
Flavonoids as Promising Antiviral Agents against SARS-CoV-2 Infection: A Mechanistic Review


Abstract
:1. Introduction
2. Results
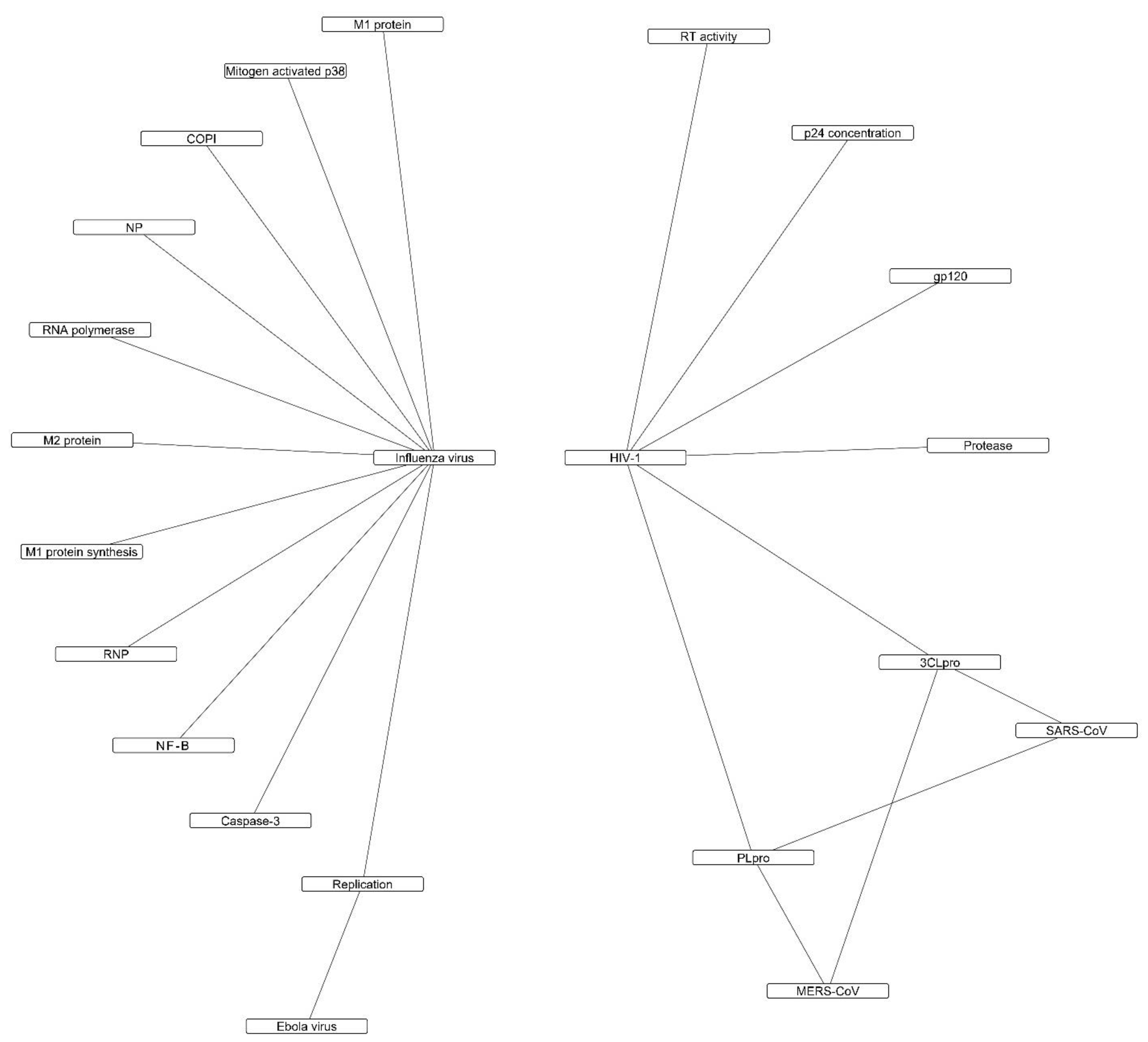
2.1. Direct Antiviral Mechanisms
2.1.1. Inhibition of Viral Protease
2.1.2. Inhibition of Viral RNA Polymerase and Viral mRNA
2.1.3. Viral Entry, Replication, and Infectivity
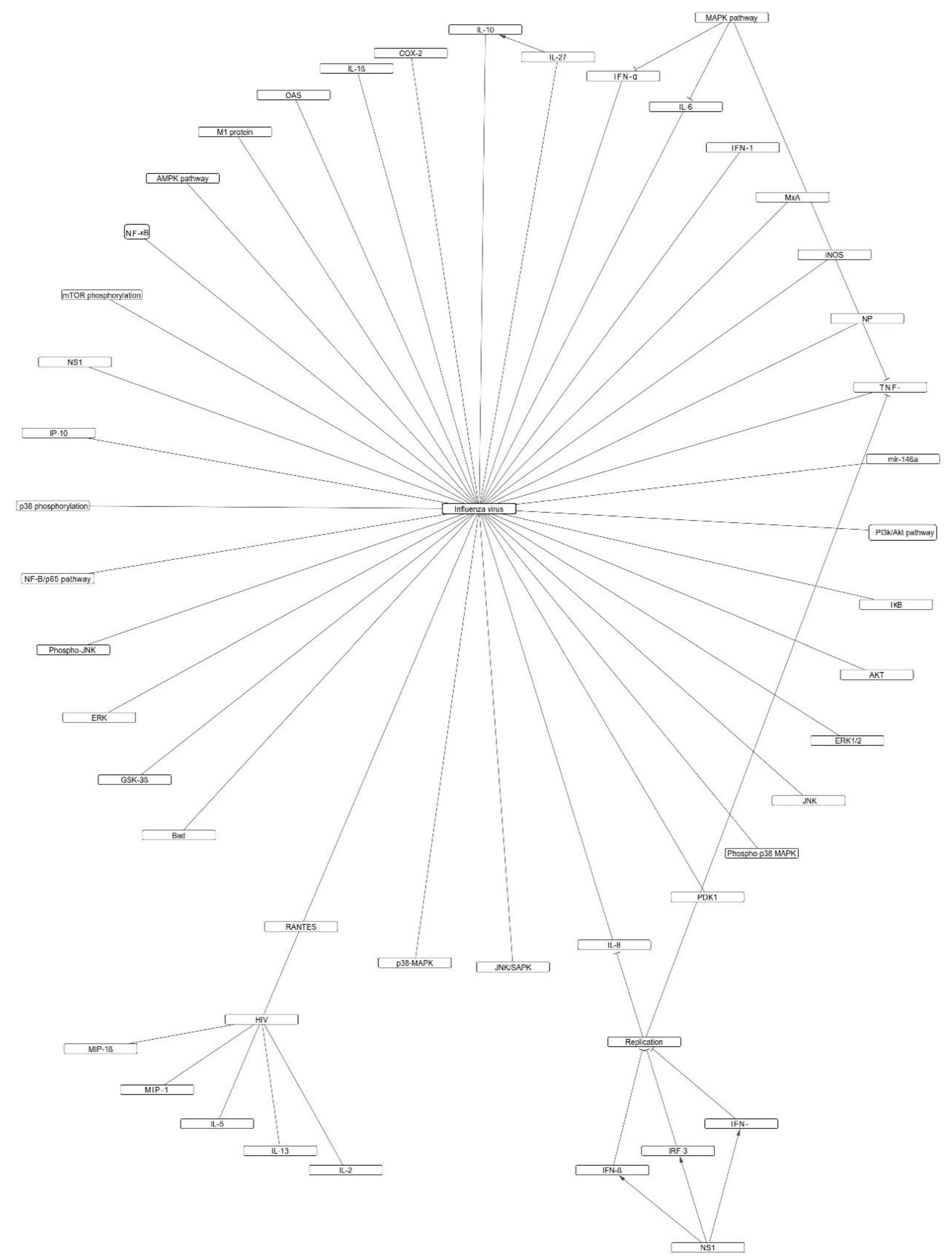
2.2. Indirect Antiviral Effects
2.2.1. Effect on Interferons
2.2.2. Effect on Pro-Inflammatory Cytokines (TNF, IL, and MCP)
2.2.3. Effect on Sub-Cellular Inflammatory Pathways (NF-kB, PI3K/Akt, and MAPK/JNK)
3. Discussion
4. Materials and Methods

{kind=link}
{kind=link}
| Phytochemical Name | Plant Source | Model | Dose/Concentration | Mechanisms/Outcomes | Reference |
|---|---|---|---|---|---|
| (−)-epigallocatechin 3-O-gallate | Limonium morisianum Arrigoni | Anti-HIV-1 RT and IN | - | ↓ HIV-1 RT-associated RNase | [128] |
| H activity: IC50 = 0.21 μM | |||||
| ↓ IN catalytic function and IN-LEDGF-dependent activity: IC50 = 1.92 μM | |||||
| 2′,4′-dihydroxy-6′-methoxy-3′,5′-dimethylchalcone | Cleistocalyx operculatus (Roxb.) Merr. and L.M.Perry | HEK293 cells infected with the plasmid H1N1 or oseltamivir-resistant novel H1N1 (H274Y) | 20–40 μM | ↑ cell viability, | [67] |
| MDCK cells infected with influenza H1N1 A/PR/8/34 and H9N2 A/Chicken/Korea/O1310/2001 | ↓ NA activity (IC50 = 5.07 to 8.84 μM), viral replication, and CPE | ||||
| 3-Deoxysappanchalcone | Caesalpinia sappan L. | MDCK, A549, and THP-1 cells infected with influenza virus A/PR/8/34 (H1N1) | 30 µM | ↓ viral genomic replication, DNA fragmentation, CCL5, CXCL10, IL-6, IL-1β, caspase 3/7, 8, and 9 activity and HA copy number (IC50 = 3.9 µM) | [68] |
| 3-deoxysappanchalcone, Sappanchalcone and rhamnetin | Caesalpinia sappan L. | MDCK cells infected with influenza A/Guangdong/243/72 (H3N2), A/PR/8/34 (H1N1) and B/Jiangsu/10/2003 | - | ↓ NA activity: IC50 = 13.9–24.1 μg/mL ↓ CPE: IC50 = 1.06–15.4 μg/mL CC50 = 12.83–115.47 μg/mL SI = 6.23–16.27 | [129] |
| 5,7,4′-Trihydroxy-8-methoxyflavone | - | MDCK cells infected with influenza A/PR/8/34 (H1N1), A/Guizhou/54/89 (H3N2), and B/Ibaraki/2/85 | - | ↓ Sialidase activity: | [130] |
| IC50 = 6.58–9.78 µg/mL | |||||
| 6-hydroxyluteolin 7-O-β-d-glucoside | Salvia plebeia R.Br. | MDCK cells infected with influenza A/PR/8/34 (H1N1) | 20, 50 μM | ↓ NA activity and CPE, ↑ cell viability | [131] |
| 8-Prenylkaempferol | Sophora flavescens Aiton | A549 cells infected with influenza A/PR/8/34 (H1N1) virus | 1–30 μM | ↓ RANTES production, NF-κB, IRF-3, PI3K activity, Akt phosphorylation, and IκB degradation | [111] |
| Agathisflavone | Anacardium occidentale L. | Mice infected with wild-type and oseltamivir-resistant influenza virus | IC50 = 20 to 2.0 µM, EC50 = 1.3 µM | [132] | |
| ↓ NA activity and virus replication | |||||
| Apigenin 7-O-β-d-(4′-caffeoyl)glucuronide | Chrysanthemum morifolium Ramat. | MT-4 cells infected with HIV-IIIIB | - | ↓ HIV-1 integrase activity: | [133] |
| IC50 = 7.2 µg/mL | |||||
| CPE: | |||||
| EC50 = 41.86µg/mL, | |||||
| SI ≥ 3.58 | |||||
| Baicalein | - | A549 cells infected with influenza H5N1 virus strains (A/Thailand/1(Kan-1)/04 and A/Vietnam/1203/04) | 40–100 µM | ↓ viral nucleoprotein: IC50 = 18.79 µM | [53] |
| CC50 = 109.41 µM | |||||
| SI = 5.82 | |||||
| Primary human monocyte-derived macrophages (MDM) infected with influenza A/Thailand/1(Kan-1)/04 | ↓ virus titer, caspase-3 activation, NA activity, | ||||
| ↓ IL-6 and IL-8 | |||||
| ↓ viral replication, IL6, CXCL10, and TNF-α | |||||
| Baicalin | - | Human lung epithelial A549 cells infected with influenza A/Jingfang/01/1986 | 20 μg/mL | ↓ viral NP, M1 protein levels, viral titer, miR-146a expression, virus replication and viral copy number (EC50 = 17.04, 19.31 μg/mL), ↑ TRAF6 level, IFN-α, and IFN-β | [54] |
| (H1N1) and A/Lufang/09/1993 (H3N2) | ↓miR-146a expression and virus copy number | ||||
| Balb/C mice inoculated intranasally with the influenza A H1N1 virus | ↑ survival rate, IFN-α and IFN-β | ||||
| Baicalin | Scutellaria baicalensis Georgi | A549 and Ana-1 cells infected with influenza virus A3/Beijing/30/95 (H3N2) | 12.5–50 μg/mL | ↑ mTOR phosphorylation, ↓ autophagy, Atg5–Atg12 complex and LC3-II expression, EC50 = 15–15.6 µg/mL | [112] |
| Baicalin | - | A549 and MDCK cells infected with influenza virus A/H1N1/Eastern India/66/pdm09 (H1N1-pdm09) | 0.5–320 µM | TD50 = 220 µM | [55] |
| IC50 = 0.5 and 18 µM | |||||
| ↓ NP transcription, RIG-1, PKR, NS1 expression, viral replication, TNF-α, IL-8, p-85b–NS1 binding, p-Akt, M1 protein, ↑ IRF-3, IFN-γ, and IFN-β | |||||
| BALB/c mice infected intranasally with H1N1-pdm09 | (10–120 mg/kg/day) twice daily for 3 days | ↓ viral titer: | |||
| MIC50 ≈ 80 mg/kg/day | |||||
| ↓ p-Akt and M1 protein expression | |||||
| Baicalin | Scutellaria baicalensis Georgi | MDCK and A549 cells infected with influenza A/FM1/1/47 (H1N1) and A/Beijing/32/92 (H3N2) | 20–80 µg/mL (in MDCK cells) | ↑ cell viability, ↓ virus replication, and CPE: | [71] |
| EC50 = 40.3 and 104.9 µg/mL | |||||
| 5–40 µg/mL (in A549 cells) | SI: 2.1–8.6 | ||||
| ICR mice infected with influenza A/FM1/1/47 (H1N1) virus | ↓ NA activity: | ||||
| 50–200 mg/kg/d for 5 days, i.v injection | IC50 = 52.3 and 85.8µg/mL | ||||
| ↓ death rate, weight loss, ↑ mean day to death, survival rates, and improved the lung parameters | |||||
| Baicalin | Scutellaria baicalensis Georgi | Hos/CD4/CCR5 or Hos/CD4/CXCR4 cells infected with recombinant vaccinia virus vTF7-3 | 0.04 to 400 µM | ↓ X4 and R5 HIV-1 Env-mediated fusion, CAT activity, CD4/CXCR4, CD4/CCR5, and HIV-1 entry | [134] |
| Baicalein, Baicalin | - | Vero CCL-81 cell line infected with SARS-CoV-2 | 20 µM | ↓ viral RdRp and viral replication | [49] |
| CC50 = 86–100 µM | |||||
| EC50 = 1–9 µM | |||||
| Baicalein | - | Vero E6 cells infected with SARS-CoV-2 | 0.1 μM | ↓ body weight loss, the replication of the virus, relieved the lesions of lung tissue, inflammatory cell infiltration, IL-1β and TNF-α | [66] |
| hACE2 transgenic mice infected with SARS-CoV-2 | 200 mg/kg | ↑ respiratory function | |||
| Biochanin A | - | A549 cells infected with influenza H5N1 virus strains (A/Thailand/1(Kan-1)/04 and A/Vietnam/1203/04) | 40 µM | ↓ viral nucleoprotein: IC50 = 8.92 µM | [53] |
| CC50 = 49.91 µM | |||||
| SI = 5.60 | |||||
| Primary human monocyte-derived macrophages (MDM) infected with influenza A/Thailand/1(Kan-1)/04 | ↓ virus titer, caspase-3 activation, NFκB p65 accumulation, IL-6, IL-8, CXCL10 production, phosphorylation of AKT and ERK 1/2 and ↑ IκB levels | ||||
| ↓ IL6, CXCL10, and TNF-α | |||||
| Catechin | - | A549 cells infected with influenza A H1N1 | 5–50 μM | ↓ NA and HA activity, viral load, and virus-induced autophagy | [135] |
| Catechins | Camellia sinensis (L.) Kuntze | MDCK cell infected with influenza A/Chile/1/83 (H1N1), A/Sydney/5/97 (H3N2), and B/Yamagata/16/88 | 30–1200 µM | ↓ plaque formation: | [136] |
| EC50 = 22.2–318 µM | |||||
| ↓ NA activity, HA activity, and viral RNA synthesis | |||||
| Catechins (EGCG, ECG and C5G) | Camellia sinensis (L.) Kuntze | MDCK cells infected with influenza A/Victoria/503/2013, | 50–100 µM | ↓ NA activity | [137] |
| IC50 = 100.3–173 µM | |||||
| A/SouthAustralia/21/2013 and A/Perth/25/2013 | CC50 = 274–551.3 µM | ||||
| ↓ plaque number | |||||
| EC50 = 28.4–34.3 µM | |||||
| Catechins with a galloyl moiety | - | HIV-1 integrase assay kit | 0.1–100 μM | ↓ HIV-1 integrase activity IC50 = 0.56–3.02 μM | [138] |
| Cirsimaritin | - | MDCK and THP-1 cells infected with influenza A/Fort Monmouth/1/1947(H1N1), A/tianjinjinnan/15/2009(H1N1) and A/JiangXi/312/2006(H3N2) | 2.5–20 μg/mL | ↓ CPE: | [59] |
| IC50 = 5.8, 6.3, 11.1 μg/mL, SI = 24.3, 26.4, 13.8, and TC50 = 153.3 | |||||
| ↓ viral replication, M2 viral protein expression, intracellular | |||||
| p65/NF-κB protein, p65/NF-κB phosphorylation, TNF-α, IL-1β, | |||||
| IL-8, IL-10, COX-2 expression, phospho-p38 MAPK, and ↓ phospho-JNK | |||||
| C-methylated flavonoids | Cleistocalyx operculatus (Roxb.) Merr. and L.M.Perry | MDCK and 293T cells infected with influenza A/PR/8/34 (H1N1), A/Chicken/Korea/O1310/2001 (H9N2), A/Sw/Kor/CAH1/04 (H1N1, KCTC 11165BP), novel H1N1 (WT) and Oseltamivir-resistant novel H1N1 (H274Y) | - | ↓ CPE: | [139] |
| EC50 = 4.90–8.79 µM | |||||
| SI = 10.15 to > 24.49 | |||||
| ↓ NA activity: | |||||
| IC50 = 2.55–93.77 µM | |||||
| EGCG | Camellia sinensis (L.) Kuntze (green tea) | MDCK cells infected with influenza A/Puerto Rico/8/34 (H1N1) (PR8), A/Hong Kong/8/68 (H3N2) (HK), A/Brisbane/59/2007 (H1N1) (BB), A/Taiwan/1/1986 (H1N1) (TW), A/Korea/01/2009 (H1N1) (KR) and B/Panama/45/1990 (PNM) | 10–100 µM | ↓ CPE: | [140] |
