
Doxycycline Interferes with Zika Virus Serine Protease and Inhibits Virus Replication in Human Skin Fibroblasts



Abstract
:1. Introduction
2. Results
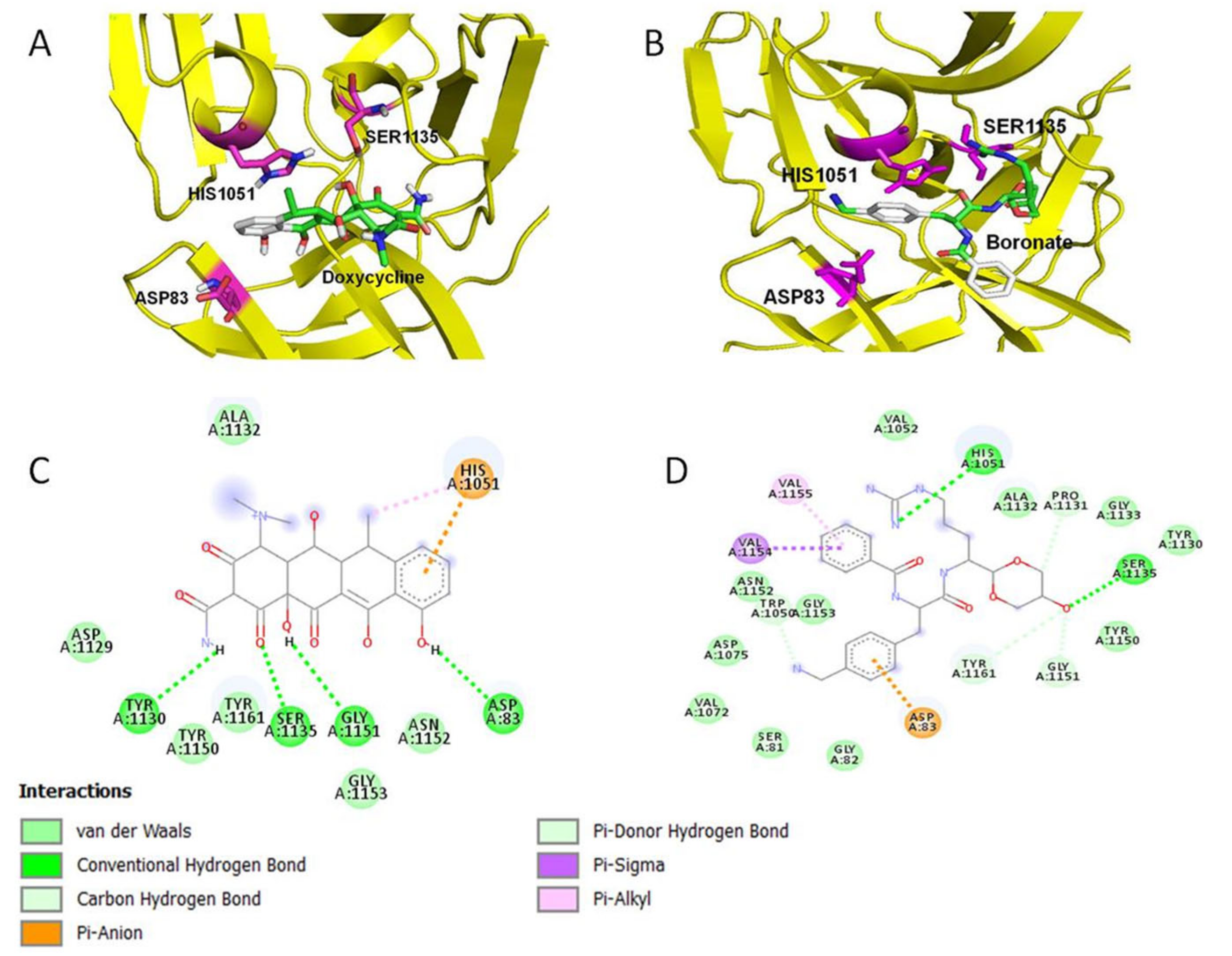
2.1. Doxycycline Binds to the Active Site of ZIKV NS2B-NS3 Protease
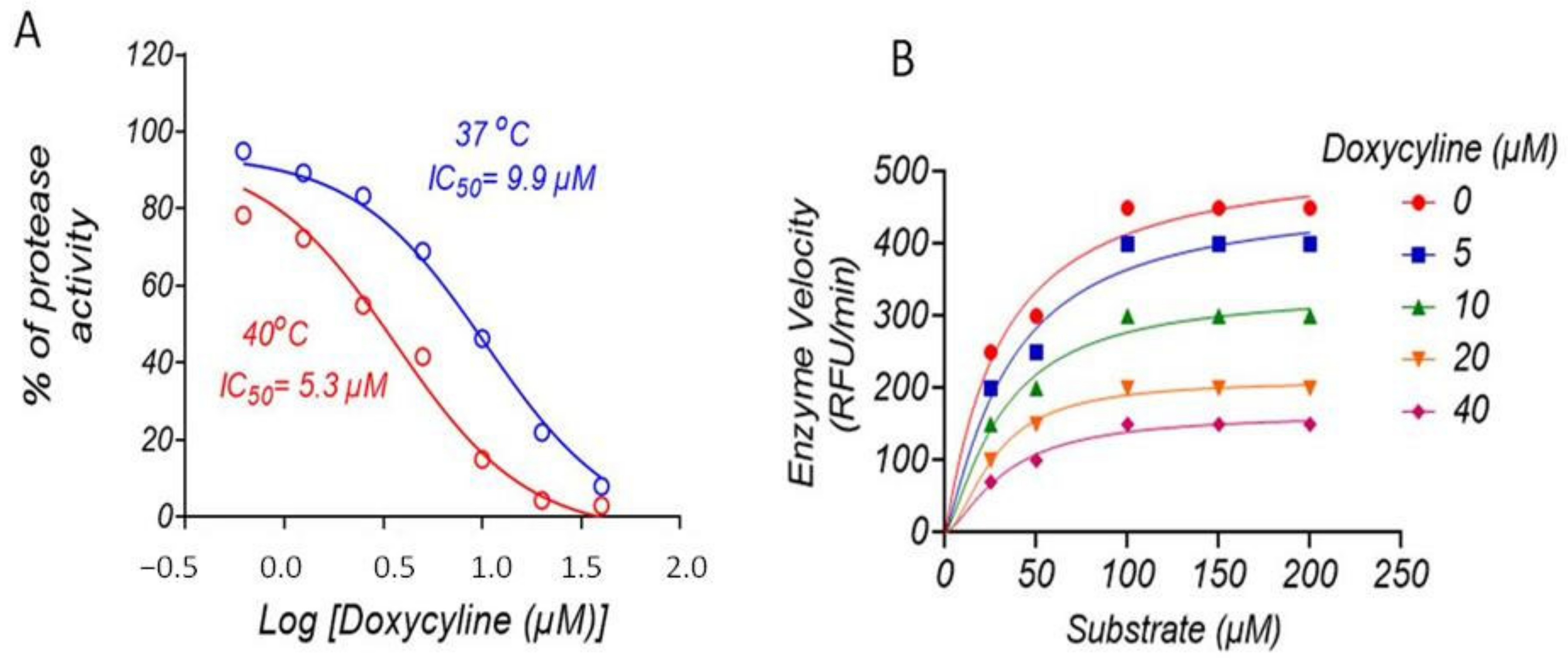
2.2. Doxycycline Inhibits ZIKV NS2B-NS3 Protease
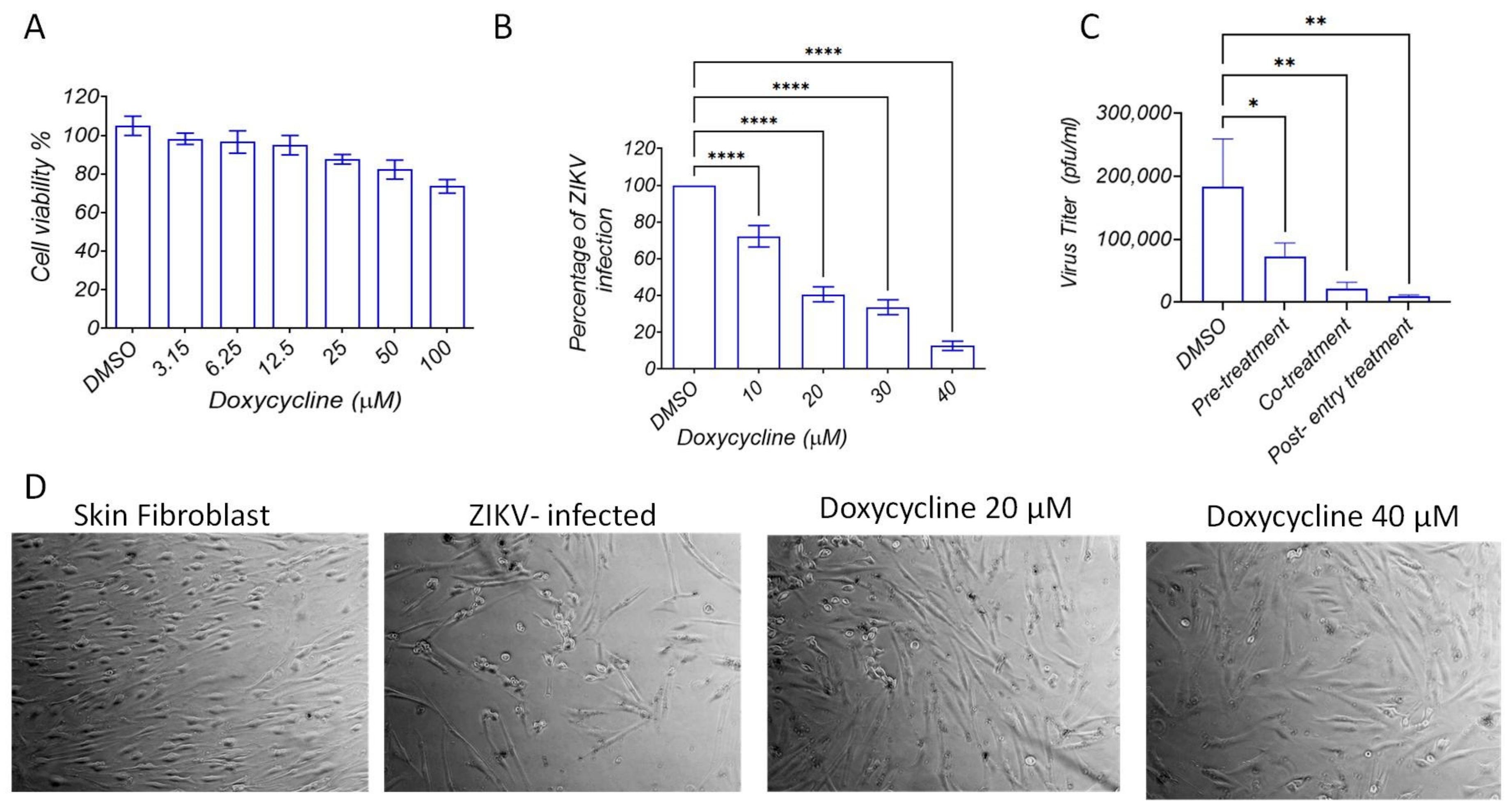
2.3. Doxycycline Inhibits ZIKV Entry and Replication in Human Dermal Fibroblasts
3. Discussion
4. Materials and Methods
4.1. The Interaction between the Doxycycline and ZIKV NS2B-NS3pro
4.2. Cells and Virus
4.3. Production and Purification of ZIKV NS2B-NS3 Protease
4.4. NS2B-NS3pro Protease Assay
4.5. Cytotoxicity Assay
4.6. Virus Infection and Quantification
4.7. Time-of-Addition Assay
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Petersen, L.R.; Jamieson, D.J.; Honein, M.A. Zika Virus. N. Engl. J. Med. 2016, 375, 294–295. [Google Scholar] [CrossRef]
- Dick, G.W. Zika virus. II. Pathogenicity and physical properties. Trans. R. Soc. Trop. Med. Hyg. 1952, 46, 521–534. [Google Scholar] [CrossRef]
- Rothan, H.A.; Bidokhti, M.R.M.; Byrareddy, S.N. Current concerns and perspectives on Zika virus co-infection with arboviruses and HIV. J. Autoimmun. 2018, 89, 11–20. [Google Scholar] [CrossRef]
- Simpson, D.I. Zika Virus Infection in Man. Trans. R. Soc. Trop. Med. Hyg. 1964, 58, 335–338. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Kosoy, O.O.; Laven, J.J.; Velez, J.O. Genetic and serologic properties of Zika virus associated with an epidemic, Yap State, Micronesia, 2007. Emerg. Infect. Dis. 2008, 14, 1232–1239. [Google Scholar] [CrossRef] [PubMed]
