
Gundelia tournefortii: Fractionation, Chemical Composition and GLUT4 Translocation Enhancement in Muscle Cell Line

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Extract Preparation
2.2. Flash Chromatography Extract Fractionation
2.3. Silylation Derivatization
2.4. Gas Chromatography–Mass Spectrometry Analysis
2.5. Identification of Components
2.6. MTT Assay
2.7. Determination of Surface GLUT4myc
Statistical Analysis
3. Results
3.1. GT Methanol Extract Fractionation and Chemical Detection
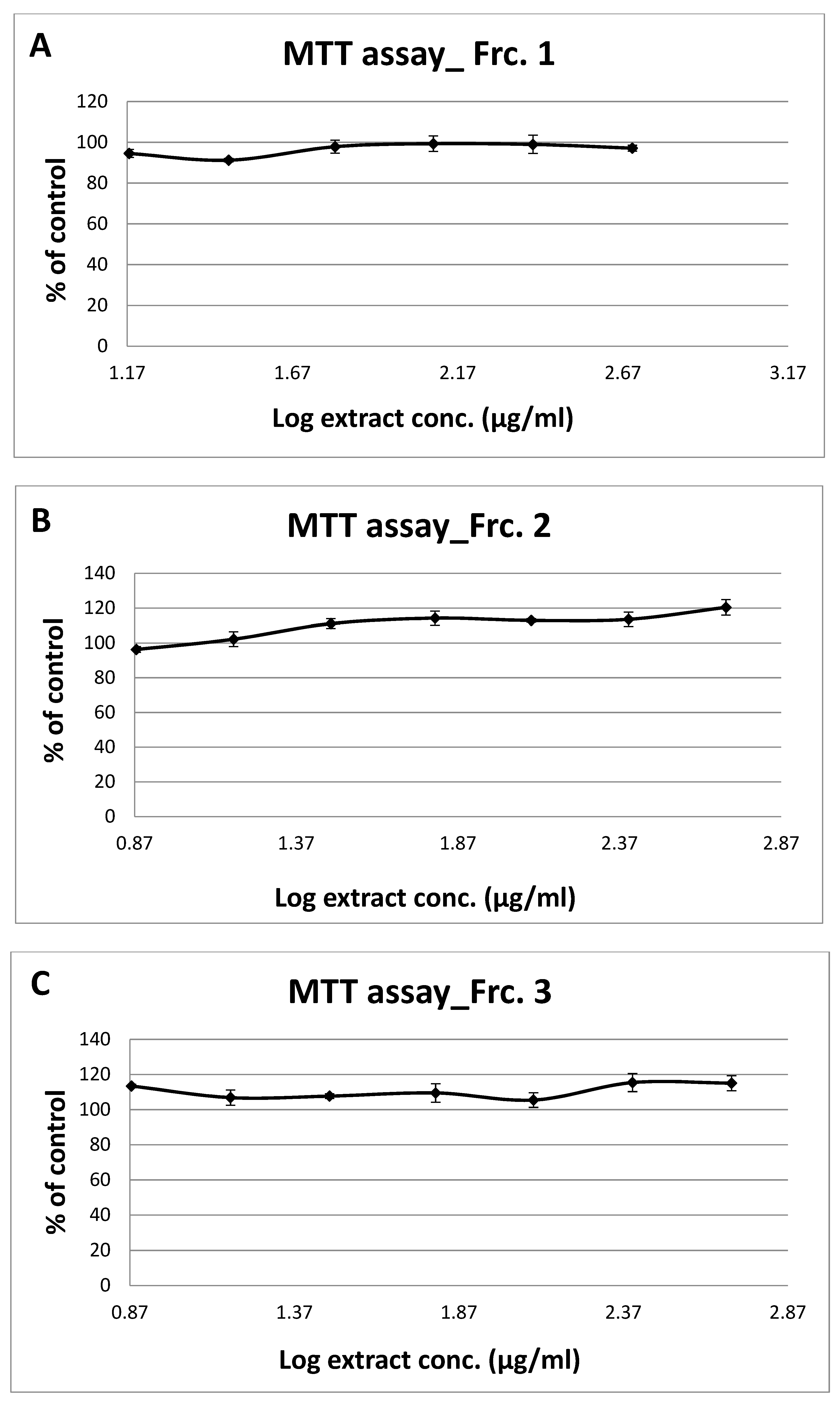
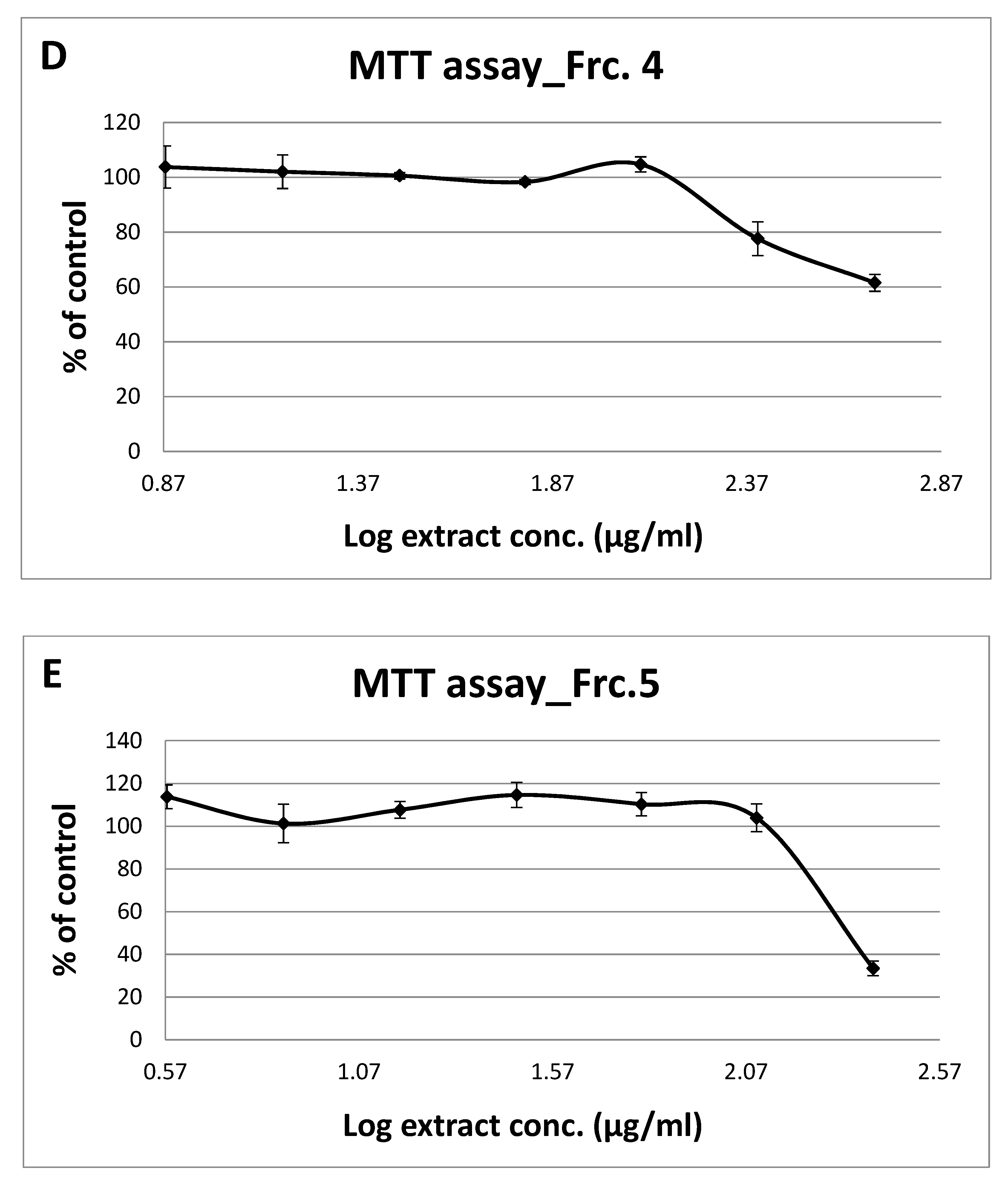
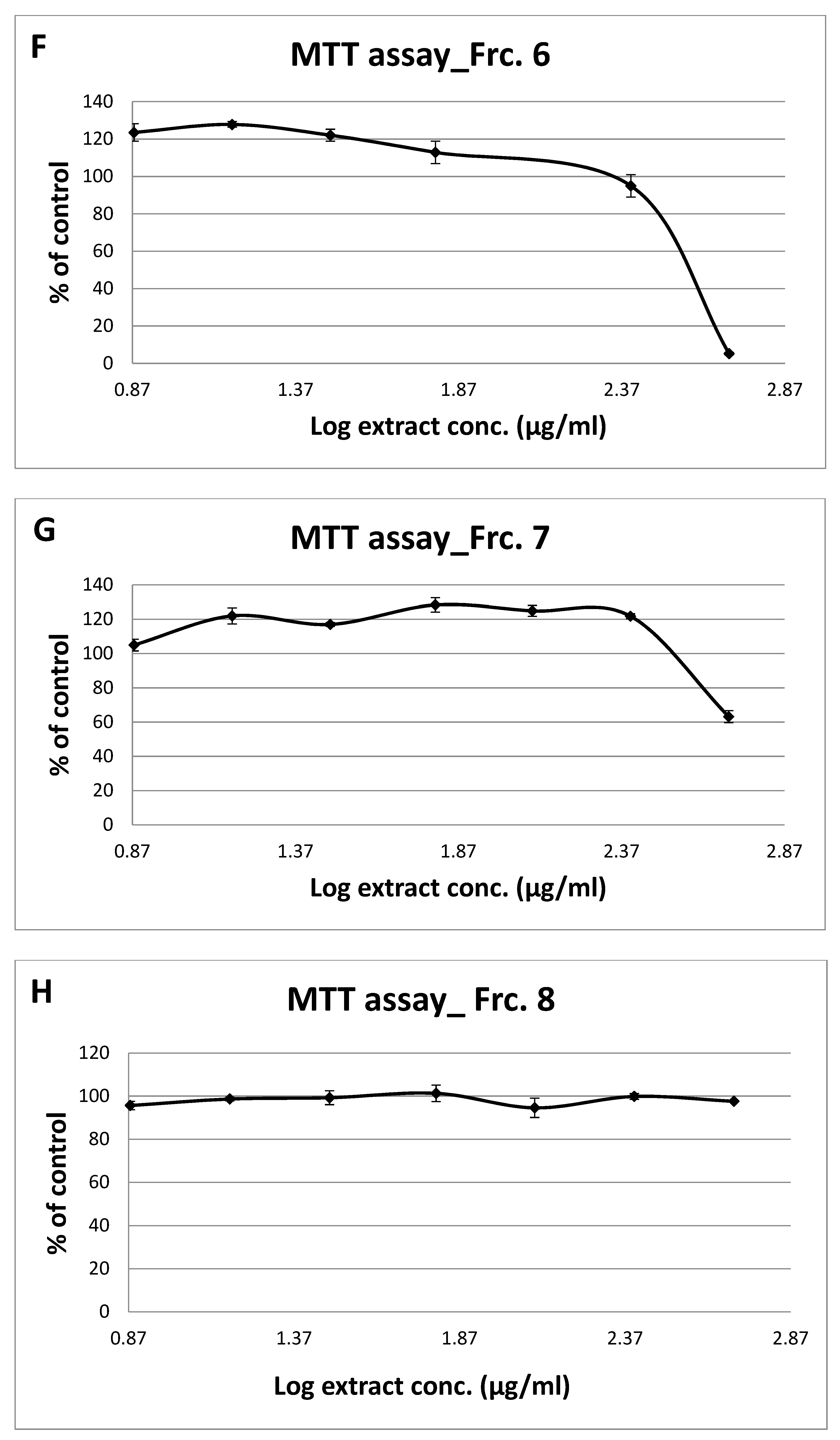
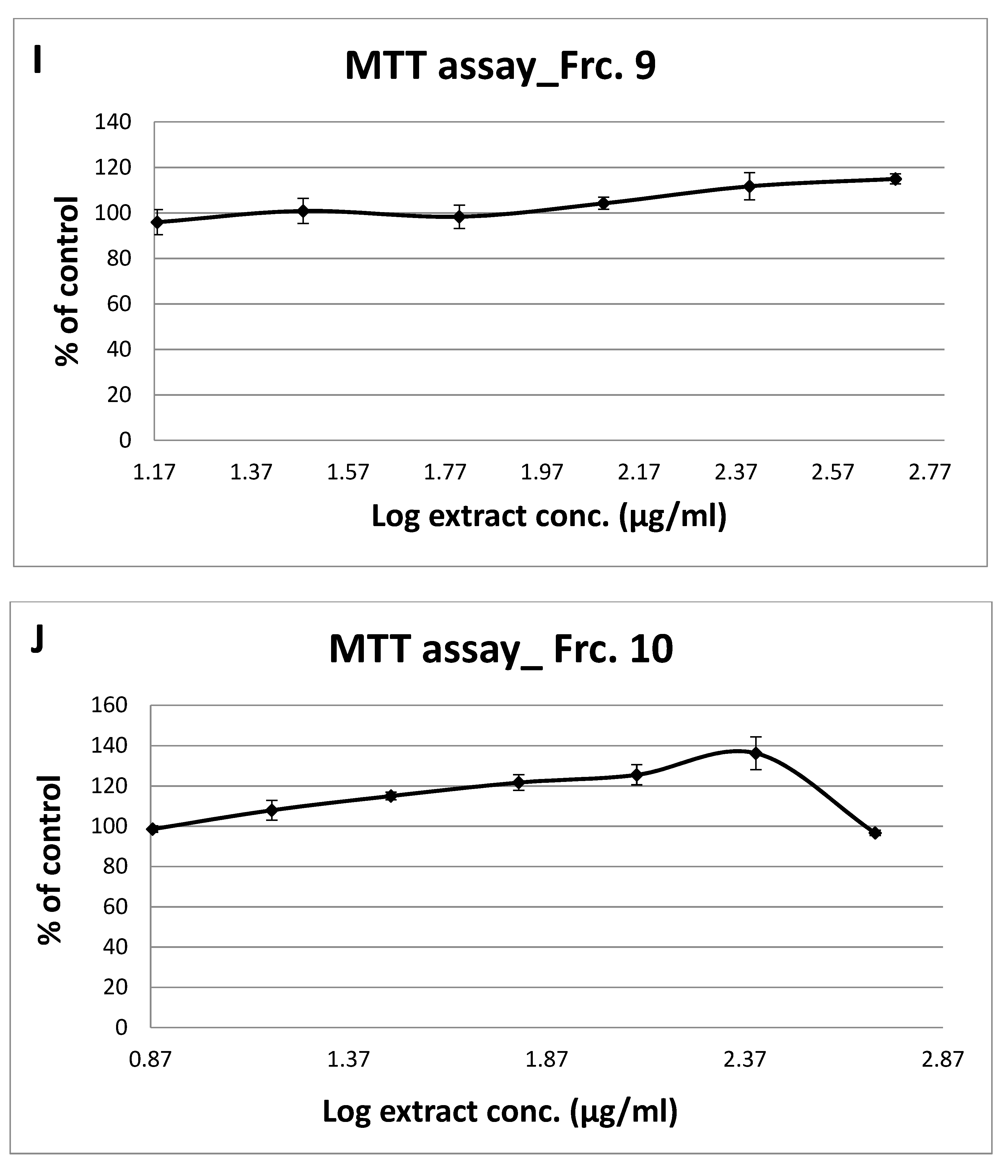
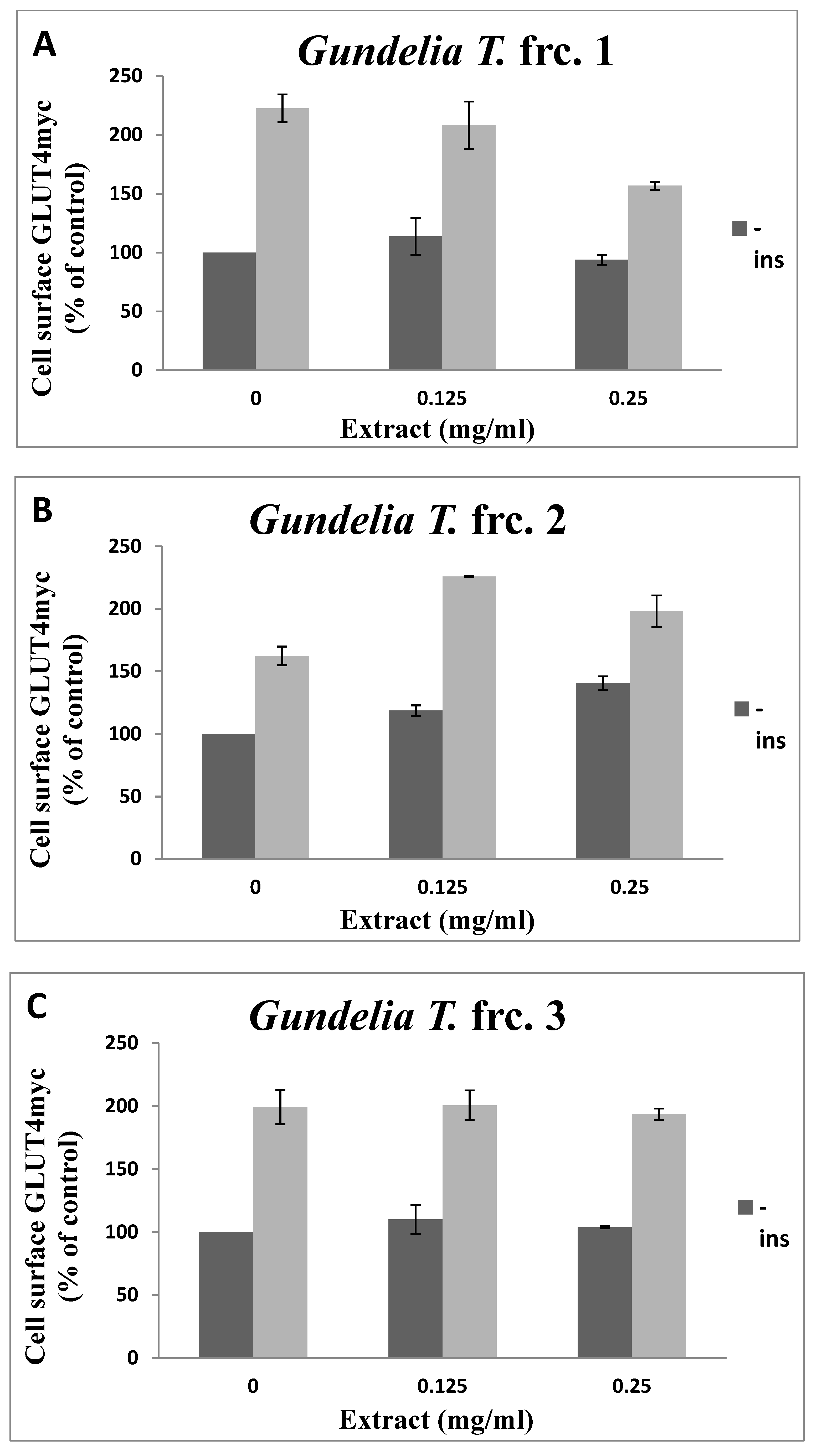
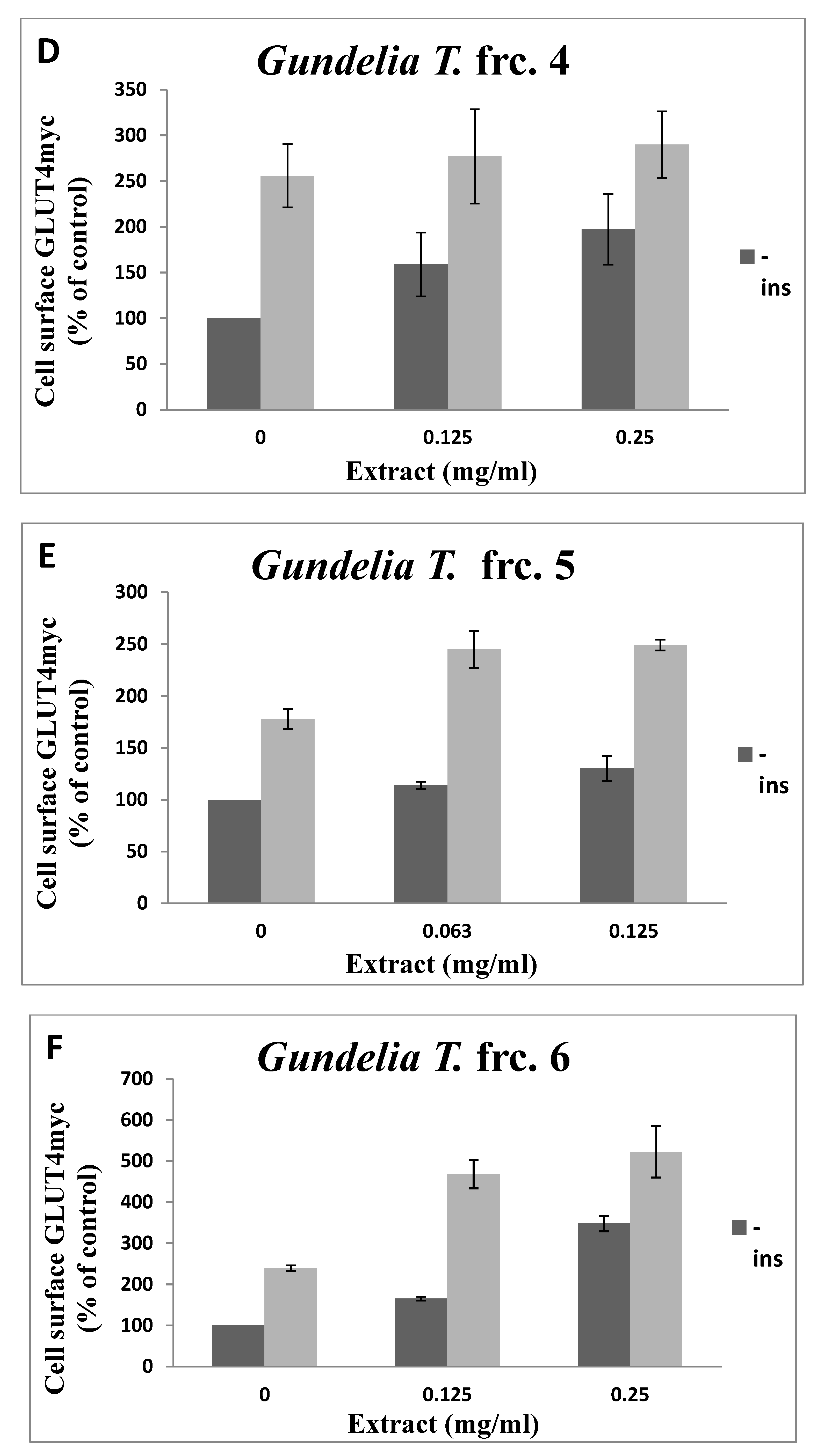
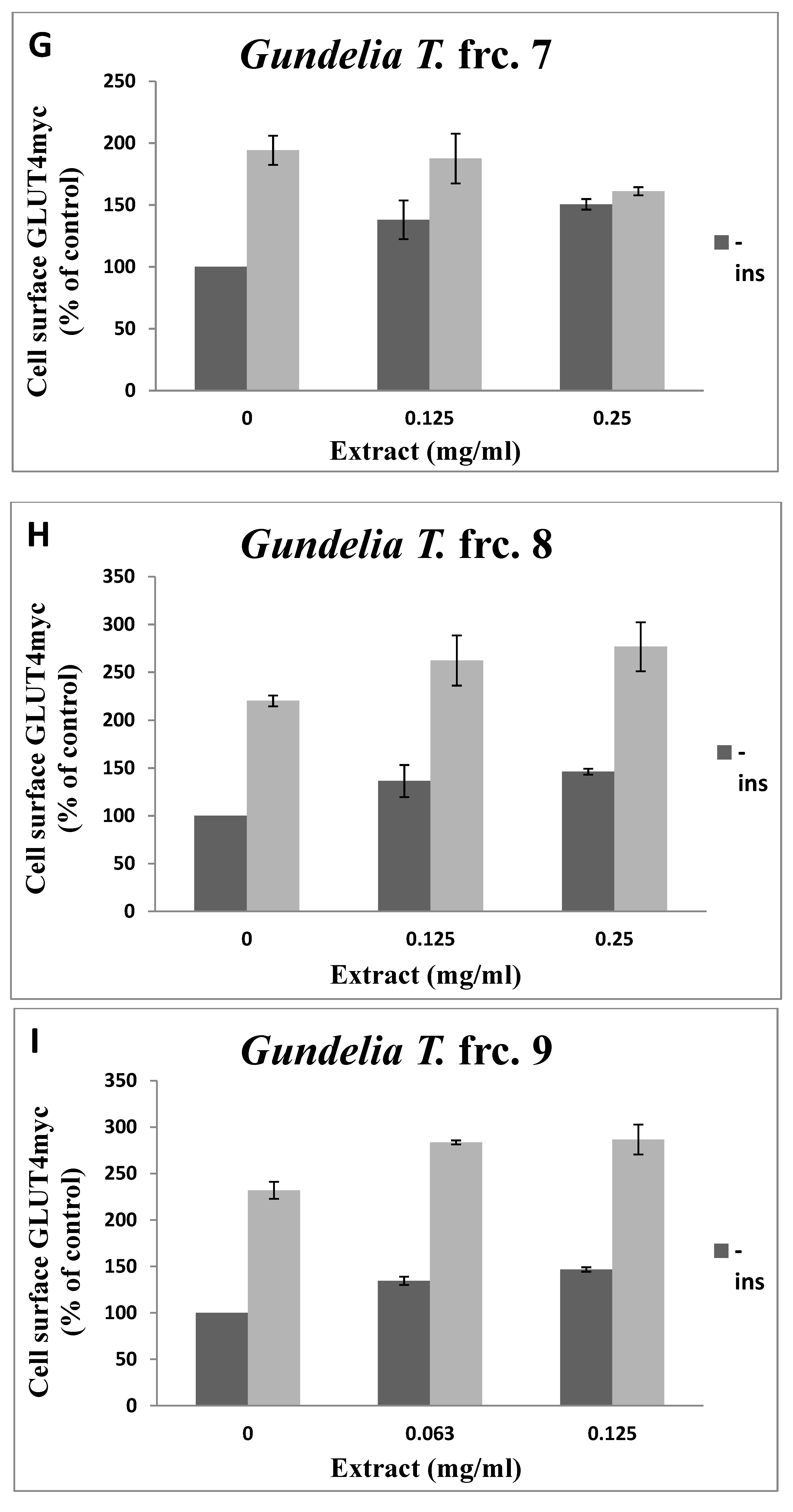
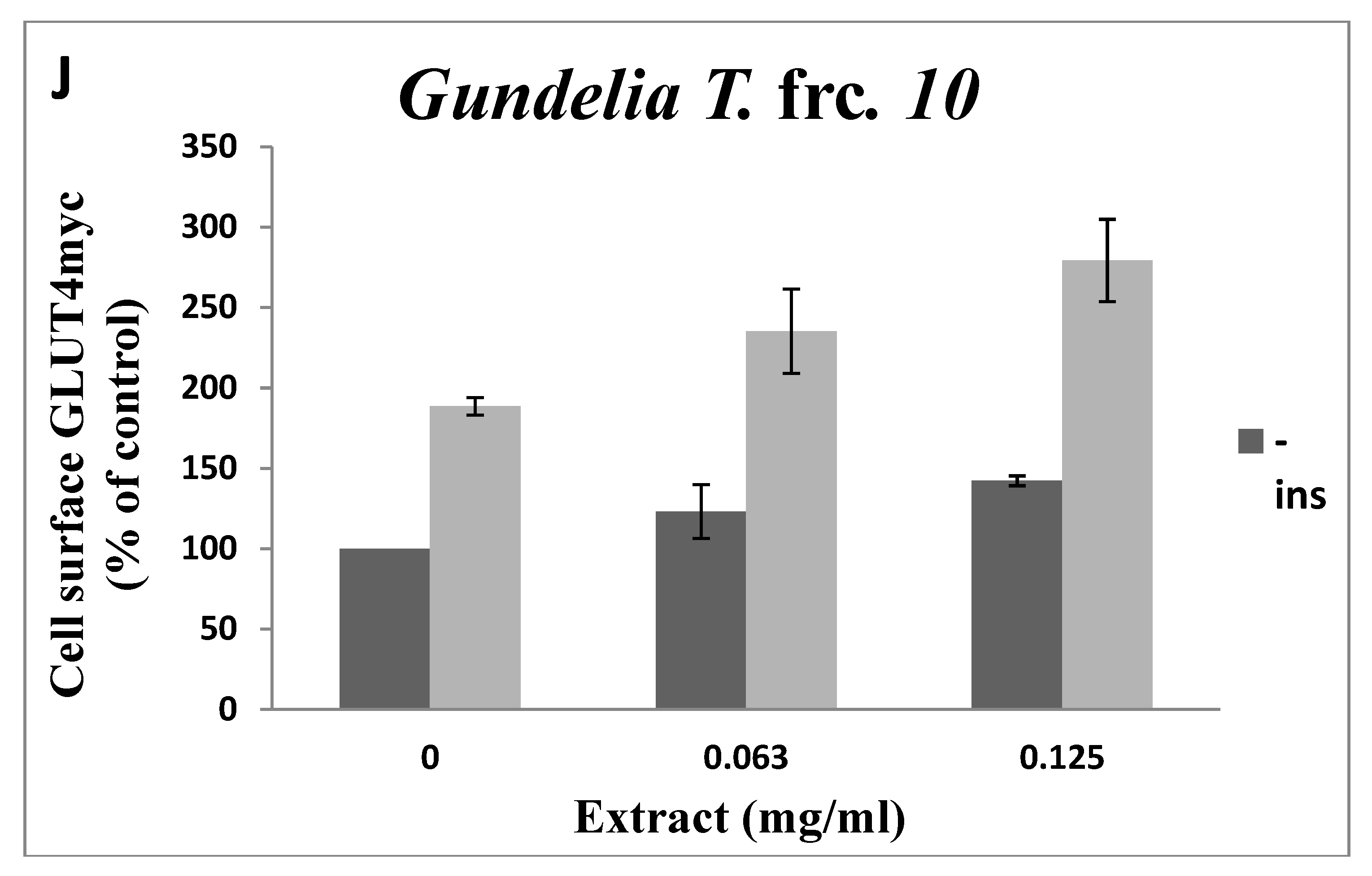
3.2. Toxicity of GT Fractions and Effects on GLUT4 Translocation
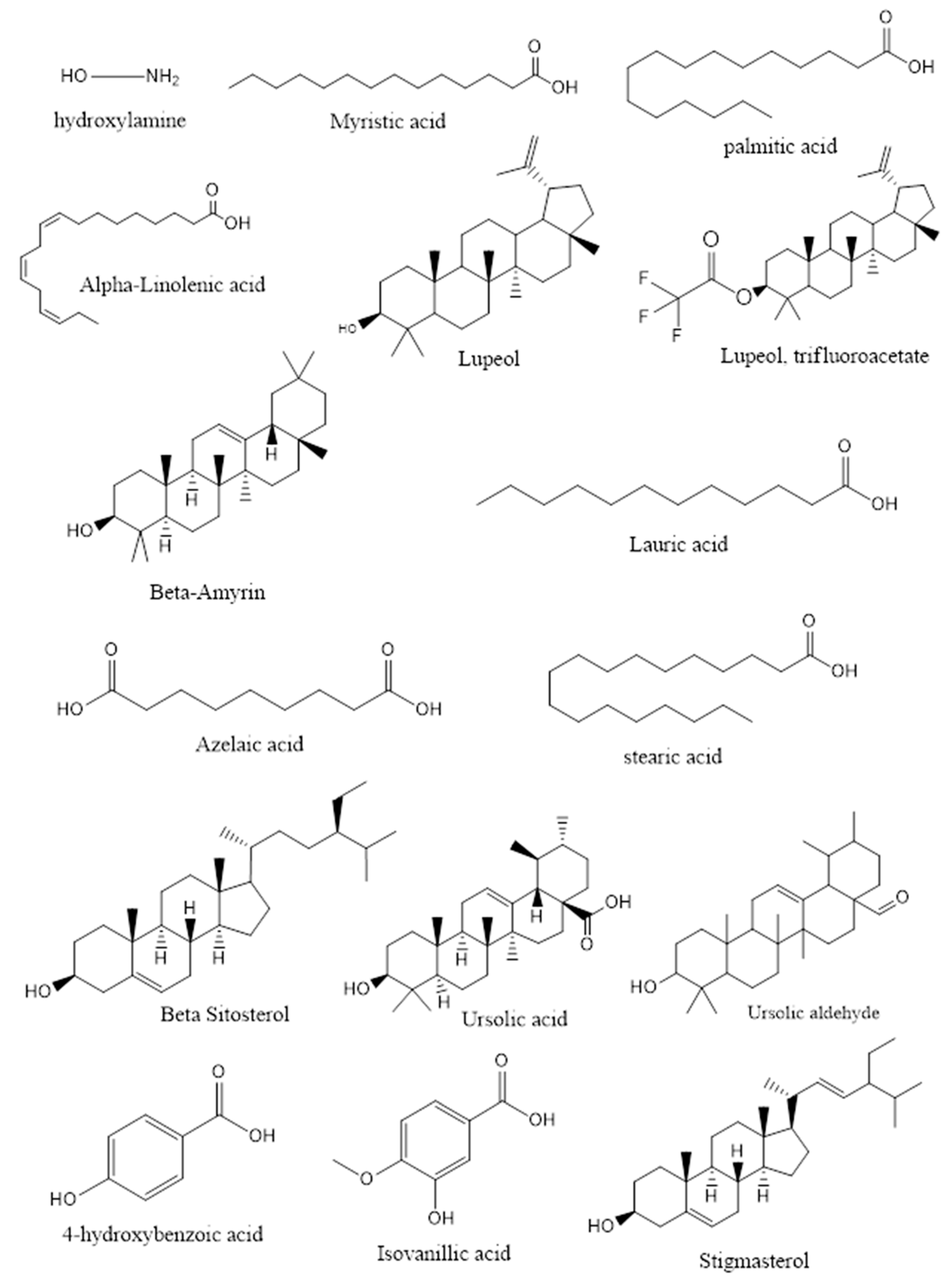
3.3. GT Fraction Components Attributed to GLUT4 Translocation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraction 1 | ||||||
| Peak | Name | Rt | % Area | Match Factor | Association with Diabetes | References |
| 1 | Hydroxylamine | 15.28 | 0.60 | 93.2 | Enhances glucose uptake in C2C12 skeletal muscle cells | [19] |
| 2 | Glycerol | 20.34 | 0.10 | 86.6 | ||
| 3 | Neophytadiene | 33.02 | 0.46 | 93.3 | ||
| 4 | Myristic acid | 33.22 | 0.08 | 81.2 | Enhances basal glucose uptake in myotubes | [20] |
| 5 | 3,7,11,15-Tetramethyl-2-hexadecen-1-ol | 33.67 | 0.20 | 90.8 | ||
| 6 | Methyl palmitate | 34.28 | 1.80 | 94.6 | ||
| 7 | Palmitic acid | 35.81 | 0.70 | 95.7 | Enhances basal glucose uptake in myotubes | [21,22] |
| 8 | 9,12-Octadecadienoic acid, methyl ester, (E,E)- | 36.34 | 0.67 | 93.1 | ||
| 9 | 9-Octadecenoic acid (Z)-methyl ester | 36.41 | 0.87 | 89.4 | ||
| 10 | Methyl stearate | 36.68 | 0.15 | 88.9 | ||
| 11 | α-Linolenic acid | 37.04 | 0.30 | 86.3 | Enhances insulin secretion from pancreatic beta cells | [23] |
| 12 | Stearic acid | 37.86 | 0.43 | 93.7 | Enhances basal glucose uptake in myotubes | [23] |
| 13 | Dinonyl phthalate | 45.19 | 0.11 | 82.2 | ||
| 14 | 24-Noroleana-3,12-diene | 47.32 | 0.48 | 83.7 | ||
| 15 | β-amyrin acetate | 54.89 | 18.74 | 93.3 | ||
| 16 | Lupeol | 55.40 | 49.25 | 82.4 | Stimulates glucose utilization by skeletal muscles | [24] |
| 17 | Cycloartenyl acetate | 56.37 | 8.19 | 83.3 | ||
| 18 | Lupeol-trifluoroacetate | 56.53 | 16.88 | 82.8 | Stimulates glucose utilization by skeletal muscles | [24] |
| Fraction 2 | ||||||
| Peak | Name | Rt | % Area | Match Factor | Association with Diabetes | References |
| 1 | Palmitic acid | 35.81 | 11.2 | 91.6 | Enhances basal glucose uptake in myotubes | [21,22] |
| 2 | β-Amyrin | 53.60 | 35.00 | 90.3 | Reduces elevated plasma glucose levels during the oral glucose tolerance test in mice and α-glucosidase inhibitor | [25,26] |
| 3 | Lupeol | 55.31 | 53.79 | 86.1 | Stimulates glucose utilization by skeletal muscles | [24] |
| Fraction 3 | ||||||
| Peak | Name | Rt | % Area | Match Factor | Association with Diabetes | References |
| 1 | Benzoic acid | 19.13 | 0.19 | 91.1 | ||
| 2 | Glycerol | 20.34 | 0.11 | 88.9 | ||
| 3 | 4-(Methoxycarbonyl)phenol | 25.67 | 1.88 | 93.4 | ||
| 4 | Lauric acid | 29.39 | 0.39 | 82.3 | Enhances glucose-stimulated insulin secretion | [27] |
| 5 | Azelaic acid | 32.45 | 1.13 | 88.1 | Restores normal levels of plasma glucose, insulin, HbA1c, Hb, liver glycogen, and carbohydrate in diabetic mice | [28] |
| 6 | Myristic acid | 33.21 | 0.53 | 94.6 | Enhances basal glucose uptake in myotubes | [20] |
| 7 | Pentadecanoic acid | 34.61 | 0.07 | 86.2 | ||
| 8 | Palmitic acid | 35.82 | 8.22 | 98.7 | Enhances basal glucose uptake in myotubes | [21,22] |
| 9 | Heptadecanoic acid | 36.88 | 0.22 | 86.8 | ||
| 10 | Linoelaidic acid | 37.57 | 1.41 | 92.8 | ||
| 11 | 9-Octadecenoic acid, (E)- | 37.62 | 2.88 | 96 | ||
| 12 | Stearic acid | 37.86 | 2.56 | 97.1 | Enhances basal glucose uptake in myotubes | [23] |
| 13 | Eicosanoic acid | 39.61 | 0.48 | 88.6 | ||