| EC50 = 8.9–17.3 µM | |||||
| ↓ hemifusion events, viral membrane integrity, cell penetration capacity, NP protein, viral entry, and NA activity: IC50 = 133.2 to > 500 µM | |||||
| EGCG | - | PBMCs, CD4+ T cells and macrophages infected with several clinical isolates of HIV-1 | 6–100 μM | ↓ HIV-1 p24 antigen: IC50 = 4.5–12 μM | [141] |
| ↓ HIV-1 infectivity, | |||||
| HIV-1–glycoprotein 120 attachment to the CD4 molecule, | |||||
| CC50 > 100 μM | |||||
| EGCG | - | CD4+ T cells | 0.2–20 μM | ↓ HIV-1 gp 120 binding to the CD4+ T cells | [142] |
| EGCG | Camelia sinensis (L.) Kuntze | Peripheral blood CD4+ T cells (by flow cytometry) | 25–100 µM | ↓ CD4 expression, anti-CD4 antibody binding to its antigen, gp120 binding to CD4, and HIV infection | [143] |
| EGCG | Camelia sinensis (L.) Kuntze | Peripheral blood lymphocytes infected with either LAI/IIIB or Bal HIV strains | 1–50 µM | ↓ virus replication, RT activity, and p24 | [75] |
| EGCG | Camelia sinensis (L.) Kuntze | THP-1 and H9 cells infected with HIV-1 | 1–100 µM | ↓ RT activity, protease activity, p24, viral entry, and viral production | [40] |
| Flavonoid aglycones (demethoxymatteucinol, matteucinol, matteucin, methoxymatteucin, and 3′-hydroxy-5′-methoxy-6,8-dimethylhuazhongilexone) | Pentarhizidium orientale (Hook.) Hayata | MDCK cells infected with influenza A/PR/8/34 (H1N1) or A/chicken/Korea/01210/2001 (H9N2) | - | ↓ NA activities and CPE: | [144] |
| IC50 = 23.1–31.3 μM, EC50 = 21.4–30.7 μM, CC50 = 77.6 μM (demethoxymatteucinol), CC50 => 100 μM (matteucin, methoxymatteucin, and 3′-hydroxy-5′-methoxy-6,8-dimethylhuazhongilexone) | |||||
| Flavonoid compounds | - | SARS-CoV proteases (recombinant 3CLpro) expressed in Pichia pastoris GS115 | 200 µM | ↓ 3CLpro activity: | [145] |
| IC50 = 47–381 µM | |||||
| Gallocatechin-7-gallate | Pithecellobium clypearia Benth | A549 and MDCK cells infected with influenza A/PR/8/34 (H1N1) | 3–30 μM | ↓ CPE: | [58] |
| CC50 = above 100 μM, EC50 = 1.69 μM | |||||
| ↓ NP and M2 expression levels, HA mRNA expression, M2/M1 levels, and phosphorylation of SF2/ASF and SC35 | |||||
| ICR mice infected intranasally with influenza A H1N1 virus | 30 mg/kg/d, i.v, for 5 days | ↓ viral NP mRNA expression, TNF-α, IL-1β, IL-6, bodyweight loss, acute lung injury and lung virus titer, ↑ survival rate, T-lymphocyte stimulation index, B-lymphocyte stimulation index, and spleen and thymus indices | |||
| Genistein | - | Heterologously expression of viral protein U of HIV in Xenopus oocyte | 20 µM | ↓ Ba2+-sensitive current and blocked Vpu ion channels | [146] |
| Genistein | - | Primary human macrophages infected with HIV-1Ba-L Env expressed on 293 T cells | 5–10 µg/mL | ↓ R5 Env pseudotyped virus infection, HIV-1Ba-L Env expressing cells and macrophages cell-fusion, reporter gene expression, virus penetration, gp120-induced TNF-α secretion, virus replication | [76] |
| Ginkgetin | Ginkgo biloba L. and Cephalotaxus harringtonia K. Koch | MDCK cells infected with influenza A/PR/8/34 (H1N1), A/Guizhou/54/89 (H3N2), and B/Ibaraki/2/85 | - | ↓ sialidase activity: | [130] |
| IC50 = 9.78 to > 100 µg/mL | |||||
| Herbacitrin | Drosera peltata Thunb. | HIV-1 infected MT-4 and MT-2 cell culture | 21.5 µM | ↓ HIV-1 replication, HIV-1 RT activity, IN activity, and p24 level | [77] |
| Hesperidin | - | R5-type HIV-1 in CD4+ NKT cells and human Vδ1+ cells in PBMCs | 30–100 µg/mL | ↑ IL-2, IL-5, IL-13, MIP-1α, MIP-1β, RANTES, CFSE, and CD25 expression, and ↓ viral replication | [104] |
| Hesperidin | - | Influenza A virus (H1N1) induced lung injury in male Sprague-Dawley rats, by the intrathecal route | 200 and 500 mg/kg/d, i.p., for 5 days | ↑ pulmonary function, | [94] |
| ↓ Local numbers of immune cells, TNF-α, IL-6, and IFN-α | |||||
| H1N1 infected pulmonary microvascular endothelial cells | 1 mg/mL | ↓ TNF-α, IL-6, IFN-α, phosphorylated p38 and JNK | |||
| Hexamethoxyflavone (5-Hydroxy-3,6,7,8,3′,4′-hexamethoxyflavone) | Marcetia taxifolia (A. St.-Hil.) DC. | MT4 cells infected with HIV-1 (HTLV-IIIB/H9) | 45 µM | ↓ HIV-1 RT activity: IC50 = 4.1 µM, | [147] |
| EC50 = 0.04 μM, CC50 > 50 μM | |||||
| Hispidulin | Salvia plebeia R. Br. | MDCK cells infected with influenza strain H1N1 A/PR/8/34 virus | 20–50 μM | ↓ CPE and NA activity: | [148] |
| IC50 = 19.83 μM, EC50 = 22.62 μM, SI > 8.90 | |||||
| ↑ cell survival rate recovered the chromosome condensation | |||||
| Homoplantaginin | Salvia plebeia R.Br. | MDCK cells infected with influenza A/PR/8/34 (H1N1) | 20, 50 μM | ↓ NA activity and CPE, ↑ cell viability | [131] |
| IND02 | Cinnamomum zeylanicum Blume | MAGI cells and PBMCs infected with HIV-1 LAI and NL4-3 | 5–30 μM | ↓ gp120 binding to HS (IC50 = 7 μM), gp120 binding to CD4 (IC50 = 20 μM), and envelope binding to CD4 | [149] |
| IND02-trimer | Cinnamomum zeylanicum Blume | MAGI cells and PBMCs infected with HIV-1 LAI, NL4-3, Ba-L and clinical isolates (HIV-1 92UG029(A-X4), HIV-1 92HT599 (B-X4), HIV-1 96USHIPS4 (B-X4/R5) and HIV-1 98IN017 (C-X4)) | 2–20 μM | ↓ gp120 binding to HS (IC50 = 7.5 μM), EC50 = 0.8–7 μM, CC50 = 96 and 23 | [149] |
| HIV-1-infected CD4+ and CD8+ T cells | 0.46–46.3 μM | ↓ up-modulation of Tim-3 and PD-1 | |||
| Isoliquiritigenin | - | SARS-CoV proteases (3CLpro and PLpro) expressed in E. coli BL21 | - | ↓ PLpro activity: IC50 = 24.6 µM, Deubiquitination activity = 17.2, DeISGylation activity = 12.6, | [37] |
| ↓ 3CLpro activity: | |||||
| MERS-CoV proteases (3CLpro and PLpro) expressed in E. coli BL21 | IC50 = 61.9 µM | ||||
| ↓ PLpro activity: IC50 = 82.2 µM, | |||||
| ↓ 3CLpro activity: IC50 = 33.9 µM | |||||
| Isoquercetin | - | MDCK or Vero cells infected with influenza A viruses from pigs (A/swine/OH/511445/2007 [H1N1], Oh7) and human (A/PR/8/34 [H1N1], PR8), and human influenza B virus (B/Lee/40) | 1–5 µM | ↓ viral replication and CPE: | [79] |
| ED50 = 1.2 µM | |||||
| 2–10 mg/kg/day, i.p. | TD50 = 45 µM | ||||
| BALB/c mice infected with influenza A/PR/8/34, H1N1virus | SI = 38 | ||||
| ↓ IFN-γ, iNOS, RANTES, virus titers, viral bronchitis, and bronchiolitis | |||||
| Isorhamnetin | - | MDCK cells infected with influenza virus A/PR/08/34 (H1N1)Embryonated chicken eggs infected with influenza virus A/PR/08/34 (H1N1) | 50 µM | ↑ cell viability | [113] |
| EC50 = 23 µM | |||||
| CC50 > 280 µM | |||||
| SI > 12 | |||||
| C57BL/6 mice infected with influenza A/PR/8/34 (H1N1) | 1 mg/kg/day for 5 days (intranasal route) | ↓ autophagy, ROS generation, ERK phosphorylation, cytoplasmic lysosome acidification, NA and HA expression, and NA activity | |||
| ↓ virus titer, adsorption onto RBCs and RBCs hemolysis | |||||
| ↓ lung virus titer and body weight loss, | |||||
| ↑ survival rate | |||||
| Quercetin | Elaeagnus rhamnoides (L.) A.Nelson (synonym: Hippophae rhamnoides L.) | ACE2h cells infected with SARS-CoV-2 | 50 µM | ↓ Viral entry | [65] |
| Isorhamnetin | ↓ Viral Binding affinity to ACE2 | ||||
| Kaempferol | - | SARS-CoV proteases (3CLpro and PLpro) expressed in E. coli BL21 | - | ↓ PLpro activity: IC50 = 16.3 µM, Deubiquitination activity = 61.7, DeISGylation activity = 71.7, | [37] |
| ↓ 3CLpro activity: | |||||
| MERS-CoV proteases (3CLpro and PLpro) expressed in E. coli BL21 | IC50 = 116.3 µM | ||||
| ↓ PLpro activity: IC50 = 206.6 µM, | |||||
| ↓ 3CLpro activity: IC50 = 35.3 µM | |||||
| Kaempferol derivatives | - | Heterologously expression of 3a protein of SARS-CoV in Xenopus oocyte | 10–20 µM | ↓ Ba2+-sensitive current and 3a-mediated current, blocked 3a-protein channel IC50 = 2.3 µM (for juglanin) | [36] |
| Linarin | - | R5-type HIV-1 in CD4+ NKT cells and human Vδ1+ cells in PBMCs | 10–100 µg/mL | ↑ IL-2, IL-5, IL-13, MIP-1α, MIP-1β, RANTES, CFSE, and CD25 expression, ↓ viral replication | [104] |
| Luteolin | - | MDCK, Calu-3, and Vero cells infected with influenza A/Jiangxi/312/2006 (H3N2) and A/Fort Monmouth/1/1947 (H1N1) | 3.75–240 μM | ↓ CPE: | [60] |
| IC50 = 6.89, 7.15 μM, CC50 = 148–240 μM | |||||
| ↓ M2 viral protein expression, virus absorption, and internalization | |||||
| Luteolin | Salvia plebeia R. Br. | MDCK cells infected with influenza strain H1N1 A/PR/8/34 virus | 50 μM | ↓ NA activity: IC50 = 17.96 μM, EC50 = cytotoxic | [148] |
| Luteolin | - | PBMCs, TZM-bl reporter, and Jurkat cellsinfected with wild–type HIV (NLENY1) or VSV-HIV-1 | 5–10 µM | ↓ clade-B- and -C -Tat-driven LTR transactivation, reactivation of latent HIV-1 infection, HIV-1 gene expression, LTR activity, PBMC cell aggregation/syncytia, viral entry | [150] |
| Luteolin and luteolin 7-methyl ether | Coleus parvifolius Benth. | MT-4 cells infected with HIV-1 (HTLV IIIB) | - | ↓ HIV-1 integrase activity: | [151] |