- Kuno, G.; Chang, G.J. Full-length sequencing and genomic characterization of Bagaza, Kedougou, and Zika viruses. Arch. Virol. 2007, 152, 687–696. [Google Scholar] [CrossRef]
- Hou, W.; Cruz-Cosme, R.; Armstrong, N.; Obwolo, L.A.; Wen, F.; Hu, W.; Luo, M.-H.; Tang, Q. Molecular cloning and characterization of the genes encoding the proteins of Zika virus. Gene 2017, 628, 117–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothan, H.A.; Abdulrahman, A.Y.; Sasikumer, P.G.; Othman, S.; Rahman, N.A.; Yusof, R. Protegrin-1 inhibits dengue NS2B-NS3 serine protease and viral replication in MK2 cells. J. Biomed. Biotechnol. 2012, 2012, 251482. [Google Scholar] [CrossRef] [PubMed]
- Rothan, H.A.; Han, H.C.; Ramasamy, T.S.; Othman, S.; Rahman, N.A.; Yusof, R. Inhibition of dengue NS2B-NS3 protease and viral replication in Vero cells by recombinant retrocyclin-1. BMC Infect. Dis. 2012, 12, 314. [Google Scholar] [CrossRef] [Green Version]
- Bessaud, M.; Pastorino, B.A.M.; Peyrefitte, C.N.; Rolland, D.; Grandadam, M.; Tolou, H.J. Functional characterization of the NS2B/NS3 protease complex from seven viruses belonging to different groups inside the genus Flavivirus. Virus Res. 2006, 120, 79–90. [Google Scholar] [CrossRef]
- Rothan, H.A.; Mohamed, Z.; Paydar, M.; Rahman, N.A.; Yusof, R. Inhibitory effect of doxycycline against dengue virus replication in vitro. Arch. Virol. 2014, 159, 711–718. [Google Scholar] [CrossRef]
- Hamel, R.; Dejarnac, O.; Wichit, S.; Ekchariyawat, P.; Neyret, A.; Luplertlop, N.; Perera-Lecoin, M.; Surasombatpattana, P.; Talignani, L.; Thomas, F. Biology of Zika Virus Infection in Human Skin Cells. J. Virol. 2015, 89, 8880–8896. [Google Scholar] [CrossRef] [Green Version]