| 14 | Glyceryl palmitate | 40.96 | 3.54 | 86 | ||
| 15 | Glycerol monostearate | 43.08 | 1.28 | 93.5 | ||
| 16 | Lignoceric acid | 43.68 | 0.50 | 87.1 | ||
| 17 | Stigmasterol | 52.84 | 2.62 | 94.5 | Increases GLUT4 translocation and expression | [29] |
| 18 | β-Sitosterol | 53.65 | 2.54 | 90.9 | Improves glycemic control through activation of insulin receptors and GLUT4 in adipose tissue | [30] |
| 19 | Ursolic acid | 56.32 | 38.46 | 91.3 | Stimulates glucose uptake in 3T3-L1 adipocytes and α-glucosidase inhibitor | [31] |
| 20 | Ursolic aldehyde | 56.86 | 19.33 | 59.4 | Ursolic acid analogs are α-glucosidase inhibitors | [25] |
| 21 | Brucine | 57.35 | 11.65 | 91.7 | ||
| Fraction 4 | ||||||
| Peak | Name | Rt | % Area | Match Factor | Association with Diabetes | References |
| 1 | Hydroxylamine | 15.28 | 0.22 | 83.9 | Enhances glucose uptake in C2C12 skeletal muscle cells | [19] |
| 2 | Glycerol | 20.34 | 0.61 | 91 | ||
| 3 | Benzeneacetic acid | 20.60 | 2.83 | 94.7 | ||
| 4 | Suberic acid | 30.51 | 0.46 | 83.8 | ||
| 5 | Azelaic acid | 32.45 | 5.87 | 92.1 | Restores normal levels of plasma glucose, insulin, HbA1c, Hb, liver glycogen, and carbohydrate in diabetic mice | [28] |
| 6 | Myristic acid | 33.21 | 0.64 | 88.5 | Enhances basal glucose uptake in myotubes | [20] |
| 7 | Gallic acid | 34.99 | 0.74 | 81.9 | ||
| 8 | Palmitic acid | 35.82 | 11.79 | 98.6 | Enhances basal glucose uptake in myotubes | [21,22] |
| 9 | Linoelaidic acid | 37.57 | 3.04 | 83.9 | ||
| 10 | Stearic acid | 37.86 | 5.64 | 95 | Enhances basal glucose uptake in myotubes | [23] |
| 11 | Glyceryl palmitate | 40.96 | 7.82 | 90 | ||
| 12 | Glycerol monostearate | 43.08 | 4.77 | 91.2 | ||
| 13 | Stigmasterol | 52.84 | 2.14 | 85.1 | Increases GLUT4 translocation and expression | [29] |
| 14 | Brucine | 57.36 | 53.42 | 91.7 | ||
| Fraction 5 | ||||||
| Peak | Name | Rt | % Area | Match Factor | Association with Diabetes | References |
| 1 | Lactic Acid | 13.71 | 0.28 | 86.2 | ||
| 2 | Glycerol | 20.34 | 0.26 | 92.6 | ||
| 3 | Benzeneacetic acid | 20.59 | 1.10 | 95.6 | ||
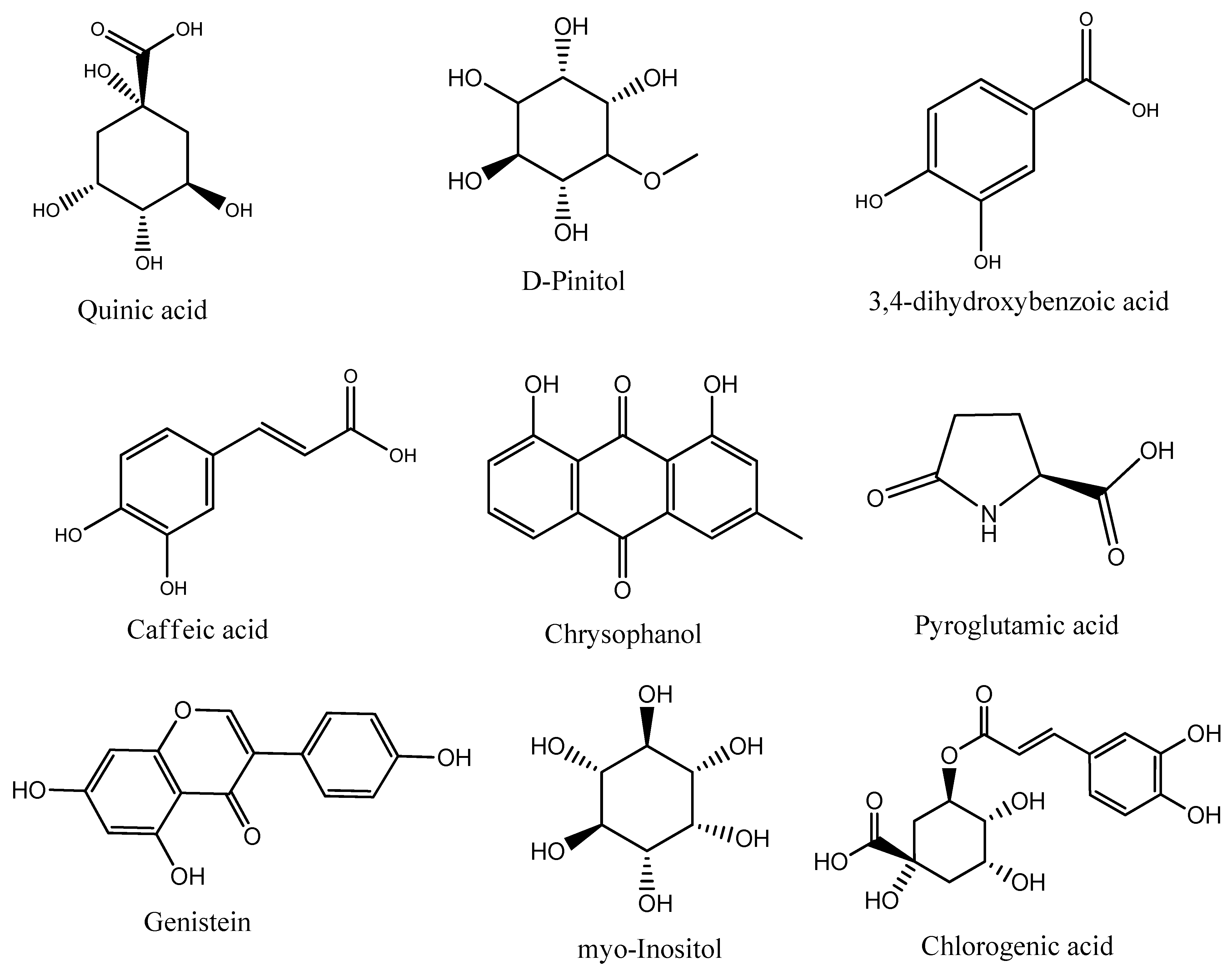
| 4 | 4-Hydroxybenzoic acid | 28.93 | 76.20 | 98.9 | Increases glucose consumption in normal and diabetic rats | [16,17] |
| 5 | Isovanillic acid | 31.89 | 0.29 | 83 | Stimulates a dose-dependent increase in glucose transport through GLUT4 | [32] |
| 6 | Azelaic acid | 32.45 | 0.55 | 89.9 | Restores normal levels of plasma glucose, insulin, HbA1c, Hb, liver glycogen, and carbohydrate in diabetic mice | [28] |
| 7 | D-Pinitol | 33.93 | 0.39 | 86.4 | Stimulates translocation of GLUT4 in skeletal muscle of C57BL/6 mice and induces translocation of GLUT4 to the plasma membrane | [18,33] |
| 8 | Palmitic acid | 35.81 | 3.18 | 98.6 | Enhances basal glucose uptake in myotubes | [21,22] |
| 9 | Stearic acid | 37.86 | 1.99 | 94.7 | Enhances basal glucose uptake in myotubes | [23] |
| 10 | Glyceryl palmitate | 40.96 | 4.56 | 96 | ||
| 11 | Glycerol monostearate | 43.08 | 3.45 | 94.1 | ||
| 12 | Brucine | 57.30 | 7.75 | 90.2 | ||
| Fraction 6 | ||||||
| Peak | Name | Rt | % Area | Match Factor | Association with Diabetes | References |
| 1 | Lactic Acid | 13.69 | 0.66 | 92.7 | ||
| 2 | Glycerol | 20.34 | 0.62 | 91.8 | ||
| 3 | 4-Hydroxybenzoic acid | 28.89 | 7.67 | 98.2 | Increases glucose consumption in normal and diabetic rats | [16,17] |
| 4 | Isovanillic acid | 31.89 | 8.35 | 96.8 | Stimulates a dose-dependent increase in glucose transport through GLUT4 | [32] |
| 5 | Azelaic acid | 32.45 | 1.14 | 92.9 | Restores normal levels of plasma glucose, insulin, HbA1c, Hb, liver glycogen, and carbohydrate in diabetic mice | [28] |
| 6 | Quinic acid | 33.88 | 4.21 | 87.7 | Enhances glucose-stimulated insulin secretion in both INS-1E cells and mouse islets | [34] |
| 7 | Dihydroferulic acid | 34.01 | 2.61 | 94.2 | ||
| 8 | 4-Coumaric acid | 34.57 | 7.73 | 96.5 | ||
| 9 | Indole-5-carboxylic acid | 35.67 | 16.26 | 84.9 | ||
| 10 | Palmitic acid | 35.81 | 6.18 | 98 | Enhances basal glucose uptake in myotubes | [21,22] |
| 11 | Isoferulic acid | 36.42 | 9.52 | 94.6 | ||
