
Metabolomic Analysis of The Chemical Diversity of South Africa Leaf Litter Fungal Species Using an Epigenetic Culture-Based Approach

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Fungal Fermentations
2.2. Influence of Media Composition on Fungal Fermentations
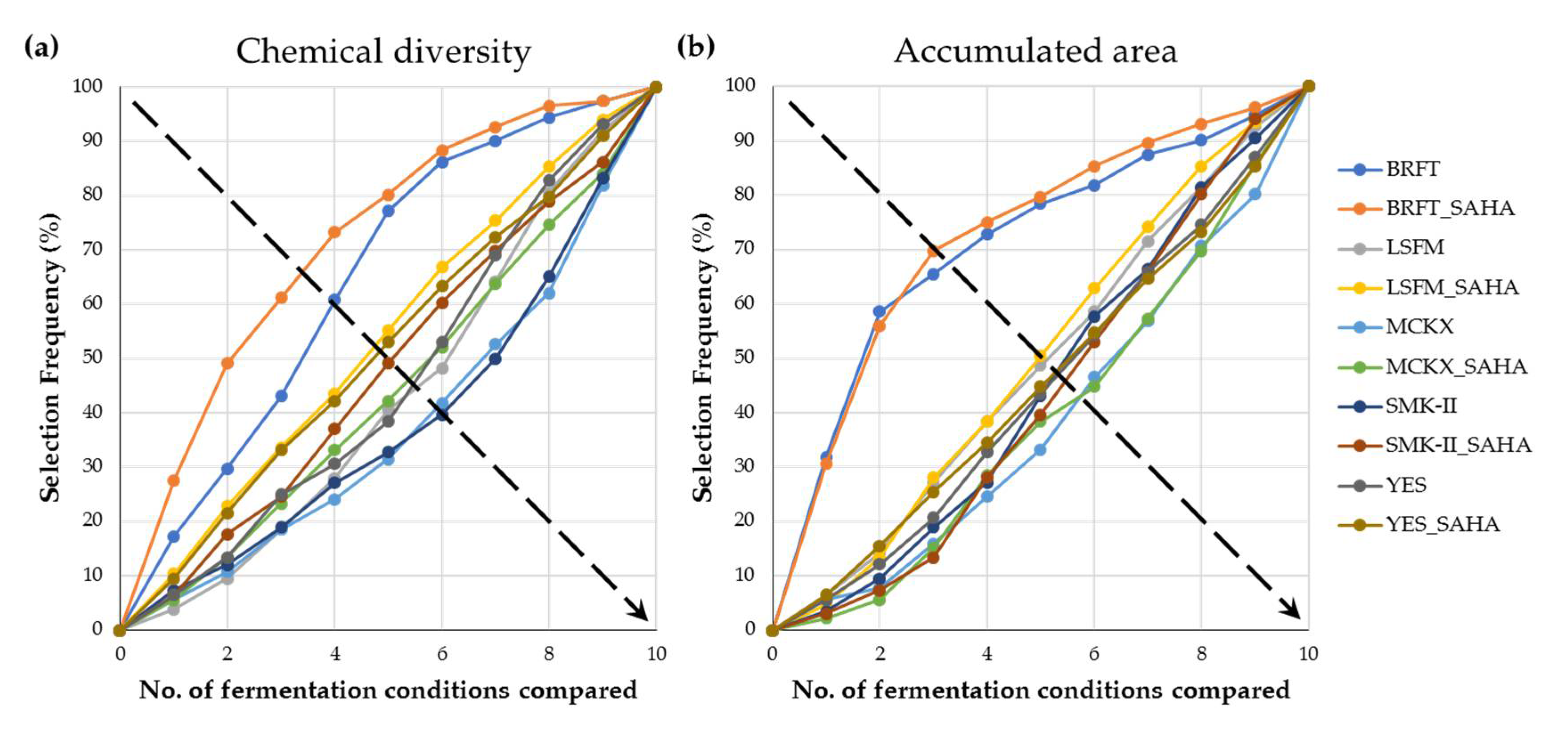
2.3. Chemical Evaluation of SAHA Addition during Fungal Fermentations
2.4. Effect of SAHA Addition to Different Taxonomic Orders
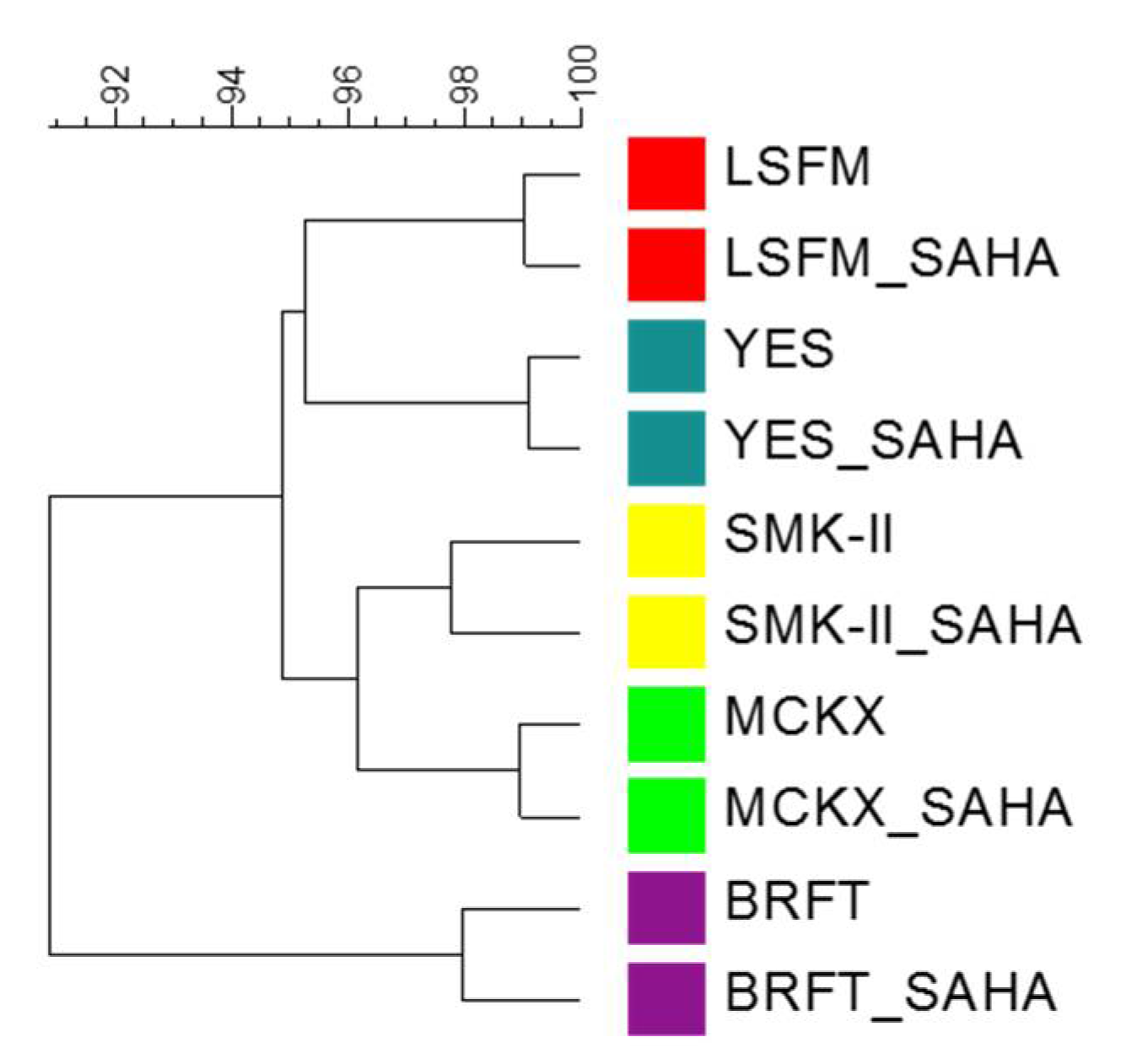
2.5. Chemometrical Analysis of Chemical Profiles
3. Materials and Methods
3.1. Fungal Strains
3.2. Fermentation Conditions
3.3. Chemical Extraction
3.4. LC-MS Profile Analysis and Database Matching of Known Metabolites
3.5. MASS Studio 2.3 Processing
3.6. HR-MS Profile Analysis and Database Matching of Known Metabolites
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Li, C.Y.; Chung, Y.M.; Wu, Y.C.; Hunyadi, A.; Wang, C.C.; Chang, F.R. Natural products development under epigenetic modulation in fungi. Phytochem. Rev. 2020, 19, 1323–1340. [Google Scholar] [CrossRef]
- Brice, A.P.; Wilson, B.A.; Thornburg, C.C.; Henrich, C.J.; Grkovic, T.; O’Keefe, B.R. Creating and screening natural product libraries. Nat. Prod. Rep. 2020, 37, 893–918. [Google Scholar]
- Hernandez, A.; Nguyen, L.T.; Dhakal, R.; Murphy, B.T. The need to innovate sample collection and library generation in microbial drug discovery: A focus on academia. Nat. Prod. Rep. 2021, 38, 292–300. [Google Scholar] [CrossRef]
- Bader, C.D.; Haack, P.A.; Panter, F.; Krug, D.; Müller, R. Expanding the scope of detectable microbial natural products by complementary analytical methods and cultivation systems. J. Nat. Prod. 2021, 84, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.B.; Henrikson, J.C.; Hoover, A.R.; Lee, A.E.; Cichewicz, R.H. Epigenetic remodeling of the fungal secondary metabolome. Org. Biomol. Chem. 2008, 6, 1895–1897. [Google Scholar] [CrossRef]
- Scherlach, K.; Hertweck, C. Triggering cryptic natural product biosynthesis in microorganisms. Org. Biomol. Chem. 2009, 7, 1753–1760. [Google Scholar] [CrossRef] [PubMed]