| IC50 = 11–70 µM | |||||
| ↓ viral replication | |||||
| Myricetin | - | TZM-bl cell infected with HIV-1 BaL (R5 tropic), H9 and PBMC cells infected with HIV-1 MN (X4 tropic), and the dual tropic (X4R5) HIV-1 89.6, | 0.01–100 μM | ↓ p24 antigen: | [152] |
| IC50 = 1.76–22.91 μM, CC50 = 804.94–1214.72 μM, | |||||
| Anti-HIV-1 RT | ↓ HIV-1 RT: IC50 = 203.65 μM | ||||
| Myricetin | - | TZM-bl cell infected with HIV-1 BaL (R5 tropic), H9 and PBMC cells infected with HIV-1 MN (X4 tropic), and the dual tropic (X4R5) HIV-1 89.6, | 0.01–100 µM | ↑ cell viability, | [152] |
| ↓ p24 antigen: | |||||
| IC50 = 1.76–22.91 μM, CC50 = 804.94–1214.72 μM, | |||||
| ↓ HIV-1 RT: | |||||
| Anti-HIV-1 RT | IC50 = 203.65 μM | ||||
| Myricetin-3-O-(6″-O-galloyl)-β-D-galactopyranoside | Limonium morisianum Arrigoni | Anti-HIV-1 RT and IN | - | ↓ HIV-1 RT-associated RNase | [128] |
| H activity: IC50 = 10.9 μM | |||||
| ↓ IN catalytic function and IN-LEDGF-dependent activity: IC50 = 6.47 μM | |||||
| Myricetin derivatives | Marcetia taxifolia (A. St.-Hil.) DC. | MT4 cells infected with HIV-1 (HTLV-IIIB/H9) | - | ↓ HIV-1 RT activity: IC50 = 7.6–13.8 µM, EC50 = 45–230 μM, SI > 1.3–7 | [153] |
| Myricetin-3′,5′-dimethylether 3-O-β-D-galactopyranoside | Cleistocalyx operculatus (Roxb.) Merr. and L.M.Perry | HEK293 cells infected with the plasmid H1N1 or oseltamivir-resistant novel H1N1 (H274Y) | 40 μM | ↑ cell viability, | [67] |
| MDCK cells infected with influenza H1N1 A/PR/8/34 and H9N2 A/Chicken/Korea/O1310/2001 | ↓ NA activity (IC50 = 6.50 to 9.34 μM), viral replication and CPE | ||||
| Naringenin | - | Vero E6 cells infected with HCoVOC43, HCoV229E, and SARS-CoV-2 | 62.5, 250 μM | ↓ TPC2, CPE activity | [74] |
| Nepetin | Salvia plebeia R. Br. | MDCK cells infected with influenza strain H1N1 A/PR/8/34 virus | 20–50 μM | ↓ CPE and NA activity: | [148] |
| IC50 = 11.18 μM, EC50 = 17.45 μM, SI = ~11.47, | |||||
| ↑ cell survival rate | |||||
| Nepitrin | Salvia plebeia R.Br. | MDCK cells infected with influenza A/PR/8/34 (H1N1) | 20, 50 μM | ↓ NA activity and CPE, ↑ cell viability | [131] |
| Oroxylin A | - | MDCK and A549 cells infected with influenza A/FM/1/47 (H1N1), A/Beijing/32/92 (H3N2), and oseltamivir-resistant | 40–50 μM | ↓ CPE: | [52] |
| IC50 = 270.9, 245.0, 241.4 μM, EC50 = 44.6, 36.1, 109.4 μM | |||||
| A/FM/1/47-H275Y (H1N1-H275Y) viruses H1N1-H275Y and A/Anhui/1/2013-R294 K (H7N9-R294 K) | 100 μM | ↓ viral mRNA and M1 protein expression | |||
| ↓ NA activity, IC50 = 241.4 and 203.6 μM | |||||
| ICR mice infected intranasally with the A/FM/1/47 (H1N1) | 100 mg/kg/d, p.o. | ↑ IFN-β, IFN-γ and survival rate ↓ body weight loss, lung injury, lung indexes and lung scores | |||
| Oroxylin A | Scutellaria baicalensis Georgi | CHME5 cells and primary human macrophages infected with HIV-1-D3 | 5–20 µM | ↓ phosphorylation of PI3K, PDK1, Akt, activation of GSK3β, m-TOR, and Bad | [114] |
| Pentamethoxyflavone(5,3′-dihydroxy-3,6,7,8,4′-pentamethoxyflavone) | Marcetia taxifolia (A. St.-Hil.) DC. | MT4 cells infected with HIV-1 (HTLV-IIIB/H9) | 45 µM | ↓ HIV-1 RT activity: IC50 = 0.4 µM, EC50 = 0.05 μM, CC50 > 50 μM | [154] |
| Pongamones A–E | Pongamia pinnata (L.) Pierre | In vitro inhibitory activity against HIV-1 RT | - | ↓ RT activity | [155] |
| IC50 > 10 µg/mL | |||||
| Prenylisoflavonoids | Erythrina senegalensis DC. | In vitro inhibitory activity against recombinant HIV-1 protease | - | ↓ HIV-1 protease activity | [44] |
| IC50 = 0.5–30.1 µM | |||||
| Purified chalcones | Angelica keiskei (Miq.) Koidz. | SARS-CoV proteases (3CLpro and PLpro) expressed in E. coli BL21 | - | ↓ 3CLpro activity: | [156] |
| Cell-free cleavage: IC50 = 11.4–129.8 µM, | |||||
| Cell-based cleavage: IC50 = 5.8–50.8 µM, SI = 0.4–9.2 | |||||
| ↓ PLpro activity: IC50 = 1.2–46.4 µM, Deubiquitination activity = 2.6–44.1, DeISGylation activity = 1.1–11.3 | |||||
| Purified flavanone glucosides | Thevetia peruviana SCHUM. | HIV-1 IN protein expressed in E. coli, | ↓ HIV-1 RDDP activity: IC50 = 20–43 µM | [157] | |
| RDDP and DDDP inhibitory activity assay | ↓ HIV-1 DDDP activity: IC50 = 42 and 69 µM | ||||
| ↓ HIV-1 IN activity: IC50 = 5–45 µM | |||||
| Purified flavones | Kaempferia parviflora Wall. ex Baker | In vitro inhibitory activity against HIV-1 protease | - | ↓ HIV-1 protease | [158] |
| IC50 = 19.04–160.07 µM | |||||
| Purified flavonoids | Pithecellobium clypearia Benth | A549 cells infected with influenza A/PR/8/34 (H1N1), A/Sydney/5/97 (H3N2) and B/Jiangsu/10/2003 | 3–30 μg/mL | ↓ NA activity: | [105] |
| IC50 = 29.77–39.15 µg/mL | |||||
| ↓ IL-6 and MCP-1 | |||||
| Purified flavonoids | - | NAs from influenza A/PR/8/34 (H1N1), A/Jinan/15/90 (H3N2), andB/Jiangshu/10/2003 | - | ↓ NA activity: | [159] |
| IC50 = 22–87.6 μM | |||||
| ↓ CPE: | |||||
| MDCK cells infected with influenza A/Jinan/15/90 (H3N2) | IC50 = 4.74–24.70 µM | ||||
| SI = 1.82–9.64 | |||||
| Purified flavonoids | Elsholtzia rugulosa Hemsl. | NAs from influenza viruses A/PR/8/34(H1N1), A/Jinan/15/90(H3N2) and B/Jiangsu/10/2003 | - | ↓ NA activity: | [160] |
| IC50 = 7.81–28.49 μM | |||||
| ↓ viral replication and CPE: | |||||
| MDCK cells infected with influenza A/Jinan/15/90 (H3N2) | IC50 = 1.43 to > 500 µM, | ||||
| SI = 1.73–7.48 | |||||
| Purified flavonoids (Ochnaflavone 7″-O-methyl ether and 2″,3″dihydroochnaflavone 7″ methyl ether) | Ochna integerrima (Lour.) Merr. | 1A2 cell line infected with ∆Tat/revMC99 virus | 200 µg/mL | ↓ RT activity: | [161] |
| IC50 = 2.0 and 2.4 µg/mL | |||||
| ↓ HIV-1 activities: | |||||
| EC50 = 2 and 0.9 µg/mL | |||||
| IC50 = 6.3 and 2.9 µg/mL | |||||
| SI = 3.1 and 3.2 | |||||
| Purified flavonol glycosides | Zanthoxylum piperitum (L.) | MDCK cells infected with influenza A/NWS/33 (H1N1) | 7.8–1000 μg/mL | ↓ NA activity: | [162] |
| IC50 = 211–434 μg/mL | |||||
| ↓ PFU | |||||
| Purified flavonols | Rhodiola rosea L. | MDCK cells infected with influenza A/PR/8/34 (H1N1) and A/Chicken/Korea/MS96/96 (H9N2) | - | ↓ CPE: | [163] |
| EC50 = 6.25–145.4 µM | |||||
| SI = 1.6 to > 48 | |||||
| Recombinant influenza A virus H1N1(rvH1N1) | ↓ NA activity: | ||||
| IC50 = 2.2–56.9 μg/mL | |||||
| Purified flavonoids | Broussonetia papyrifera (L.) L’Hér. ex Vent. | SARS-CoV proteases (3CLpro and PLpro) expressed in E. coli BL21 | - | ↓ PLpro activity: IC50 = 3.7–66.2 µM, Deubiquitination activity = 7.6–74.8, DeISGylation activity = 8.5–70.8, | [37] |
| ↓ 3CLpro activity: | |||||
| IC50 = 30.2–233.3 µM | |||||
| MERS-CoV proteases (3CLpro and PLpro) expressed in E. coli BL21 | ↓ PLpro activity: IC50 = 39.5–171.6 µM, | ||||
| ↓ 3CLpro activity: IC50 = 27.9–193.7 µM | |||||
| Quercetin | - | SARS-CoV proteases (3CLpro and PLpro) expressed in E. coli BL21 | - | ↓ PLpro activity: IC50 = 8.6 µM, Deubiquitination activity = 20.7, DeISGylation activity = 34.4, | [37] |
| ↓ 3CLpro activity: | |||||
| MERS-CoV proteases (3CLpro and PLpro) expressed in E. coli BL21 | IC50 = 52.7 µM | ||||
| ↓ 3CLpro activity: IC50 = 34.8 µM | |||||
| Kaempferol | Vero E6 cells infected with SARS-CoV | 125, 62.5, and 31.25 μM | ↓ Virus-induced cell death, 3CLprotease | [35] | |
| Quercetin | - | MDCK and A549 cells infected with influenza A/Puerto Rico/8/34 (H1N1), A/FM-1/47/1 (H1N1) and A/Aichi/2/68 (H3N2) | 12.5–100 µg/mL (50 µg/mL) | ↓ CPE: | [62] |
| IC50 = 2.738–7.756 µg/mL, IC90 = 8.24–24.58 µg/mL, | |||||
| ↓ HA mRNA transcription, viral NP protein synthesis, viral HA expression and virus infection rate, target the membrane fusion process during virus entry | |||||
| Quercetin | - | Inhibitory activity against recombinant HIV-1 protease | - | ↓ HIV-1 protease activity: | [45] |
| IC50 = 58.8 µM | |||||
| Quercetin 3-O-(6″-feruloyl)-β-D-galactopyranoside | Polygonum viscosum Buch.-Ham. ex D. Don | In vitro anti-HIV-1 activity | - | ↓ RT activity | [164] |
| IC50 = 25.61 µg/mL | |||||
| Quercetin 3-rhamnoside | Houttuynia cordata Thunb. | MDCK cells infected with influenza A/WS/33 | 10–100 µg/mL | ↓ CPE, viral mRNA synthesis, virus replication and virus infection | [80] |
| Quercetin 3-β-O-D-glucoside | - | Vero E6 epithelial | 10 μM | ↓ virus replication (EC50 = 5.3 µM, EC90 = 9.3 µM), viral titers and entry of Ebola viruses | [81] |
| cells infected with EBOV-Kikwit-GFP, EBOV-Makona | |||||
| and SUDV or VSV-EBOV, and VSV-SUDV | 50 mg/kg every other day, i.p. | ↓ virus replication and body weight loss, | |||
| BALB/c or C57BL/6 mice infected with mouse-adapted Ebola virus | ↑ survival rate | ||||
| Quercetin-3-O-α-L-rhamnopyranoside | Rapanea melanophloeos (L.) Mez | MDCK cells infected with A/Puerto Rico/8/1934 (H1N1) | 150 μg/mL | ↓ CPE: | [106] |