| 12 | Stearic acid | 37.86 | 5.18 | 96.3 | Enhances basal glucose uptake in myotubes | [23] |
| 13 | Glyceryl palmitate | 40.95 | 11.71 | 96.3 | ||
| 14 | Glycerol monostearate | 43.08 | 9.65 | 97.1 | ||
| 15 | Questinol | 52.85 | 8.50 | 82.1 | ||
| Fraction 7 | ||||||
| Peak | Name | Rt | % Area | Match Factor | Association with Diabetes | References |
| 1 | Propanoic acid | 16.86 | 2.74 | 91.1 | ||
| 2 | Glycerol | 20.34 | 1.40 | 93.8 | ||
| 3 | Succinic acid | 21.20 | 47.86 | 98.1 | ||
| 4 | 4-Hydroxybenzoic acid | 28.89 | 0.36 | 87.8 | Increases glucose consumption in normal and diabetic rats | [16,17] |
| 5 | Azelaic acid | 32.45 | 7.81 | 94.6 | Restores normal levels of plasma glucose, insulin, HbA1c, Hb, liver glycogen, and carbohydrate in diabetic mice | [28] |
| 6 | D-Ribonic acid | 32.66 | 1.45 | 90 | ||
| 7 | Protocatechuic acid | 32.94 | 12.24 | 95.9 | Protects mesangial cells against high glucose damage via inhibition of the p38 MAPK signaling pathway | [35,36,37] |
| 8 | Quinic acid | 33.88 | 1.01 | 83.8 | Enhances glucose-stimulated insulin secretion in both INS-1E cells and mouse islets | [34] |
| 9 | Syringic acid | 34.09 | 1.49 | 94.3 | ||
| 10 | Caffeic acid | 36.93 | 12.81 | 95.8 | Reduces insulin resistance and modulates glucose uptake in HepG2 cells | [38] |
| 11 | 1,2-Hexadecanediol | 37.56 | 5.02 | 82.7 | ||
| 12 | Stearic acid | 37.86 | 2.03 | 95.2 | Enhances basal glucose uptake in myotubes | [23] |
| 13 | Glyceryl palmitate | 40.95 | 1.37 | 90.3 | ||
| 14 | Chrysophanol | 43.05 | 1.60 | 85.2 | Increases GLUT4 expression in myotubes | [39] |
| 15 | Decanedioic acid, bis(2-ethylhexyl) ester | 43.27 | 0.82 | 81.9 | ||
| Fraction 8 | ||||||
| Peak | Name | Rt | % Area | Match Factor | Association with Diabetes | References |
| 1 | Hydroxylamine | 15.28 | 0.28 | 90.8 | Enhances glucose uptake in C2C12 skeletal muscle cells | [19] |
| 2 | Hydracrylic acid | 16.33 | 1.65 | 94.7 | ||
| 3 | Glycerol | 20.34 | 0.46 | 93.7 | ||
| 4 | Succinic acid | 21.22 | 42.22 | 98.3 | ||
| 5 | Uracil | 21.92 | 14.05 | 97.1 | ||
| 6 | 5-Methylpyrimidine-2,4-diol | 23.59 | 5.37 | 91.8 | ||
| 7 | 4,5-Dihydro-4-hydroxy-5-(hydroxymethyl)-2(3H)-furanone | 26.22 | 7.73 | 86.3 | ||
| 8 | Pyroglutamic acid | 26.52 | 2.87 | 83.2 | Reduces oral glucose tolerance and serum insulin levels in rats | [40] |
| 9 | 3,4,5-Trihydroxytetrahydro-2H-pyran-2-one | 29.22 | 4.04 | 87.1 | ||
| 10 | 3,4-Dihydroxy-5-(hydroxymethyl)dihydrofuran-2(3H)-one | 29.31 | 1.34 | 93.6 | ||
| 11 | D-(+)-Ribono-1,4-lactone | 30.39 | 0.77 | 89 | ||
| 12 | Xylonic acid | 32.65 | 0.70 | 90.9 | ||
| 13 | Protocatechuic acid | 32.94 | 0.76 | 91.7 | Protects mesangial cells against high glucose damage via inhibition of the p38 MAPK signaling pathway | [35,36,37] |
| 14 | Quinic acid | 33.89 | 1.70 | 84.9 | Enhances glucose-stimulated insulin secretion in both INS-1E cells and mouse islets | [34] |
| 15 | Gulonic acid gamma-lactone | 34.33 | 1.34 | 83.5 | ||
| 16 | D-Gluconic acid | 35.76 | 1.84 | 92.9 | ||
| 17 | Caffeic acid | 36.93 | 0.66 | 92.9 | Reduces insulin resistance and modulates glucose uptake in HepG2 cells | [38] |
| 18 | Stearic acid | 37.86 | 1.67 | 96.7 | Enhances basal glucose uptake in myotubes | [23] |
| 19 | Glyceryl palmitate | 40.95 | 0.83 | 86.4 | ||
| 20 | Glycerol monostearate | 43.08 | 0.64 | 84.1 | ||
| 21 | Decanedioic acid, bis(2-ethylhexyl) ester | 43.28 | 0.97 | 89.9 | ||
| 22 | Genistein | 50.21 | 0.26 | 83.4 | Improves insulin secretion from pancreatic beta cells | [41] |
| 23 | Brucine | 57.30 | 7.84 | 91.6 | ||
| Fraction 9 | ||||||
| Peak | Name | Rt | % Area | Match Factor | Association with Diabetes | References |
| 1 | Hydracrylic acid | 16.34 | 0.44 | 93 | ||
| 2 | Urea | 19.16 | 2.42 | 95.1 | ||
| 3 | Glycerol | 20.37 | 35.50 | 97.8 | ||
| 4 | Butanedioic acid | 21.20 | 1.98 | 97.7 | ||
| 5 | Meso-erythritol | 26.60 | 1.97 | 97.8 | ||
| 6 | 2-Isopropylmalic acid | 27.99 | 0.67 | 92.4 | ||
| 7 | 2-Deoxy-D-ribitol | 28.73 | 0.73 | 93 | ||