- Pettit, R.K. Small-molecule elicitation of microbial secondary metabolites. Microb. Biotechnol. 2011, 4, 471–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bigelis, R.; He, H.; Yang, H.Y.; Chan, L.; Greenstein, M. Production of fungal antibiotics using polymeric solid supports in solid-state and liquid fermentation. J. Ind. Microbiol. Biotechnol. 2006, 33, 815–826. [Google Scholar] [CrossRef]
- Bills, G.F.; Platas, G.; Fillola, A.; Jiménez, M.R.; Collado, J.; Vicente, F.; Martín, J.; González, A.; Bur-Zimmermann, J.; Tormo, J.R.; et al. Enhancement of antibiotic and secondary metabolite detection from filamentous fungi by growth on nutritional arrays. J. Appl. Microbiol. 2008, 104, 1644–1658. [Google Scholar] [CrossRef]
- González-Menéndez, V.; Asensio, F.; Moreno, C.; de Pedro, N.; Monteiro, M.C.; de la Cruz, M.; Vicente, F.; Bills, G.F.; Reyes, F.; Genilloud, O.; et al. Assessing the effects of adsorptive polymeric resin additions on fungal secondary metabolite chemical diversity. Mycology 2014, 5, 179–191. [Google Scholar] [CrossRef]
- González-Menéndez, V.; Pérez-Bonilla, M.; Pérez-Victoria, I.; Martín, J.; Muñoz, F.; Reyes, F.; Tormo, J.R.; Genilloud, O. Multicomponent analysis of the differential induction of secondary metabolite profiles in fungal endophytes. Molecules 2016, 21, 234. [Google Scholar] [CrossRef] [Green Version]
- González-Menéndez, V. Enhancement of chemical diversity in fungal endophytes from arid plants of Andalusia. Ph.D. Thesis, University of Granada, Granada, Spain, 2018. [Google Scholar]
- Schneider, T.; Keiblinger, K.M.; Schmid, E.; Sterflinger-Gleixner, K.; Ellersdorfer, G.; Roschitzki, B.; Richter, A.; Eberl, L.; Zechmeister-Boltenstern, S.; Riedel, K. Who is who in litter decomposition? Metaproteomics reveals major microbial players and their biogeochemical functions. ISME J. 2012, 6, 1749–1762. [Google Scholar] [CrossRef] [Green Version]
- Bills, G.F.; Polishook, J.D. Abundance and diversity of microfungi in leaf litter of a lowland rain forest in Costa Rica. Mycologia 1994, 86, 187–198. [Google Scholar] [CrossRef]
- Joanne, E.; Taylor, J.E.; Lee, S.; Crous, P.W. Biodiversity in the Cape Floral Kingdom: Fungi occurring on Proteaceae. Mycol. Res. 2001, 105, 1480–1484. [Google Scholar]