- Buckley, A.; Gould, E.A. Detection of virus-specific antigen in the nuclei or nucleoli of cells infected with Zika or Langat virus. J. Gen. Virol. 1988, 69, 1913–1920. [Google Scholar] [CrossRef] [PubMed]
- Rothan, H.A.; Kumar, M. Role of Endoplasmic Reticulum-Associated Proteins in Flavivirus Replication and Assembly Complexes. Pathogens 2019, 8, 148. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Saucedo-Cuevas, L.; Yuan, L.; Ross, D.; Johansen, A.; Sands, D.; Stanley, V.; Guemex-Gamboa, A.; Gregor, A.; Evans, T. Zika Virus Protease Cleavage of Host Protein Septin-2 Mediates Mitotic Defects in Neural Progenitors. Neuron. 2019, 101, 1089–1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, D.; Schroder, K.; Fang, N.X.; Stoermer, M.J.; Abbenante, G.; Martin, J.L.; Young, P.R.; Fairlie, D.P. Activity of recombinant dengue 2 virus NS3 protease in the presence of a truncated NS2B co-factor, small peptide substrates, and inhibitors. J. Biol. Chem. 2001, 276, 45762–45771. [Google Scholar] [CrossRef] [Green Version]
- Rothan, H.A.; Buckle, M.J.; Ammar, Y.A.; Mohammadjavad, P.; Shatrah, O.; Noorsaadah, A.R.; Rohana, Y. Study the antiviral activity of some derivatives of tetracycline and non-steroid anti-inflammatory drugs towards dengue virus. Trop. Biomed. 2013, 30, 681–690. [Google Scholar]
- Topno, R.; Khan, S.A.; Chowdhury, P.; Mahanta, J. Pharmacodynamics of aminoglycosides and tetracycline derivatives against Japanese encephalitis virus. Asian Pac. J. Trop. Med. 2016, 9, 241–246. [Google Scholar] [CrossRef] [Green Version]