| CC50 = 200 μg/mL, EC50 = 25 μg/mL, EC90 = 100 μg/mL | |||||
| ↓ NP and M2 genes copy numbers, viral titer, HA titer and TNF-α, | |||||
| ↑ IL-27 protein level and cell viability | |||||
| Quercetin-7-O-glucoside | Dianthus superbus var. longicalycinus (Maxim.) F.N.Williams | MDCK cells infected with influenza A/Vic/3/75 (H3N2), A/PR/8/34 (H1N1), B/Maryland/1/59 and B/Lee/40 viruses | 10 μg/mL | ↓ CPE: | [50] |
| IC50 = 3.10 μg/mL to 8.19 μg/mL, CC50 > 100 μg/mL, SI = 12.21 to 32.25, | |||||
| ↓ ROS, autophagy, viral RNA synthesis and viral RNA polymerase | |||||
| Quercetin-β-galactoside | - | SARS-CoV proteases (3CLpro and PLpro) expressed in E. coli BL21 | - | ↓ PLpro activity: IC50 = 51.9 µM, Deubiquitination activity = 136.9, DeISGylation activity = 67.7, | [37] |
| ↓ 3CLpro activity: | |||||
| IC50 = 128.8 µM | |||||
| MERS-CoV proteases (3CLpro and PLpro) expressed in E. coli BL21 | ↓ PLpro activity: IC50 = 129.4 µM, | ||||
| ↓ 3CLpro activity: IC50 = 68.0 µM | |||||
| Santin | Artemisia rupestris L. | MDCK and THP-1 cells infected with influenza strain A/Fort Monmouth/1/1947 (H1N1) and A/Wuhan/359/1995 | 60 µM | ↓ CPE: | [61] |
| IC50 = 27.68, 37.20 μM, SI = 14.45, 10.75, TC50 > 400 μM | |||||
| ↓ M2 viral protein expression, phosphorylation of p38 MAPK, | |||||
| (H3N2) | JNK/SAPK, ERK, NF-κB, TNF-α, IL-1β, IL-6, IL-8, and IL-10 production | ||||
| Scutellarin | Erigeron breviscapus (Vant.) Hand.-Mazz | C8166 cells infected with HIV-1IIIB and HIV-1IIIB/H9, | 54–541 µM | ↓ HIV-1 replication: | [165] |
| MT-2 cells infected with HIV-174V, | EC50 = 15–253 µM | ||||
| PBMC cells infected with HIV-1KM018 | CC50 = 336 -> 1082 µM | ||||
| Purified recombinant HIV-1 RT | ↓ RT activity, HIV-1 particle attachment and fusion | ||||
| Tectorigenin | Pueraria thunbergiana (Siebold and Zucc.) Benth. | CHME5 cells and primary human macrophages infected with HIV-1-D3 | 5–20 µM | ↓ phosphorylation of PI3K, PDK1, Akt, activation of GSK3β, m-TOR, and Bad | [114] |
| Theaflavins | Camellia sinensis (L.) Kuntze (black tea) | MDCK andA549 cells infected with influenza A/PR/8/34(A/H1N1), A/Sydney/5/97(A/H3N2) and B/Jiangsu/10/2003 | 0.1 to 30 µg/mL | ↓ NA: | [108] |
| IC50 = 10.67–49.6 µM | |||||
| ↓ CPE: | |||||
| CC50 = 76.7–177.1 µM | |||||
| ↓ HA activity, IL-6 and vRNP nuclear localization, | |||||
| Theaflavins | - | SARS-CoV proteases (3CLpro) expressed in E. coli | - | ↓ 3CLpro activity | [43] |
| IC50 = 3–9.5 µM | |||||
| Tricin | - | MDCK cells infected with influenza A/Solomon islands/3/2006 (H1N1), A/Hiroshima/52/2005 (H3N2), A/California/07/2009 (H1N1pdm), A/Narita/1/2009 (H1N1pdm) and B/Malaysia/2506/2004 | 3.3–30 µM | ↓ HA and matrix protein, mRNA expression, virus titer (EC50 = 3.4–10.2 µM) | [63] |
| DBA/2 Cr mice infected intranasally with influenza A/PR/8/34 virus | 20–100 µg/kg, p.o. | ↓ Body weight loss, | |||
| ↑ Survival rate | |||||
| Wogonin | Scutellaria baicalensis Georgi | MDCK and A549 cells infected with human influenza virus A/Puerto-Rico/8/34 (H1N1) PR8, seasonal H1N1, H3N2 and B (yamagata lineage) | 10 μg/mL | ↓ NA and NS1 levels, viral replication, Akt phosphorylation, ↑ IFN-β, IFN-λ1, MxA, OAS, AMPK phosphorylation, phospho-IRF-3 expression, cleaved PARP, and caspase-3 expression and apoptosis | [95] |
| ↓ Plaque formation: IC50 = 10 μg/mL |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, Y.F.; Bai, C.; He, F.; Xie, Y.; Zhou, H. Review on the potential action mechanisms of Chinese medicines in treating Coronavirus Disease 2019 (COVID-19). Pharmacol. Res. 2020, 158, 104939. [Google Scholar] [CrossRef]
- Samieefar, N.; Yari Boroujeni, R.; Jamee, M.; Lotfi, M.; Golabchi, M.R.; Afshar, A.; Miri, H.; Khazeei Tabari, M.A.; Darzi, P.; Abdullatif Khafaie, M.; et al. Country Quarantine During COVID-19: Critical or Not? Disaster Med. Public Health Prep. 2020, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Park, S.E. Epidemiology, virology, and clinical features of severe acute respiratory syndrome -coronavirus-2 (SARS-CoV-2; Coronavirus Disease-19). Clin. Exp. Pediatr. 2020, 63, 119–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singhal, T. A review of coronavirus disease-2019 (COVID-19). Indian J. Pediatr. 2020, 87, 281–286. [Google Scholar] [CrossRef] [Green Version]
- Novel, C.P.E.R.E. The epidemiological characteristics of an outbreak of 2019 novel coronavirus diseases (COVID-19) in China. Zhonghua Liu Xing Bing Xue Za Zhi 2020, 41, 145. [Google Scholar] [CrossRef]
- Bosch, B.J.; Van der Zee, R.; De Haan, C.A.; Rottier, P.J.M. The coronavirus spike protein is a class I virus fusion protein: Structural and functional characterization of the fusion core complex. J. Virol. 2003, 77, 8801–8811. [Google Scholar] [CrossRef] [Green Version]
- Li, H.Y.; Li, F.; Sun, H.Z.; Qian, Z.M. Membrane-inserted conformation of transmembrane domain 4 of divalent-metal transporter. Biochem. J. 2003, 372, 757–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Khazeei Tabari, M.A.; Khoshhal, H.; Tafazoli, A.; Khandan, M.; Bagheri, A. Applying computer simulations in battling with COVID-19, using pre-analyzed molecular and chemical data to face the pandemic. Inf. Med. Unlocked 2020, 21, 100458. [Google Scholar] [CrossRef]
- Chan, J.F.; Kok, K.H.; Zhu, Z.; Chu, H.; To, K.K.; Yuan, S.; Yuen, K.Y. Genomic characterization of the 2019 novel human-pathogenic coronavirus isolated from a patient with atypical pneumonia after visiting Wuhan. Emerg. Microbes Infect. 2020, 9, 221–236. [Google Scholar] [CrossRef] [Green Version]
- Grein, J.; Ohmagari, N.; Shin, D.; Diaz, G.; Asperges, E.; Castagna, A.; Feldt, T.; Green, G.; Green, M.L.; Lescure, F.X.; et al. Compassionate Use of Remdesivir for Patients with Severe Covid-19. N. Engl. J. Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Sissoko, D.; Laouenan, C.; Folkesson, E.; M’lebing, A.B.; Beavogui, A.H.; Baize, S.; Camara, A.M.; Maes, P.; Shepherd, S.; Danel, C. Experimental treatment with favipiravir for Ebola virus disease (the JIKI Trial): A historically controlled, single-arm proof-of-concept trial in Guinea. PLoS. Med. 2016, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuta, Y.; Komeno, T.; Nakamura, T. Favipiravir (T-705), a broad spectrum inhibitor of viral RNA polymerase. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2017, 93, 449–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Y.X.; Chen, X.P. Favipiravir: Pharmacokinetics and Concerns about Clinical Trials for 2019-nCoV Infection. Clin. Pharmacol. Ther. 2020, 108, 242–247. [Google Scholar] [CrossRef] [Green Version]
- Choy, K.T.; Wong, A.Y.; Kaewpreedee, P.; Sia, S.F.; Chen, D.; Hui, K.P.Y.; Chu, D.K.W.; Chan, M.C.W.; Cheung, P.P.; Huang, X.; et al. Remdesivir, lopinavir, emetine, and homoharringtonine inhibit SARS-CoV-2 replication in vitro. Antivir. Res. 2020, 178, 104786. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Wang, Y.; Wen, D.; Liu, W.; Wang, J.; Fan, G.; Ruan, L.; Song, B.; Cai, Y.; Wei, M.; et al. A Trial of Lopinavir-Ritonavir in Adults Hospitalized with Severe Covid-19. N. Engl. J. Med. 2020, 382, 1787–1799. [Google Scholar] [CrossRef]
- Yousefifard, M.; Zali, A.; Mohamed Ali, K.; Madani Neishaboori, A.; Zarghi, A.; Hosseini, M.; Safari, S. Antiviral therapy in management of COVID-19: A systematic review on current evidence. Arch. Acad. Emerg. Med. 2020, 8, e45. [Google Scholar]
- Bahramsoltani, R.; Sodagari, H.R.; Farzaei, M.H.; Abdolghaffari, A.H.; Gooshe, M.; Rezaei, N. The preventive and therapeutic potential of natural polyphenols on influenza. Expert. Rev. Anti Infect. Ther. 2016, 14, 57–80. [Google Scholar] [CrossRef]
- Bahramsoltani, R.; Rahimi, R. An Evaluation of Traditional Persian Medicine for the Management of SARS-CoV-2. Front. Pharmacol. 2020, 11, 571434. [Google Scholar] [CrossRef]
- Siahpoosh, M.B. How Can Persian Medicine (Traditional Iranian Medicine) Be Effective to Control COVID-19? Tradit. Integr. Med. 2020, 5, 46–48. [Google Scholar]
- Siahpoosh, M.B. Clinically Improvement in a Case of COVID-19 with Gastrointestinal Manifestations through Using Natural Therapy based on Persian Medicine: A Case Report. Tradit. Integr. Med. 2020, 5, 114–117. [Google Scholar]
- Zargaran, A.; Karimi, M.; Rezaeizadeh, H. COVID 19: Natural Products and Traditional Medicines; Opportunity or Threat? Tradit. Integr. Med. 2021, 6, 1–2. [Google Scholar]
- Zhang, Z.L.; Zhong, H.; Liu, Y.X.; Le, K.J.; Cui, M.; Yu, Y.T.; Gu, Z.C.; Gao, Y.; Lin, H.W. Current therapeutic options for coronavirus disease 2019 (COVID-19)-lessons learned from severe acute respiratory syndrome (SARS) and Middle East Respiratory Syndrome (MERS) therapy: A systematic review protocol. Ann. Transl. Med. 2020, 8, 1527. [Google Scholar] [CrossRef]