- Crous, P.W.; Rong, I.H.; Wood, A.; Lee, S.; Glen, H.; Botha, W.; Slippers, B.; de Beer, W.Z.; Wingfield, M.J.; Hawksworth, D.L. How many species of fungi are there at the tip of Africa? Stud. Mycol. 2006, 55, 13–33. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Mel Nik, V.; Taylor, J.E.; Crous, P.W. Diversity of saprobic hyphomycetes on Proteaceae and Restionaceae from South Africa. Fungal Diversity 2004, 17, 91–114. [Google Scholar]
- Mel Nik, V.; Lee, S.; Groenewal, J.Z.; Crous, P.W. New hyphomycetes from Restionaceae in fynbos: Parasarcopodium ceratocaryi gen. et sp. nov., and Rhexodenticula elegiae sp. nov. Mycol. Progress 2004, 3, 19–28. [Google Scholar]
- Magotra, A.; Kumar, M.; Kushwaha, M.; Awasthi, P.; Raina, C.; Gupta, A.P.; Shah, B.A.; Gandhi, S.G.; Chaubey, A. Epigenetic modifier induced enhancement of fumiquinazoline C production in Aspergillus fumigatus (GA-L7): An endophytic fungus from Grewia asiatica L. AMB Expr. 2017, 7, 43. [Google Scholar] [CrossRef]
- Zhu, J.X.; Ding, L.; He, S. Discovery of a new biphenyl derivative by epigenetic manipulation of marine-derived fungus Aspergillus versicolor. Nat. Prod. Res. 2019, 33, 1191–1195. [Google Scholar] [CrossRef]
- Kouipou, R.; Sahal, D.; Boyom, F. Recent advances in inducing endophytic fungal specialized metabolites using small molecule elicitors including epigenetic modifiers. Phytochemistry 2020, 174, 112338. [Google Scholar]
- Shwab, E.K.; Bok, J.W.; Tribus, M.; Galehr, J.; Graessle, S.; Keller, N.P. Histone deacetylase activity regulates chemical diversity in Aspergillus. Eukaryot. Cell 2007, 6, 1656–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Hawary, S.S.; Sayed, A.M.; Mohammed, R.; Hassan, H.M.; Zaki, M.A.; Mostafa, E.; Rateb, M.E.; Mohammed, T.A.; Amin, E.; Abdelmohsen, U.R. Epigenetic modifiers induce bioactive phenolic metabolites in the marine-derived fungus Penicillium brevicompactum. Mar. Drugs 2018, 16, 253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henrikson, J.C.; Hoover, A.R.; Joyner, P.M.; Cichewicz, R.H. A chemical epigenetics approach for engineering the in situ biosynthesis of a cryptic natural product from Aspergillu niger. Org. Biomol. Chem. 2009, 7, 435–438. [Google Scholar] [CrossRef] [PubMed]