| 8 | Quinic acid | 33.89 | 7.75 | 89.4 | Enhances glucose-stimulated insulin secretion in both INS-1E cells and mouse islets | [34] |
| 9 | D-(-)-Fructose | 34.11 | 8.16 | 95.9 | ||
| 10 | L-(-)-Sorbose | 34.25 | 4.30 | 96 | ||
| 11 | D-Sorbitol | 34.88 | 4.79 | 98 | ||
| 12 | Myo-Inositol | 35.18 | 22.82 | 96.3 | Stimulates translocation of GLUT4 in skeletal muscle of C57BL/6 mice and induces translocation of GLUT4 to the plasma membrane | [18,33] |
| 13 | D-Gluconic acid | 35.76 | 0.75 | 91.5 | ||
| 14 | Caffeic acid | 36.93 | 1.71 | 95.1 | Reduces insulin resistance and modulates glucose uptake in HepG2 cells | [38] |
| 15 | D-(+)-Trehalose | 43.33 | 3.94 | 95.1 | ||
| 16 | Chlorogenic acid | 50.61 | 2.06 | 80.5 | Reduces insulin resistance and modulates glucose uptake in HepG2 cells | [38] |
| Fraction 10 | ||||||
| Peak | Name | Rt | % Area | Match Factor | Association with Diabetes | References |
| 1 | L-Proline | 16.99 | 4.91 | 91.8 | ||
| 2 | L-Valine | 18.50 | 1.11 | 95.5 | ||
| 3 | Urea | 19.08 | 0.69 | 96.4 | ||
| 4 | L-Leucine | 20.14 | 0.08 | 90.1 | ||
| 5 | Glycerol | 20.34 | 2.69 | 97.3 | ||
| 6 | Butanedioic acid | 21.19 | 0.43 | 96.1 | ||
| 7 | Glyceric acid | 21.88 | 0.99 | 95.8 | ||
| 8 | Serine | 22.66 | 0.32 | 95.3 | ||
| 9 | L-Threonine | 23.37 | 0.71 | 90.7 | ||
| 10 | 3-Aminoisobutyric acid | 25.01 | 0.26 | 91.9 | ||
| 11 | Pyroglutamic acid | 26.52 | 3.94 | 96.7 | Reduces oral glucose tolerance and serum insulin levels in rats | [40] |
| 12 | 4-Aminobutanoic acid | 26.75 | 3.15 | 94.8 | ||
| 13 | Threonic acid | 27.44 | 0.25 | 92.6 | ||
| 14 | L-Threonic acid | 27.86 | 0.31 | 93.3 | ||
| 15 | Phenylalanine | 28.98 | 0.67 | 96.1 | ||
| 16 | Asparagine | 30.11 | 0.18 | 85.2 | ||
| 17 | D-(+)-Arabitol | 31.00 | 0.35 | 88.8 | ||
| 18 | Xylitol | 31.00 | 0.35 | 87.4 | ||
| 19 | Quinic acid | 33.90 | 58.03 | 89.9 | Enhances glucose-stimulated insulin secretion in both INS-1E cells and mouse islets | [34] |
| 20 | D-(-)-Fructose | 34.11 | 2.16 | 91.8 | ||
| 21 | L-(-)-Sorbose | 34.25 | 0.66 | 93.8 | ||
| 22 | D-(+)-Talose | 34.45 | 0.37 | 86.1 | ||
| 23 | L-Tyrosine | 34.72 | 0.32 | 85 | ||
| 24 | D-Sorbitol | 34.87 | 0.92 | 96.9 | ||
| 25 | Myo-inositol | 35.18 | 12.01 | 96 | Stimulates translocation of GLUT4 in skeletal muscle of C57BL/6 mice and induces translocation of GLUT4 to the plasma membrane | [18,33] |
| 26 | D-Gluconic acid | 35.81 | 0.70 | 84.9 | ||
| 27 | Caffeic acid | 36.93 | 0.67 | 94 | Reduces insulin resistance and modulates glucose uptake in HepG2 cells | [38] |
| 28 | D-(+)-Trehalose | 43.33 | 4.34 | 96.2 | ||
| 29 | Chlorogenic acid | 50.61 | 1.56 | 80.8 | Reduces insulin resistance and modulates glucose uptake in HepG2 cells | [38] |
| Fraction Number | Cytotoxicity (µg/mL), Safe Up to: | GLUT4 Translocation (% Relative to Controls) at 125 µg/mL | |
|---|---|---|---|
| − Insulin Relative to Control without Insulin | + Insulin Relative to Control with Insulin | ||
| 1 | 500 | 1.14 | 0.94 |
| 2 | 500 | 1.18 | 1.40 |
| 3 | 500 | 1.10 | 1.00 |
| 4 | 250 | 1.59 | 1.08 |
| 5 | 125 | 1.30 | 1.40 |
| 6 | 250 | 1.65 | 1.95 |
| 7 | 250 | 1.38 | 0.96 |
| 8 | 500 | 1.36 | 1.19 |
| 9 | 500 | 1.46 | 1.24 |
| 10 | 500 | 1.42 | 1.48 |
4. Discussion
4.1. Lupeol and Lupeol-Trifluoroacetate
4.2. Palmitic Acid
4.3. β-Amyrin
4.4. Ursolic Acid and Ursolic Aldehyde
4.5. 4-Hydroxybenzoic Acid and 3,4-Dihydroxybenzoic Acid
4.6. Caffeic Acid
4.7. Myo-Inositol and Quinic Acid
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zaid, H.; Tamrakar, A.K.; Razzaque, M.S.; Efferth, T. Diabetes and metabolism disorders medicinal plants: A glance at the past and a look to the future 2018. Evid. Based Complement. Altern. Med. 2018, 2018, 5843298. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Ma, Z.F.; Lu, Y.; Du, Y.; Shao, J.; Wang, L.; Wu, Q.; Pan, B.; Zhu, W.; Zhao, Q.; et al. Elevated serum uric acid, hyperuricaemia and dietary patterns among adolescents in mainland china. J. Pediatr. Endocrinol. Metab. 2020, 33, 487–493. [Google Scholar] [CrossRef]