- Gurung, A.B.; Ali, M.A.; Lee, J.; Farah, M.A.; Al-Anazi, K.M. Unravelling lead antiviral phytochemicals for the inhibition of SARS-CoV-2 M(pro) enzyme through in silico approach. Life. Sci. 2020, 255, 117831. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Choudhir, G.; Shukla, S.K.; Sharma, M.; Tyagi, P.; Bhushan, A.; Rathore, M. Identification of phytochemical inhibitors against main protease of COVID-19 using molecular modeling approaches. J. Biomol. Struct. Dyn. 2020, 1–21. [Google Scholar] [CrossRef]
- Sadati, S.M.; Gheibi, N.; Ranjbar, S.; Hashemzadeh, M.S. Docking study of flavonoid derivatives as potent inhibitors of influenza H1N1 virus neuraminidase. Biomed. Rep. 2019, 10, 33–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmadian, R.; Rahimi, R.; Bahramsoltani, R. Kaempferol: An encouraging flavonoid for COVID-19. Bol. Latinoam. Caribe Plantas Med. Aromat. 2020, 19. [Google Scholar] [CrossRef]
- Suwannarach, N.; Kumla, J.; Sujarit, K.; Pattananandecha, T.; Saenjum, C.; Lumyong, S. Natural Bioactive Compounds from Fungi as Potential Candidates for Protease Inhibitors and Immunomodulators to Apply for Coronaviruses. Molecules 2020, 25, 1800. [Google Scholar] [CrossRef] [Green Version]
- Jäger, S.; Cimermancic, P.; Gulbahce, N.; Johnson, J.R.; McGovern, K.E.; Clarke, S.C.; Shales, M.; Mercenne, G.; Pache, L.; Li, K.; et al. Global landscape of HIV-human protein complexes. Nature 2011, 481, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V.; Hayashi, Y.; Jung, S.H. An Overview of Severe Acute Respiratory Syndrome-Coronavirus (SARS-CoV) 3CL Protease Inhibitors: Peptidomimetics and Small Molecule Chemotherapy. J. Med. Chem. 2016, 59, 6595–6628. [Google Scholar] [CrossRef] [PubMed]
- Liang, P.H. Characterization and inhibition of SARS-coronavirus main protease. Curr. Top. Med. Chem. 2006, 6, 361–376. [Google Scholar] [CrossRef] [PubMed]
- Rut, W.; Lv, Z.; Zmudzinski, M.; Patchett, S.; Nayak, D.; Snipas, S.J.; El Oualid, F.; Huang, T.T.; Bekes, M.; Drag, M.; et al. Activity profiling and crystal structures of inhibitor-bound SARS-CoV-2 papain-like protease: A framework for anti-COVID-19 drug design. Sci. Adv. 2020, 6, eabd4596. [Google Scholar] [CrossRef]
- Kurt Yilmaz, N.; Swanstrom, R.; Schiffer, C.A. Improving Viral Protease Inhibitors to Counter Drug Resistance. Trends Microbiol. 2016, 24, 547–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.; Heng, W.; Wang, Y.; Qiu, J.; Wei, X.; Peng, S.; Saleem, S.; Khan, M.; Ali, S.S.; Wei, D.Q. In silico and in vitro evaluation of kaempferol as a potential inhibitor of the SARS-CoV-2 main protease (3CLpro). Phytother. Res. 2021. [Google Scholar] [CrossRef]
- Schwarz, S.; Sauter, D.; Wang, K.; Zhang, R.; Sun, B.; Karioti, A.; Bilia, A.R.; Efferth, T.; Schwarz, W. Kaempferol derivatives as antiviral drugs against the 3a channel protein of coronavirus. Planta Med. 2014, 80, 177. [Google Scholar] [CrossRef] [Green Version]
- Park, J.Y.; Yuk, H.J.; Ryu, H.W.; Lim, S.H.; Kim, K.S.; Park, K.H.; Ryu, Y.B.; Lee, W.S. Evaluation of polyphenols from Broussonetia papyrifera as coronavirus protease inhibitors. J. Enzyme Inhib. Med. Chem. 2017, 32, 504–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aggarwal, V.; Tuli, H.S.; Tania, M.; Srivastava, S.; Ritzer, E.E.; Pandey, A.; Aggarwal, D.; Barwal, T.S.; Jain, A.; Kaur, G.; et al. Molecular mechanisms of action of epigallocatechin gallate in cancer: Recent trends and advancement. Semin. Cancer Biol. 2020. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Kaihatsu, K.; Nishino, K.; Ogawa, M.; Kato, N.; Yamaguchi, A. Antibacterial and antifungal activities of new acylated derivatives of epigallocatechin gallate. Front. Microbiol. 2012, 3, 53. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, K.; Honda, M.; Ikigai, H.; Hara, Y.; Shimamura, T. Inhibitory effects of (-)-epigallocatechin gallate on the life cycle of human immunodeficiency virus type 1 (HIV-1). Antivir. Res. 2002, 53, 19–34. [Google Scholar] [CrossRef]
- Ramalingam, M.; Kim, H.; Lee, Y.; Lee, Y.I. Phytochemical and Pharmacological Role of Liquiritigenin and Isoliquiritigenin From Radix Glycyrrhizae in Human Health and Disease Models. Front. Aging Neurosci. 2018, 10, 348. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, N.; Venkatesh, P.; Ganguli, S.; Sinkar, V.P. Role of polyphenol oxidase and peroxidase in the generation of black tea theaflavins. J. Agric. Food Chem. 1999, 47, 2571–2578. [Google Scholar] [CrossRef]
- Chen, C.N.; Lin, C.P.C.; Huang, K.K.; Chen, W.C.; Hsieh, H.P.; Liang, P.H.; Hsu, J.T.A. Inhibition of SARS-CoV 3C-like protease activity by theaflavin-3,3′- digallate (TF3). Evid. Based Complement. Alternat. Med. 2005, 2, 209–215. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Oh, W.K.; Ahn, J.S.; Kim, Y.H.; Mbafor, J.T.; Wandji, J.; Fomum, Z.T. Prenylisoflavonoids from Erythrina senegalensis as novel HIV-1 protease inhibitors. Planta Med. 2009, 75, 268–270. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.X.; Wan, M.; Dong, H.; But, P.P.H.; Foo, L.Y. Inhibitory activity of flavonoids and tannins against HIV-1 protease. Biol. Pharm. Bull. 2000, 23, 1072–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peersen, O.B. A Comprehensive Superposition of Viral Polymerase Structures. Viruses 2019, 11, 745. [Google Scholar] [CrossRef] [Green Version]
- Koonin, E.V.; Gorbalenya, A.E.; Chumakov, K.M. Tentative identification of RNA-dependent RNA polymerases of dsRNA viruses and their relationship to positive strand RNA viral polymerases. FEBS Lett. 1989, 252, 42–46. [Google Scholar] [CrossRef] [Green Version]
- Walsh, D.; Mathews, M.B.; Mohr, I. Tinkering with translation: Protein synthesis in virus-infected cells. Cold Spring Harb. Perspect. Biol. 2013, 5, a012351. [Google Scholar] [CrossRef]
- Zandi, K.; Musall, K.; Oo, A.; Cao, D.; Liang, B.; Hassandarvish, P.; Lan, S.; Slack, R.L.; Kirby, K.A.; Bassit, L. Baicalein and Baicalin Inhibit SARS-CoV-2 RNA-Dependent-RNA Polymerase. Microorganisms 2021, 9, 893. [Google Scholar] [CrossRef] [PubMed]
- Gansukh, E.; Kazibwe, Z.; Pandurangan, M.; Judy, G.; Kim, D.H. Probing the impact of quercetin-7-O-glucoside on influenza virus replication influence. Phytomedicine 2016, 23, 958–967. [Google Scholar] [CrossRef] [PubMed]
- Sha, B.; Luo, M. Structure of a bifunctional membrane-RNA binding protein, influenza virus matrix protein M1. Nat. Struct. Biol. 1997, 4, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Chen, S.; Wang, D.C.; Chen, Y.J.; Wang, Y.X.; Guo, M.; Zhou, C.L.; Dou, J. Oroxylin A suppresses influenza A virus replication correlating with neuraminidase inhibition and induction of IFNs. Biomed. Pharmacother. 2018, 97, 385–394. [Google Scholar] [CrossRef]
- Sithisarn, P.; Michaelis, M.; Schubert-Zsilavecz, M.; Cinatl, J., Jr. Differential antiviral and anti-inflammatory mechanisms of the flavonoids biochanin A and baicalein in H5N1 influenza A virus-infected cells. Antivir. Res. 2013, 97, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Wang, L.X. Baicalin inhibits influenza virus A replication via activation of type I IFN signaling by reducing miR-146a. Mol. Med. Rep. 2019, 20, 5041–5049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nayak, M.K.; Agrawal, A.S.; Bose, S.; Naskar, S.; Bhowmick, R.; Chakrabarti, S.; Sarkar, S.; Chawla-Sarkar, M. Antiviral activity of baicalin against influenza virus H1N1-pdm09 is due to modulation of NS1-mediated cellular innate immune responses. J. Antimicrob. Chemother. 2014, 69, 1298–1310. [Google Scholar] [CrossRef] [Green Version]
- Zu, M.; Li, C.; Fang, J.-S.; Lian, W.-W.; Liu, A.-L.; Zheng, L.-S.; Du, G.-H. Drug Discovery of Host CLK1 Inhibitors for Influenza Treatment. Molecules 2015, 20, 19735–19747. [Google Scholar] [CrossRef] [Green Version]
- Pielak, R.M.; Chou, J.J. Influenza M2 proton channels. Biochim. Biophys. Acta 2011, 1808, 522–529. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Xu, L.J.; Lian, W.W.; Pang, X.C.; Jia, H.; Liu, A.L.; Du, G.H. Anti-influenza effect and action mechanisms of the chemical constituent gallocatechin-7-gallate from Pithecellobium clypearia Benth. Acta Pharmacol. Sin. 2018, 39, 1913–1922. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.Y.; Wang, H.Q.; Ma, L.L.; Ma, X.P.; Yin, J.Q.; Wu, S.; Huang, H.; Li, Y.H. Cirsimaritin inhibits influenza A virus replication by downregulating the NF-kappa B signal transduction pathway. Virol. J. 2018, 15. [Google Scholar] [CrossRef]