- Beau, J.; Mahid, N.; Burda, W.N.; Harrington, L.; Shaw, L.N.; Mutka, T.; Kyle, D.E.; Barisic, B.; Van Olphen, A.; Baker, B.J. Epigenetic tailoring for the production of anti-infective cytosporones from the marine fungus Leucostoma persoonia. Mar. Drugs 2012, 10, 762–774. [Google Scholar] [CrossRef] [PubMed]
- Gubiani, J.R.; Kithsiri, E.M.; Shi, T.; Araujob, A.R.; Arnold, E.; Chapmanc, E.; Gunatilaka, A.A. An epigenetic modifier induces production of (10′S)-verruculide B, an inhibitor of protein tyrosine phosphatases by Phoma sp. nov. LG0217, a fungal endophyte of Parkinsonia microphylla. Bioorg. Med. Chem. 2017, 25, 1860–1866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Fang, H.; Yin, C.; Wei, C.; Hu, J.; Zhang, Y. Antimicrobial metabolites produced by Penicillium mallochii CCH01 isolated from the gut of Ectropis oblique, cultivated in the presence of a histone deacetylase inhibitor. Front. Microbiol. 2019, 10, 2186. [Google Scholar] [CrossRef] [PubMed]
- Martínez, G.; González-Menéndez, V.; Martín, J.; Reyes, F.; Genilloud, O.; Tormo, J.R. MASS Studio: A novel software utility to simplify LC-MS analyses of large sets of samples for metabolomics. In Proceedings of the Bioinformatics and Biomedical Engineering: 5th International Work-Conference, IWBBIO, Granada, Spain, 26–28 April 2017; Ignacio Rojas, Francisco Ortuño; Lecture Notes in Computer Science, volume 10208; Springer: Cham, Switzerland, 2017. [Google Scholar]
- González-Menéndez, V.; Martínez, G.; Serrano, R.; Muñoz, F.; Martín, J.; Genilloud, O.; Tormo, J.R. Ultraviolet (IUV) and mass spectrometry (IMS) imaging for the deconvolution of microbial interactions. BMC Syst. Biol. 2018, 12, 99. [Google Scholar] [CrossRef] [Green Version]
- Bills, G.F.; Dombrowski, A.W.; Goetz, M.A. The “FERMEX” Method for Metabolite-Enriched Fungal Extracts. In Fungal Secondary Metabolism: Methods and Protocols, Methods in Molecular Biology; Keller, N.P., Turner, G., Eds.; Springer Science+Business Media: Totowa, NJ, USA, 2012; Volume 944, pp. 79–96. [Google Scholar]
