
An Untargeted Metabolomics Approach for Correlating Pulse Crop Seed Coat Polyphenol Profiles with Antioxidant Capacity and Iron Chelation Ability




Abstract
:1. Introduction
2. Results and Discussion
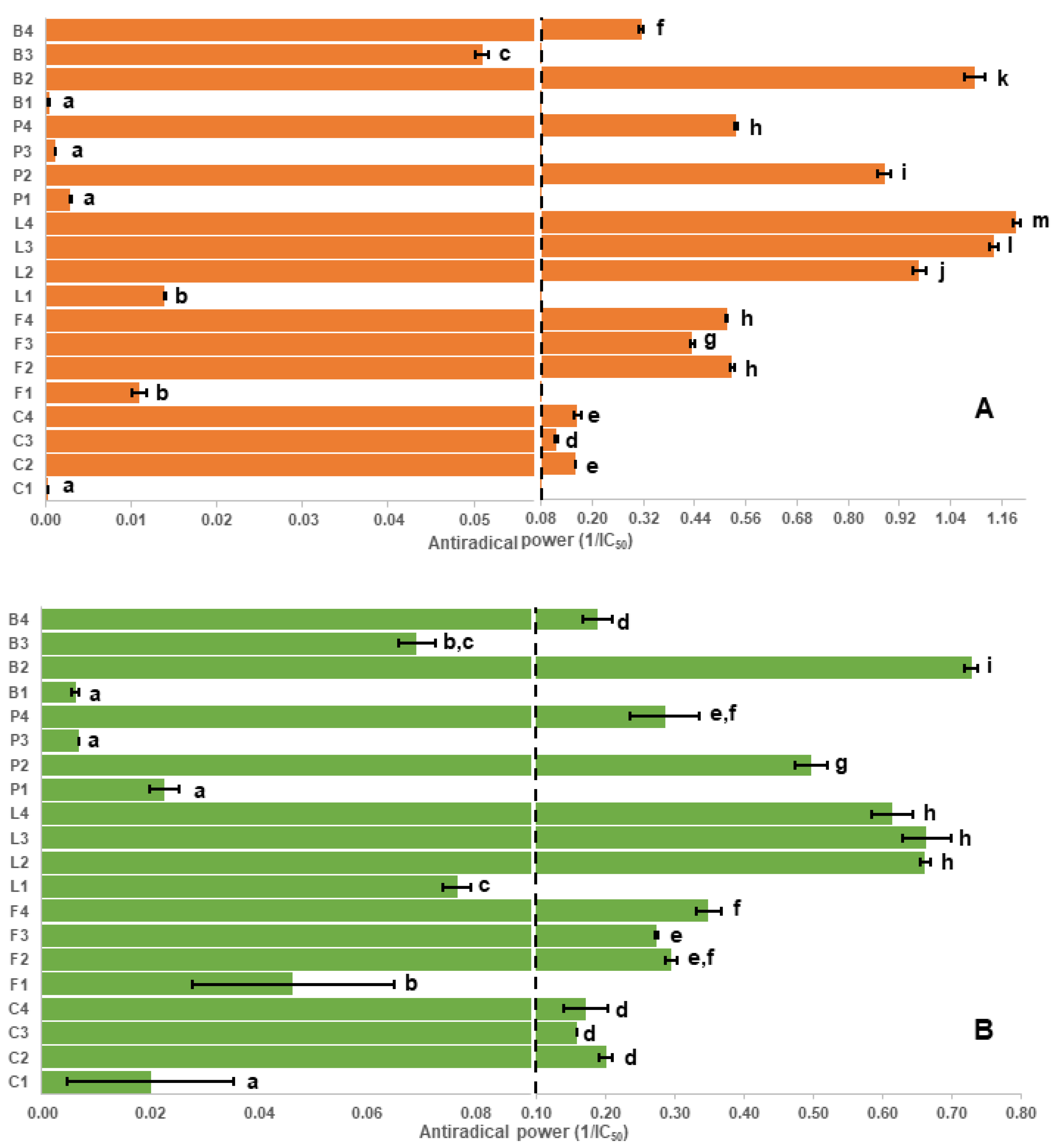
2.1. Assay Results and Evaluation of the Antioxidant Capacity Assays
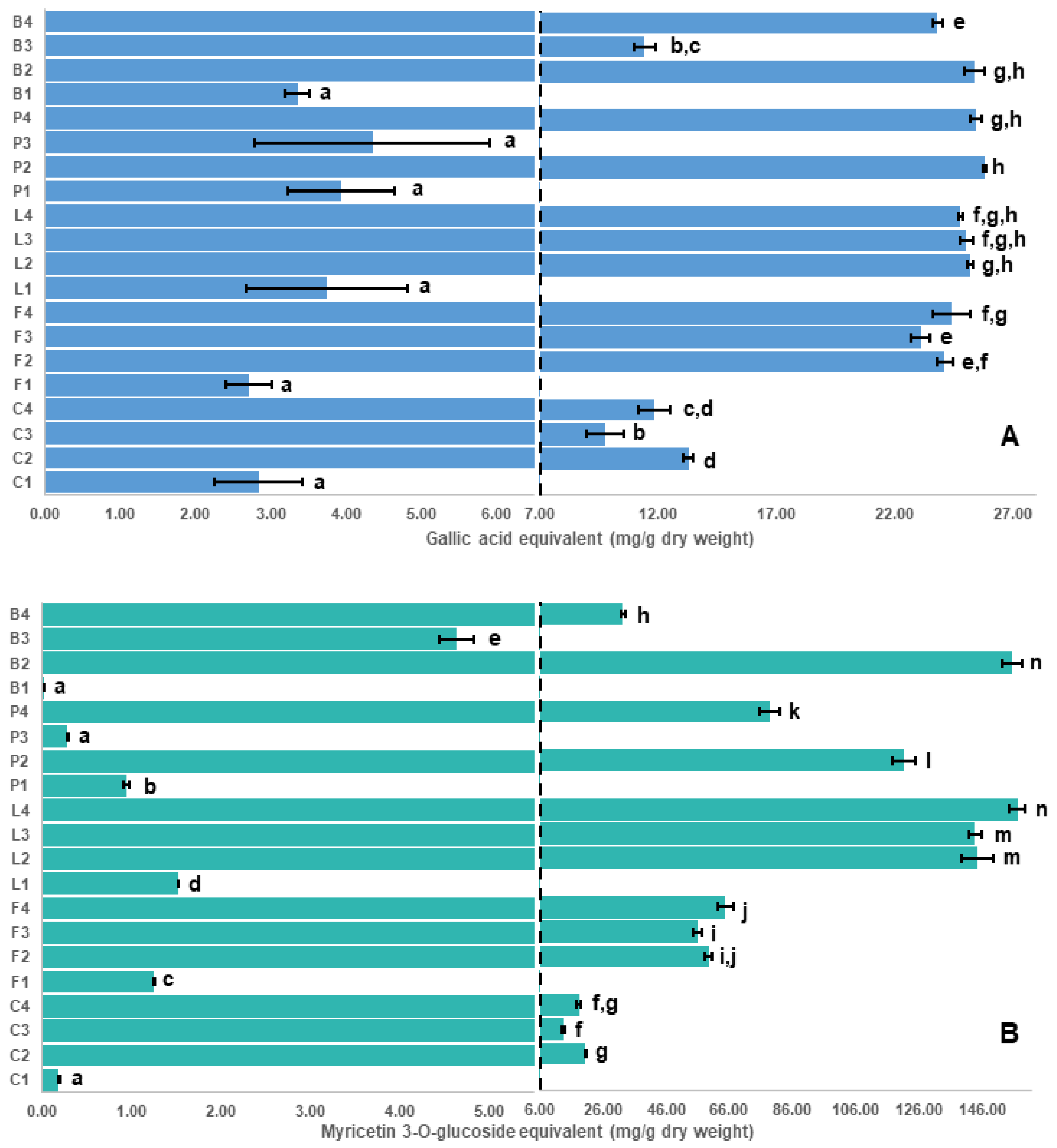
2.2. Correlation of Antioxidant Capacity to Polyphenol Classes
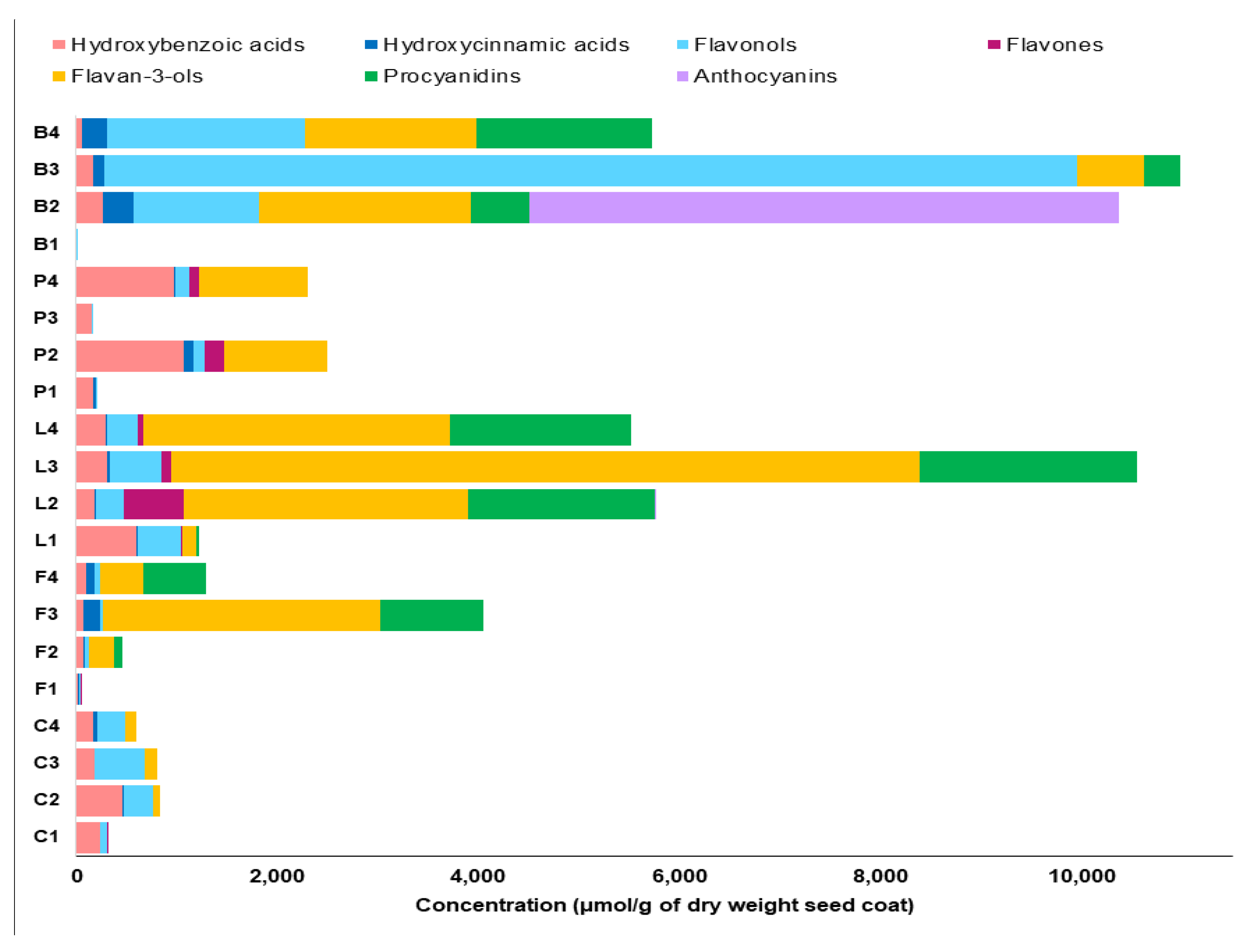
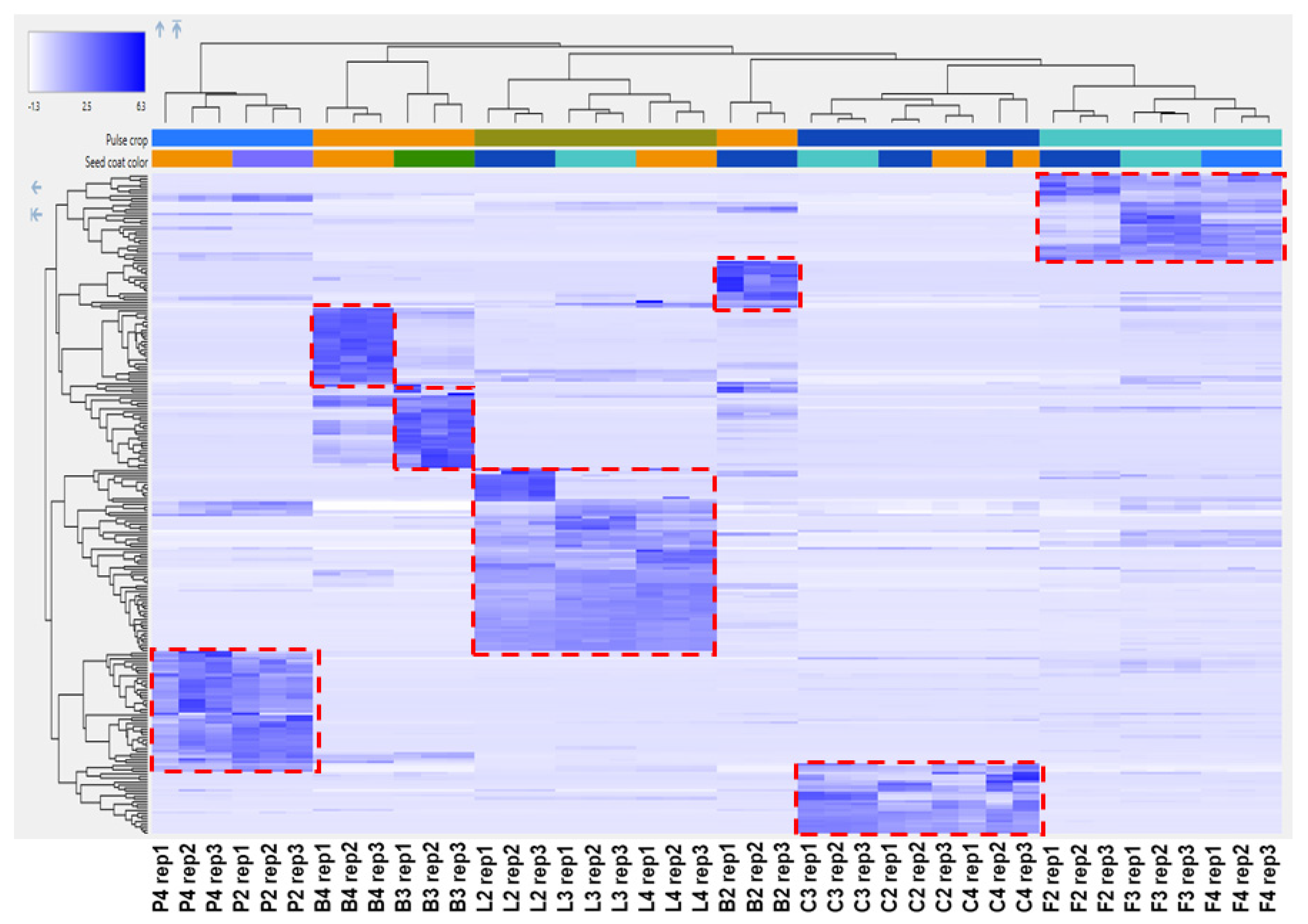
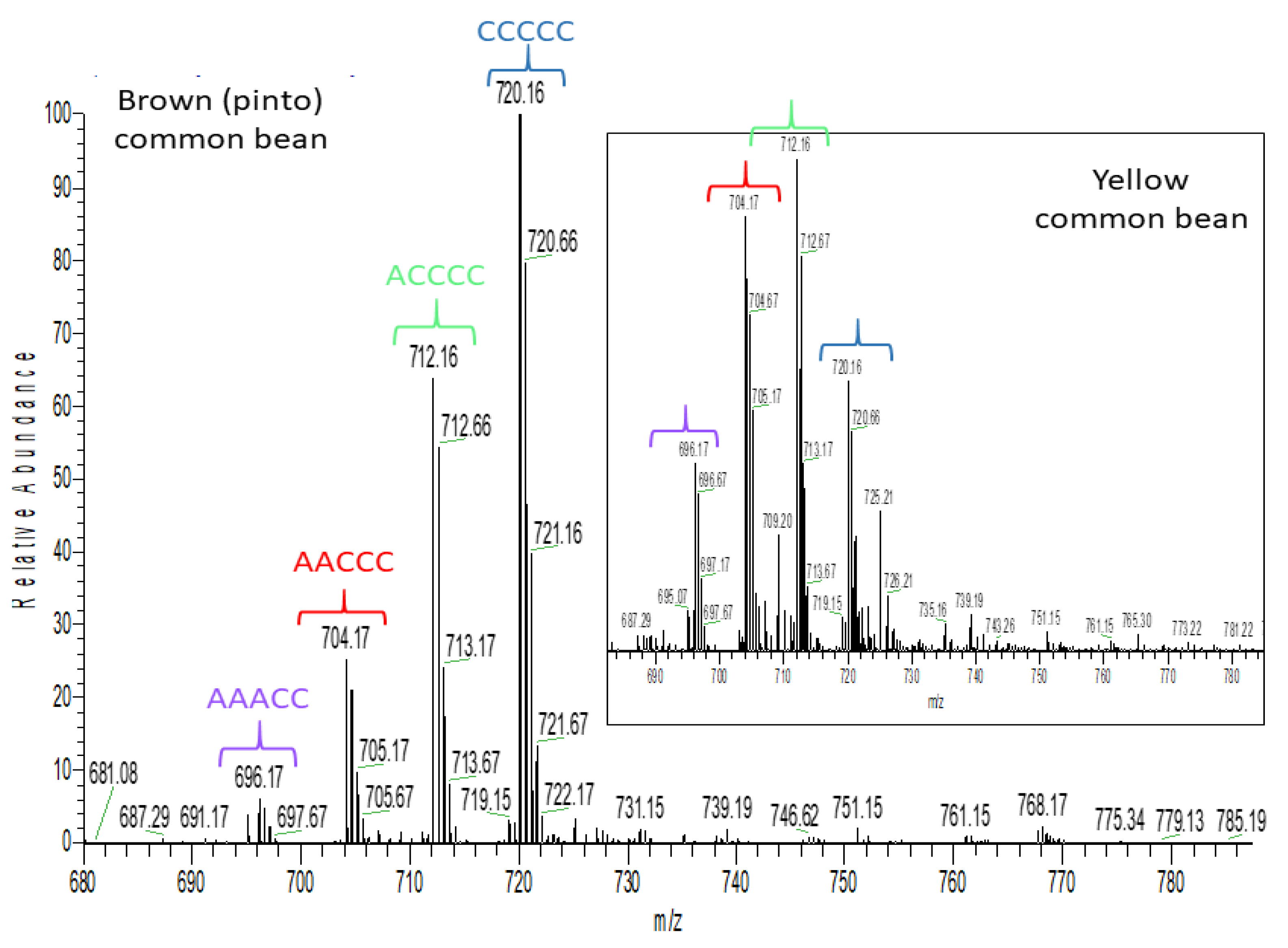
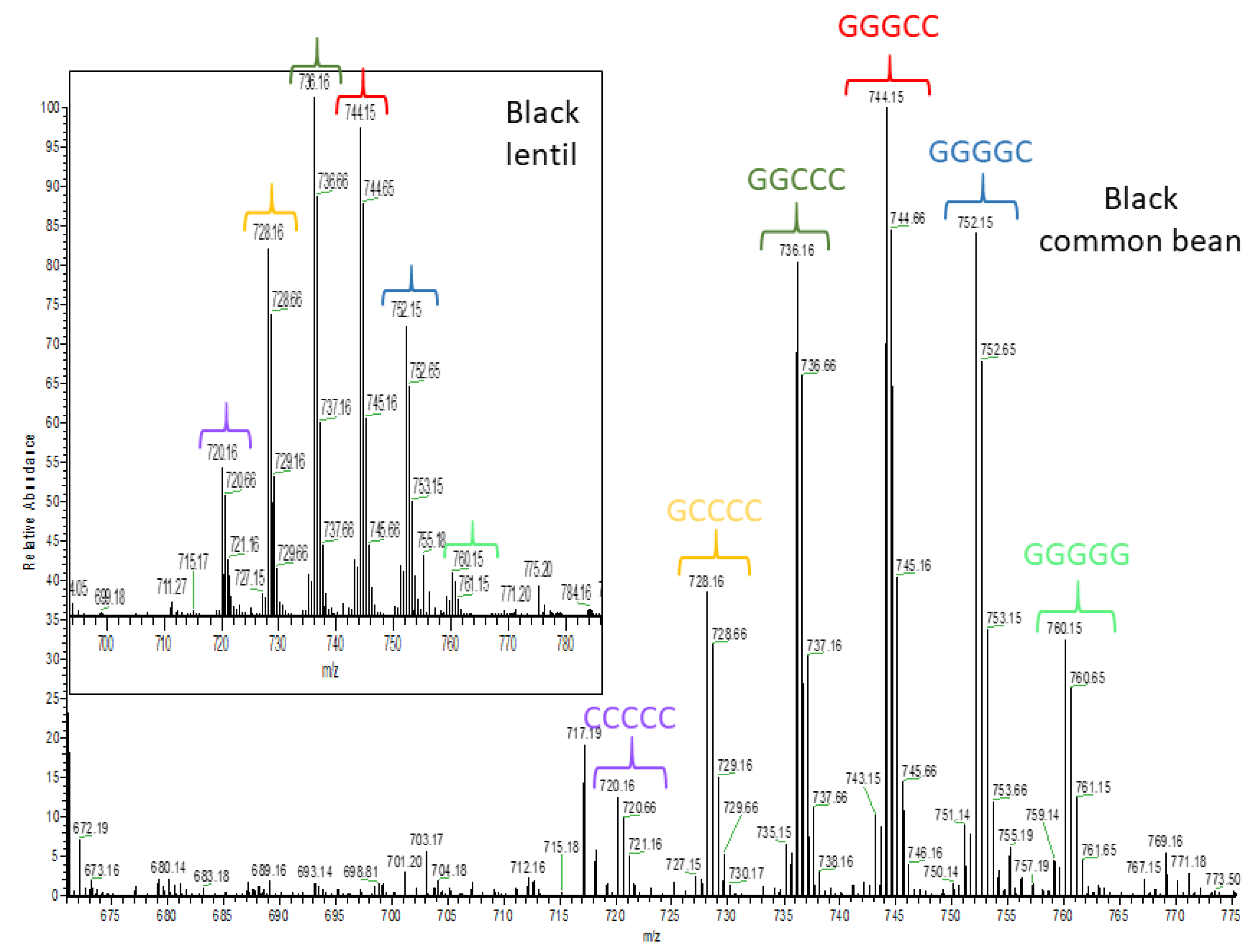
2.3. Untargeted Analysis of the Seed Coat Samples
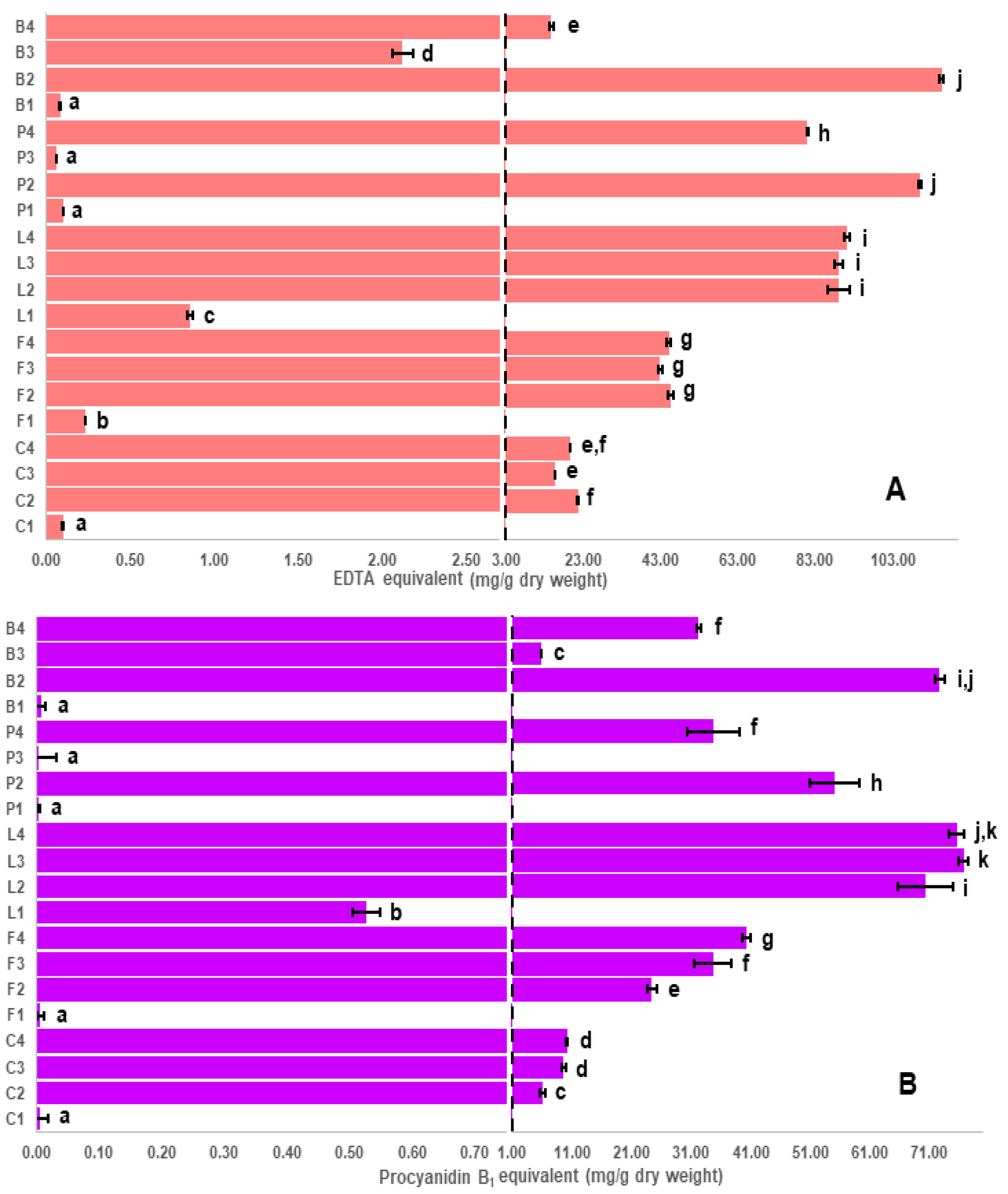
2.4. Correlation of Iron Chelation to Polyphenol Classes
3. Materials and Methods
3.1. Plant Material
3.2. Chemicals and Reagents
3.3. Preparation of Seed Coat Extracts
3.4. Ferrozine Iron Chelating Assay
3.5. Antioxidant Capacity Assays
3.5.1. DPPH Assay (Mixed-Mode HAT and ET Based)
3.5.2. TBARS Assay
3.5.3. Folin–Ciocalteu Assay (ET-Based)
3.5.4. Ferric Reducing Antioxidant Power (FRAP) Assay (ET-Based)
3.6. Spectrophotometric Measurement of Polymeric Polyphenols
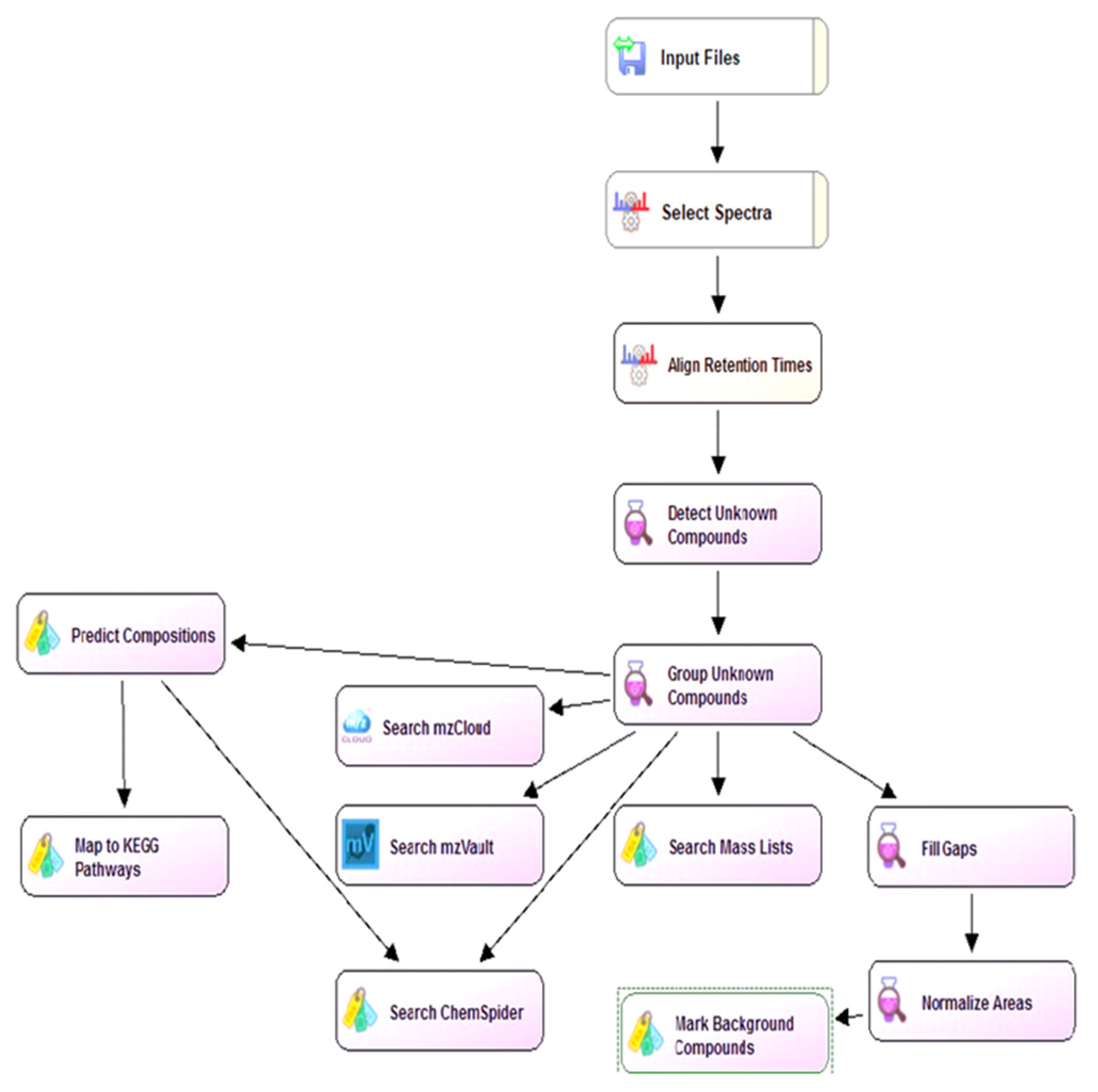
3.7. Untargeted Analysis of Seed Coat Extracts Using LC-HRMS
3.8. Data Analysis
3.9. Statistics
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Iriti, M.; Varoni, E.M. Pulses, healthy, and sustainable food sources for feeding the planet. Int. J. Mol. Sci. 2017, 18, 255. [Google Scholar] [CrossRef]
- Singh, B.; Singh, J.P.; Shevkani, K.; Singh, N.; Kaur, A. Bioactive constituents in pulses and their health benefits. J. Food Sci. Technol. 2017, 54, 858–870. [Google Scholar] [CrossRef] [Green Version]
- Crozier, A.; Jaganath, I.B.; Clifford, M.N. Dietary phenolics: Chemistry, bioavailability and effects on health. Nat. Prod. Rep. 2009, 26, 1001–1043. [Google Scholar] [CrossRef] [PubMed]
- Ignat, I.; Volf, I.; Popa, V.I. A critical review of methods for characterisation of polyphenolic compounds in fruits and vegetables. Food Chem. 2011, 126, 1821–1835. [Google Scholar] [CrossRef] [PubMed]
- Kasote, D.M.; Katyare, S.S.; Hegde, M.V.; Bae, H. Significance of Antioxidant Potential of Plants and its Relevance to Therapeutic Applications. Int. J. Biol. Sci. 2015, 11, 982–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants in plants: Location and functional significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The Role of Polyphenols in Human Health and Food Systems: A Mini-Review. Front. Nutr. 2018, 5. [Google Scholar] [CrossRef] [Green Version]
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Tiwari, R.; Chakraborty, S.; Dhama, K. Oxidative stress, prooxidants, and antioxidants: The interplay. BioMed Res. Int. 2014, 2014, 761264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Petry, N.; Egli, I.; Zeder, C.; Walczyk, T.; Hurrell, R. Polyphenols and phytic acid contribute to the low iron bioavailability from common beans in young women. J. Nutr. 2010, 140, 1977–1982. [Google Scholar] [CrossRef]
- Miller, J.L. Iron deficiency anemia: A common and curable disease. Cold Spring Harb. Perspect. Med. 2013, 3. [Google Scholar] [CrossRef] [Green Version]
- Hart, J.J.; Tako, E.; Kochian, L.V.; Glahn, R.P. Identification of black bean (Phaseolus vulgaris L.) polyphenols that inhibit and promote iron iptake by Caco-2 cells. J. Agric. Food Chem. 2015, 63, 5950–5956. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.L.; Blachier, F.; Tossou, M.C.B.; Rahu, N. Oxidative stress and inflammation: What polyphenols can do for us? Oxid. Med. Cell. Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Peng, H.; Deng, Z.; Tsao, R. Phytochemicals of lentil (Lens culinaris) and their antioxidant and anti-inflammatory effects. J. Food Bioact. 2018, 1, 93–103. [Google Scholar] [CrossRef] [Green Version]