- Yan, H.; Ma, L.; Wang, H.; Wu, S.; Huang, H.; Gu, Z.; Jiang, J.; Li, Y. Luteolin decreases the yield of influenza A virus in vitro by interfering with the coat protein I complex expression. J. Nat. Med. 2019, 73, 487–496. [Google Scholar] [CrossRef]
- Zhong, M.; Wang, H.Q.; Yan, H.Y.; Wu, S.; Gu, Z.Y.; Li, Y.H. Santin inhibits influenza A virus replication through regulating MAPKs and NF-kappaB pathways. J. Asian Nat. Prod. Res. 2019, 21, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.J.; Li, R.C.; Li, X.L.; He, J.; Jiang, S.B.; Liu, S.W.; Yang, J. Quercetin as an Antiviral Agent Inhibits Influenza A Virus (IAV) Entry. Viruses 2016, 8, 6. [Google Scholar] [CrossRef] [PubMed]
- Yazawa, K.; Kurokawa, M.; Obuchi, M.; Li, Y.; Yamada, R.; Sadanari, H.; Matsubara, K.; Watanabe, K.; Koketsu, M.; Tuchida, Y.; et al. Anti-influenza virus activity of tricin, 4’,5,7-trihydroxy-3’,5’-dimethoxyflavone. Antivir. Chem. Chemother. 2011, 22, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- König, R.; Stertz, S.; Zhou, Y.; Inoue, A.; Hoffmann, H.H.; Bhattacharyya, S.; Alamares, J.G.; Tscherne, D.M.; Ortigoza, M.B.; Liang, Y.; et al. Human host factors required for influenza virus replication. Nature 2010, 463, 813–817. [Google Scholar] [CrossRef]
- Zhan, Y.; Ta, W.; Tang, W.; Hua, R.; Wang, J.; Wang, C.; Lu, W. Potential antiviral activity of isorhamnetin against SARS-CoV-2 spike pseudotyped virus in vitro. Drug. Dev. Res. 2021. [Google Scholar] [CrossRef]
- Song, J.; Zhang, L.; Xu, Y.; Yang, D.; Yang, S.; Zhang, W.; Wang, J.; Tian, S.; Yang, S.; Yuan, T. The comprehensive study on the therapeutic effects of baicalein for the treatment of COVID-19 in vivo and in vitro. Biochem. Pharmacol. 2021, 183, 114302. [Google Scholar] [CrossRef]
- Ha, T.K.Q.; Dao, T.T.; Nguyen, N.H.; Kim, J.; Kim, E.; Cho, T.O.; Oh, W.K. Antiviral phenolics from the leaves of Cleistocalyx operculatus. Fitoterapia 2016, 110, 135–141. [Google Scholar] [CrossRef]
- Yang, F.; Zhou, W.L.; Liu, A.L.; Qin, H.L.; Lee, S.M.; Wang, Y.T.; Du, G.H. The protective effect of 3-deoxysappanchalcone on in vitro influenza virus-induced apoptosis and inflammation. Planta Med. 2012, 78, 968–973. [Google Scholar] [CrossRef]
- Wurzer, W.J.; Planz, O.; Ehrhardt, C.; Giner, M.; Silberzahn, T.; Pleschka, S.; Ludwig, S. Caspase 3 activation is essential for efficient influenza virus propagation. EMBO J. 2003, 22, 2717–2728. [Google Scholar] [CrossRef] [Green Version]
- Geiler, J.; Michaelis, M.; Naczk, P.; Leutz, A.; Langer, K.; Doerr, H.W.; Cinatl, J., Jr. N-acetyl-L-cysteine (NAC) inhibits virus replication and expression of pro-inflammatory molecules in A549 cells infected with highly pathogenic H5N1 influenza A virus. Biochem. Pharmacol. 2010, 79, 413–420. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Dou, J.; Teng, Z.J.; Yu, J.; Wang, T.T.; Lu, N.; Wang, H.; Zhou, C.L. Antiviral activity of baicalin against influenza A (H1N1/H3N2) virus in cell culture and in mice and its inhibition of neuraminidase. Arch. Virol. 2014, 159, 3269–3278. [Google Scholar] [CrossRef] [PubMed]
- Santoro, M.G.; Rossi, A.; Amici, C. NF-kappaB and virus infection: Who controls whom. EMBO J. 2003, 22, 2552–2560. [Google Scholar] [CrossRef]
- Kumar, N.; Xin, Z.T.; Liang, Y.; Ly, H.; Liang, Y. NF-kappaB signaling differentially regulates influenza virus RNA synthesis. J. Virol. 2008, 82, 9880–9889. [Google Scholar] [CrossRef] [Green Version]
- Clementi, N.; Scagnolari, C.; D’Amore, A.; Palombi, F.; Criscuolo, E.; Frasca, F.; Pierangeli, A.; Mancini, N.; Antonelli, G.; Clementi, M. Naringenin is a powerful inhibitor of SARS-CoV-2 infection in vitro. Pharmacol. Res. 2020, 163, 105255. [Google Scholar] [CrossRef] [PubMed]
- Fassina, G.; Buffa, A.; Benelli, R.; Varnier, O.E.; Noonan, D.M.; Albini, A. Polyphenolic antioxidant (-)-epigallocatechin-3-gallate from green tea as a candidate anti-HIV agent. AIDS 2002, 16, 939–941. [Google Scholar] [CrossRef]
- Stantchev, T.S.; Markovic, I.; Telford, W.G.; Clouse, K.A.; Broder, C.C. The tyrosine kinase inhibitor genistein blocks HIV-1 infection in primary human macrophages. Virus Res. 2007, 123, 178–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Áy, É.; Hunyadi, A.; Mezei, M.; Minárovits, J.; Hohmann, J. Flavonol 7-O-Glucoside Herbacitrin Inhibits HIV-1 Replication through Simultaneous Integrase and Reverse Transcriptase Inhibition. Evid. Based Complement. Alternat. Med. 2019, 2019, 1064793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.J.; Nguyen, V.H.; Nguyen, M.C.; Soejarto, D.D.; Pezzuto, J.M.; Fong, H.H.; Tan, G.T. Sesquiterpenes and butenolides, natural anti-HIV constituents from Litsea verticillata. Planta Med. 2005, 71, 452–457. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Narayanan, S.; Chang, K.O. Inhibition of influenza virus replication by plant-derived isoquercetin. Antivir. Res. 2010, 88, 227–235. [Google Scholar] [CrossRef]
- Choi, H.J.; Song, J.H.; Park, K.S.; Kwon, D.H. Inhibitory effects of quercetin 3-rhamnoside on influenza A virus replication. Eur. J. Pharm. Sci. 2009, 37, 329–333. [Google Scholar] [CrossRef]
- Qiu, X.G.; Kroeker, A.; He, S.H.; Kozak, R.; Audet, J.; Mbikay, M.; Chretienc, M. Prophylactic Efficacy of Quercetin 3-beta-O-D-Glucoside against Ebola Virus Infection. Antimicrob. Agents Chemother. 2016, 60, 5182–5188. [Google Scholar] [CrossRef] [Green Version]
- Abbas, A.K.; Lichtman, A.H.; Pillai, S. Cellular and Molecular Immunology; Elsevier: Amsterdam, The Netherlands, 2021. [Google Scholar]
- Bramhachari, P.V. Dynamics of Immune Activation in Viral Diseases; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Sadler, A.J.; Williams, B.R.J. Interferon-inducible antiviral effectors. Nat. Rev. Immunol. 2008, 8, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F. Molecular mechanisms of IFN-γ to up-regulate MHC class I antigen processing and presentation. Int. Rev. Immunol. 2009, 28, 239–260. [Google Scholar] [CrossRef]
- Abdolvahab, M.H.; Moradi-Kalbolandi, S.; Zarei, M.; Bose, D.; Majidzadeh-A, K.; Farahmand, L. Potential role of interferons in treating COVID-19 patients. Int. Immunopharmacol. 2021, 90, 107171. [Google Scholar] [CrossRef] [PubMed]
- Parmar, S.; Platanias, L.C. Interferons: Mechanisms of action and clinical applications. Curr. Opin. Oncol. 2003, 15, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, S.; Balkhy, H.; Gabere, M.N. Current treatment options and the role of peptides as potential therapeutic components for Middle East respiratory syndrome (MERS): A review. J. Infect. Public Health 2018, 11, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Morgenstern, B.; Michaelis, M.; Baer, P.C.; Doerr, H.W.; Cinatl, S., Jr. Ribavirin and interferon-β synergistically inhibit SARS-associated coronavirus replication in animal and human cell lines. Biochem. Biophys. Res. Commun. 2005, 326, 905–908. [Google Scholar] [CrossRef]
- Acosta, P.L.; Byrne, A.B.; Hijano, D.R.; Talarico, L.B. Human type I interferon antiviral effects in respiratory and reemerging viral infections. J. Immunol. Res. 2020, 2020. [Google Scholar] [CrossRef]
- Acharya, D.; Liu, G.; Gack, M.U. Dysregulation of type I interferon responses in COVID-19. Nat. Rev. Immunol. 2020, 20, 397–398. [Google Scholar] [CrossRef]
- Mishan, M.A.; Tabari, M.A.K.; Parnian, J.; Fallahi, J.; Mahrooz, A.; Bagheri, A.J.G. Chromosomes; Cancer, Functional mechanisms of miR-192 family in cancer. Genes Chromosomes Cancer 2020, 59, 722–735. [Google Scholar] [CrossRef]
- Mishan, M.A.; Tabari, M.A.K.; Zargari, M.; Bagheri, A. MicroRNAs in the anticancer effects of celecoxib: A systematic review. Eur. J. Pharmacol. 2020, 173325. [Google Scholar] [CrossRef]
- Ding, Z.; Sun, G.; Zhu, Z. Hesperidin attenuates influenza A virus (H1N1) induced lung injury in rats through its anti-inflammatory effect. Antivir. Ther. 2018, 23, 611–615. [Google Scholar] [CrossRef]
- Seong, R.K.; Kim, J.A.; Shin, O.S. Wogonin, a flavonoid isolated from Scutellaria baicalensis, has anti-viral activities against influenza infection via modulation of AMPK pathways. Acta Virol. 2018, 62, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.X.; Fung, T.S.; Chong, K.K.L.; Shukla, A.; Hilgenfeld, R. Accessory proteins of SARS-CoV and other coronaviruses. Antivir. Res. 2014, 109, 97–109. [Google Scholar] [CrossRef]