- Huang, J.; Ma, Z.F.; Zhang, Y.; Wan, Z.; Li, Y.; Zhou, H.; Chu, A.; Lee, Y.Y. Geographical distribution of hyperuricemia in mainland china: A comprehensive systematic review and meta-analysis. Glob. Health Res. Policy 2020, 5, 52. [Google Scholar] [CrossRef]
- Zaid, H.; Mahdi, A.A.; Tamrakar, A.K.; Saad, B.; Razzaque, M.S.; Dasgupta, A. Natural active ingredients for diabetes and metabolism disorders treatment. Evid. Based Complement. Altern. Med. 2016, 2016, 2965214. [Google Scholar] [CrossRef]
- Ma, Z.F.; Zhou, H.; Ma, J.; Lu, Y.; Pan, B. Prevalence of diabetes mellitus and hypertension during pregnancy in eastern china after the implementation of universal two-child policy. Int. J. Diabetes Dev. Ctries 2020, 41, 221–227. [Google Scholar] [CrossRef]
- Klip, A.; McGraw, T.E.; James, D.E. Thirty sweet years of glut4. J. Biol. Chem. 2019, 294, 11369–11381. [Google Scholar] [CrossRef] [Green Version]
- Zaid, H.; Antonescu, C.N.; Randhawa, V.K.; Klip, A. Insulin action on glucose transporters through molecular switches, tracks and tethers. Biochem. J. 2008, 413, 201–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu-Lafi, S.; Rayan, B.; Kadan, S.; Abu-Lafi, M.; Rayan, A. Anticancer activity and phytochemical composition of wild gundelia tournefortii. Oncol. Lett. 2019, 17, 713–717. [Google Scholar] [CrossRef] [Green Version]
- Kadan, S.; Sasson, Y.; Saad, B.; Zaid, H. Gundelia tournefortii antidiabetic efficacy: Chemical composition and glut4 translocation. Evid. Based Complement. Altern. Med. 2018, 2018, 8294320. [Google Scholar] [CrossRef] [Green Version]
- Azeez, O.H.; Kheder, A.E. Effect of gundelia tournefortii on some biochemical parameters in dexamethasone-induced hyperglycemic and hyperlipidemic mice. Iraqi J. Vet. Sci. 2012, 26, 7. [Google Scholar]
- Asadi-Samani, M.; Rafieian-Kopaei, M.; Azimi, N. Gundelia: A systematic review of medicinal and molecular perspective. Pak. J. Biol. Sci. 2013, 16, 1238–1247. [Google Scholar] [CrossRef]
- Mahajna, S.; Kadan, S.; Tietel, Z.; Saad, B.; Khasib, S.; Tumeh, A.; Ginsberg, D.; Zaid, H. In vitro evaluation of chemically analyzed hypericum triquetrifolium extract efficacy in apoptosis induction and cell cycle arrest of the hct-116 colon cancer cell line. Molecules 2019, 24, 4139. [Google Scholar] [CrossRef] [Green Version]
- Kadan, S.; Saad, B.; Sasson, Y.; Zaid, H. In vitro evaluation of anti-diabetic activity and cytotoxicity of chemically analysed ocimum basilicum extracts. Food Chem. 2016, 196, 1066–1074. [Google Scholar] [CrossRef] [PubMed]
- Zaid, H.; Talior-Volodarsky, I.; Antonescu, C.; Liu, Z.; Klip, A. Gapdh binds glut4 reciprocally to hexokinase-ii and regulates glucose transport activity. Biochem. J. 2009, 419, 475–484. [Google Scholar] [CrossRef] [Green Version]
- Saraswati, S.; Agrawal, S.S. Brucine, an indole alkaloid from strychnos nux-vomica attenuates vegf-induced angiogenesis via inhibiting vegfr2 signaling pathway in vitro and in vivo. Cancer Lett. 2013, 332, 83–93. [Google Scholar] [CrossRef]
- Peungvicha, P.; Temsiririrkkul, R.; Prasain, J.K.; Tezuka, Y.; Kadota, S.; Thirawarapan, S.S.; Watanabe, H. 4-hydroxybenzoic acid: A hypoglycemic constituent of aqueous extract of pandanus odorus root. J. Ethnopharmacol. 1998, 62, 79–84. [Google Scholar] [CrossRef]
- Peungvicha, P.; Thirawarapan, S.S.; Watanabe, H. Possible mechanism of hypoglycemic effect of 4-hydroxybenzoic acid, a constituent of pandanus odorus root. Jpn. J. Pharmacol. 1998, 78, 395–398. [Google Scholar] [CrossRef] [Green Version]
- Dang, N.T.; Mukai, R.; Yoshida, K.; Ashida, H. D-pinitol and myo-inositol stimulate translocation of glucose transporter 4 in skeletal muscle of c57bl/6 mice. Biosci. Biotechnol. Biochem. 2010, 74, 1062–1067. [Google Scholar] [CrossRef] [Green Version]
- Kimura, T.; Kato, E.; Machikawa, T.; Kimura, S.; Katayama, S.; Kawabata, J. Hydroxylamine enhances glucose uptake in c2c12 skeletal muscle cells through the activation of insulin receptor substrate 1. Biochem. Biophys. Res. Commun. 2014, 445, 6–9. [Google Scholar] [CrossRef] [Green Version]
