
Functional Characterization of Porcine NK-Lysin: A Novel Immunomodulator That Regulates Intestinal Inflammatory Response

 , ,
, , 
Abstract
:1. Introduction
2. Results
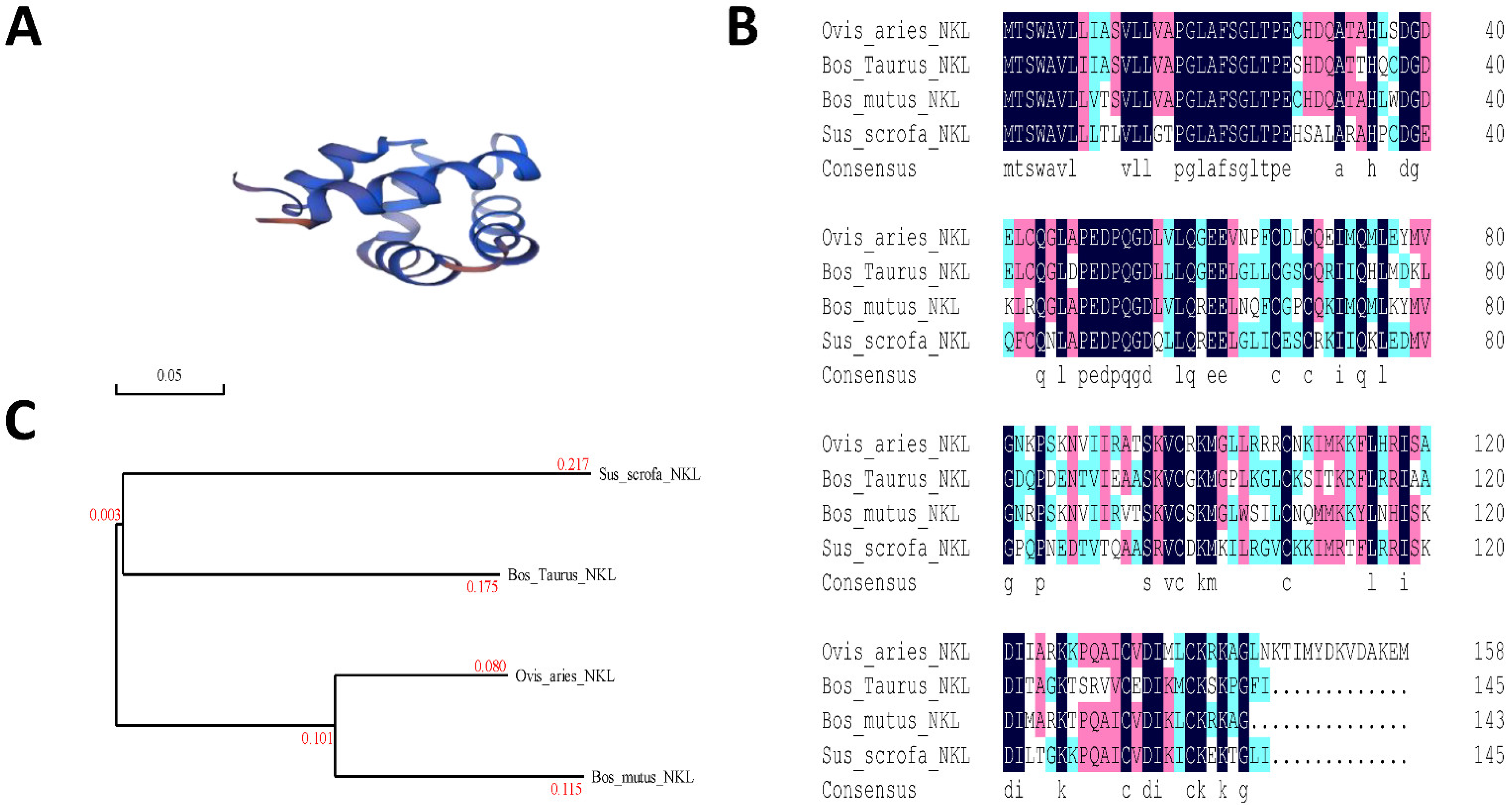
2.1. Comparison of the PNKL Nucleotide Sequences
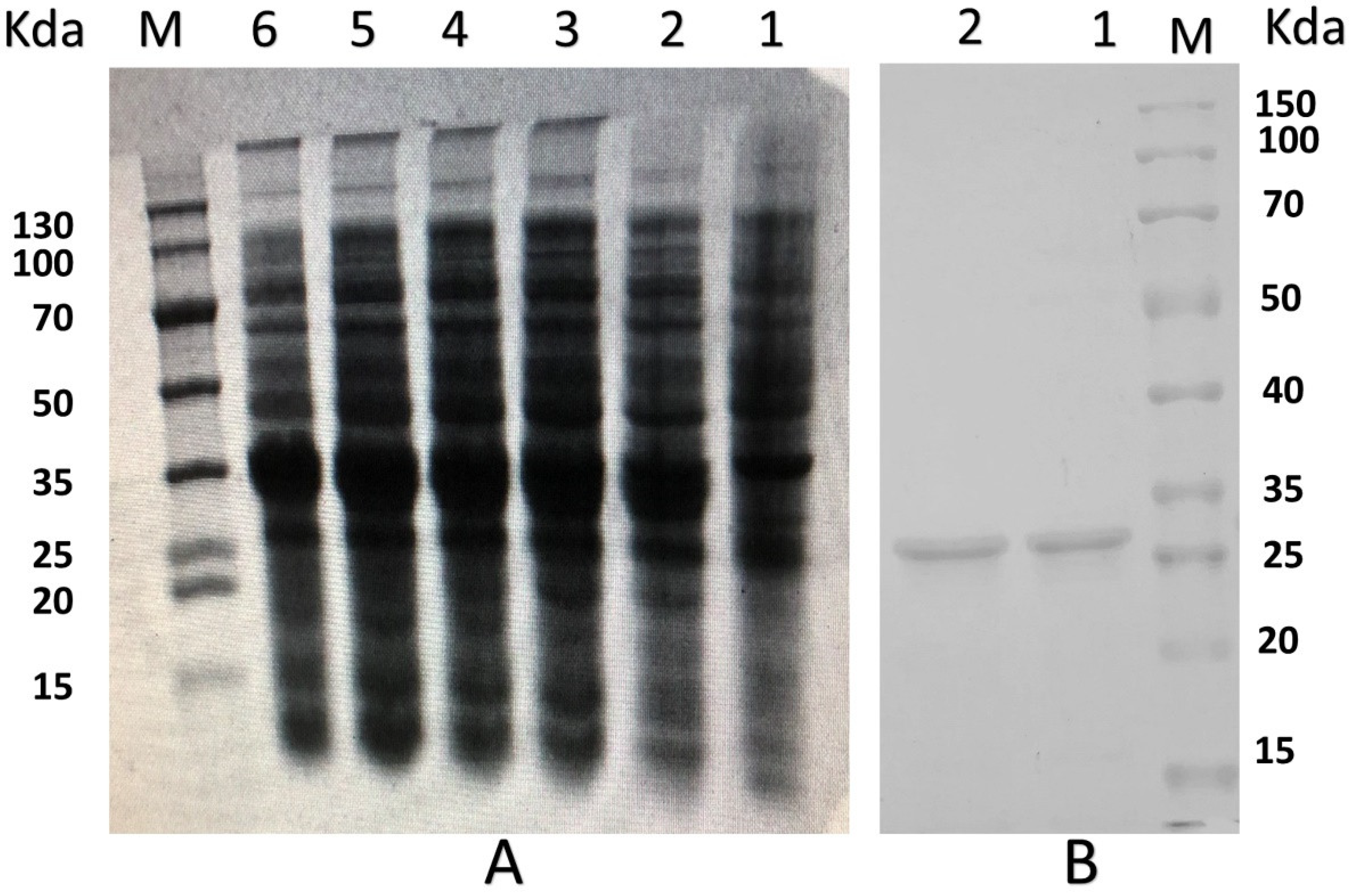
2.2. Expression and Purification of the Recombinant PNKL
2.3. Antibacterial Activity of the PNKL
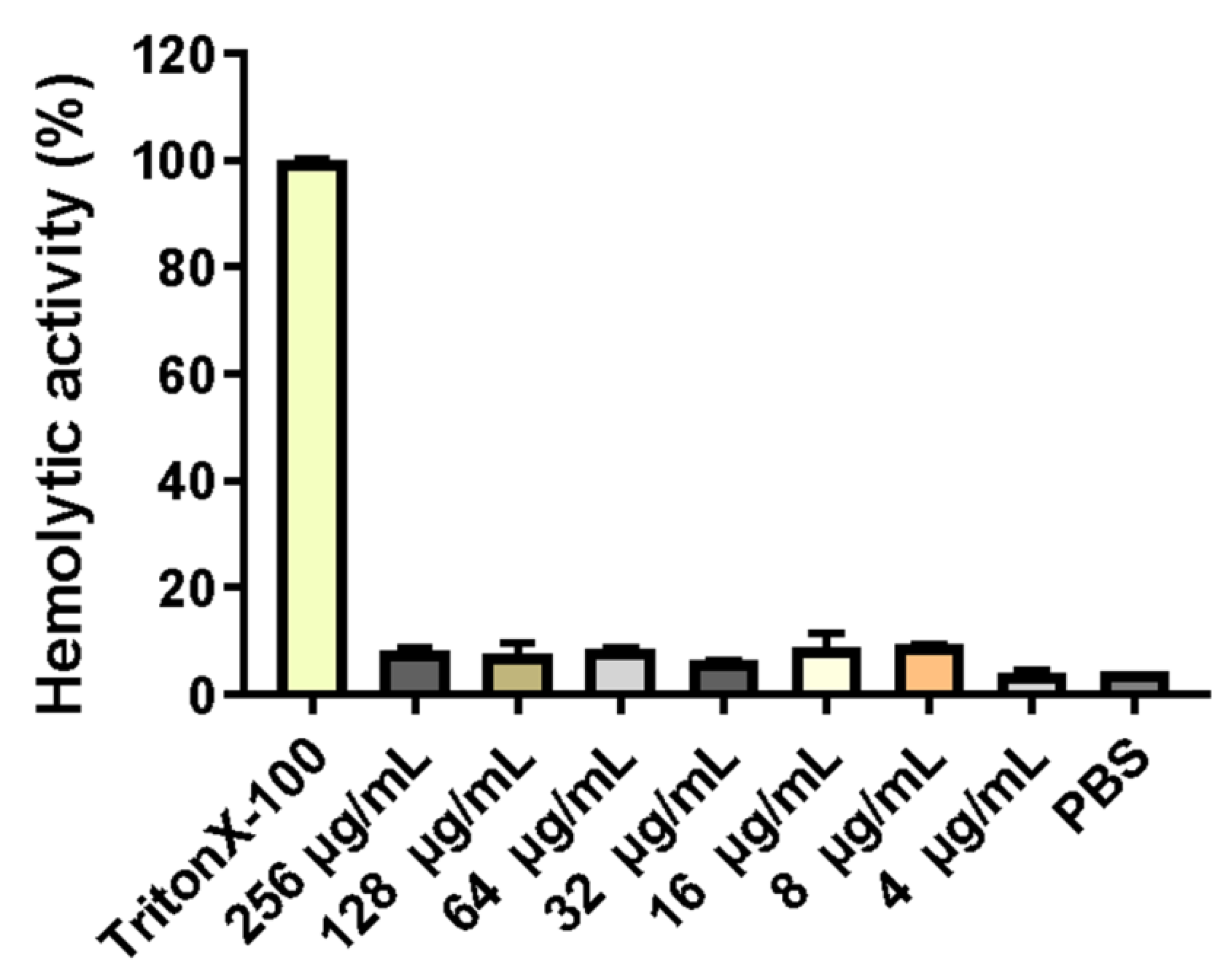
2.4. Hemolytic Activity of the PNKL
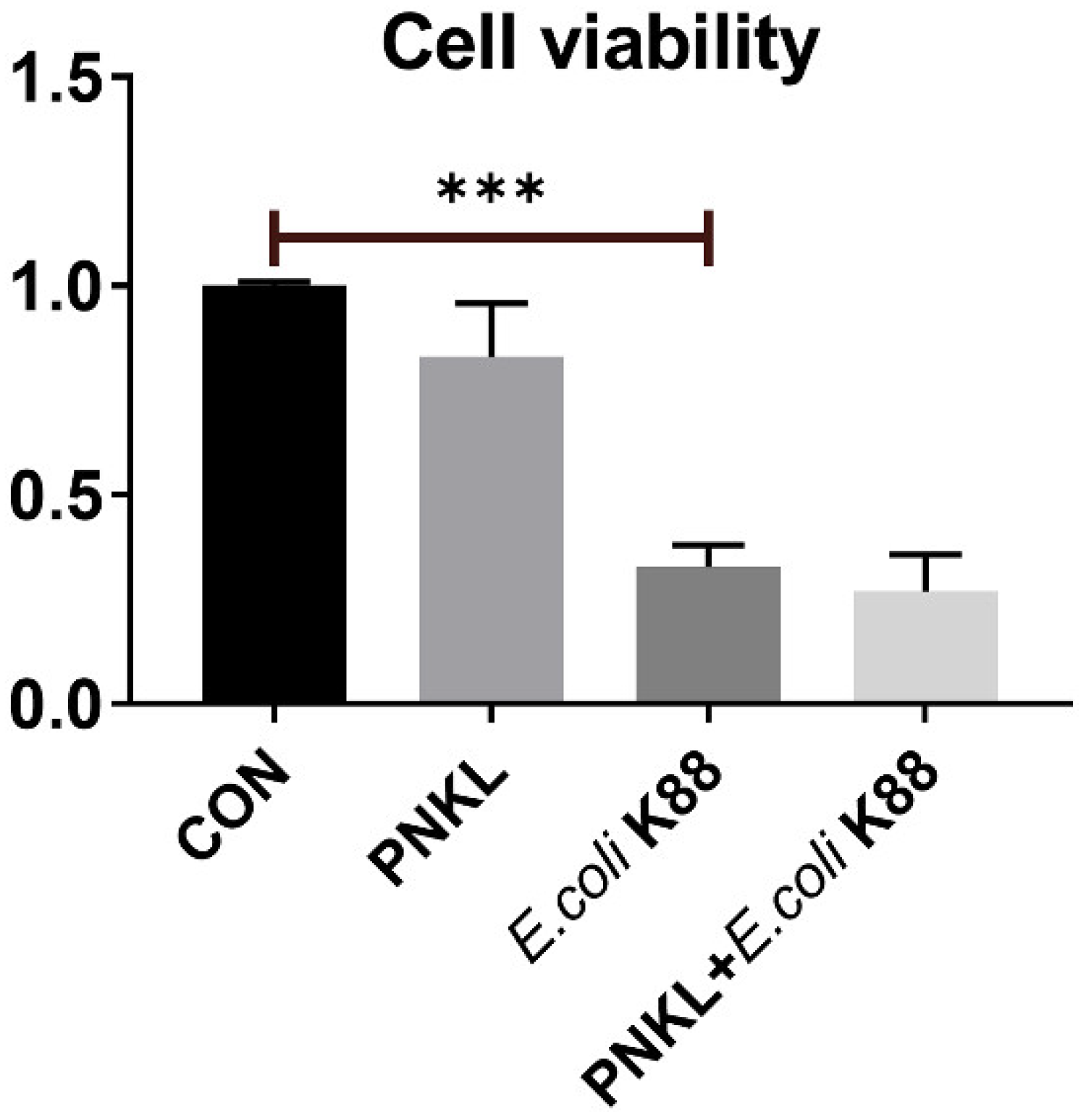
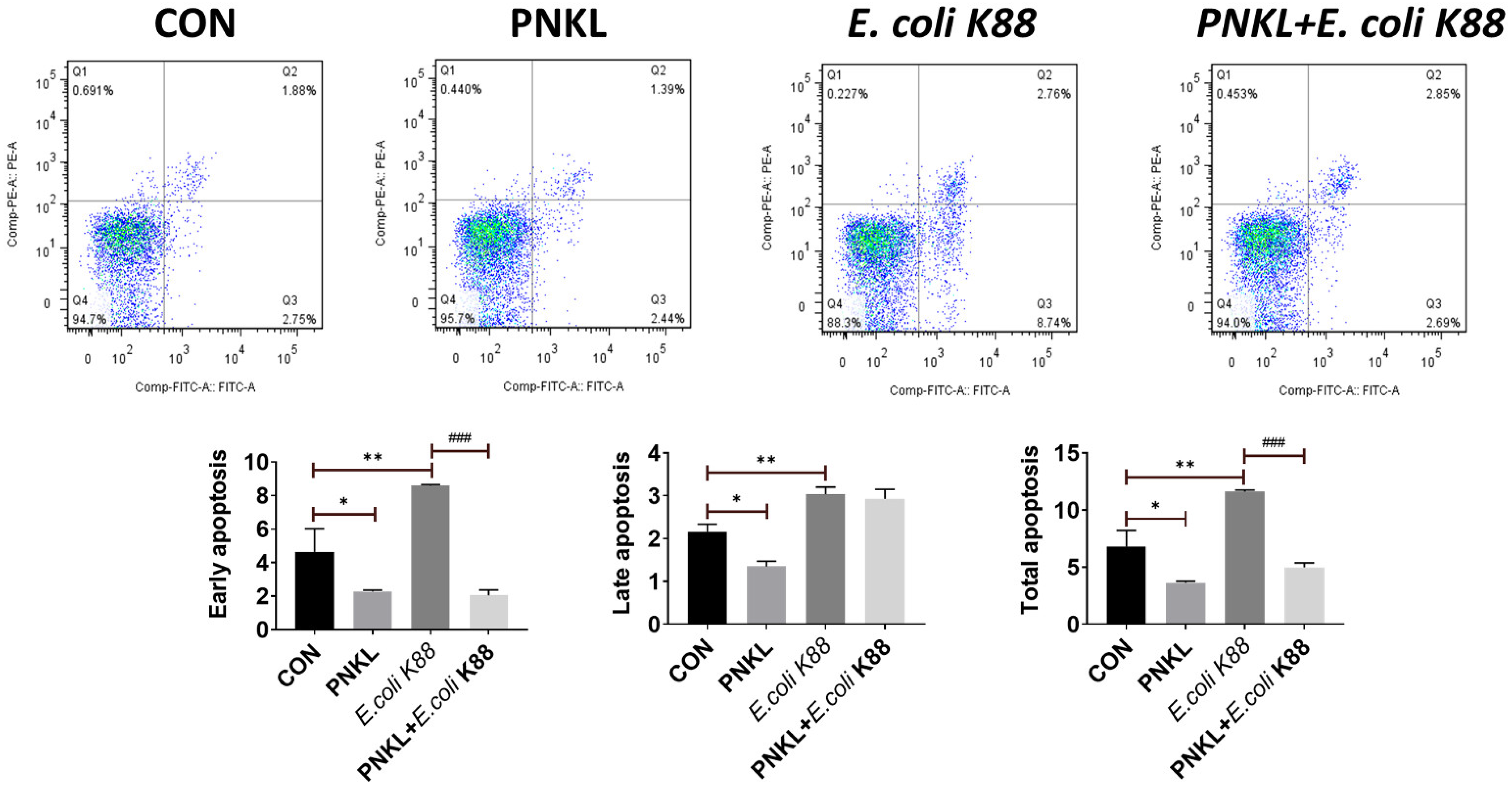
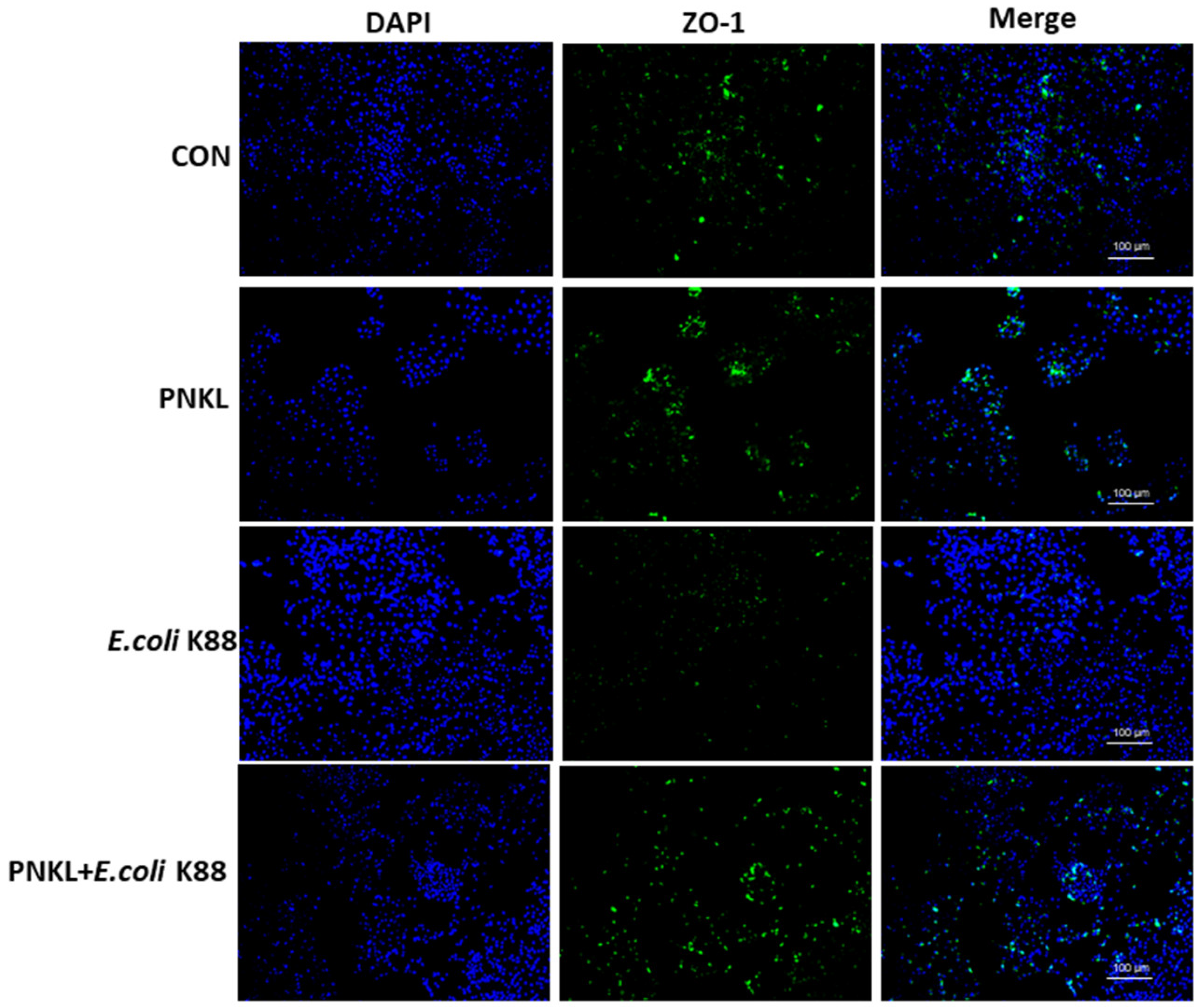
2.5. Effect of PNKL on Cell Viability, Apoptosis, and Localization of ZO-1 Protein in IPEC-J2 Cells Exposure to E. coli K88
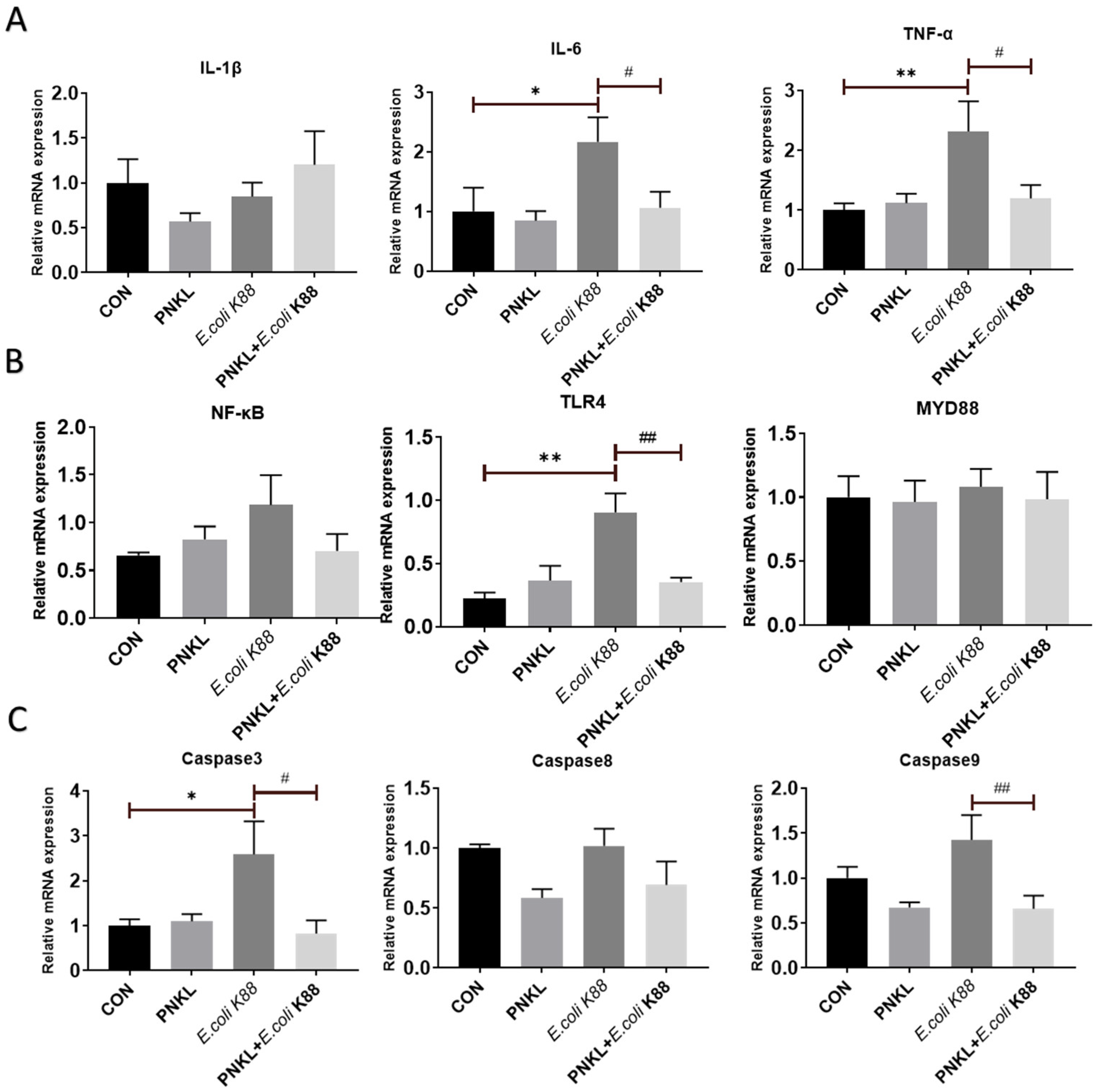
2.6. Effect of PNKL on Expressions of Genes Involved in Inflammatory Response and Cell Apoptosis
3. Discussion
4. Materials and Methods
4.1. Strains and Vectors
4.2. Plasmid Construction
4.3. Inducible Expression of PNKL
4.4. Affinity Purification
4.5. Antimicrobial Activity Assays
4.6. Hemolytic Activity Assay
4.7. Cell Culture
4.8. Cytotoxicity Assay
4.9. Assessment of Apoptosis by Flow Cytometry
4.10. RNA Extraction and RT-PCR
4.11. Immunofluorescence
4.12. Statistics Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviation
| MIC | minimal inhibitory concentration |
References
- Bosák, J.; Hrala, M.; Pirková, V.; Micenková, L.; Čížek, A.; Smola, J.; Kučerová, D.; Vacková, Z.; Budinská, E.; Koláčková, I.; et al. Porcine pathogenic Escherichia coli strains differ from human fecal strains in occurrence of bacteriocin types. Vet. Microbiol. 2019, 232, 121–127. [Google Scholar] [CrossRef]
- Chen, C.; Wang, L.-P.; Yu, J.-X.; Chen, X.; Wang, R.-N.; Yang, X.-Z.; Zheng, S.-F.; Yu, F.; Zhang, Z.-K.; Liu, S.-J.; et al. Prevalence of Enteropathogens in Outpatients with Acute Diarrhea from Urban and Rural Areas, Southeast China, 2010–2014. Am. J. Trop. Med. Hyg. 2019, 101, 310–318. [Google Scholar] [CrossRef]
- Al-Alo, K.Z.K.; Brujeni, G.N.; Lotfollahzadeh, S.; Moosakhani, F.; Gharabaghi, A. Correlation between neonatal calf diarrhea and the level of maternally derived antibodies. Iran. J. Vet. Res. 2018, 19, 3–8. [Google Scholar] [CrossRef]
- Gao, P.; Mao, D.; Luo, Y.; Wang, L.; Xu, B.; Xu, L. Occurrence of sulfonamide and tetracycline-resistant bacteria and resistance genes in aquaculture environment. Water Res. 2012, 46, 2355–2364. [Google Scholar] [CrossRef] [PubMed]