- de Camargo, A.C.; Favero, B.T.; Morzelle, M.C.; Franchin, M.; Alvarez-Parrilla, E.; de la Rosa, L.A.; Geraldi, M.V.; Marostica, M.R.; Shahidi, F.; Schwember, A.R. Is Chickpea a Potential Substitute for Soybean? Phenolic Bioactives and Potential Health Benefits. Int. J. Mol. Sci. 2019, 20, 2644. [Google Scholar] [CrossRef] [Green Version]
- Dueñas, M.; Hernández, T.; Estrella, I. Assessment of in vitro antioxidant capacity of the seed coat and the cotyledon of legumes in relation to their phenolic contents. Food Chem. 2006, 98, 95–103. [Google Scholar] [CrossRef]
- Oomah, B.D.; Caspar, F.; Malcolmson, L.J.; Bellido, A.S. Phenolics and antioxidant activity of lentil and pea hulls. Food Res. Int. 2011, 44, 436–441. [Google Scholar] [CrossRef]
- Collar, C.; Santos, E.; Rosell, C.M. Significance of dietary fiber on the viscometric pattern of pasted and gelled flour-fiber blends. Cereal Chem. 2006, 83, 370–376. [Google Scholar] [CrossRef]
- Piteira, M.F.; Maia, J.M.; Raymundo, A.; Sousa, I. Extensional flow behaviour of natural fibre-filled dough and its relationship with structure and properties. J. Non-Newton. Fluid Mech. 2006, 137, 72–80. [Google Scholar] [CrossRef] [Green Version]
- Thakur, S.; Scanlon, M.G.; Tyler, R.T.; Milani, A.; Paliwal, J. Pulse Flour Characteristics from a Wheat Flour Miller’s Perspective: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 775–797. [Google Scholar] [CrossRef] [Green Version]
- Dias, R.; Oliveira, H.; Fernandes, I.; Simal-Gandara, J.; Perez-Gregorio, R. Recent advances in extracting phenolic compounds from food and their use in disease prevention and as cosmetics. Crit. Rev. Food Sci. Nutr. 2020, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Pathiraja, P.M.H.D.; Shand, P.J. Effectiveness of lentil (Lens culinaris) components for replacing phosphates in mechanically separated chicken bologna. In Proceedings of the 64th International Congress on Meat Science and Technology, Melbourne, Australia, 12–17 August 2018. [Google Scholar]
- Cherubim, D.J.D.; Martins, C.V.B.; Farina, L.O.; de Lucca, R.A.D. Polyphenols as natural antioxidants in cosmetics applications. J. Cosmet. Dermatol. 2020, 19, 33–37. [Google Scholar] [CrossRef]
- Ben-Othman, S.; Joudu, I.; Bhat, R. Bioactives from Agri-Food Wastes: Present Insights and Future Challenges. Molecules 2020, 25, 510. [Google Scholar] [CrossRef] [Green Version]
- Shahidi, F.; Varatharajan, V.; Oh, W.Y.; Peng, H. Phenolic compounds in agri-food by-products, their bioavailability and health effects. J. Food Bioact. 2019, 5, 57–119. [Google Scholar] [CrossRef] [Green Version]
- Elessawy, F.M.; Bazghaleh, N.; Vandenberg, A.; Purves, R.W. Polyphenol profile comparisons of seed coats of five pulse crops using a semi-quantitative liquid chromatography-mass spectrometric method. Phytochem. Anal. 2020, 31, 458–471. [Google Scholar] [CrossRef]
- Zhang, X.; Li, X.; Su, M.; Du, J.; Zhou, H.; Li, X.; Ye, Z. A comparative UPLC-Q-TOF/MS-based metabolomics approach for distinguishing peach (Prunus persica (L.) Batsch) fruit cultivars with varying antioxidant activity. Food Res. Int. 2020, 137. [Google Scholar] [CrossRef]
