease. Can. J. Physiol. Pharmacol. 2017, 95, 1141–1148. [Google Scholar] [CrossRef] [PubMed]
- Mahamid, M.; Mahroum, N.; Bragazzi, N.L.; Shalaata, K.; Yavne, Y.; Adawi, M.; Amital, H.; Watad, A. Folate and B12 levels correlate with histological severity in NASH patients. Nutrients 2018, 10. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.; Zhu, J.; Meng, D.; Yu, C.; Li, Y. Association of homocysteine level with biopsy-proven non-alcoholic fatty liver disease: A meta-analysis. J. Clin. Biochem. Nutr. 2016, 58, 76–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, X.; Li, J.; Cui, Y.; Liu, Z.; Zhao, Z.; Ge, J.; Guan, D.; Hu, J.; Wang, Y.; Zhang, F.; et al. Effect of folic acid intervention on ALT concentration in hypertensives without known hepatic disease: A randomized, double-blind, controlled trial. Eur. J. Clin. Nutr. 2012, 66, 541–548. [Google Scholar] [CrossRef]
- Samodelov, S.L.; Gai, Z.; Kullak-Ublick, G.A.; Visentin, M. Renal reabsorption of folates: Pharmacological and toxicological snapshots. Nutrients 2019, 11, 2353. [Google Scholar] [CrossRef] [Green Version]
- Capelli, I.; Cianciolo, G.; Gasperoni, L.; Zappulo, F.; Tondolo, F.; Cappuccilli, M.; La Manna, G. Folic acid and vitamin B12 administration in CKD, why not? Nutrients 2019, 11, 383. [Google Scholar] [CrossRef] [Green Version]
- Khalifa, A.M.; Almoallem, R.A.; Hamoud, S.; Mohammed, A.E.; Alanazi, Y.N.R. Prevalence of anemia: A study on inpatients’ records with chronic kidney disease at prince mutaib hospital, Al-Jouf province, Saudi Arabia. Int. J. Med. Dev. Ctries. 2020, 4, 206–212. [Google Scholar] [CrossRef]
- Sies, H.; Stahl, W. Vitamins E and C, beta-carotene, and other carotenoids as antioxidants. Am. J. Clin. Nutr. 1995, 62, 1315S–1321S. [Google Scholar] [CrossRef]
- Beydoun, M.A.; Dore, G.A.; Canas, J.A.; Beydoun, H.A.; Zonderman, A.B. Helicobacter pylori seropositivity’s association with markers of iron, 1-carbon metabolism, and antioxidant status among US adults: A structural equations modeling approach. PLoS ONE 2015, 10, e0121390. [Google Scholar] [CrossRef] [Green Version]
- Raut, S.C.; Patil, V.W.; Dalvi, S.M.; Bakhshi, G.D. Helicobacter pylori gastritis, a presequeale to coronary plaque. Clin. Pract. 2015, 5, 717. [Google Scholar] [CrossRef] [Green Version]
- Akcam, M. Helicobacter pylori and micronutrients. Indian Pediatr. 2010, 47, 119–126. [Google Scholar] [CrossRef]
- Nejatinamini, S.; Debenham, B.J.; Clugston, R.D.; Mawani, A.; Parliament, M.; Wismer, W.V.; Mazurak, V.C. Poor vitamin status is associated with skeletal muscle loss and mucositis in head and neck cancer patients. Nutrients 2018, 10, 1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demirkiran, D.; Kurt, H.; Yüksel, M.; Sari, Y. Pancytopenia and mucositis encountered in the early stages of methotrexate treatment. Eur. J. Health Sci. 2016, 2, 1–3. [Google Scholar]
- Hwang, S.Y.; Kang, Y.J.; Sung, B.; Jang, J.Y.; Hwang, N.L.; Oh, H.J.; Ahn, Y.R.; Kim, H.J.; Shin, J.H.; Yoo, M.-A.; et al. Folic acid is necessary for proliferation and differentiation of C2C12 myoblasts. J. Cell Physiol. 2018, 233, 736–747. [Google Scholar] [CrossRef] [PubMed]