- Barrios-González, J.; Mejía, A. Production of secondary metabolites by solid-state fermentation. Biotechnol. Annu. Rev. 1996, 2, 85–121. [Google Scholar]
- Manan, M.A.; Webb, C. Design aspects of solid state fermentation as applied to microbial bioprocessing. J. Appl. Biotechnol. Bioeng. 2017, 4, 511–532. [Google Scholar]
- Yao, F.; Liu, S.C.; Wang, D.N.; Liu, Z.J.; Hua, Q.; Wei, L.J. Engineering oleaginous yeast Yarrowia lipolytica for enhanced limonene production from xylose and lignocellulosic hydrolysate. FEMS Yeast Res. 2020, 20, 6. [Google Scholar] [CrossRef]
- Petersen, L.A.; Hughes, D.L.; Hughes, R.; Di Michele, L.; Salmon, P.; Connors, N. Effects of amino acid and trace element supplementation on pneumocandin production by Glarea lozoyensis: Impact on titer, analogue levels, and the identification of the new analogues of pneumocandin B0. J. Ind. Microbiol. Biotechnol. 2001, 26, 216–221. [Google Scholar] [CrossRef]
- Fernández-Miranda, E.; Casares, A. Influence of the culture media and the organic matter in the growth of paxillus ammoniavirescens (Contu & Dessi). Mycobiology 2017, 45, 172–177. [Google Scholar]
- Pérez-Victoria, I.; Martín, J.; Reyes, F. Combined LC/UV/MS and NMR strategies for the dereplication of marine natural products. Planta. Med. 2016, 82, 857–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García, J.B.; Tormo, J.R. HPLC-studio: A novel software utility to perform HPLC chromatogram comparison for screening purposes. J. Biomol. Screen 2003, 8, 305–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tormo, J.R.; García, J.B.; De Antonio, M.; Feliz, J.; Mira, A.; Díez, M.T.; Hernández, P.; Peláez, F. A method for the selection of production media for actinomycete strains based on their metabolite HPLC profiles. J. Ind. Microbiol. Biotechnol. 2003, 30, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Tormo, J.R.; Garcia, J.B. Automated Analyses of HPLC Profiles of Microbial Extracts: A new tool for drug discovery screening. In Natural Products: Drug Discovery and Therapeutic Medicine; Zhang, L., Demain, A.L., Eds.; Humana Press: Totowa, NJ, USA, 2005; Volume 3, pp. 57–75. [Google Scholar]