- Lieverse, R.I.Y.; Marcus, D.; Van Der Wiel, A.M.; Van Limbergen, E.J.; Theys, J.; Yaromina, A.; Lambin, P.; Dubois, L.J. Human fibronectin extra domain B as a biomarker for targeted therapy in cancer. Mol. Oncol. 2020, 14, 1555–1568. [Google Scholar] [CrossRef]
- Bessone, F.A.; Bessone, G.; Marini, S.; Conde, M.B.; Alustiza, F.E.; Zielinski, G. Presence and characterization of Escherichia coli virulence genes isolated from diseased pigs in the central region of Argentina. Vet. World 2017, 10, 939–945. [Google Scholar] [CrossRef] [Green Version]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Su, G.; Tang, F.; Chen, D.; Yu, B.; Huang, Z.; Luo, Y.; Mao, X.; Zheng, P.; Yu, J.; Luo, J.; et al. Expression, Purification and Characterization of a Novel Antimicrobial Peptide: Gloverin A2 from Bombyx mori. Int. J. Pept. Res. Ther. 2019, 25, 827–833. [Google Scholar] [CrossRef]
- Jenssen, H.; Hamill, P.; Hancock, R. Peptide Antimicrobial Agents. Clin. Microbiol. Rev. 2006, 19, 491–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semeraro, E.F.; Mandl, J.; Marx, L.; Narayanan, T.; Prévost, S.; Bergler, H.; Lohner, K.; Pabst, G. Antimicrobial Peptides Impair Bacteria Cell Structures within Seconds. Biophys. J. 2020, 118, 234A. [Google Scholar] [CrossRef]
- Hammami, R.; Ben Hamida, J.; Vergoten, G.; Lacroix, J.-M.; Slomianny, M.-C.; Mohamed, N.; Fliss, I. A new antimicrobial peptide isolated from Oudneya africana seeds. Microbiol. Immunol. 2009, 53, 658–666. [Google Scholar] [CrossRef]
- Martínez, M.; Polizzotto, A.; Flores, N.; Semorile, L.; Maffía, P.C. Antibacterial, anti-biofilm and in vivo activities of the antimicrobial peptides P5 and P6.2. Microb. Pathog. 2020, 139, 103886. [Google Scholar] [CrossRef]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef]
- Deng, Z.; Xu, C. Role of the neuroendocrine antimicrobial peptide catestatin in innate immunity and pain. Acta Biochim. Biophys. Sin. 2017, 49, 967–972. [Google Scholar] [CrossRef] [Green Version]
- Grassi, L.; Maisetta, G.; Maccari, G.; Esin, S.; Batoni, G. Analogs of the Frog-skin Antimicrobial Peptide Temporin 1Tb Exhibit a Wider Spectrum of Activity and a Stronger Antibiofilm Potential as Compared to the Parental Peptide. Front. Chem. 2017, 5, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, M.; Mukhopadhyay, K. Alpha-Melanocyte Stimulating Hormone: An Emerging Anti-Inflammatory Antimicrobial Peptide. BioMed Res. Int. 2014, 2014, 874610. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.; Curstedt, T.; Jörnvall, H.; Johansson, J. An amphipathic helical motif common to tumourolytic polypeptide NK-lysin and pulmonary surfactant polypeptide SP-B. FEBS Lett. 1995, 362, 328–332. [Google Scholar] [CrossRef] [Green Version]