- Shamkani, W.A.; Jafar, N.S.; Narayanan, S.R.; Rajappan, A.K. Acute myocardial infarction in a young lady due to vitamin B12 deficiency induced hyperhomocysteinemia. Heart Views 2015, 16, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Boushey, C.J.; Beresford, S.A.; Omenn, G.S.; Motulsky, A.G. A quantitative assessment of plasma homocysteine as a risk factor for vascular disease. probable benefits of increasing folic acid intakes. JAMA 1995, 274, 1049–1057. [Google Scholar] [CrossRef] [PubMed]
- Welch, G.N.; Loscalzo, J. Homocysteine and atherothrombosis. N. Engl. J. Med. 1998, 338, 1042–1050. [Google Scholar] [CrossRef]
- Ciaccio, M.; Bivona, G.; Bellia, C. Therapeutical approach to plasma homocysteine and cardiovascular risk reduction. Ther. Clin. Risk Manag. 2008, 4, 219–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Homocysteine Studies Collaboration. Homocysteine and risk of ischemic heart disease and stroke: A meta-analysis. JAMA 2002, 288, 2015–2022. [Google Scholar] [CrossRef] [PubMed]
- Logan, R.F.A.; Grainge, M.J.; Shepherd, V.C.; Armitage, N.C.; Muir, K.R.; ukCAP Trial Group. Aspirin and folic acid for the prevention of recurrent colorectal adenomas. Gastroenterology 2008, 134, 29–38. [Google Scholar] [CrossRef]
- VITATOPS Trial Study Group; Hankey, G.J.; Algra, A.; Chen, C.; Wong, M.C.; Cheung, R.; Wong, L.; Divjak, I.; Ferro, J.; de Freitas, G.; et al. VITATOPS, the vitamins to prevent stroke trial: Rationale and design of a randomised trial of B-vitamin therapy in patients with recent transient ischaemic attack or stroke (NCT00097669) (ISRCTN74743444). Int. J. Stroke 2007, 2, 144–150. [Google Scholar] [CrossRef]
- Wang, Y.; Jin, Y.; Wang, Y.; Li, L.; Liao, Y.; Zhang, Y.; Yu, D. The effect of folic acid in patients with cardiovascular disease: A systematic review and meta-analysis. Medicine 2019, 98, e17095. [Google Scholar] [CrossRef] [PubMed]
- Scazzone, C.; Bono, A.; Tornese, F.; Arsena, R.; Schillaci, R.; Butera, D.; Cottone, S. Correlation between low folate levels and hyperhomocysteinemia, but not with vitamin B12 in hypertensive patients. Ann. Clin. Lab. Sci. 2014, 44, 286–290. [Google Scholar]
- Yang, X.; Zhang, M.; Song, R.; Liu, C.; Huo, Y.; Qian, G. The modifying effect of the MTHFR genotype on the association between folic acid supplementation and pulse wave velocity: Findings from the CSPPT. Cardiovasc. Ther. 2018, 36, e12473. [Google Scholar] [CrossRef] [Green Version]
- Wollack, J.B.; Makori, B.; Ahlawat, S.; Koneru, R.; Picinich, S.C.; Smith, A.; Goldman, I.D.; Qiu, A.; Cole, P.D.; Glod, J.; et al. Characterization of folate uptake by choroid plexus epithelial cells in a rat primary culture model. J. Neurochem. 2008, 104, 1494–1503. [Google Scholar] [CrossRef]
- Jakaria, M.; Park, S.-Y.; Haque, M.E.; Karthivashan, G.; Kim, I.-S.; Ganesan, P.; Choi, D.-K. Neurotoxic agent-induced injury in neurodegenerative disease model: Focus on involvement of glutamate receptors. Front. Mol. Neurosci. 2018, 11, 307. [Google Scholar] [CrossRef]