- Commisso, M.; Bianconi, M.; Poletti, S.; Negri, S.; Munari, F.; Ceoldo, S.; Guzzo, F. Metabolomic Profiling and Antioxidant Activity of Fruits Representing Diverse Apple and Pear Cultivars. Biology 2021, 10, 380. [Google Scholar] [CrossRef]
- Schaich, K.M.; Tian, X.; Xie, J. Hurdles and pitfalls in measuring antioxidant efficacy: A critical evaluation of ABTS, DPPH, and ORAC assays. J. Funct. Foods 2015, 14, 111–125. [Google Scholar] [CrossRef]
- Granato, D.; Shahidi, F.; Wrolstad, R.; Kilmartin, P.; Melton, L.D.; Hidalgo, F.J.; Miyashita, K.; van Camp, J.; Alasalvar, C.; Ismail, A.B.; et al. Antioxidant activity, total phenolics and flavonoids contents: Should we ban in vitro screening methods? Food Chem. 2018, 264, 471–475. [Google Scholar] [CrossRef]
- Santos, J.S.; Brizola, V.R.A.; Granato, D. High-throughput assay comparison and standardization for metal chelating capacity screening: A proposal and application. Food Chem. 2017, 214, 515–522. [Google Scholar] [CrossRef]
- Wallace, T.C.; Giusti, M.M. Evaluation of Parameters that Affect the 4-Dimethylaminocinnamaldehyde Assay for Flavanols and Proanthocyanidins. J. Food Sci. 2010, 75, C619–C625. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Oxid. Antioxid. Part A 1999, 299, 152–178. [Google Scholar]
- Apak, R.; Guclu, K.; Demirata, B.; Ozyurek, M.; Celik, S.E.; Bektasoglu, B.; Berker, K.I.; Ozyurt, D. Comparative evaluation of various total antioxidant capacity assays applied to phenolic compounds with the CUPRAC assay. Molecules 2007, 12, 1496–1547. [Google Scholar] [CrossRef] [Green Version]
- Marathe, S.A.; Rajalakshmi, V.; Jamdar, S.N.; Sharma, A. Comparative study on antioxidant activity of different varieties of commonly consumed legumes in India. Food Chem. Toxicol. 2011, 49, 2005–2012. [Google Scholar] [CrossRef]
- Troszynska, A.; Ciska, E. Phenolic compounds of seed coats of white and coloured varieties of pea (Pisum sativum L.) and their total antioxidant activity. Czech J. Food Sci. 2002, 20, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Jha, A.B.; Purves, R.W.; Elessawy, F.M.; Zhang, H.X.; Vandenberg, A.; Warkentin, T.D. Polyphenolic Profile of Seed Components of White and Purple Flower Pea Lines. Crop Sci. 2019, 59, 2711–2719. [Google Scholar] [CrossRef] [Green Version]
- Verstraeten, S.V.; Keen, C.L.; Schmitz, H.H.; Fraga, C.G.; Oteiza, P.I. Flavan-3-ols and procyanidins protect liposomes against lipid oxidation and disruption of the bilayer structure. Free Radic. Biol. Med. 2003, 34, 84–92. [Google Scholar] [CrossRef]
- Hagerman, A.E.; Riedl, K.M.; Jones, G.A.; Sovik, K.N.; Ritchard, N.T.; Hartzfeld, P.W.; Riechel, T.L. High molecular weight plant polyphenolics (tannins) as biological antioxidants. J. Agric. Food Chem. 1998, 46, 1887–1892. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, T.; Shiga, K.; Ohshima, K.; Kawakishi, S.; Osawa, T. Inhibition of lipid peroxidation and the active oxygen radical scavenging effect of anthocyanin pigments isolated from Phaseolus vulgaris L. Biochem. Pharmacol. 1996, 52, 1033–1039. [Google Scholar] [CrossRef]
- Aggarwal, R.; Ranganathan, P. Common pitfalls in statistical analysis: The use of correlation techniques. Perspect. Clin. Res. 2016, 7, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.P.; Chang, S.K.C.; Gu, Y.; Qian, S.Y. Antioxidant activity and phenolic compositions of lentil (Lens culinaris var. Morton) extract and its fractions. J. Agric. Food Chem. 2011, 59, 2268–2276. [Google Scholar] [CrossRef] [Green Version]