- Hirono, I.; Kondo, H.; Koyama, T.; Arma, N.R.; Hwang, J.Y.; Nozaki, R.; Midorikawa, N.; Aoki, T. Characterization of Japanese flounder (Paralichthys olivaceus) NK-lysin, an antimicrobial peptide. Fish Shellfish Immunol. 2007, 22, 567–575. [Google Scholar] [CrossRef]
- Andersson, M.; Gunne, H.; Agerberth, B.; Boman, A.; Bergman, T.; Sillard, R.; Jörnvall, H.; Mutt, V.; Olsson, B.; Wigzell, H. NK-lysin, a novel effector peptide of cytotoxic T and NK cells. Structure and cDNA cloning of the porcine form, induction by interleukin 2, antibacterial and antitumour activity. EMBO J. 1995, 14, 1615–1625. [Google Scholar] [CrossRef]
- Hong, Y.H.; Lillehoj, H.S.; Siragusa, G.R.; Bannerman, D.D.; Lillehoj, E.P. Antimicrobial Activity of Chicken NK-Lysin against Eimeria Sporozoites. Avian Dis. 2008, 52, 302–305. [Google Scholar] [CrossRef]
- Zhang, M.; Long, H.; Sun, L. A NK-lysin from Cynoglossus semilaevis enhances antimicrobial defense against bacterial and viral pathogens. Dev. Comp. Immunol. 2013, 40, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.-X.; Wang, K.-R.; Chen, R.; Song, J.-J.; Zhang, B.-Z.; Dang, W.; Zhang, W.; Wang, R. Membrane active antitumor activity of NK-18, a mammalian NK-lysin-derived cationic antimicrobial peptide. Biochimie 2012, 94, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Banković, J.; Andrä, J.; Todorović, N.; Podolski-Renić, A.; Milošević, Z.; Miljković, Đ.; Krause, J.; Ruždijić, S.; Tanić, N.; Pešić, M. The elimination of P-glycoprotein over-expressing cancer cells by antimicrobial cationic peptide NK-2: The unique way of multi-drug resistance modulation. Exp. Cell Res. 2013, 319, 1013–1027. [Google Scholar] [CrossRef]
- Lee, M.O.; Kim, E.-H.; Jang, H.-J.; Na Park, M.; Woo, H.-J.; Han, J.Y.; Womack, J.E. Effects of a single nucleotide polymorphism in the chicken NK-lysin gene on antimicrobial activity and cytotoxicity of cancer cells. Proc. Natl. Acad. Sci. USA 2012, 109, 12087–12092. [Google Scholar] [CrossRef] [Green Version]
- Fan, K.; Du, W.; Wang, Z.; Khan, A.; Li, H.; Jiang, J. Anti-Invasion and Antimetastatic Effects of Porcine Recombinant NK-lysin on SMMC-7721 Human Hepatocellular Carcinoma Cells. BioMed Res. Int. 2019, 2019, 5318729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Q.; Su, G.; Wu, A.; Chen, D.; Yu, B.; Huang, Z.; Luo, Y.; Mao, X.; Zheng, P.; Yu, J.; et al. Bombyx mori gloverin A2 alleviates enterotoxigenic Escherichia coli-induced inflammation and intestinal mucosa disruption. Antimicrob. Resist. Infect. Control 2019, 8, 189. [Google Scholar] [CrossRef] [Green Version]