- Pezacka, E.H.; Jacobsen, D.W.; Luce, K.; Green, R. Glial cells as a model for the role of cobalamin in the nervous system: Impaired synthesis of cobalamin coenzymes in cultured human astrocytes following short-term cobalamin-deprivation. Biochem. Biophys. Res. Commun. 1992, 184, 832–839. [Google Scholar] [CrossRef]
- Pramod, A.B.; Foster, J.; Carvelli, L.; Henry, L.K. SLC6 transporters: Structure, function, regulation, disease association and therapeutics. Mol. Asp. Med. 2013, 34, 197–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charlton, C.G. Methylation reactions at dopaminergic nerve endings, serving as biological off-switches in managing dopaminergic functions. Neural. Regen. Res. 2014, 9, 1110–1111. [Google Scholar] [CrossRef]
- Pavel, K.; Damulin, I. Cognitive impairments in vitamin b12 and folic acid deficiencies and hyperhomocysteinemia. Clinician 2015, 9, 18. [Google Scholar] [CrossRef] [Green Version]
- Ducker, G.S.; Rabinowitz, J.D. One-carbon metabolism in health and disease. Cell Metab. 2017, 25, 27–42. [Google Scholar] [CrossRef] [Green Version]
- Miranpuri, G.S.; Nguyen, J.; Moreno, N.; Yutuc, N.A.; Kim, J.; Buttar, S.; Brown, G.R.; Sauer, S.E.; Singh, C.K.; Kumar, S.; et al. Folic acid modulates matrix metalloproteinase-9 expression following spinal cord injury. Ann. Neurosci. 2019, 26, 60–65. [Google Scholar] [CrossRef]
- MacFarlane, A.J.; Behan, N.A.; Field, M.S.; Williams, A.; Stover, P.J.; Yauk, C.L. Dietary folic acid protects against genotoxicity in the red blood cells of mice. Mutat. Res. 2015, 779, 105–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mesolella, M.; Iengo, M.; Testa, D.; Ricciardiello, F.; Iorio, B. Chemoprevention using folic acid for dysplastic lesions of the larynx. Mol. Clin. Oncol. 2017, 7, 843–846. [Google Scholar] [CrossRef]
- Koul, A.; Kaur, N.; Chugh, N.A. Folic acid modulates DMBA/TPA-induced changes in skin of mice: A study relevant to carcinogenesis. J. Diet. Suppl. 2018, 15, 72–87. [Google Scholar] [CrossRef]
- Fan, C.; Yu, S.; Zhang, S.; Ding, X.; Su, J.; Cheng, Z. Association between folate intake and risk of head and neck squamous cell carcinoma: An overall and dose-response PRISMA meta-analysis. Medicine 2017, 96, e8182. [Google Scholar] [CrossRef]
- Guo, X.; Dai, X.; Ni, J.; Cao, N.; Yang, G.; Xue, J.; Wang, X. High concentration of sugars is genotoxic to folate-deficient cells. Mutat. Res. 2019, 814, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Scocchera, E.; Reeve, S.M.; Keshipeddy, S.; Lombardo, M.N.; Hajian, B.; Sochia, A.E.; Alverson, J.B.; Priestley, N.D.; Anderson, A.C.; Wright, D.L. Charged nonclassical antifolates with activity against gram-positive and gram-negative pathogens. ACS Med. Chem. Lett. 2016, 7, 692–696. [Google Scholar] [CrossRef] [Green Version]
- Hoody, J.; Alverson, J.B.; Keshipeddy, S.; Barney, P.A.; Walker, L.; Anderson, A.C.; Wright, D.L.; Priestley, N.D. Pharmaceutical analysis of a novel propargyl-linked antifolate antibiotic in the mouse. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2017, 1051, 54–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Graaf, D.; Sharma, R.C.; Mechetner, E.B.; Schimke, R.T.; Roninson, I.B. P-glycoprotein confers methotrexate resistance in 3T6 cells with deficient carrier-mediated methotrexate uptake. Proc. Natl. Acad. Sci. USA 1996, 93, 1238–1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, Y.C.; Bernal, S.D. Reversal of methotrexate resistance in human squamous carcinoma cells by SQM1-liposome. Anticancer. Res. 1999, 19, 251–254. [Google Scholar] [PubMed]