- Wada, Y.; Sakiyama, S.; Sakai, H.; Sakane, F. Myristic acid enhances diacylglycerol kinase delta-dependent glucose uptake in myotubes. Lipids 2016, 51, 897–903. [Google Scholar] [CrossRef]
- Bhattacharjee, S.; Das, N.; Mandala, A.; Mukhopadhyay, S.; Roy, S.S. Fenofibrate reverses palmitate induced impairment in glucose uptake in skeletal muscle cells by preventing cytosolic ceramide accumulation. Cell. Physiol. Biochem. 2015, 37, 1315–1328. [Google Scholar] [CrossRef] [PubMed]
- Pu, J.; Peng, G.; Li, L.; Na, H.; Liu, Y.; Liu, P. Palmitic acid acutely stimulates glucose uptake via activation of akt and erk1/2 in skeletal muscle cells. J. Lipid Res. 2011, 52, 1319–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yore, M.M.; Syed, I.; Moraes-Vieira, P.M.; Zhang, T.; Herman, M.A.; Homan, E.A.; Patel, R.T.; Lee, J.; Chen, S.; Peroni, O.D.; et al. Discovery of a class of endogenous mammalian lipids with anti-diabetic and anti-inflammatory effects. Cell 2014, 159, 318–332. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.F.; Maurya, C.K.; Dev, K.; Arha, D.; Rai, A.K.; Tamrakar, A.K.; Maurya, R. Design and synthesis of lupeol analogues and their glucose uptake stimulatory effect in l6 skeletal muscle cells. Bioorg. Med. Chem. Lett. 2014, 24, 2674–2679. [Google Scholar] [CrossRef]
- Etsassala, N.; Badmus, J.A.; Waryo, T.T.; Marnewick, J.L.; Cupido, C.N.; Hussein, A.A.; Iwuoha, E.I. Alpha-glucosidase and alpha-amylase inhibitory activities of novel abietane diterpenes from salvia africana-lutea. Antioxidants 2019, 8, 421. [Google Scholar] [CrossRef] [Green Version]
- Santos, F.A.; Frota, J.T.; Arruda, B.R.; de Melo, T.S.; da Silva, A.A.; Brito, G.A.; Chaves, M.H.; Rao, V.S. Antihyperglycemic and hypolipidemic effects of alpha, beta-amyrin, a triterpenoid mixture from protium heptaphyllum in mice. Lipids Health Dis. 2012, 11, 98. [Google Scholar] [CrossRef] [Green Version]
- McVeay, C.; Fitzgerald, P.C.E.; Horowitz, M.; Feinle-Bisset, C. Effects of duodenal infusion of lauric acid and l-tryptophan, alone and combined, on fasting glucose, insulin and glucagon in healthy men. Nutrients 2019, 11, 2697. [Google Scholar] [CrossRef] [Green Version]
- Muthulakshmi, S.; Saravanan, R. Efficacy of azelaic acid on hepatic key enzymes of carbohydrate metabolism in high fat diet induced type 2 diabetic mice. Biochimie 2013, 95, 1239–1244. [Google Scholar] [CrossRef]
- Wang, J.; Huang, M.; Yang, J.; Ma, X.; Zheng, S.; Deng, S.; Huang, Y.; Yang, X.; Zhao, P. Anti-diabetic activity of stigmasterol from soybean oil by targeting the glut4 glucose transporter. Food Nutr. Res. 2017, 61, 1364117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponnulakshmi, R.; Shyamaladevi, B.; Vijayalakshmi, P.; Selvaraj, J. In silico and in vivo analysis to identify the antidiabetic activity of beta sitosterol in adipose tissue of high fat diet and sucrose induced type-2 diabetic experimental rats. Toxicol. Mech. Methods 2019, 29, 276–290. [Google Scholar] [CrossRef]
- He, Y.; Li, W.; Li, Y.; Zhang, S.; Wang, Y.; Sun, C. Ursolic acid increases glucose uptake through the pi3k signaling pathway in adipocytes. PLoS ONE 2014, 9, e110711. [Google Scholar] [CrossRef] [Green Version]
- Houghton, M.J.; Kerimi, A.; Mouly, V.; Tumova, S.; Williamson, G. Gut microbiome catabolites as novel modulators of muscle cell glucose metabolism. FASEB J. 2019, 33, 1887–1898. [Google Scholar] [CrossRef] [Green Version]
- Yap, A.; Nishiumi, S.; Yoshida, K.; Ashida, H. Rat l6 myotubes as an in vitro model system to study glut4-dependent glucose uptake stimulated by inositol derivatives. Cytotechnology 2007, 55, 103–108. [Google Scholar] [CrossRef] [Green Version]
- Heikkila, E.; Hermant, A.; Thevenet, J.; Bermont, F.; Kulkarni, S.S.; Ratajczak, J.; Santo-Domingo, J.; Dioum, E.H.; Canto, C.; Barron, D.; et al. The plant product quinic acid activates Ca(2+)-dependent mitochondrial function and promotes insulin secretion from pancreatic beta cells. Br. J. Pharmacol. 2019, 176, 3250–3263. [Google Scholar] [CrossRef]