- Spranger, I.; Sun, B.; Mateus, A.M.; de Freitas, V.; Ricardo-Da-Silva, J.M. Chemical characterization and antioxidant activities of oligomeric and polymeric procyanidin fractions from grape seeds. Food Chem. 2008, 108, 519–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.Y.; Zhang, L.; Zong, S.L.; Xu, S.F.; Li, X.Y.; Ye, Y.P. Antioxidant Capacity and Proanthocyanidin Composition of the Bark of Metasequoia glyptostroboides. Evid.-Based Complement. Altern. Med. 2014, 2014. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.H.; Lin, L.Z.; Chen, P. Study of the mass spectrometric behaviors of anthocyanins in negative ionization mode and its applications for characterization of anthocyanins and non-anthocyanin polyphenols. Rapid Commun. Mass Spectrom. 2012, 26, 1123–1133. [Google Scholar] [CrossRef]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar] [CrossRef]
- Choi, J.S.; Chung, H.Y.; Kang, S.S.; Jung, M.J.; Kim, J.W.; No, J.K.; Jung, H.A. The structure-activity relationship of flavonoids as scavengers of peroxynitrite. Phytother. Res. 2002, 16, 232–235. [Google Scholar] [CrossRef]
- Cai, Y.Z.; Sun, M.; Xing, J.; Luo, Q.; Corke, H. Structure-radical scavenging activity relationships of phenolic compounds from traditional Chinese medicinal plants. Life Sci. 2006, 78, 2872–2888. [Google Scholar] [CrossRef]
- Plumb, G.W.; Price, K.R.; Williamson, G. Antioxidant properties of flavonol glycosides from tea. Redox Rep. 1999, 4, 13–16. [Google Scholar] [CrossRef]
- Iglesias, J.; Pazos, M.; Lois, S.; Medina, I. Contribution of Galloylation and Polymerization to the Antioxidant Activity of Polyphenols in Fish Lipid Systems. J. Agric. Food Chem. 2010, 58, 7423–7431. [Google Scholar] [CrossRef] [Green Version]
- Plumb, G.W.; de Pascual-Teresa, S.; Santos-Buelga, C.; Rivas-Gonzalo, J.C.; Williamson, G. Antioxidant properties of gallocatechin and prodelphinidins from pomegranate peel. Redox Rep. 2002, 7, 41–46. [Google Scholar] [CrossRef] [Green Version]
- Atrahimovich, D.; Vaya, J.; Khatib, S. The effects and mechanism of flavonoid-rePON1 interactions. Structure-activity relationship study. Bioorganic Med. Chem. 2013, 21, 3348–3355. [Google Scholar] [CrossRef]
- RiceEvans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Ellis, N.; Hattori, C.; Cheema, J.; Donarski, J.; Charlton, A.; Dickinson, M.; Venditti, G.; Kalo, P.; Szabo, Z.; Kiss, G.B.; et al. NMR Metabolomics Defining Genetic Variation in Pea Seed Metabolites. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef]
- Duenas, M.; Estrella, I.; Hernandez, T. Occurrence of phenolic compounds in the seed coat and the cotyledon of peas (Pisum sativum L.). Eur. Food Res. Technol. 2004, 219, 116–123. [Google Scholar] [CrossRef]
- Ferraro, K.; Jin, A.L.; Nguyen, T.D.; Reinecke, D.M.; Ozga, J.A.; Ro, D.K. Characterization of proanthocyanidin metabolism in pea (Pisum sativum) seeds. BMC Plant Biol. 2014, 14. [Google Scholar] [CrossRef] [Green Version]
- Jovanovic, S.V.; Simic, M.G.; Steenken, S.; Hara, Y. Iron complexes of gallocatechins. Antioxidant action or iron regulation? J. Chem. Soc. Perkin Trans. 1998, 2, 2365–2369. [Google Scholar] [CrossRef]
- Gu, L.W.; Kelm, M.A.; Hammerstone, J.F.; Zhang, Z.; Beecher, G.; Holden, J.; Haytowitz, D.; Prior, R.L. Liquid chromatographic/electrospray ionization mass spectrometric studies of proanthocyanidins in foods. J. Mass Spectrom. 2003, 38, 1272–1280. [Google Scholar] [CrossRef]
- Andjelkovic, M.; Van Camp, J.; De Meulenaer, B.; Depaemelaere, G.; Socaciu, C.; Verloo, M.; Verhe, R. Iron-chelation properties of phenolic acids bearing catechol and galloyl groups. Food Chem. 2006, 98, 23–31. [Google Scholar] [CrossRef]