- Yusuf, E. Imminent threat of antibiotic resistance and the importance of diagnostic and antibiotic stewardship. Universa Med. 2017, 36, 157–159. [Google Scholar] [CrossRef] [Green Version]
- Lewies, A.; Du Plessis, L.H.; Wentzel, J.F. Antimicrobial Peptides: The Achilles’ Heel of Antibiotic Resistance? Probiotics Antimicrob. Proteins 2019, 11, 370–381. [Google Scholar] [CrossRef]
- Giuliani, A.; Pirri, G.; Nicoletto, S. Antimicrobial peptides: An overview of a promising class of therapeutics. Cent. Eur. J. Biol. 2007, 2, 1–33. [Google Scholar] [CrossRef]
- Lee, M.O.; Jang, H.-J.; Han, J.Y.; Womack, J.E. Chicken NK-lysin is an alpha-helical cationic peptide that exerts its antibacterial activity through damage of bacterial cell membranes. Poult. Sci. 2014, 93, 864–870. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.; Gunne, H.; Agerberth, B.; Boman, A.; Bergman, T.; Olsson, B.; Dagerlind, Å.; Wigzell, H.; Boman, H.; Gudmundsson, G. NK-lysin, structure and function of a novel effector molecule of porcine T and NK cells. Vet. Immunol. Immunopathol. 1996, 54, 123–126. [Google Scholar] [CrossRef]
- Lv, X.; Ma, Q.; Zhu, D.; Shao, C.; And, Y.L.; Shan, A. The C-terminal sequences of porcine thrombin are active as antimicrobial peptides. Chem. Biol. Drug Des. 2016, 88, 905–914. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhong, F.; Zhang, Y.; Zhang, J.; Huo, S.; Lin, H.; Wang, L.; Cui, D.; Li, X. Construction of Bacillus subtilis strain engineered for expression of porcine β-defensin-2/cecropin P1 fusion antimicrobial peptides and its growth-promoting effect and antimicrobial activity. Asian Australas. J. Anim. Sci. 2016, 30, 576–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, K.; Li, H.; Wang, Z.; Du, W.; Yin, W.; Sun, Y.; Jiang, J. Expression and purification of the recombinant porcine NK-lysin in Pichia pastoris and observation of anticancer activity in vitro. Prep. Biochem. Biotechnol. 2016, 46, 65–70. [Google Scholar] [CrossRef]
- Zhou, Q.-J.; Wang, J.; Liu, M.; Qiao, Y.; Hong, W.-S.; Su, Y.-Q.; Han, K.-H.; Ke, Q.-Z.; Zheng, W.-Q. Identification, expression and antibacterial activities of an antimicrobial peptide NK-lysin from a marine fish Larimichthys crocea. Fish Shellfish Immunol. 2016, 55, 195–202. [Google Scholar] [CrossRef]