- Rothan, H.A.; Bahrani, H.; Mohamed, Z.; Teoh, T.C. A combination of doxycycline and ribavirin alleviated chikungunya infection. PLoS ONE 2015, 10, e0126360. [Google Scholar] [CrossRef] [Green Version]
- Sturtz, F.G. Antimurine retroviral effect of doxycycline. Methods Find. Exp. Clin. Pharmacol. 1998, 20, 643–647. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.C.; Wang, X.; Wei, J.-C.; Li, B.-B.; Shao, D.-H.; Li, Y.-M.; Liu, K.; Shi, Y.-Y.; Zhou, B.; Qiu, Y.-F. Antiviral activity of doxycycline against vesicular stomatitis virus in vitro. FEMS Microbiol. Lett. 2015, 362, fnv195. [Google Scholar]
- Krieger, E.; Koraimann, G.; Vriend, G. Increasing the precision of comparative models with YASARA NOVA--a self-parameterizing force field. Proteins. 2002, 47, 393–402. [Google Scholar] [CrossRef] [PubMed]
- De Vries, S.J.; van Dijk, M.; Bonvin, A.M. The HADDOCK web server for data-driven biomolecular docking. Nat. Protoc. 2010, 5, 883–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karaca, E.; Melquiond, A.S.J.; de Vries, S.; Kastritis, P.L.; Bonvin, A.M.J.J. Building macromolecular assemblies by information-driven docking: Introducing the HADDOCK multibody docking server. Mol. Cell Proteomics. 2010, 9, 1784–1794. [Google Scholar] [CrossRef] [Green Version]
- Abdulrahman, A.Y.; Khazali, A.S.; Teoh, T.C.; Rothan, H.A.; Yusof, R. Novel Peptides Inhibit Zika NS2B-NS3 Serine Protease and Virus Replication in Human Hepatic Cell Line. Int. J. Pept. Res. Ther. 2019, 25, 1659–1668. [Google Scholar] [CrossRef]