- Hatcher, H.C.; Singh, R.N.; Torti, F.M.; Torti, S.V. Synthetic and natural iron chelators: Therapeutic potential and clinical use. Future Med. Chem. 2009, 1, 1643–1670. [Google Scholar] [CrossRef] [Green Version]
- Bartzoka, E.D.; Lange, H.; Poce, G.; Crestini, C. Stimuli-Responsive Tannin-Fe-III Hybrid Microcapsules Demonstrated by the Active Release of an Anti-Tuberculosis Agent. Chemsuschem 2018, 11, 3975–3991. [Google Scholar] [CrossRef]
- Perumalla, A.V.S.; Hettiarachchy, N.S. Green tea and grape seed extracts—Potential applications in food safety and quality. Food Res. Int. 2011, 44, 827–839. [Google Scholar] [CrossRef]
- Molino, S.; Casanova, N.A.; Henares, J.A.R.; Miyakawa, M.E.F. Natural Tannin Wood Extracts as a Potential Food Ingredient in the Food Industry. J. Agric. Food Chem. 2020, 68, 2836–2848. [Google Scholar] [CrossRef]
- Mirali, M.; Purves, R.W.; Stonehouse, R.; Song, R.; Bett, K.; Vandenberg, A. Genetics and biochemistry of zero-tannin lentils. PLoS ONE 2016, 11. [Google Scholar] [CrossRef]
- Zanotto, S.; Khazaei, H.; Elessawy, F.M.; Vandenberg, A.; Purves, R.W. Do faba bean genotypes carrying different zero-tannin genes (zt1 and zt2) differ in phenolic profiles? J. Agric. Food Chem. 2020, 68, 7530–7540. [Google Scholar] [CrossRef]
- Yao, Y.; Yang, X.S.; Tian, J.; Liu, C.Y.; Cheng, X.Z.; Ren, G.X. Antioxidant and antidiabetic activities of black mung bean (Vigna radiata L.). J. Agric. Food Chem. 2013, 61, 8104–8109. [Google Scholar] [CrossRef]
- Mirali, M.; Purves, R.W.; Vandenberg, A. Profiling the phenolic compounds of the four major seed coat types and their relation to color genes in lentil. J. Nat. Prod. 2017, 80, 1310–1317. [Google Scholar] [CrossRef]
- Mirali, M.; Ambrose, S.J.; Wood, S.A.; Vandenberg, A.; Purves, R.W. Development of a fast extraction method and optimization of liquid chromatography–mass spectrometry for the analysis of phenolic compounds in lentil seed coats. J. Chromatogr. B 2014, 969, 149–161. [Google Scholar] [CrossRef]
- Shahidi, F.; Yeo, J. Insoluble-Bound Phenolics in Food. Molecules 2016, 21, 1216. [Google Scholar] [CrossRef]
- Carter, P. Spectrophotometric determination of serum iron at submicrogram level with a new reagent (Ferrozine). Anal. Biochem. 1971, 40, 450–458. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT—Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Csepregi, K.; Neugart, S.; Schreiner, M.; Hideg, É. Comparative evaluation of total antioxidant capacities of plant polyphenols. Molecules 2016, 21, 208. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, M.; Chintalwar, G.J.; Chattopadhyay, S. Antioxidant and radioprotective properties of an Ocimum sanctum polysaccharide. Redox Rep. 2005, 10, 257–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farvin, K.H.S.; Jacobsen, C. Phenolic compounds and antioxidant activities of selected species of seaweeds from Danish coast. Food Chem. 2013, 138, 1670–1681. [Google Scholar] [CrossRef] [PubMed]
- Poudel, A.; Gachumi, G.; Wasan, K.M.; Bashi, Z.D.; El-Aneed, A.; Badea, I. Development and Characterization of Liposomal Formulations Containing Phytosterols Extracted from Canola Oil Deodorizer Distillate along with Tocopherols as Food Additives. Pharmaceutics 2019, 11, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AAT Bioquest, I. Quest Graph™ IC50 Calculator. Available online: https://www.aatbio.com/tools/ic50-calculator (accessed on 1 May 2020).