- Krzysztoforska, K.; Mirowska-Guzel, D.; Widy-Tyszkiewicz, E. Pharmacological effects of protocatechuic acid and its therapeutic potential in neurodegenerative diseases: Review on the basis of in vitro and in vivo studies in rodents and humans. Nutr. Neurosci. 2019, 22, 72–82. [Google Scholar] [CrossRef]
- Scazzocchio, B.; Vari, R.; Filesi, C.; Del Gaudio, I.; D’Archivio, M.; Santangelo, C.; Iacovelli, A.; Galvano, F.; Pluchinotta, F.R.; Giovannini, C.; et al. Protocatechuic acid activates key components of insulin signaling pathway mimicking insulin activity. Mol. Nutr. Food Res. 2015, 59, 1472–1481. [Google Scholar] [CrossRef]
- Kakkar, S.; Bais, S. A review on protocatechuic acid and its pharmacological potential. ISRN Pharmacol. 2014, 2014, 952943. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Teng, H.; Cao, H. Chlorogenic acid and caffeic acid from sonchus oleraceus linn synergistically attenuate insulin resistance and modulate glucose uptake in hepg2 cells. Food Chem. Toxicol. 2019, 127, 182–187. [Google Scholar] [CrossRef]
- Lee, M.S.; Sohn, C.B. Anti-diabetic properties of chrysophanol and its glucoside from rhubarb rhizome. Biol. Pharm. Bull. 2008, 31, 2154–2157. [Google Scholar] [CrossRef] [Green Version]
- Yoshinari, O.; Igarashi, K. Anti-diabetic effect of pyroglutamic acid in type 2 diabetic goto-kakizaki rats and kk-ay mice. Br. J. Nutr. 2011, 106, 995–1004. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.; Liu, D. Long-term exposure to genistein improves insulin secretory function of pancreatic beta-cells. Eur. J. Pharmacol. 2009, 616, 321–327. [Google Scholar] [CrossRef] [Green Version]
- Kooti, W.; Farokhipour, M.; Asadzadeh, Z.; Ashtary-Larky, D.; Asadi-Samani, M. The role of medicinal plants in the treatment of diabetes: A systematic review. Electron. Physician 2016, 8, 1832–1842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.H.; Chen, L.X.; Li, H. Progress in the discovery of naturally occurring anti-diabetic drugs and in the identification of their molecular targets. Fitoterapia 2019, 134, 270–289. [Google Scholar] [CrossRef] [PubMed]
- Xia, N.; Li, H. The role of perivascular adipose tissue in obesity-induced vascular dysfunction. Br. J. Pharmacol. 2017, 174, 3425–3442. [Google Scholar] [CrossRef] [Green Version]
- Jaimes-Hoy, L.; Gurrola, G.B.; Cisneros, M.; Joseph-Bravo, P.; Possani, L.D.; Charli, J.L. The kv1.3 channel blocker vm24 enhances muscle glucose transporter 4 mobilization but does not reduce body-weight gain in diet-induced obese male rats. Life Sci. 2017, 181, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Malik, A.; Jamil, U.; Butt, T.T.; Waquar, S.; Gan, S.H.; Shafique, H.; Jafar, T.H. In silico and in vitro studies of lupeol and iso-orientin as potential antidiabetic agents in a rat model. Drug Des. Dev. Ther. 2019, 13, 1501–1513. [Google Scholar] [CrossRef] [Green Version]
- Nugent, C.; Prins, J.B.; Whitehead, J.P.; Wentworth, J.M.; Chatterjee, V.K.; O’Rahilly, S. Arachidonic acid stimulates glucose uptake in 3t3-l1 adipocytes by increasing glut1 and glut4 levels at the plasma membrane. Evidence for involvement of lipoxygenase metabolites and peroxisome proliferator-activated receptor gamma. J. Biol. Chem. 2001, 276, 9149–9157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thode, J.; Pershadsingh, H.A.; Ladenson, J.H.; Hardy, R.; McDonald, J.M. Palmitic acid stimulates glucose incorporation in the adipocyte by a mechanism likely involving intracellular calcium. J. Lipid Res. 1989, 30, 1299–1305. [Google Scholar] [CrossRef]
- Sawada, K.; Kawabata, K.; Yamashita, T.; Kawasaki, K.; Yamamoto, N.; Ashida, H. Ameliorative effects of polyunsaturated fatty acids against palmitic acid-induced insulin resistance in l6 skeletal muscle cells. Lipids Health Dis. 2012, 11, 36. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Wei, D.; Mo, C.; Zhang, J.; Wang, X.; Han, X.; Wang, Z.; Xiao, H. Saturated fatty acid palmitate-induced insulin resistance is accompanied with myotube loss and the impaired expression of health benefit myokine genes in c2c12 myotubes. Lipids Health Dis. 2013, 12, 104. [Google Scholar] [CrossRef] [Green Version]