- Nor Rashid, N.; Teoh, T.C.; Al-Harbi, S.J.; Yusof, R.; Rothan, H.A. Evaluation of neutralizing antibodies produced by papaya mosaic virus nanoparticles fused to the E2EP3 peptide epitope of Chikungunya envelope. Trop. Biomed. 2021, 38, 36–41. [Google Scholar]
- Nor Rashid, N.; Yusof, R.; Rothan, H.A. Antiviral and virucidal activities of sulphated polysaccharides against Japanese encephalitis virus. Trop. Biomed. 2020, 37, 713–721. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Compound | Docked Energy Kcal/mol | H-Bond | vdw | Pi Interactions |
|---|---|---|---|---|
| Doxycycline | −7.8 | 4 | 6 | 2 |
| Boronate (reference) | −7.2 | 2 | 15 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chong Teoh, T.; J. Al-Harbi, S.; Abdulrahman, A.Y.; Rothan, H.A. Doxycycline Interferes with Zika Virus Serine Protease and Inhibits Virus Replication in Human Skin Fibroblasts. Molecules 2021, 26, 4321. https://doi.org/10.3390/molecules26144321
Chong Teoh T, J. Al-Harbi S, Abdulrahman AY, Rothan HA. Doxycycline Interferes with Zika Virus Serine Protease and Inhibits Virus Replication in Human Skin Fibroblasts. Molecules. 2021; 26(14):4321. https://doi.org/10.3390/molecules26144321
Chicago/Turabian StyleChong Teoh, Teow, Sawsam J. Al-Harbi, Ammar Yasir Abdulrahman, and Hussin A. Rothan. 2021. "Doxycycline Interferes with Zika Virus Serine Protease and Inhibits Virus Replication in Human Skin Fibroblasts" Molecules 26, no. 14: 4321. https://doi.org/10.3390/molecules26144321
APA StyleChong Teoh, T., J. Al-Harbi, S., Abdulrahman, A. Y., & Rothan, H. A. (2021). Doxycycline Interferes with Zika Virus Serine Protease and Inhibits Virus Replication in Human Skin Fibroblasts. Molecules, 26(14), 4321. https://doi.org/10.3390/molecules26144321