- Hong, S.-M.; Kusakabe, T.; Lee, J.-M.; Tatsuke, T.; Kawaguchi, Y.; Kang, M.-W.; Kang, S.-W.; Kim, K.-A.; Nho, S.-K. Structure and Expression Analysis of the Cecropin-E Gene from the Silkworm, Bombyx mori. Biosci. Biotechnol. Biochem. 2008, 72, 1992–1998. [Google Scholar] [CrossRef]
- Lee, S.H.; Lillehoj, H.S.; Tuo, W.; Murphy, C.A.; Hong, Y.H.; Lillehoj, E.P. Parasiticidal activity of a novel synthetic peptide from the core α-helical region of NK-lysin. Vet. Parasitol. 2013, 197, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Fischer, U.; Koppang, E.O.; Nakanishi, T. Teleost T and NK cell immunity. Fish Shellfish Immunol. 2013, 35, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Sarson, A.J.; Abdul-Careem, M.F.; Read, L.R.; Brisbin, J.T.; Sharif, S. Expression of Cytotoxicity-Associated Genes in Marek’s Disease Virus—Infected Chickens. Viral Immunol. 2008, 21, 267–272. [Google Scholar] [CrossRef]
- Rauf, A.; Khatri, M.; Murgia, M.V.; Saif, Y.M. Expression of perforin-granzyme pathway genes in the bursa of infectious bursal disease virus-infected chickens. Dev. Comp. Immunol. 2011, 35, 620–627. [Google Scholar] [CrossRef]
- Kim, J.-K.; Lee, E.; Shin, S.; Jeong, K.-W.; Lee, J.-Y.; Bae, S.-Y.; Kim, S.-H.; Lee, J.; Kim, S.R.; Lee, D.G.; et al. Structure and Function of Papiliocin with Antimicrobial and Anti-inflammatory Activities Isolated from the Swallowtail Butterfly, Papilio xuthus. J. Biol. Chem. 2011, 286, 41296–41311. [Google Scholar] [CrossRef] [Green Version]
- Valero, Y.; Chaves-Pozo, E.; Cuesta, A. NK-lysin is highly conserved in European sea bass and gilthead seabream but differentially modulated during the immune response. Fish Shellfish Immunol. 2020, 99, 435–441. [Google Scholar] [CrossRef]
- Zou, T.; Zheng, C.; Zhang, Z.; Yu, L.; Fu, C. Neoadjuvant chemotherapy efficacy and prognostic factors in 187 cervical cancer patients with IB2 and IIA2 stage. J. Cent. South Univ. 2020, 45, 297–304. [Google Scholar] [CrossRef]
- Wang, K.; Su, R.; Sun, X.; Jiang, J.; Ma, Q. Progress in applications of 18F-fluciclovine in diagnosis of prostate cancer. J. Cent. South Univ. 2020, 45, 187–192. [Google Scholar] [CrossRef]
- Li, Y.; Shen, R.; Wen, G.; Ding, R.; Du, A.; Zhou, J.; Dong, Z.; Ren, X.; Yao, H.; Zhao, R.; et al. Effects of Ketamine on Levels of Inflammatory Cytokines IL-6, IL-1β, and TNF-α in the Hippocampus of Mice Following Acute or Chronic Administration. Front. Pharmacol. 2017, 8, 139. [Google Scholar] [CrossRef] [Green Version]