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin-Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DMAC | TBARS | F-C | DPPH | FRAP | Ferrozine | |
|---|---|---|---|---|---|---|
| Assays | ||||||
| DMAC P | 1 | 0.973 * | 0.865 * | 0.986 * | 0.983 * | 0.930 * |
| TBARS A | 1 | 0.831 * | 0.979 * | 0.982 * | 0.943 * | |
| F-C A | 1 | 0.849 * | 0.832 * | 0.846 * | ||
| DPPH A | 1 | 0.995 * | 0.954 * | |||
| FRAP A | 1 | 0.966 * | ||||
| Ferrozine C | 1 | |||||
| Polyphenol classes | ||||||
| Anthocyanins | 0.370 * | 0.448 * | 0.234 | 0.377 * | 0.406 * | 0.433 * |
| Dihydroflavonols | 0.427 * | 0.496 * | 0.353 * | 0.434 * | 0.469 * | 0.541 * |
| Flavones | 0.521 * | 0.537 * | 0.402 * | 0.499 * | 0.550 * | 0.526 * |
| Flavonols | −0.082 | −0.104 | −0.025 | −0.115 | −0.122 | −0.157 |
| Flavan-3-ols | 0.771 * | 0.722 * | 0.560 * | 0.744 * | 0.716 * | 0.593 * |
| Hydroxybenzoic acids | 0.294 * | 0.314 * | 0.217 | 0.335 * | 0.350 * | 0.428 * |
| Hydroxycinnamic acids | 0.337 * | 0.299 * | 0.413 * | 0.264 * | 0.260 * | 0.269 * |
| Procyanidins | 0.742 * | 0.648 * | 0.599 * | 0.668 * | 0.643 * | 0.468 * |
| Name | Formula | Molecular Weight | RT [min] | Mass Error (ppm) | Identification Level |
|---|---|---|---|---|---|
| Yellow Bean (B3) | |||||
| Phenolic acid derivative | C11 H12 O6 | 240.06358 | 8.54 | 0.82 | 3 |
| Phenolic acid derivative | C14 H24 O5 | 272.16243 | 10.59 | 0.21 | 3 |
| Phenolic acid derivative | C14 H24 O5 | 272.1625 | 9.89 | 0.46 | 3 |
| Afzelechin | C15 H14 O5 | 274.0845 | 10.29 | 1.38 | 2 |
| Epiafzelechin | C15 H14 O5 | 274.08451 | 11.15 | 1.40 | 2 |
| Phenolic acid deoxyhexoside | C15 H16 O10 | 356.07484 | 9.66 | 1.39 | 3 |
| (Epi)afzelechin hexoside | C21 H24 O10 | 436.13738 | 7.70 | 1.00 | 2/3 |
| Kaempferol 3-O-glucoside (X, Z) | C21 H20 O11 | 448.10084 | 14.39 | 0.63 | 1 |
| Quercetin 3-O-rhamnoside | C21 H20 O11 | 448.1013 | 14.715 | 1.65 | 1 |
| 3,5-Dihydroxy-2-(4-hydroxyphenyl)-4-oxo-3,4-dihydro-2H-chromen-7-yl hexopyranoside | C21 H22 O11 | 450.11658 | 9.89 | 0.83 | 2/3 |
| Myricetin 3-O-rhamnoside | C21 H20 O12 | 464.09603 | 13.51 | 1.19 | 1 |
| Kaempferol acetyl hexoside | C23 H22 O12 | 490.11134 | 16.39 | 0.43 | 2/3 |
| Kaempferol malonyl hexoside (X) | C24 H22 O14 | 534.10118 | 16.40 | 0.41 | 2/3 |
| AA | C30 H26 O10 | 546.15358 | 11.42 | 1.80 | 2/3 |
| AA | C30 H26 O10 | 546.1536 | 10.35 | 1.84 | 2/3 |
| Kaempferol 3-O-sambioside (X) | C26 H28 O15 | 580.1433 | 13.41 | 0.82 | 2 |
| Kaempferol dihexoside | C27 H30 O16 | 610.15424 | 12.47 | 1.39 | 2/3 |
| AAC | C45 H38 O16 | 834.2176 | 10.94 | 1.93 | 2/3 |
| Brown bean (B4) | |||||
| (+)-Catechin | C15 H14 O6 | 290.07923 | 8.85 | 0.67 | 1 |
| (Epi)catechin hexoside | C21 H24 O11 | 452.13217 | 6.59 | 0.68 | 2/3 |
| AC | C30 H26 O11 | 562.148 | 9.14 | 0.87 | 2/3 |
| Procyanidin B1 | C30 H26 O12 | 578.14284 | 8.05 | 0.72 | 1 |
| CC (Z) | C30 H26 O12 | 578.14315 | 11.61 | 1.25 | 2/3 |
| Procyanidin B2 | C30 H26 O12 | 578.14329 | 9.41 | 1.50 | 1 |
| ACC | C45 H38 O17 | 850.21255 | 10.25 | 1.94 | 2/3 |
| ACC | C45 H38 O17 | 850.21255 | 9.91 | 1.94 | 2/3 |
| CCC | C45 H38 O18 | 866.207 | 6.03 | 1.37 | 2/3 |
| CCC | C45 H38 O18 | 866.20703 | 9.25 | 1.40 | 2/3 |
| CCC | C45 H38 O18 | 866.20715 | 9.73 | 1.54 | 2/3 |
| ACCC | C60 H50 O23 | 1138.27613 | 10.61 | 1.62 | 2/3 |
| CCCC | C60 H50 O24 | 1154.27068 | 9.84 | 1.28 | 2/3 |
| CCCC | C60 H50 O24 | 1154.2707 | 9.97 | 1.30 | 2/3 |
| CCCC | C60 H50 O24 | 1154.27127 | 11.69 | 1.80 | 2/3 |
| ACCCC | C75 H62 O29 | 1426.3409 | 11.45 | 2.26 | 2/3 |
| CCCCC | C75 H62 O30 | 1442.33558 | 10.99 | 2.07 | 2/3 |
| CCCCC | C75 H62 O30 | 1442.33568 | 10.45 | 2.14 | 2/3 |
| CCCCCC | C90 H74 O36 | 1730.39828 | 11.68 | 1.33 | 2/3 |
| CCCCCCC | C105 H86 O42 | 2018.46508 | 12.00 | 2.83 | 2/3 |
| Black bean (B2) | |||||
| (−)-Gallocatechin | C15 H14 O7 | 306.07417 | 6.89 | 0.71 | 1 |
| Vanillic acid 4- β -d-glucoside | C14 H18 O9 | 330.0953 | 5.632 | 0.66 | 1 |
| Delphinidin 3- β -d-Glucoside (myrtillin) (Y) | C21 H20 O12 | 464.0966 | 6.47 | 2.43 | 1 |
| (Epi)gallocatechin hexoside | C21 H24 O12 | 468.12707 | 4.85 | 0.62 | 2/3 |
| Myricetin 3-O-glucoside | C21 H20 O13 | 480.09095 | 12.36 | 1.17 | 2 |
| Quercetin hexoside derivative | C24 H26 O13 | 522.13791 | 9.48 | 1.10 | 2/3 |
| Quercetin hexoside derivative | C24 H26 O13 | 522.13792 | 10.21 | 1.11 | 2/3 |
| GC | C30 H26 O13 | 594.13794 | 6.68 | 1.00 | 2/3 |
| GG | C30 H26 O14 | 610.13304 | 4.82 | 1.28 | 2/3 |
| Phenolic acid dihexoside derivative | C28 H34 O18 | 658.17597 | 6.47 | 2.22 | 3 |
| Phenolic acid dihexoside derivative | C29 H36 O18 | 672.19118 | 7.75 | 1.51 | 3 |
| Soyasaponin A1 (X) | C59 H96 O29 | 1268.60579 | 16.05 | 1.62 | 2 |
| GGCCC | C75 H62 O32 | 1474.32534 | 9.09 | 1.98 | 2/3 |
| GGGGC | C75 H62 O34 | 1506.31495 | 7.968 | 1.79 | 2/3 |
| Lentil (L2, L3 and L4) | |||||
| Pantothenic acid | C9 H17 N O5 | 219.1107 | 3.04 | 0.13 | 2 |
| Thymidine | C10 H14 N2 O5 | 242.0905 | 3.44 | 0.96 | 2 |
| Hydroxybenzoic acid hexoside | C13 H16 O8 | 300.08462 | 6.40 | 0.36 | 2/3 |
| Hydroxybenzoic acid hexoside | C13 H16 O8 | 300.08472 | 3.94 | 0.68 | 2/3 |
| Hydroxybenzoic acid derivative | C16 H20 O11 | 388.10104 | 9.22 | 1.23 | 3 |
| 5-O-[β-apiosyl-(1-2)-O-β-xylopyranosyl]gentisic acid | C17 H22 O12 | 418.11142 | 10.98 | 0.71 | 2 |
| Dihydroxybenzoic acid pentoside hexoside | C18 H24 O13 | 448.12215 | 9.43 | 1.03 | 2/3 |
| Phenolic acid derivative | C21 H32 O13 | 492.18471 | 12.43 | 0.84 | 3 |
| Procyanidin B3 | C30 H26 O12 | 578.14298 | 8.71 | 0.95 | 1 |
| CC | C30 H26 O12 | 578.14298 | 10.40 | 0.96 | 2/3 |
| GC | C30 H26 O13 | 594.13772 | 8.67 | 0.64 | 2/3 |
| GC (Z) | C30 H26 O13 | 594.13794 | 7.53 | 1.00 | 2/3 |
