
Effect of Methyl Jasmonate on the Terpene Trilactones, Flavonoids, and Phenolic Acids in Ginkgo biloba L. Leaves: Relevance to Leaf Senescence

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
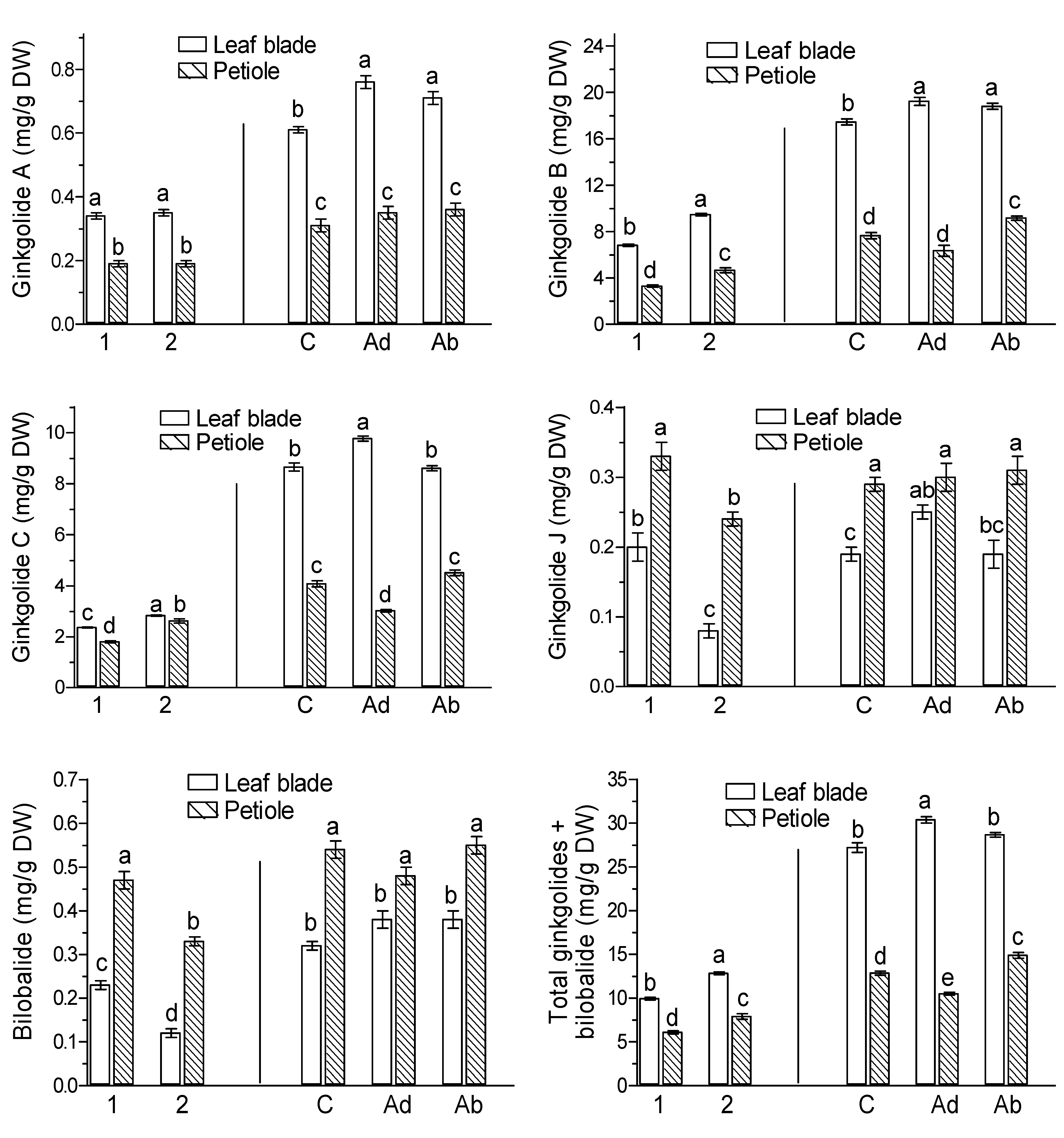
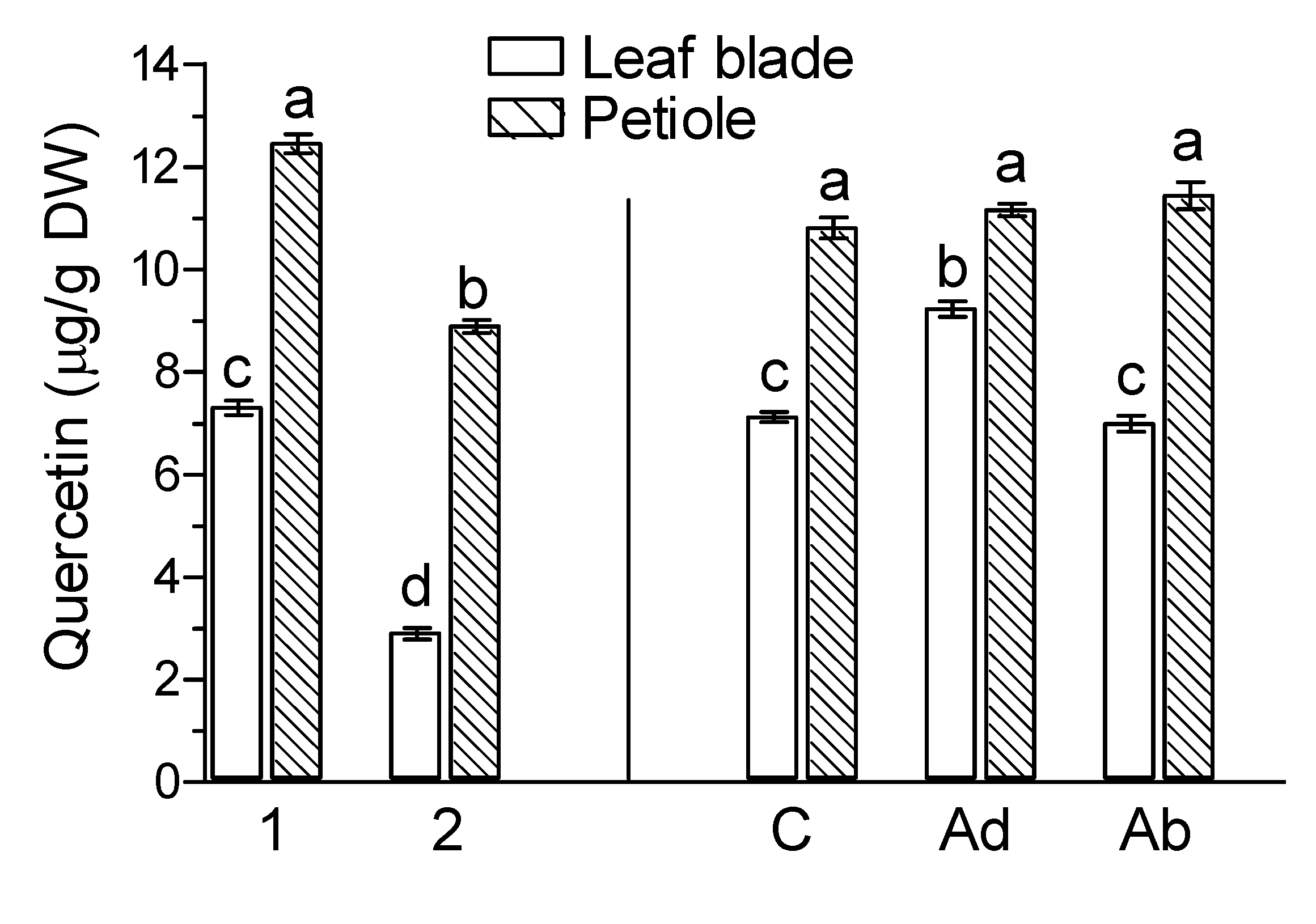
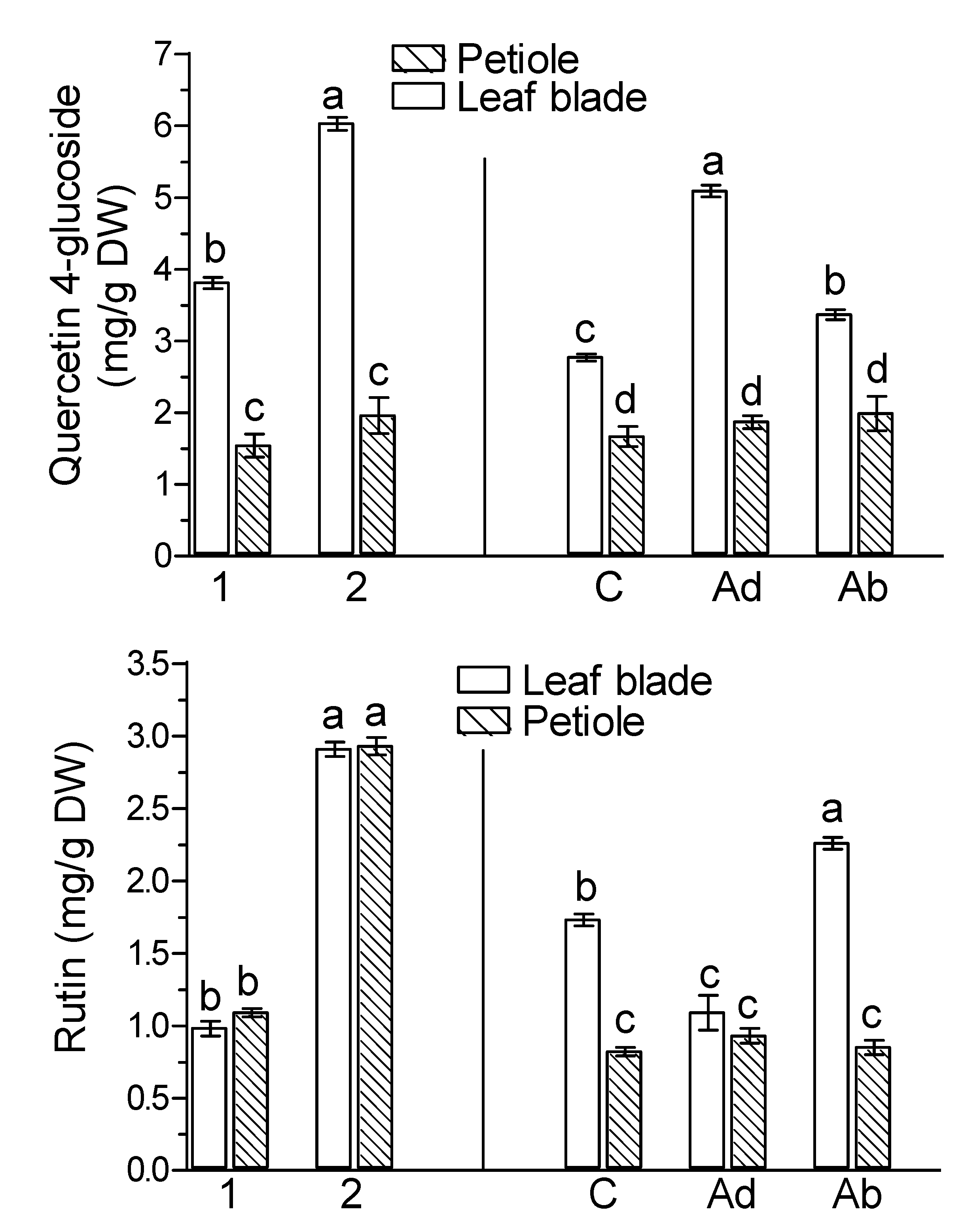
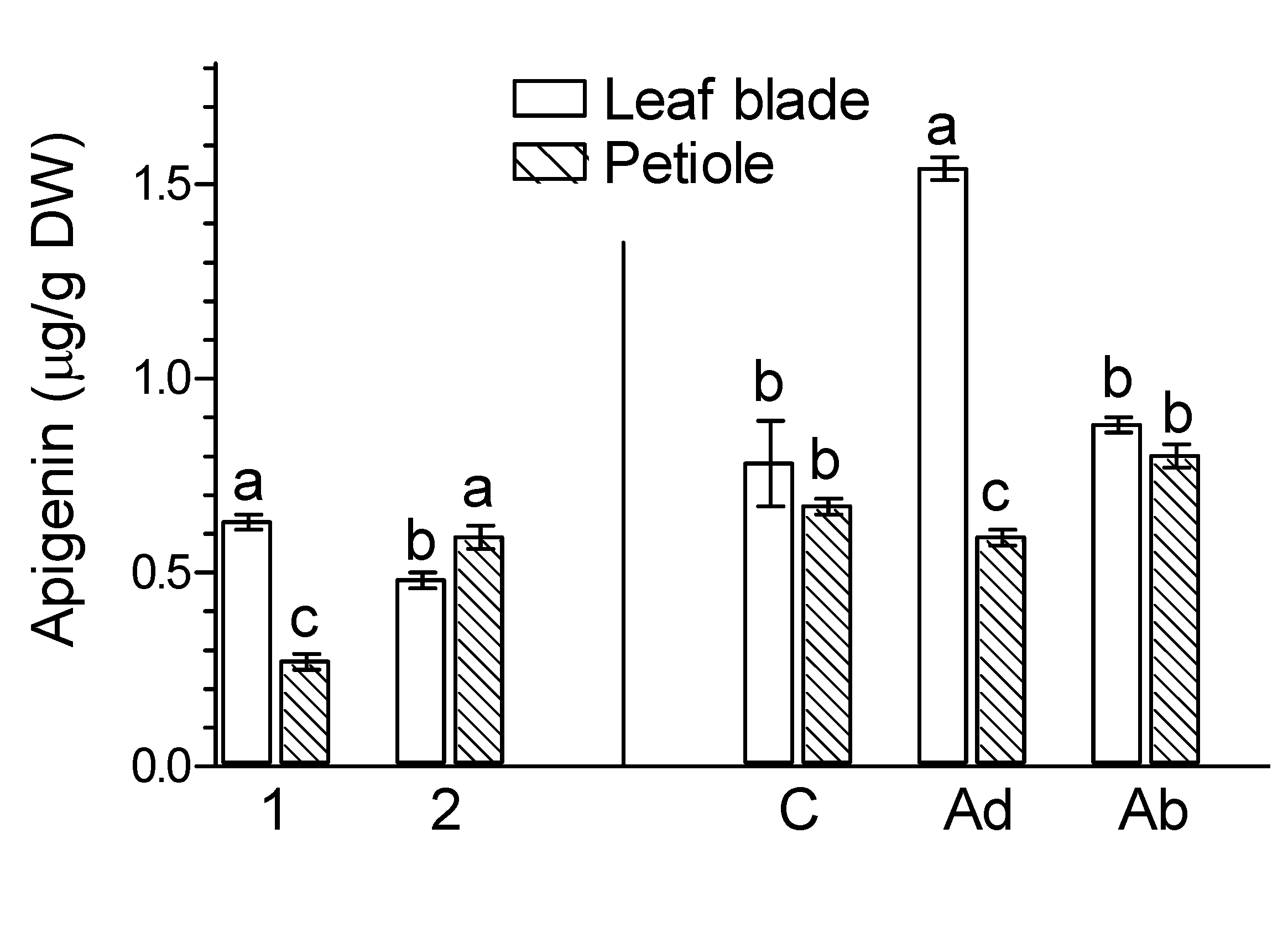