- McKay, D.M.; Baird, A.W. Cytokine regulation of epithelial permeability and ion transport. Gut 1999, 44, 283–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szczepankiewicz, D.; Skrzypski, M.; Pruszyńska-Oszmałek, E.; Kolodziejski, P.; Sassek, M.; Stefańska, B.; Nowak, K.W.; Szczepankiewicz, A. Interleukin 4 affects lipid metabolism and the expression of pro-inflammatory factors in mature rat adipocytes. Immunobiology 2018, 223, 677–683. [Google Scholar] [CrossRef] [PubMed]
- Zihni, C.; Mills, C.; Matter, K.; Balda, M. Tight junctions: From simple barriers to multifunctional molecular gates. Nat. Rev. Mol. Cell Biol. 2016, 17, 564–580. [Google Scholar] [CrossRef] [PubMed]
- Tong, L.-C.; Wang, Y.; Wang, Z.-B.; Liu, W.-Y.; Sun, S.; Li, L.; Su, D.-F.; Zhang, L.-C. Propionate Ameliorates Dextran Sodium Sulfate-Induced Colitis by Improving Intestinal Barrier Function and Reducing Inflammation and Oxidative Stress. Front. Pharmacol. 2016, 7, 253. [Google Scholar] [CrossRef]
- Taupin, D.; Podolsky, D.K. Trefoil factors: Initiators of mucosal healing. Nat. Rev. Mol. Cell Biol. 2003, 4, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yi, H.; Wang, Z.; Qiu, Y.; Wen, X.; Ma, X.; Yang, X.; Jiang, Z. 452 Effects of Lactobacillus reuteri LR1 on tight junction proteins expression in Ipec-1 cells during enterotoxigenic Escherichia coli K88 infection and its underlying mechanisms. J. Anim. Sci. 2017, 95, 222. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.-Y.; Zhu, Y.-H.; Zhang, W.; Zhou, D.; Zhai, C.-C.; Wang, J.-F. Influence of orally fed a select mixture of Bacillus probiotics on intestinal T-cell migration in weaned MUC4 resistant pigs following Escherichia coli challenge. Vet. Res. 2016, 47, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wollersheim, B.M.; Van Stam, M.-A.; Bosch, R.J.; Pos, F.J.; Tillier, C.N.; Van Der Poel, H.G.; Aaronson, N.K. Unmet expectations in prostate cancer patients and their association with decision regret. J. Cancer Surviv. 2020, 14, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Meng, X.-L.; Xu, J.-P.; Wang, J.; Wang, H.; Ma, C.-W. Production, purification, and characterization of the cecropin from Plutella xylostella, pxCECA1, using an intein-induced self-cleavable system in Escherichia coli. Appl. Microbiol. Biotechnol. 2012, 94, 1031–1039. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, J.; Li, Y.; Wu, H.; Zhao, S.; Yu, Q. Protecting intestinal epithelial cells against deoxynivalenol and E. coli damage by recombinant porcine IL-22. Vet. Microbiol. 