- de Melo, K.M.; de Oliveira, F.T.B.; Costa Silva, R.A.; Gomes Quindere, A.L.; Marinho Filho, J.D.B.; Araujo, A.J.; Barros Pereira, E.D.; Carvalho, A.A.; Chaves, M.H.; Rao, V.S.; et al. Alpha, beta-amyrin, a pentacyclic triterpenoid from protium heptaphyllum suppresses adipocyte differentiation accompanied by down regulation of ppargamma and c/ebpalpha in 3t3-l1 cells. Biomed. Pharmacother. 2019, 109, 1860–1866. [Google Scholar] [CrossRef]
- Nair, S.A.; Sabulal, B.; Radhika, J.; Arunkumar, R.; Subramoniam, A. Promising anti-diabetes mellitus activity in rats of beta-amyrin palmitate isolated from hemidesmus indicus roots. Eur. J. Pharmacol. 2014, 734, 77–82. [Google Scholar] [CrossRef]
- Na, M.; Oh, W.K.; Kim, Y.H.; Cai, X.F.; Kim, S.; Kim, B.Y.; Ahn, J.S. Inhibition of protein tyrosine phosphatase 1b by diterpenoids isolated from acanthopanax koreanum. Bioorg. Med. Chem. Lett. 2006, 16, 3061–3064. [Google Scholar] [CrossRef]
- Ramirez-Espinosa, J.J.; Rios, M.Y.; Lopez-Martinez, S.; Lopez-Vallejo, F.; Medina-Franco, J.L.; Paoli, P.; Camici, G.; Navarrete-Vazquez, G.; Ortiz-Andrade, R.; Estrada-Soto, S. Antidiabetic activity of some pentacyclic acid triterpenoids, role of ptp-1b: In vitro, in silico, and in vivo approaches. Eur. J. Med. Chem. 2011, 46, 2243–2251. [Google Scholar] [CrossRef]
- Sundaresan, A.; Radhiga, T.; Pugalendi, K.V. Ursolic acid and rosiglitazone combination improves insulin sensitivity by increasing the skeletal muscle insulin-stimulated irs-1 tyrosine phosphorylation in high-fat diet-fed c57bl/6j mice. J. Physiol. Biochem. 2016, 72, 345–352. [Google Scholar] [CrossRef]
- Wu, P.; He, P.; Zhao, S.; Huang, T.; Lu, Y.; Zhang, K. Effects of ursolic acid derivatives on caco-2 cells and their alleviating role in streptozocin-induced type 2 diabetic rats. Molecules 2014, 19, 12559–12576. [Google Scholar] [CrossRef] [Green Version]
- Harini, R.; Pugalendi, K.V. Antihyperglycemic effect of protocatechuic acid on streptozotocin-diabetic rats. J. Basic Clin. Physiol. Pharmacol. 2010, 21, 79–91. [Google Scholar] [CrossRef]
- Rebollo-Hernanz, M.; Zhang, Q.; Aguilera, Y.; Martin-Cabrejas, M.A.; Gonzalez de Mejia, E. Relationship of the phytochemicals from coffee and cocoa by-products with their potential to modulate biomarkers of metabolic syndrome in vitro. Antioxidants 2019, 8, 279. [Google Scholar] [CrossRef] [Green Version]
- Cabrera-Cruz, H.; Orostica, L.; Plaza-Parrochia, F.; Torres-Pinto, I.; Romero, C.; Vega, M. The insulin-sensitizing mechanism of myo-inositol is associated with ampk activation and glut-4 expression in human endometrial cells exposed to a pcos environment. Am. J. Physiol. Endocrinol. Metab. 2020, 318, E237–E248. [Google Scholar] [CrossRef] [PubMed]










| Hexane | EtoAc | EtOH | Time (min) |
|---|---|---|---|
| From 0% | To 100% | 30.2 | |
| 100% | 15 | ||
| From 0% | To 100% | 15 | |
| 100% | 15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kadan, S.; Melamed, S.; Benvalid, S.; Tietel, Z.; Sasson, Y.; Zaid, H. Gundelia tournefortii: Fractionation, Chemical Composition and GLUT4 Translocation Enhancement in Muscle Cell Line. Molecules 2021, 26, 3785. https://doi.org/10.3390/molecules26133785
Kadan S, Melamed S, Benvalid S, Tietel Z, Sasson Y, Zaid H. Gundelia tournefortii: Fractionation, Chemical Composition and GLUT4 Translocation Enhancement in Muscle Cell Line. Molecules. 2021; 26(13):3785. https://doi.org/10.3390/molecules26133785
Chicago/Turabian StyleKadan, Sleman, Sarit Melamed, Shoshana Benvalid, Zipora Tietel, Yoel Sasson, and Hilal Zaid. 2021. "Gundelia tournefortii: Fractionation, Chemical Composition and GLUT4 Translocation Enhancement in Muscle Cell Line" Molecules 26, no. 13: 3785. https://doi.org/10.3390/molecules26133785
APA StyleKadan, S., Melamed, S., Benvalid, S., Tietel, Z., Sasson, Y., & Zaid, H. (2021). Gundelia tournefortii: Fractionation, Chemical Composition and GLUT4 Translocation Enhancement in Muscle Cell Line. Molecules, 26(13), 3785. https://doi.org/10.3390/molecules26133785