- Knapp, D.G.; Kovács, G.M. Interspecific metabolic diversity of root-colonizing endophytic fungi revealed by enzyme activity tests. FEMS Microbiol. Ecol. 2016, 92, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peláez, F.; Cabello, A.; Platas, G.; Díez, M.T.; González, A.; Basilio, A.; Martán, I.; Vicente, F.; Bills, G.F.; Giacobbe, R.A.; et al. The discovery of enfumafungin, a novel antifungal compound produced by endophytic Hormonema species, biological activity, and taxonomy of the producing organisms. Syst. Appl. Microbiol. 2000, 23, 333–343. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SAHA | NONE | |
|---|---|---|
| Exclusive m/z (MS positive mode) | [350, 400] | [400, 450] |
| Exclusive m/z (MS negative mode) | [−350, −400] | [−350, −400] |
| Exclusive rt (MS positive mode) | [0.5, 1.0], [2.5, 3.0] | [0.5, 1.0], [3.5, 4.0] |
| Exclusive rt (MS negative mode) | [−0.5, −1.0], [−3.5, −4.0] | [−0.5, −1.0], [−3.5, −4.0] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serrano, R.; González-Menéndez, V.; Martínez, G.; Toro, C.; Martín, J.; Genilloud, O.; Tormo, .R. Metabolomic Analysis of The Chemical Diversity of South Africa Leaf Litter Fungal Species Using an Epigenetic Culture-Based Approach. Molecules 2021, 26, 4262. https://doi.org/10.3390/molecules26144262
Serrano R, González-Menéndez V, Martínez G, Toro C, Martín J, Genilloud O, Tormo R. Metabolomic Analysis of The Chemical Diversity of South Africa Leaf Litter Fungal Species Using an Epigenetic Culture-Based Approach. Molecules. 2021; 26(14):4262. https://doi.org/10.3390/molecules26144262
Chicago/Turabian StyleSerrano, Rachel, Víctor González-Menéndez, Germán Martínez, Clara Toro, Jesús Martín, Olga Genilloud, and José R. Tormo. 2021. "Metabolomic Analysis of The Chemical Diversity of South Africa Leaf Litter Fungal Species Using an Epigenetic Culture-Based Approach" Molecules 26, no. 14: 4262. https://doi.org/10.3390/molecules26144262
APA StyleSerrano, R., González-Menéndez, V., Martínez, G., Toro, C., Martín, J., Genilloud, O., & Tormo, . R. (2021). Metabolomic Analysis of The Chemical Diversity of South Africa Leaf Litter Fungal Species Using an Epigenetic Culture-Based Approach. Molecules, 26(14), 4262. https://doi.org/10.3390/molecules26144262