2019, 231, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Pace, E.; Ferraro, M.; Di Vincenzo, S.; Siena, L.; Gjomarkaj, M. Effects of ceftaroline on the innate immune and on the inflammatory responses of bronchial epithelial cells exposed to cigarette smoke. Toxicol. Lett. 2016, 258, 216–226. [Google Scholar] [CrossRef]
- Scott, M.G.; Dullaghan, E.; Mookherjee, N.; Glavas, N.; Waldbrook, M.; Thompson, A.; Wang, A.; Lee, K.; Doria, S.; Hamill, P.; et al. An anti-infective peptide that selectively modulates the innate immune response. Nat. Biotechnol. 2007, 25, 465–472. [Google Scholar] [CrossRef]
- Mookherjee, N.; Brown, K.; Bowdish, D.M.E.; Doria, S.; Falsafi, R.; Hokamp, K.; Roche, F.M.; Mu, R.; Doho, G.H.; Pistolic, J.; et al. Modulation of the TLR-Mediated Inflammatory Response by the Endogenous Human Host Defense Peptide LL-37. J. Immunol. 2006, 176, 2455–2464. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.H.; Lillehoj, H.S.; Min, W. Evaluation of the Immunomodulatory Activity of the Chicken NK-Lysin-Derived Peptide cNK-2. Sci. Rep. 2017, 7, 45099. [Google Scholar] [CrossRef] [Green Version]
- Van Opdenbosch, N.; Lamkanfi, M. Caspases in Cell Death, Inflammation, and Disease. Immunity 2019, 50, 1352–1364. [Google Scholar] [CrossRef]
- Ramirez, M.L.G.; Salvesen, G.S. A primer on caspase mechanisms. Semin. Cell Dev. Biol. 2018, 82, 79–85. [Google Scholar] [CrossRef]
- Ponder, K.G.; Boise, L.H. The prodomain of caspase-3 regulates its own removal and caspase activation. Cell Death Discov. 2019, 5, 56. [Google Scholar] [CrossRef]
- Xia, Y.; Bin, P.; Liu, S.; Chen, S.; Yin, J.; Liu, G.; Tang, Z.; Ren, W. Enterotoxigenic Escherichia coli infection promotes apoptosis in piglets. Microb. Pathog. 2018, 125, 290–294. [Google Scholar] [CrossRef]
- Kim, J.; Xu, M.; Xo, R.; Mates, A.; Wilson, G.; Pearsall, A.; Grishko, V. Mitochondrial DNA damage is involved in apoptosis caused by pro-inflammatory cytokines in human OA chondrocytes. Osteoarthr. Cartil. 2010, 18, 424–432. [Google Scholar] [CrossRef] [Green Version]