- Zhang, X.; Zhang, D.; Huang, L.; Li, G.; Chen, L.; Ma, J.; Li, M.; Wei, M.; Zhou, W.; Zhou, C.; et al. Discovery of novel biomarkers of therapeutic responses in han chinese pemetrexed-based treated advanced NSCLC patients. Front. Pharmacol. 2019, 10, 944. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Food | Daily Value (%) |
|---|---|
| Beef liver, braised, 3 ounces | 54 |
| Spinach, boiled, ½ cup | 33 |
| Black-eyed peas, boiled, ½ cup | 26 |
| Breakfast cereals, fortified with 25% of the DV | 25 |
| Rice, white, medium-grain, cooked, ½ cup | 22 |
| Asparagus, boiled, 4 spears | 22 |
| Brussels sprouts, frozen, boiled, ½ cup | 20 |
| Spaghetti, cooked, enriched, ½ cup | 19 |
| Lettuce, romaine, shredded, 1 cup | 16 |
| Avocado, raw, sliced, ½ cup | 15 |
| Spinach, raw, 1 cup | 15 |
| Broccoli, chopped, frozen, cooked, ½ cup | 13 |
| Mustard greens, chopped, frozen, boiled, ½ cup | 13 |
| Bread, white, 1 slice * | 13 |
| Green peas, frozen, boiled, ½ cup | 12 |
| Kidney beans, canned, ½ cup | 12 |
| Wheat germ, 2 tablespoons | 10 |
| Tomato juice, canned, ¾ cup | 9 |
| Crab, Dungeness, 3 ounces | 9 |
| Orange juice, ¾ cup | 9 |
| Age, Years | Adequate Intake, µg/Day | Tolerable Upper Intake Level, µg/Day |
|---|---|---|
| 1–3 | 150 | 300 |
| 4–8 | 200 | 400 |
| 9–13 | 300 | 600 |
| 14–18 | 400 | 800 |
| 19+ * | 400–600 | 1000 |
| Serum/Plasma Folate ng/mL (nmol/L) | Ber Blood Cells Folate ng/mL (nmol/L) | Interpretation |
|---|---|---|
| >20 (>45.3) | Increased | |
| 6−20 (13.5−45.3) | Normal | |
| 3−5.9 (6.8−13.4) | Probable deficiency | |
| <3 (<6.8) | <100 (<226.5) | Deficiency |
| Parameter | Homocysteine Level ng/mL (nmol/L) |
|---|---|
| Serum/plasma folate | <4 (<10) |
| Red blood cell folate | <151 (<340) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shulpekova, Y.; Nechaev, V.; Kardasheva, S.; Sedova, A.; Kurbatova, A.; Bueverova, E.; Kopylov, A.; Malsagova, K.; Dlamini, J.C.; Ivashkin, V. The Concept of Folic Acid in Health and Disease. Molecules 2021, 26, 3731. https://doi.org/10.3390/molecules26123731
Shulpekova Y, Nechaev V, Kardasheva S, Sedova A, Kurbatova A, Bueverova E, Kopylov A, Malsagova K, Dlamini JC, Ivashkin V. The Concept of Folic Acid in Health and Disease. Molecules. 2021; 26(12):3731. https://doi.org/10.3390/molecules26123731
Chicago/Turabian StyleShulpekova, Yulia, Vladimir Nechaev, Svetlana Kardasheva, Alla Sedova, Anastasia Kurbatova, Elena Bueverova, Arthur Kopylov, Kristina Malsagova, Jabulani Clement Dlamini, and Vladimir Ivashkin. 2021. "The Concept of Folic Acid in Health and Disease" Molecules 26, no. 12: 3731. https://doi.org/10.3390/molecules26123731
APA StyleShulpekova, Y., Nechaev, V., Kardasheva, S., Sedova, A., Kurbatova, A., Bueverova, E., Kopylov, A., Malsagova, K., Dlamini, J. C., & Ivashkin, V. (2021). The Concept of Folic Acid in Health and Disease. Molecules, 26(12), 3731. https://doi.org/10.3390/molecules26123731