| GG (Z) | C30 H26 O14 | 610.13321 | 6.48 | 1.57 | 2/3 |
| CCC | C45 H38 O18 | 866.20698 | 10.87 | 1.35 | 2/3 |
| GCC (Z) | C45 H38 O19 | 882.20148 | 9.54 | 0.86 | 2/3 |
| GCC | C45 H38 O19 | 882.20187 | 8.24 | 1.29 | 2/3 |
| GGC | C45 H38 O20 | 898.19641 | 7.34 | 0.86 | 2/3 |
| GGC (Z) | C45 H38 O20 | 898.19674 | 9.98 | 1.22 | 2/3 |
| GGG | C45 H38 O21 | 914.19169 | 6.63 | 1.24 | 2/3 |
| GCCC | C60 H50 O25 | 1170.26587 | 9.38 | 1.50 | 2/3 |
| GGCC | C60 H50 O26 | 1186.261455 | 8.48 | 2.04 | 2/3 |
| GGCC | C60 H50 O26 | 1186.26057 | 9.66 | 1.30 | 2/3 |
| GGGC | C60 H50 O27 | 1202.25548 | 7.85 | 1.27 | 2/3 |
| GCCCC | C75 H62 O31 | 1458.33039 | 10.23 | 1.98 | 2/3 |
| GGCCC | C75 H62 O32 | 1474.32422 | 10.53 | 1.22 | 2/3 |
| GGCCC | C75 H62 O32 | 1474.32441 | 9.35 | 1.35 | 2/3 |
| GGGCC | C75 H62 O33 | 1490.31944 | 8.78 | 1.42 | 2/3 |
| GGGCC | C75 H62 O33 | 1490.31961 | 9.02 | 1.53 | 2/3 |
| GGGGC | C75 H62 O34 | 1506.31474 | 8.12 | 1.65 | 2/3 |
| GGGCCC | C90 H74 O39 | 1778.38314 | 9.71 | 1.36 | 2/3 |
| GGGGCC | C90 H74 O40 | 1794.37975 | 9.19 | 2.29 | 2/3 |
| GGGGGC | C90 H74 O41 | 1810.37349 | 8.47 | 1.62 | 2/3 |
| GGGCCCC | C105 H86 O45 | 2066.4502 | 10.55 | 2.95 | 2/3 |
| GGGGCCC | C105 H86 O46 | 2082.44459 | 9.92 | 2.67 | 2/3 |
| Green lentil (L3) | |||||
| Quercetin pentoside | C17 H22 O13 | 434.10639 | 14.31 | 0.80 | 2/3 |
| Catechin 3-O-glucoside (Z) | C21 H24 O11 | 452.13219 | 8.02 | 0.73 | 2 |
| Kaempferol di-rutinoside | C39 H50 O24 | 902.27023 | 10.11 | 1.14 | 2/3 |
| Brown lentil (L4) | |||||
| Phenolic acid derivative | C13 H20 O4 | 240.13636 | 12.97 | 0.85 | 3 |
| Trihydroxy-megastigmadien-one hexoside | C19 H30 O9 | 402.18923 | 11.57 | 0.61 | 2/3 |
| Trihydroxy-megastigmadien-one hexoside (Z) | C19 H30 O9 | 402.18928 | 11.47 | 0.75 | 2/3 |
| Trihydroxy-megastigma-en-one hexoside | C19 H32 O9 | 404.20496 | 10.20 | 0.80 | 2/3 |
| Black lentil (L2) | |||||
| Quercetin deoxyhexoside | C21 H20 O11 | 448.10092 | 16.75 | 0.79 | 2/3 |
| Luteolin 4′-O-glucoside | C21 H20 O11 | 448.10104 | 16.14 | 1.07 | 1 |
| Tricetin hexoside | C21 H20 O12 | 464.09605 | 16.75 | 1.24 | 2/3 |
| Tricetin hexoside | C21 H20 O12 | 464.09606 | 15.16 | 1.26 | 2/3 |
| Phenolic acid derivative | C20 H28 O14 | 492.14852 | 7.91 | 1.26 | 3 |
| Delphinidin 3-O-(2-O-β-d-Glucopyranosyl-α-l-arabinopyranoside) (Y) | C26 H28 O16 | 596.13879 | 6.68 | 1.77 | 2 |
| GCC | C45 H38 O19 | 882.2021 | 8.36 | 1.55 | 2/3 |
| GGC | C45 H38 O20 | 898.19622 | 7.43 | 0.64 | 2/3 |
| GGCC | C60 H50 O26 | 1186.26075 | 8.56 | 1.45 | 2/3 |
| GGGGC | C75 H62 O34 | 1506.31576 | 8.30 | 2.33 | 2/3 |
| Pea (P2 and P4) | |||||
| 3,4-Dihydroxybenzoic acid | C7 H6 O4 | 154.02662 | 5.59 | 0.07 | 1 |
| Aspartic acid derivative | C9 H15 N O5 | 217.09494 | 5.95 | −0.36 | 3 |
| N-phenylacetyl-aspartic acid | C12 H13 N O5 | 251.07947 | 9.26 | 0.39 | 2 |
| N-salicyloyl-aspartic acid | C11 H11 N O6 | 253.05873 | 7.28 | 0.35 | 2 |
| Aspartic acid derivative | C11 H17 N O7 | 275.10071 | 6.55 | 0.76 | 3 |
| N-coumaroyl-aspartic acid isomer | C13 H13 N O6 | 279.07431 | 10.45 | 0.08 | 2/3 |
| N-coumaroyl-aspartic acid isomer | C13 H13 N O6 | 279.07438 | 8.80 | 0.32 | 2/3 |
| N-[(2,4-Dihydroxyphenyl)acetyl]-l-aspartic acid | C12 H13 N O7 | 283.0693 | 8.31 | 0.34 | 2 |
| Amino acid derivative | C13 H15 N O7 | 297.08486 | 7.21 | 0.03 | 3 |
| N-feruloyl-aspartic acid isomer | C14 H15 N O7 | 309.0849 | 9.73 | 0.17 | 2/3 |
| N-feruloyl-aspartic acid isomer | C14 H15 N O7 | 309.08498 | 11.31 | 0.41 | 2/3 |
| N-(2,4,6-Trimethoxybenzyl)-l-aspartic acid | C14 H19 N O7 | 313.11643 | 11.49 | 0.88 | 2 |
| Aspartic acid derivative | C14 H21 N O7 | 315.13213 | 10.91 | 1.05 | 3 |
| Aspartic acid derivative | C13 H21 N O9 | 335.12185 | 5.48 | 0.66 | 3 |
| Gibberellic acid or isomer | C19 H22 O6 | 346.14182 | 10.99 | 0.53 | 2/3 |
| Unidentified | 404.13211 | 3.81 | 4 | ||
| Phenolic acid derivative | C18 H28 O10 | 404.16868 | 8.59 | 1.07 | 3 |
| Unidentified | 406.1843 | 8.75 | 4 | ||
| Amino acid derivative | C17 H21 N O11 | 415.11196 | 8.96 | 1.21 | 3 |
| Aspartic acid derivative | C20 H24 N2 O11 | 468.13798 | 6.78 | −0.06 | 3 |
| Aspartic acid derivative | C20 H24 N2 O11 | 468.13798 | 5.64 | −0.05 | 3 |
| N-[[3-(β -d-Glucopyranosyloxy)-2,3-dihydro-2-oxo-1H-indol-3-yl]acetyl]aspartic acid | C20 H24 N2 O12 | 484.13328 | 4.19 | 0.74 | 2 |
| Chlorinated aspartic acid derivative | C21 H27 Cl N2 O6 S2 | 502.09938 | 8.65 | −1.06 | 3 |
| Chlorinated aspartic acid derivative (Z) | C21 H27 Cl N2 O6 S2 | 502.09938 | 7.45 | −1.05 | 3 |
| Chlorinated aspartic acid derivative | C21 H27 Cl N2 O7 S2 | 518.09414 | 5.23 | −1.31 | 3 |
| Unidentified | 589.27425 | 11.76 | 4 | ||
| GG | C30 H26 O14 | 610.13302 | 6.17 | 1.25 | 2/3 |
| GG (Z) | C30 H26 O14 | 610.13303 | 8.24 | 1.27 | 2/3 |
| GGG | C45 H38 O21 | 914.1916 | 7.72 | 1.14 | 2/3 |
| GGG | C45 H38 O21 | 914.19184 | 4.37 | 1.40 | 2/3 |
| GGG | C45 H38 O21 | 914.19192 | 7.82 | 1.49 | 2/3 |
| GGGG | C60 H50 O28 | 1218.25088 | 9.58 | 1.66 | 2/3 |
| GGGGG | C75 H62 O35 | 1522.3086 | 10.05 | 0.95 | 2/3 |
| GGGGGG | C90 H74 O42 | 1826.36757 | 8.90 | 1.15 | 2/3 |
| GGGGGGG | C105 H86 O49 | 2130.43022 | 9.17 | 3.03 | 2/3 |
| Chickpea (C2, C3 and C4) | |||||
| Dihydroxybenzoic acid | C7 H6 O4 | 154.02667 | 10.99 | 0.39 | 3 |
| Gallic acid | C7 H6 O5 | 170.02166 | 3.74 | 0.80 | 1 |
| Glutamyl phenylalanine | C14 H18 N2 O5 | 294.12176 | 6.12 | 0.64 | 2 |
| Glutamyl tyrosine | C14 H18 N2 O6 | 310.11664 | 4.61 | 0.48 | 2 |
| Dihydroxybenzoic acid hexoside | C13 H16 O9 | 316.07957 | 8.48 | 0.43 | 2/3 |
| Dihydroxybenzoic acid hexoside | C13 H16 O9 | 316.07964 | 8.97 | 0.65 | 2/3 |
| Trihydroxybenzoic acid hexoside | C13 H16 O10 | 332.07459 | 13.38 | 0.72 | 2/3 |