- Costela-Ruiz, V.J.; Illescas-Montes, R.; Puerta-Puerta, J.M.; Ruiz, C.; Melguizo-Rodríguez, L. SARS-CoV-2 infection: The role of cytokines in COVID-19 disease. Cytokine Growth Factor Rev. 2020, 54, 62–75. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Melo, D.; Nilsson-Payant, B.E.; Liu, W.-C.; Uhl, S.; Hoagland, D.; Møller, R.; Jordan, T.X.; Oishi, K.; Panis, M.; Sachs, D.J.C. Imbalanced host response to SARS-CoV-2 drives development of COVID-19. Cell 2020, 181, 1036–1045.e9. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Q.; Yang, K.; Wang, W.; Jiang, L.; Song, J. Clinical predictors of mortality due to COVID-19 based on an analysis of data of 150 patients from Wuhan, China. Intensive Care Med. 2020, 46, 846–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jose, R.J.; Manuel, A. COVID-19 cytokine storm: The interplay between inflammation and coagulation. Lancet Respir. Med. 2020, 8, e46–e47. [Google Scholar] [CrossRef]
- Roshanravan, N.; Seif, F.; Ostadrahimi, A.; Pouraghaei, M.; Ghaffari, S. Targeting cytokine storm to manage patients with COVID-19: A mini-review. Arch. Med. Res. 2020, 51, 608–612. [Google Scholar] [CrossRef] [PubMed]
- Fara, A.; Mitrev, Z.; Rosalia, R.A.; Assas, B.M. Cytokine storm and COVID-19: A chronicle of pro-inflammatory cytokines. Open Biol. 2020, 10, 200160. [Google Scholar] [CrossRef]
- Karki, R.; Sharma, B.R.; Tuladhar, S.; Williams, E.P.; Zalduondo, L.; Samir, P.; Zheng, M.; Sundaram, B.; Banoth, B.; Malireddi, R.K.S.; et al. COVID-19 cytokines and the hyperactive immune response: Synergism of TNF-α and IFN-γ in triggering inflammation, tissue damage, and death. Cell 2020, 184, 149–168.e17. [Google Scholar] [CrossRef]
- Yonekawa, M.; Shimizu, M.; Kaneko, A.; Matsumura, J.; Takahashi, H. Suppression of R5-type of HIV-1 in CD4(+) NKT cells by Vdelta1(+) T cells activated by flavonoid glycosides, hesperidin and linarin. Sci. Rep. 2019, 9, 7506. [Google Scholar] [CrossRef] [Green Version]
- Kang, J.; Liu, C.; Wang, H.; Li, B.; Li, C.; Chen, R.; Liu, A. Studies on the bioactive flavonoids isolated from Pithecellobium clypearia Benth. Molecules 2014, 19, 4479–4490. [Google Scholar] [CrossRef] [Green Version]
- Mehrbod, P.; Abdalla, M.A.; Fotouhi, F.; Heidarzadeh, M.; Aro, A.O.; Eloff, J.N.; McGaw, L.J.; Fasina, F.O. Immunomodulatory properties of quercetin-3-O-alpha-L-rhamnopyranoside from Rapanea melanophloeos against influenza a virus. BMC Complement. Altern. Med. 2018, 18, 184. [Google Scholar] [CrossRef] [Green Version]
- Barthelemy, A.; Ivanov, S.; Fontaine, J.; Soulard, D.; Bouabe, H.; Paget, C.; Faveeuw, C.; Trottein, F. Influenza A virus-induced release of interleukin-10 inhibits the anti-microbial activities of invariant natural killer T cells during invasive pneumococcal superinfection. Mucosal Immunol. 2017, 10, 460–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zu, M.; Yang, F.; Zhou, W.L.; Liu, A.L.; Du, G.H.; Zheng, L.S. In vitro anti-influenza virus and anti-inflammatory activities of theaflavin derivatives. Antivir. Res. 2012, 94, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Janeway Jr, C.A.; Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hur, S. Double-stranded RNA sensors and modulators in innate immunity. Annu. Rev. Immunol. 2019, 37, 349–375. [Google Scholar] [CrossRef] [PubMed]
- Chiou, W.F.; Chen, C.C.; Wei, B.L. 8-Prenylkaempferol Suppresses Influenza A Virus-Induced RANTES Production in A549 Cells via Blocking PI3K-Mediated Transcriptional Activation of NF-kappaB and IRF3. Evid. Based Complement. Alternat. Med. 2011, 2011, 920828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, H.Y.; Han, L.; Shi, X.L.; Wang, B.L.; Huang, H.; Wang, X.; Chen, D.F.; Ju, D.W.; Feng, M.Q. Baicalin inhibits autophagy induced by influenza A virus H3N2. Antivir. Res. 2015, 113, 62–70. [Google Scholar] [CrossRef]
- Abdal Dayem, A.; Choi, H.Y.; Kim, Y.B.; Cho, S.G. Antiviral effect of methylated flavonol isorhamnetin against influenza. PLoS ONE 2015, 10, e0121610. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.J.; Kim, D.H. 5,7-Dihydroxy-6-Methoxy-Flavonoids Eliminate HIV-1 D3-transfected Cytoprotective Macrophages by Inhibiting the PI3K/Akt Signaling Pathway. Phytother. Res. 2015, 29, 1355–1365. [Google Scholar] [CrossRef] [PubMed]
- Solnier, J.; Fladerer, J.P. Flavonoids: A complementary approach to conventional therapy of COVID-19? Phytochem. Rev. 2020, 1–23. [Google Scholar] [CrossRef]
- Russo, M.; Moccia, S.; Spagnuolo, C.; Tedesco, I.; Russo, G.L. Roles of flavonoids against coronavirus infection. Chem. Biol. Interact. 2020, 109211. [Google Scholar] [CrossRef]
- Jo, S.; Kim, S.; Shin, D.H.; Kim, M.S. Inhibition of SARS-CoV 3CL protease by flavonoids. J. Enzyme Inhib. Med. Chem. 2020, 35, 145–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jo, S.; Kim, H.; Kim, S.; Shin, D.H.; Kim, M.S. Characteristics of flavonoids as potent MERS-CoV 3C-like protease inhibitors. Chem. Biol. Drug. Des. 2019, 94, 2023–2030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhowmik, D.; Nandi, R.; Prakash, A.; Kumar, D. Evaluation of flavonoids as 2019-nCoV cell entry inhibitor through molecular docking and pharmacological analysis. Heliyon 2021, 7, e06515. [Google Scholar] [CrossRef]
- Bora, A.; Pacureanu, L.; Crisan, L. In silico study of some natural flavonoids as potential agents against COVID-19: Preliminary results. Chem. Proc. 2020, 3, 25. [Google Scholar] [CrossRef]
- Gorla, U.S.; Rao, G.K.; Kulandaivelu, U.S.; Alavala, R.R.; Panda, S.P. Lead Finding from Selected Flavonoids with Antiviral (SARS-CoV-2) Potentials against COVID-19: An in-silico Evaluation. Comb. Chem. High Throughput Screen. 2020. [Google Scholar] [CrossRef]
- Jain, A.S.; Sushma, P.; Dharmashekar, C.; Beelagi, M.S.; Prasad, S.K.; Shivamallu, C.; Prasad, A.; Syed, A.; Marraiki, N.; Prasad, K.S. In silico evaluation of flavonoids as effective antiviral agents on the spike glycoprotein of SARS-CoV-2. Saudi J. Biol. Sci. 2021, 28, 1040–1051. [Google Scholar] [CrossRef]
- Varughese, J.K.; Joseph Libin, K.; Sindhu, K.; Rosily, A.; Abi, T. Investigation of the inhibitory activity of some dietary bioactive flavonoids against SARS-CoV-2 using molecular dynamics simulations and MM-PBSA calculations. J. Biomol. Struct. Dyn. 2021, 1–16. [Google Scholar] [CrossRef]
- Gour, A.; Manhas, D.; Bag, S.; Gorain, B.; Nandi, U. Flavonoids as potential phytotherapeutics to combat cytokine storm in SARS-CoV-2. Phytother. Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- Liskova, A.; Samec, M.; Koklesova, L.; Samuel, S.M.; Zhai, K.; Al-Ishaq, R.K.; Abotaleb, M.; Nosal, V.; Kajo, K.; Ashrafizadeh, M. Flavonoids against the SARS-CoV-2 induced inflammatory storm. Biomed. Pharmacother. 2021, 111430. [Google Scholar] [CrossRef]
- Muchtaridi, M.; Fauzi, M.; Khairul Ikram, N.K.; Mohd Gazzali, A.; Wahab, H.A. Natural flavonoids as potential angiotensin-converting enzyme 2 inhibitors for anti-SARS-CoV-2. Molecules 2020, 25, 3980. [Google Scholar] [CrossRef]
- Santana, F.P.R.; Thevenard, F.; Gomes, K.S.; Taguchi, L.; Câmara, N.O.S.; Stilhano, R.S.; Ureshino, R.P.; Prado, C.M.; Lago, J.H.G. New perspectives on natural flavonoids on COVID-19-induced lung injuries. Phytother. Res. 2021. [Google Scholar] [CrossRef]
- Sanna, C.; Rigano, D.; Corona, A.; Piano, D.; Formisano, C.; Farci, D.; Franzini, G.; Ballero, M.; Chianese, G.; Tramontano, E.; et al. Dual HIV-1 reverse transcriptase and integrase inhibitors from Limonium morisianum Arrigoni, an endemic species of Sardinia (Italy). Nat. Prod. Res. 2019, 33, 1798–1803. [Google Scholar] [CrossRef]
- Liu, A.L.; Shu, S.H.; Qin, H.L.; Lee, S.M.; Wang, Y.T.; Du, G.H. In vitro anti-influenza viral activities of constituents from Caesalpinia sappan. Planta Med. 2009, 75, 337–339. [Google Scholar] [CrossRef] [Green Version]
- Miki, K.; Nagai, T.; Suzuki, K.; Tsujimura, R.; Koyama, K.; Kinoshita, K.; Furuhata, K.; Yamada, H.; Takahashi, K. Anti-influenza virus activity of biflavonoids. Bioorg. Med. Chem. Lett. 2007, 17, 772–775. [Google Scholar] [CrossRef]
- Bang, S.; Li, W.; Ha, T.K.Q.; Lee, C.; Oh, W.K.; Shim, S.H. Anti-influenza effect of the major flavonoids from Salvia plebeia R.Br. via inhibition of influenza H1N1 virus neuraminidase. Nat. Prod. Res. 2018, 32, 1224–1228. [Google Scholar] [CrossRef]
- De Freitas, C.S.; Rocha, M.E.N.; Sacramento, C.Q.; Marttorelli, A.; Ferreira, A.C.; Rocha, N.; de Oliveira, A.C.; de Oliveira Gomes, A.M.; Dos Santos, P.S.; da Silva, E.O.; et al. Agathisflavone, a Biflavonoid from Anacardium occidentale L. Inhibits Influenza Virus Neuraminidase. Curr. Top. Med. Chem. 2020, 20, 111–120. [Google Scholar] [CrossRef]
- Lee, J.S.; Kim, H.J.; Lee, Y.S. A new anti-HIV flavonoid from glucoronide from Chrysanthemum marifolium. Planta Med. 2003, 69, 859–861. [Google Scholar] [CrossRef]
- Li, B.Q.; Fu, T.; Yao, D.Y.; Mikovits, J.A.; Ruscetti, F.W.; Wang, J.M. Flavonoid baicalin inhibits HIV-1 infection at the level of viral entry. Biochem. Biophys. Res. Commun. 2000, 276, 534–538. [Google Scholar] [CrossRef]