- Grunnet, L.G.; Aikin, R.; Tonnesen, M.F.; Paraskevas, S.; Blaabjerg, L.; Størling, J.; Rosenberg, L.; Billestrup, N.; Maysinger, D.; Mandrup-Poulsen, T. Proinflammatory Cytokines Activate the Intrinsic Apoptotic Pathway in β-Cells. Diabetes 2009, 58, 1807–1815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Yang, Y.; Teng, D.; Tian, Z.; Wang, S.; Wang, J. Expression of plectasin in Pichia pastoris and its characterization as a new antimicrobial peptide against Staphyloccocus and Streptococcus. Protein Expr. Purif. 2011, 78, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Liu, C.; Guo, J.; Song, X.; Li, J.; Xu, W.; Li, Z. Recombinant expression, purification, and antimicrobial activity of a novel hybrid antimicrobial peptide LFT33. Appl. Microbiol. Biotechnol. 2012, 95, 1191–1198. [Google Scholar] [CrossRef]
- Deng, X.; Cao, M.; Zhang, J.; Hu, K.; Yin, Z.; Zhou, Z.; Xiao, X.; Yang, Y.; Sheng, W.; Wu, Y.; et al. Hyaluronic acid-chitosan nanoparticles for co-delivery of MiR-34a and doxorubicin in therapy against triple negative breast cancer. Biomaterials 2014, 35, 4333–4344. [Google Scholar] [CrossRef] [PubMed]
- Kusminski, C.M.; Park, J.; Scherer, P.E. MitoNEET-mediated effects on browning of white adipose tissue. Nat. Commun. 2014, 5, 3962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | MIC (µg/mL) |
|---|---|
| Gram-negative bacteria | |
| E. coli DH5α 32 | 32 |
| Pathogenic E. coli K88 | 4 |
| Salmonella typhimurium CICC14028 | 8 |
| Gram-positive bacteria | |
| Streptococcus | 16 |
| Staphylococcus aureus CICC23656 | 2 |
| Bacillus subtilis | 8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Q.; Fu, Q.; Chen, D.; Yu, B.; Luo, Y.; Huang, Z.; Zheng, P.; Mao, X.; Yu, J.; Luo, J.; et al. Functional Characterization of Porcine NK-Lysin: A Novel Immunomodulator That Regulates Intestinal Inflammatory Response. Molecules 2021, 26, 4242. https://doi.org/10.3390/molecules26144242
Lin Q, Fu Q, Chen D, Yu B, Luo Y, Huang Z, Zheng P, Mao X, Yu J, Luo J, et al. Functional Characterization of Porcine NK-Lysin: A Novel Immunomodulator That Regulates Intestinal Inflammatory Response. Molecules. 2021; 26(14):4242. https://doi.org/10.3390/molecules26144242
Chicago/Turabian StyleLin, Qian, Qingqing Fu, Daiwen Chen, Bing Yu, Yuheng Luo, Zhiqing Huang, Ping Zheng, Xiangbing Mao, Jie Yu, Junqiu Luo, and et al. 2021. "Functional Characterization of Porcine NK-Lysin: A Novel Immunomodulator That Regulates Intestinal Inflammatory Response" Molecules 26, no. 14: 4242. https://doi.org/10.3390/molecules26144242
APA StyleLin, Q., Fu, Q., Chen, D., Yu, B., Luo, Y., Huang, Z., Zheng, P., Mao, X., Yu, J., Luo, J., Yan, H., & He, J. (2021). Functional Characterization of Porcine NK-Lysin: A Novel Immunomodulator That Regulates Intestinal Inflammatory Response. Molecules, 26(14), 4242. https://doi.org/10.3390/molecules26144242