| 11-hydroxy-9,10-dihydrojasmonic acid 11-β-d-glucoside | C18 H30 O9 | 390.18933 | 7.14 | 0.88 | 2 |
| Unidentified | 396.16328 | 6.18 | 4 | ||
| Hydroxybenzoic acid hexoside pentoside | C18 H24 O12 | 432.12679 | 5.74 | 0.04 | 2/3 |
| Kaempferol 3-O-rutinoside (Nicotiflorin) | C27 H30 O15 | 594.15915 | 13.95 | 1.15 | 1 |
| Quercetin 3-O-rutinoside (Rutin) (X) | C27 H30 O16 | 610.15399 | 13.03 | 1.00 | 1 |
| Myricetin hexoside deoxyhexoside | C27 H30 O17 | 626.14864 | 12.19 | 0.55 | 2/3 |
| Myricetin hexoside deoxyhexoside | C27 H30 O17 | 626.14869 | 12.11 | 0.63 | 2/3 |
| Kaempferol pentoside-hexoside-deoxyhexoside | C32 H38 O19 | 726.20143 | 12.95 | 0.96 | 2/3 |
| Quercetin pentoside-hexoside-deoxyhexoside | C32 H38 O20 | 742.19611 | 12.15 | 0.62 | 2/3 |
| Quercetin pentoside-hexoside-deoxyhexoside | C32 H38 O20 | 742.19625 | 12.24 | 0.81 | 2/3 |
| GG-deoxyhexoside | C33 H40 O20 | 756.21216 | 13.14 | 1.15 | 2/3 |
| Myricetin pentoside-hexoside-deoxyhexoside | C32 H38 O21 | 758.19094 | 11.32 | 0.50 | 2/3 |
| Myricetin pentoside-hexoside-deoxyhexoside | C32 H38 O21 | 758.19098 | 11.55 | 0.55 | 2/3 |
| Myricetin hexoside dideoxyhexoside | C33 H40 O21 | 772.20664 | 12.29 | 0.56 | 2/3 |
| Myricetin derivative | C34 H42 O22 | 802.21764 | 11.20 | 1.08 | 3 |
| GGGG | C60 H50 O28 | 1218.2508 | 7.33 | 1.60 | 2/3 |
| GGGGG | C75 H62 O35 | 1522.30948 | 7.66 | 1.52 | 2/3 |
| Faba bean (F2, F3 and F4) | |||||
| Phenolic acid derivative | C11 H12 O5 | 224.06867 | 10.99 | 0.89 | 3 |
| Hydroxyjasmonic acid | C12 H18 O4 | 226.12081 | 8.65 | 1.35 | 2 |
| Prolyl aspartic acid | C9 H14 N2 O5 | 230.09031 | 5.48 | 0.16 | 2 |
| Phenolic acid derivative | C12 H16 O5 | 240.10005 | 9.19 | 1.14 | 3 |
| Phenolic acid derivative | C12 H18 O5 | 242.1157 | 5.86 | 1.15 | 3 |
| Phenolic acid derivative | C12 H18 O6 | 258.11044 | 5.42 | 0.39 | 3 |
| Phenolic acid derivative | C12 H18 O6 | 258.11047 | 6.24 | 0.51 | 3 |
| Phenolic acid derivative | C12 H14 O7 | 270.07392 | 9.62 | −0.14 | 3 |
| (-)-Epicatechin | C15 H14 O6 | 290.07922 | 10.04 | 0.64 | 1 |
| (-)-Epigallocatechin | C15 H14 O7 | 306.0742 | 8.49 | 0.80 | 1 |
| Aspartic acid derivative | C14 H14 N2 O7 | 322.08015 | 6.47 | 0.16 | 3 |
| Aspartic acid derivative (Z) | C14 H14 N2 O7 | 322.08016 | 4.75 | 0.18 | 3 |
| Phenolic acid derivative | C15 H18 N O9 P | 387.07262 | 12.08 | 1.81 | 3 |
| Phenolic acid derivative | C18 H28 O9 | 388.17373 | 7.18 | 1.04 | 3 |
| Phenolic acid derivative | C18 H28 O10 | 404.1687 | 11.13 | 1.11 | 3 |
| Unidentified | 427.22128 | 13.11 | 4 | ||
| Caffeic acid malonyl hexoside (X) | C16 H28 O13 | 428.15318 | 2.24 | 0.44 | 2/3 |
| (Epi)catechin hexoside | C21 H24 O11 | 452.13245 | 9.34 | 1.31 | 2/3 |
| (Epi)gallocatechin hexoside | C21 H24 O12 | 468.12722 | 7.88 | 0.95 | 2/3 |
| N-[[3-(β -d-Glucopyranosyloxy)-2,3-dihydro-2-oxo-1H-indol-3-yl]acetyl]aspartic acid | C20 H24 N2 O12 | 484.13314 | 4.03 | 0.45 | 2 |
| CC (Z) | C30 H26 O12 | 578.14271 | 9.83 | 0.50 | 2/3 |
| GC | C30 H26 O13 | 594.13794 | 7.22 | 1.01 | 2/3 |
| GC | C30 H26 O13 | 594.13798 | 6.32 | 1.08 | 2/3 |
| GC | C30 H26 O13 | 594.13803 | 9.01 | 1.16 | 2/3 |
| Unidentified | 594.15926 | 10.25 | 4 | ||
| GCC | C45 H38 O19 | 882.20213 | 5.17 | 1.59 | 2/3 |
| GGC | C45 H38 O20 | 898.19605 | 4.78 | 0.45 | 2/3 |
| Pulse Crop | Sample Code | Seed Coat Genotype | Seed Coat Color | High/Low Tannin | Seed Pictures |
|---|---|---|---|---|---|
| Chickpea (Cicer arietinum L.) | C1 | CDC Xena | White | Low tannin |  |
| C2 | CDC Ebony | Black | High tannin |  | |
| C3 | CDC Jade | Green | High tannin |  | |
| C4 | CDC Cory | Brown | High tannin |  | |
| Faba bean (Vicia faba L.) | F1 | CDC Snowdrop | White | Low tannin |  |
| F2 | Black Fava | Black | High tannin |  | |
| F3 | Masterpiece | Green | High tannin |  | |
| F4 | 749-13-2015 | Beige | High tannin |  | |
| Lentil (Lens culinaris Medik.) | L1 | 6500ZT-4 | Grey | Low tannin |  |
| L2 | Indianhead | Black | High tannin |  | |
| L3 | CDC Kermit | Green | High tannin |  | |
| L4 | CDC Robin | Brown | High tannin |  | |
| Pea (Pisum sativum L.) | P1 | CDC Meadow | White | Low tannin |  |
| P2 | 30/2-ILT | Maple (patterned) | High tannin |  | |
| P3 | MFR042 | Green | Low tannin |  | |
| P4 | CDC Dakota | Dun (brown) | High tannin |  | |
| Common bean (Phaseolus vulgaris L.) | B1 | Envoy | White | Low tannin |  |
| B2 | CDC Jet | Black | High tannin |  | |
| B3 | CDC Sol | Yellow | High tannin |  | |
| B4 | CDC WM-2 | Brown (pinto) | High tannin |  |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elessawy, F.M.; Vandenberg, A.; El-Aneed, A.; Purves, R.W. An Untargeted Metabolomics Approach for Correlating Pulse Crop Seed Coat Polyphenol Profiles with Antioxidant Capacity and Iron Chelation Ability. Molecules 2021, 26, 3833. https://doi.org/10.3390/molecules26133833
Elessawy FM, Vandenberg A, El-Aneed A, Purves RW. An Untargeted Metabolomics Approach for Correlating Pulse Crop Seed Coat Polyphenol Profiles with Antioxidant Capacity and Iron Chelation Ability. Molecules. 2021; 26(13):3833. https://doi.org/10.3390/molecules26133833
Chicago/Turabian StyleElessawy, Fatma M., Albert Vandenberg, Anas El-Aneed, and Randy W. Purves. 2021. "An Untargeted Metabolomics Approach for Correlating Pulse Crop Seed Coat Polyphenol Profiles with Antioxidant Capacity and Iron Chelation Ability" Molecules 26, no. 13: 3833. https://doi.org/10.3390/molecules26133833
APA StyleElessawy, F. M., Vandenberg, A., El-Aneed, A., & Purves, R. W. (2021). An Untargeted Metabolomics Approach for Correlating Pulse Crop Seed Coat Polyphenol Profiles with Antioxidant Capacity and Iron Chelation Ability. Molecules, 26(13), 3833. https://doi.org/10.3390/molecules26133833