- Chang, C.C.; You, H.L.; Huang, S.T. Catechin inhibiting the H1N1 influenza virus associated with the regulation of autophagy. J. Chin. Med. Assoc. 2020, 83, 386–393. [Google Scholar] [CrossRef]
- Song, J.M.; Lee, K.H.; Seong, B.L. Antiviral effect of catechins in green tea on influenza virus. Antivir. Res. 2005, 68, 66–74. [Google Scholar] [CrossRef]
- Muller, P.; Downard, K.M. Catechin inhibition of influenza neuraminidase and its molecular basis with mass spectrometry. J. Pharm. Biomed. Anal. 2015, 111, 222–230. [Google Scholar] [CrossRef]
- Jiang, F.; Chen, W.; Yi, K.; Wu, Z.; Si, Y.; Han, W.; Zhao, Y. The evaluation of catechins that contain a galloyl moiety as potential HIV-1 integrase inhibitors. Clin. Immunol. 2010, 137, 347–356. [Google Scholar] [CrossRef]
- Dao, T.T.; Tung, B.T.; Nguyen, P.H.; Thuong, P.T.; Yoo, S.S.; Kim, E.H.; Kim, S.K.; Oh, W.K. C-Methylated Flavonoids from Cleistocalyx operculatus and Their Inhibitory Effects on Novel Influenza A (H1N1) Neuraminidase. J. Nat. Prod. 2010, 73, 1636–1642. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Kim, S.Y.; Lee, H.W.; Shin, J.S.; Kim, P.; Jung, Y.S.; Jeong, H.S.; Hyun, J.K.; Lee, C.K. Inhibition of influenza virus internalization by (-)-epigallocatechin-3-gallate. Antivir. Res. 2013, 100, 460–472. [Google Scholar] [CrossRef] [PubMed]
- Nance, C.L.; Siwak, E.B.; Shearer, W.T. Preclinical development of the green tea catechin, epigallocatechin gallate, as an HIV-1 therapy. J. Allergy Clin. Immunol. 2009, 123, 459–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williamson, M.P.; McCormick, T.G.; Nance, C.L.; Shearer, W.T. Epigallocatechin gallate, the main polyphenol in green tea, binds to the T-cell receptor, CD4: Potential for HIV-1 therapy. J. Allergy Clin. Immunol. 2006, 118, 1369–1374. [Google Scholar] [CrossRef]
- Kawai, K.; Tsuno, N.H.; Kitayama, J.; Okaji, Y.; Yazawa, K.; Asakage, M.; Hori, N.; Watanabe, T.; Takahashi, K.; Nagawa, H. Epigallocatechin gallate, the main component of tea polyphenol, binds to CD4 and interferes with gp120 binding. J. Allergy Clin. Immunol. 2003, 112, 951–957. [Google Scholar] [CrossRef]
- Huh, J.; Ha, T.K.Q.; Kang, K.B.; Kim, K.H.; Oh, W.K.; Kim, J.; Sung, S.H. C-Methylated Flavonoid Glycosides from Pentarhizidium orientale Rhizomes and Their Inhibitory Effects on the H1N1 Influenza Virus. J. Nat. Prod. 2017, 80, 2818–2824. [Google Scholar] [CrossRef]
- Nguyen, T.T.H.; Kang, H.K.; Kim, Y.M.; Jang, T.S.; Kim, D. Inhibition effect of flavonoid compounds against neuraminidase expressed in Pichia pastoris. Biotechnol. Bioprocess Eng. 2014, 19, 70–75. [Google Scholar] [CrossRef]
- Sauter, D.; Schwarz, S.; Wang, K.; Zhang, R.H.; Sun, B.; Schwarz, W. Genistein as Antiviral Drug against HIV Ion Channel. Planta Med. 2014, 80, 682–687. [Google Scholar] [CrossRef] [PubMed]
- Ortega, J.T.; Serrano, M.L.; Suarez, A.I.; Baptista, J.; Pujol, F.H.; Rangel, H.R. Methoxyflavones from Marcetia taxifolia as HIV-1 Reverse Transcriptase Inhibitors. Nat. Prod. Commun. 2017, 12, 1677–1680. [Google Scholar] [CrossRef] [Green Version]
- Bang, S.; Ha, T.K.Q.; Lee, C.; Li, W.; Oh, W.K.; Shim, S.H. Antiviral activities of compounds from aerial parts of Salvia plebeia R. Br. J. Ethnopharmacol. 2016, 192, 398–405. [Google Scholar] [CrossRef]
- Connell, B.J.; Chang, S.Y.; Prakash, E.; Yousfi, R.; Mohan, V.; Posch, W.; Wilflingseder, D.; Moog, C.; Kodama, E.N.; Clayette, P.; et al. A Cinnamon-Derived Procyanidin Compound Displays Anti-HIV-1 Activity by Blocking Heparan Sulfate- and Co-Receptor- Binding Sites on gp120 and Reverses T Cell Exhaustion via Impeding Tim-3 and PD-1 Upregulation. PLoS ONE 2016, 11, e0165386. [Google Scholar] [CrossRef] [PubMed]
- Mehla, R.; Bivalkar-Mehla, S.; Chauhan, A. A flavonoid, luteolin, cripples HIV-1 by abrogation of tat function. PLoS ONE 2011, 6, e27915. [Google Scholar] [CrossRef] [Green Version]
- Tewtrakul, S.; Miyashiro, H.; Nakamura, N.; Hattori, M.; Kawahata, T.; Otake, T.; Yoshinaga, T.; Fujiwara, T.; Supavita, T.; Yuenyongsawad, S.; et al. HIV-1 integrase inhibitory substances from Coleus parvifolius. Phytother. Res. 2003, 17, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Pasetto, S.; Pardi, V.; Murata, R.M. Anti-HIV-1 activity of flavonoid myricetin on HIV-1 infection in a dual-chamber in vitro model. PLoS ONE 2014, 9, e115323. [Google Scholar] [CrossRef]
- Ortega, J.T.; Suarez, A.I.; Serrano, M.L.; Baptista, J.; Pujol, F.H.; Rangel, H.R. The role of the glycosyl moiety of myricetin derivatives in anti-HIV-1 activity in vitro. AIDS Res. Ther. 2017, 14, 57. [Google Scholar] [CrossRef] [Green Version]
- Ortega, J.T.; Estrada, O.; Serrano, M.L.; Contreras, W.; Orsini, G.; Pujol, F.H.; Rangel, H.R. Glycosylated Flavonoids from Psidium guineense as Major Inhibitors of HIV-1 Replication in vitro. Nat. Prod. Commun. 2017, 12, 1049–1052. [Google Scholar] [CrossRef] [Green Version]
- Li, L.Y.; Li, X.; Shi, C.; Deng, Z.W.; Fu, H.Z.; Proksch, P.; Lin, W.H. Pongamone A-E, five flavonoids from the stems of a mangrove plant, Pongamia pinnata. Phytochemistry 2006, 67, 1347–1352. [Google Scholar] [CrossRef]
- Park, J.Y.; Ko, J.A.; Kim, D.W.; Kim, Y.M.; Kwon, H.J.; Jeong, H.J.; Kim, C.Y.; Park, K.H.; Lee, W.S.; Ryu, Y.B. Chalcones isolated from Angelica keiskei inhibit cysteine proteases of SARS-CoV. J. Enzyme Inhib. Med. Chem. 2016, 31, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Tewtrakul, S.; Nakamura, N.; Hattori, M.; Fujiwara, T.; Supavita, T. Flavanone and flavonol glycosides from the leaves of Thevetia peruviana and their HIV-1 reverse transcriptase and HIV-1 integrase inhibitory activities. Chem. Pharm. Bull. 2002, 50, 630–635. [Google Scholar] [CrossRef] [Green Version]
- Sookkongwaree, K.; Geitmann, M.; Roengsumran, S.; Petsom, A.; Danielson, U.H. Inhibition of viral proteases by Zingiberaceae extracts and flavones isolated from Kaempferia parviflora. Pharmazie 2006, 61, 717–721. [Google Scholar]
- Liu, A.L.; Wang, H.D.; Lee, S.M.Y.; Wang, Y.T.; Du, G.H. Structure-activity relationship of flavonoids as influenza virus neuraminidase inhibitors and their in vitro anti-viral activities. Bioorg. Med. Chem. 2008, 16, 7141–7147. [Google Scholar] [CrossRef]
- Liu, A.L.; Liu, B.; Qin, H.L.; Lee, S.M.Y.; Wang, Y.T.; Du, G.H. Anti-influenza virus activities of flavonoids from the medicinal plant Elsholtzia rugulosa. Planta Med. 2008, 74, 847–851. [Google Scholar] [CrossRef] [Green Version]
- Reutrakul, V.; Ningnuek, N.; Pohmakotr, M.; Yoosook, C.; Napaswad, C.; Kasisit, J.; Santisuk, T.; Tuchinda, P. Anti HIV-1 flavonoid glycosides from Ochna integerrima. Planta Med. 2007, 73, 683–688. [Google Scholar] [CrossRef] [Green Version]
- Ha, S.Y.; Youn, H.; Song, C.S.; Kang, S.C.; Bae, J.J.; Kim, H.T.; Lee, K.M.; Eom, T.L.; Kim, I.S.; Kwak, J.H. Antiviral Effect of Flavonol Glycosides Isolated from the Leaf of Zanthoxylum piperitum on Influenza Virus. J. Microbiol. 2014, 52, 340–344. [Google Scholar] [CrossRef]
- Jeong, H.J.; Ryu, Y.B.; Park, S.J.; Kim, J.H.; Kwon, H.J.; Kim, J.H.; Park, K.H.; Rho, M.C.; Lee, W.S. Neuraminidase inhibitory activities of flavonols isolated from Rhodiola rosea roots and their in vitro anti-influenza viral activities. Bioorg. Med. Chem. 2009, 17, 6816–6823. [Google Scholar] [CrossRef] [PubMed]
- Datta, B.K.; Datta, S.K.; Khan, T.H.; Kundu, J.K.; Rashid, M.A.; Nahar, L.; Sarker, S.D. Anti-cholinergic, cytotoxic and anti-HIV-1 activities of sesquiterpenes and a flavonoid glycoside from the aerial parts of Polygonum viscosum. Pharm. Biol. 2004, 42, 18–23. [Google Scholar] [CrossRef]
- Zhang, G.H.; Wang, Q.; Chen, J.J.; Zhang, X.M.; Tam, S.C.; Zheng, Y.T. The anti-HIV-1 effect of scutellarin. Biochem. Biophys. Res. Commun. 2005, 334, 812–816. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khazeei Tabari, M.A.; Iranpanah, A.; Bahramsoltani, R.; Rahimi, R. Flavonoids as Promising Antiviral Agents against SARS-CoV-2 Infection: A Mechanistic Review. Molecules 2021, 26, 3900. https://doi.org/10.3390/molecules26133900
Khazeei Tabari MA, Iranpanah A, Bahramsoltani R, Rahimi R. Flavonoids as Promising Antiviral Agents against SARS-CoV-2 Infection: A Mechanistic Review. Molecules. 2021; 26(13):3900. https://doi.org/10.3390/molecules26133900
Chicago/Turabian StyleKhazeei Tabari, Mohammad Amin, Amin Iranpanah, Roodabeh Bahramsoltani, and Roja Rahimi. 2021. "Flavonoids as Promising Antiviral Agents against SARS-CoV-2 Infection: A Mechanistic Review" Molecules 26, no. 13: 3900. https://doi.org/10.3390/molecules26133900
APA StyleKhazeei Tabari, M. A., Iranpanah, A., Bahramsoltani, R., & Rahimi, R. (2021). Flavonoids as Promising Antiviral Agents against SARS-CoV-2 Infection: A Mechanistic Review. Molecules, 26(13), 3900. https://doi.org/10.3390/molecules26133900