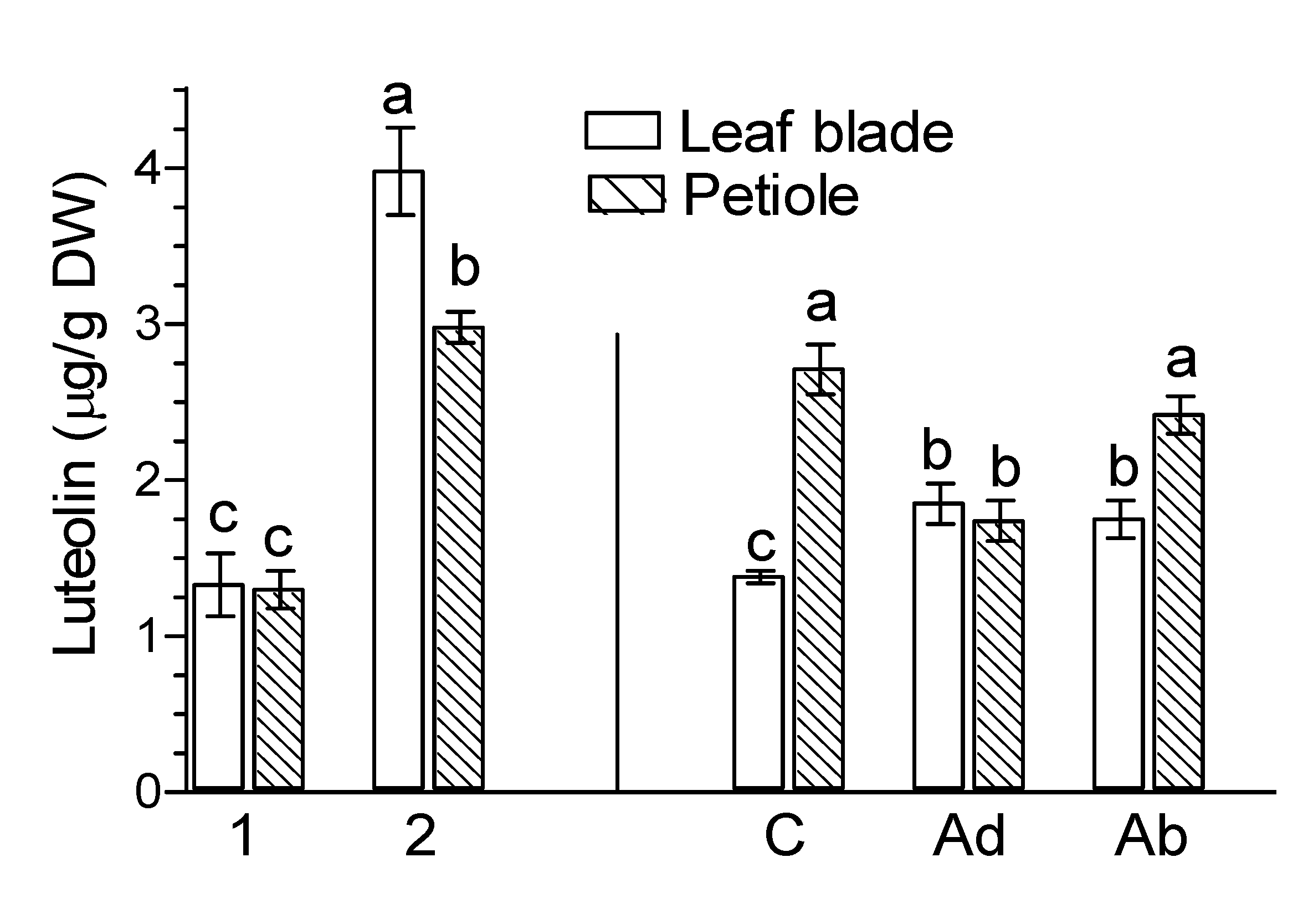
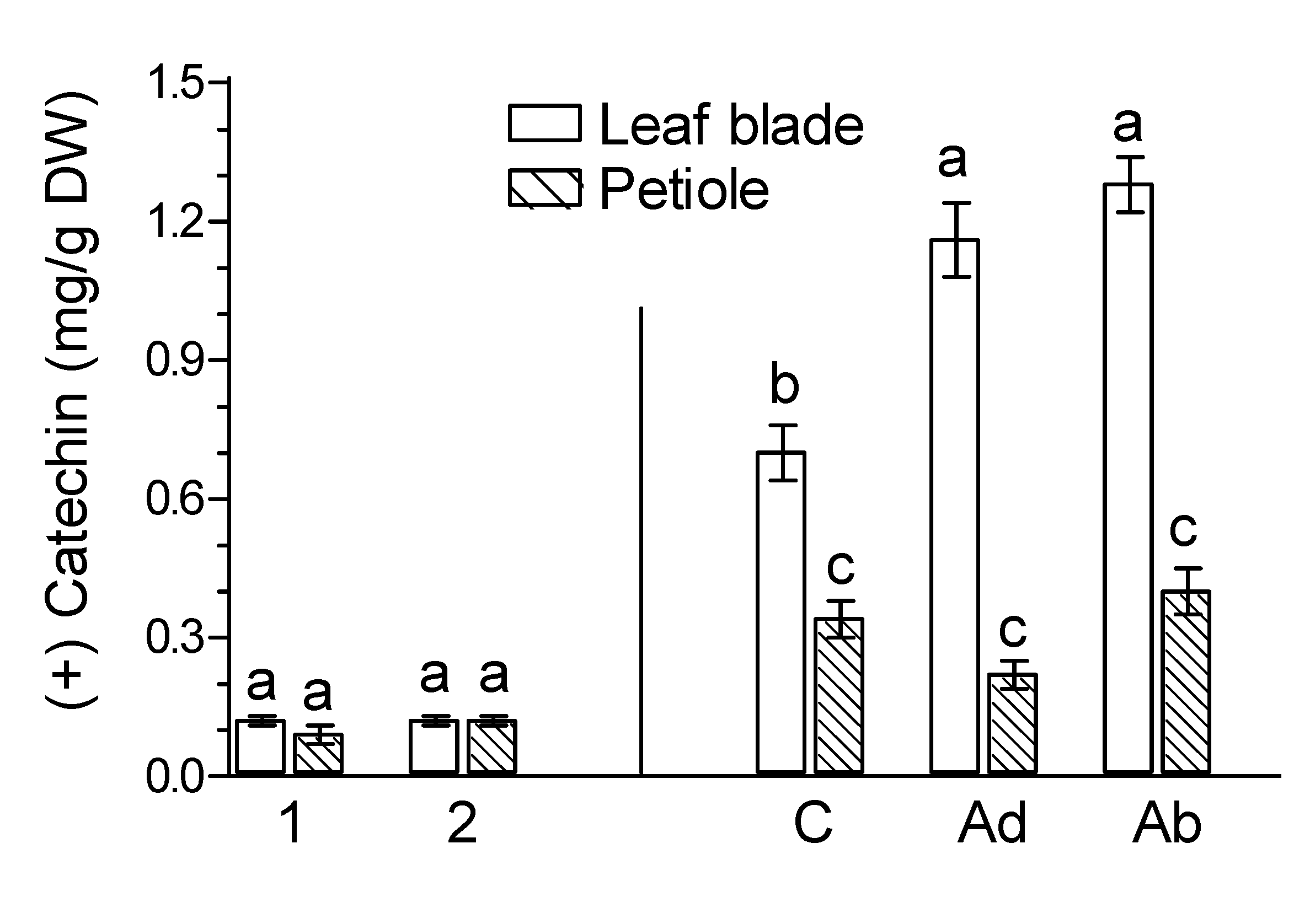
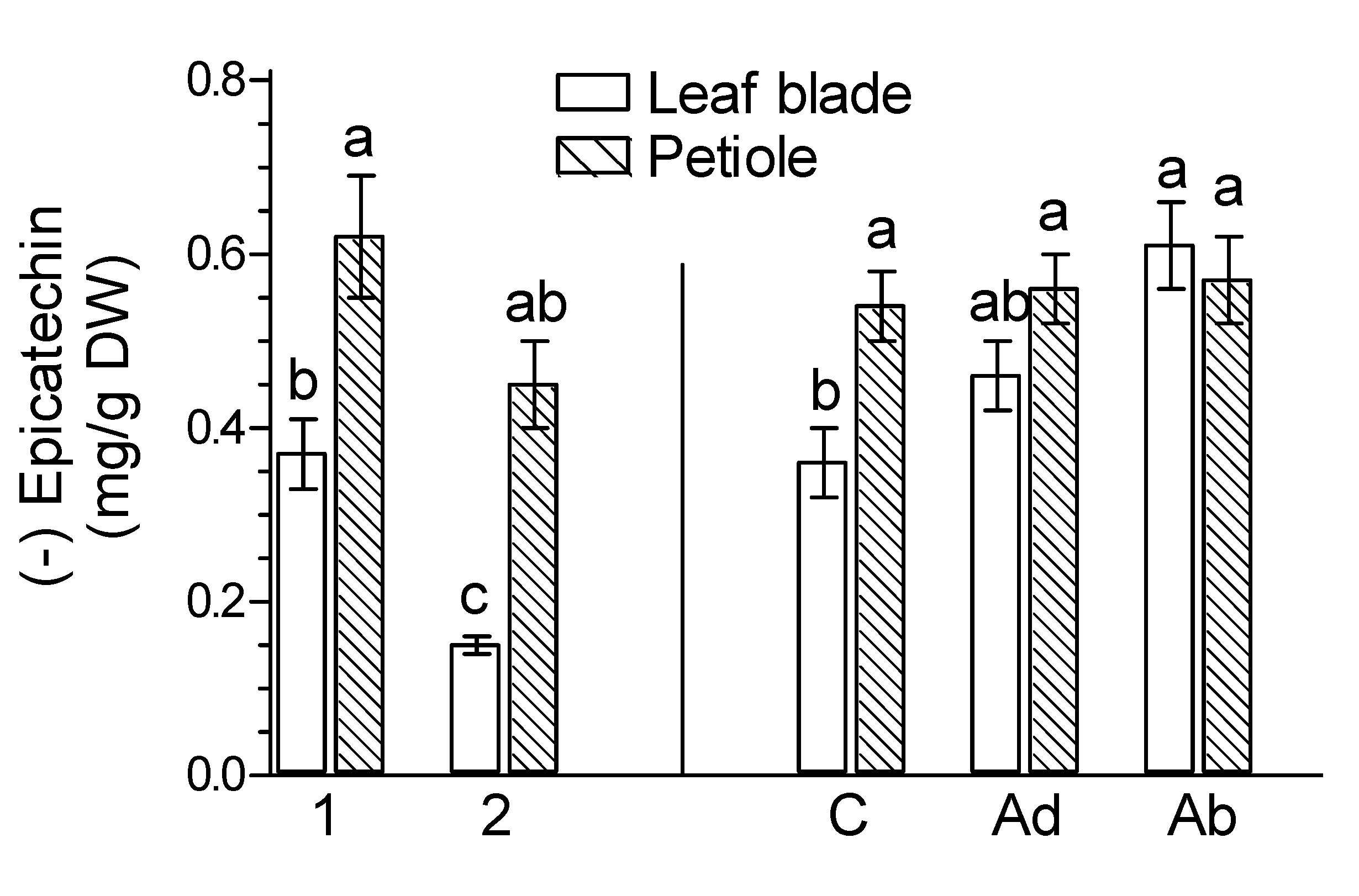
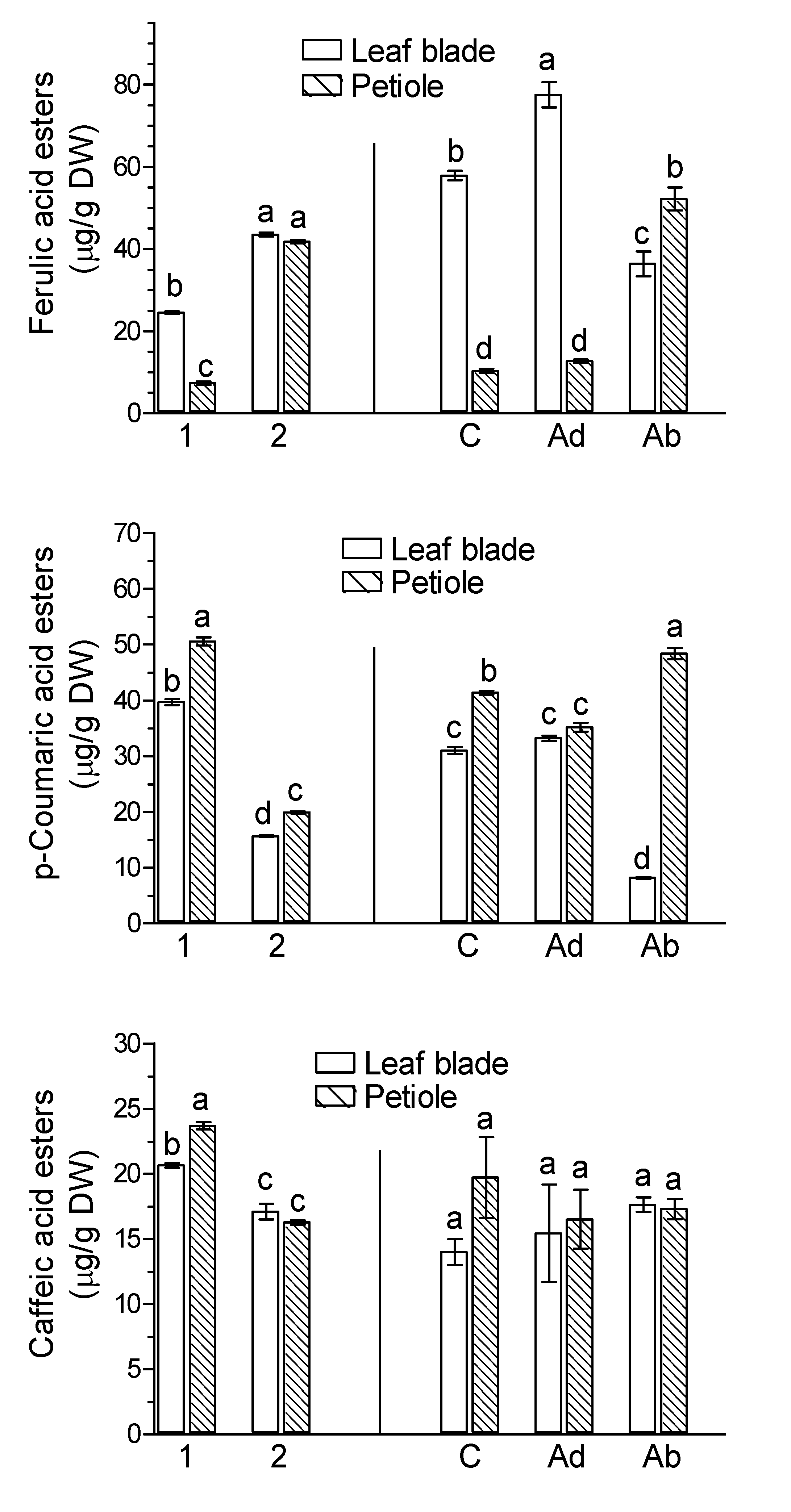
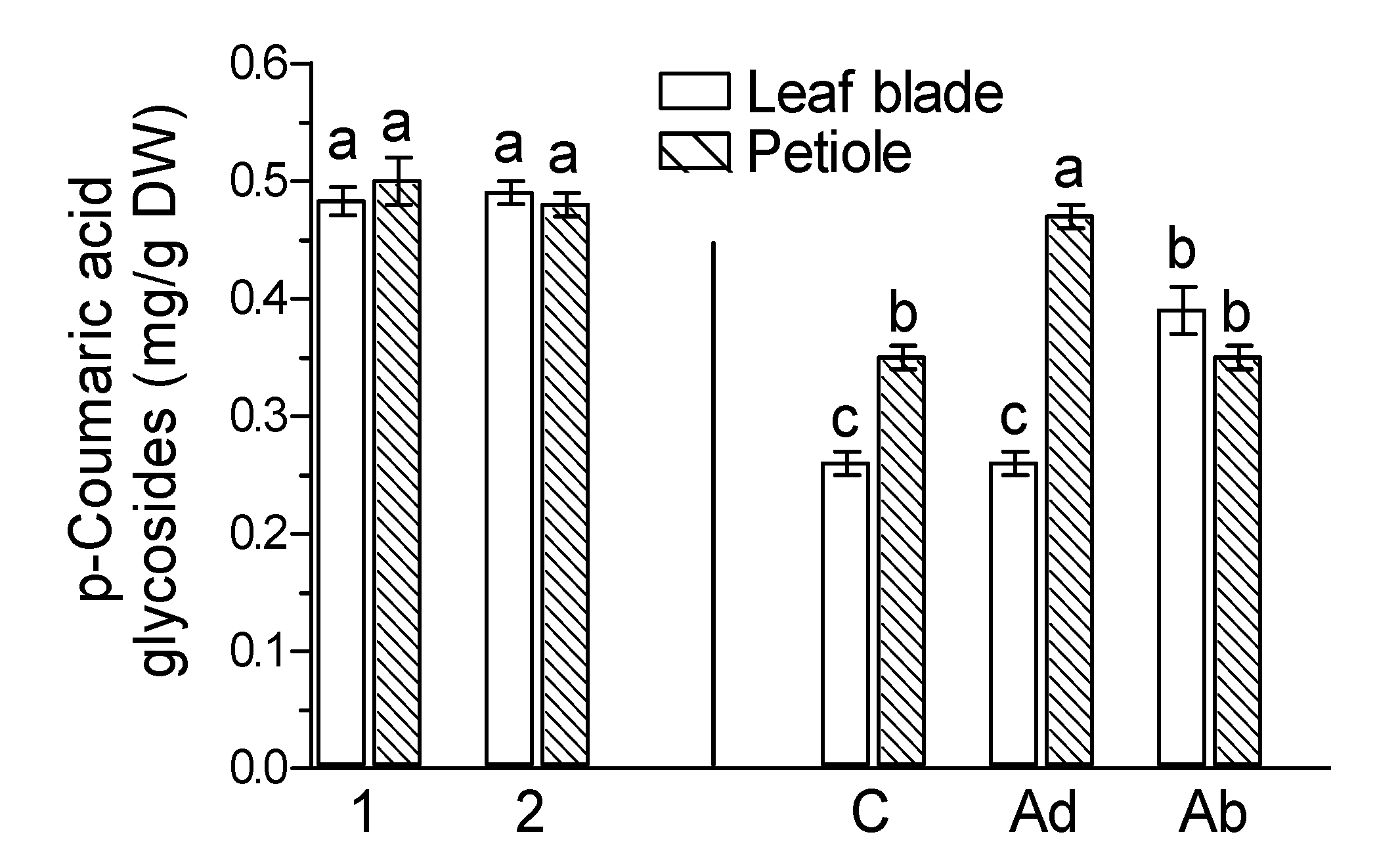
2. Results
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Analyses of Terpenoic Trilactones (Gingkolides, Bilobalide), Flavonoids, and Phenolic Acids
4.3. Statistical Analysis
5. Summary
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Dmuchowski, W.; Brągoszewska, P.; Gozdowski, D.; Baczewska-Dąbrowska, A.B.; Chojnacki, T.; Jozwiak, A.; Swiezewska, E.; Gworek, B.; Suwara, I. Strategy of Ginkgo biloba L. in the mitigation of salt stress in the urban environment. Urban. For. Urban Gree. 2019, 38, 223–231. [Google Scholar] [CrossRef]
- Ahlemeyer, B.; Krieglstein, J. Neuroprotective effects of Ginkgo biloba extract. Cell. Mol. Life Sci. 2003, 60, 1779–1792. [Google Scholar] [CrossRef] [PubMed]
- Hasler, A. Chemical constituents of Ginkgo biloba. In Ginkgo biloba; van Beek, T.A., Ed.; Harvard Academic: Cambridge, MA, USA, 2005; pp. 124–164. Available online: https://www.taylorfrancis.com/books/edit/10.1201/9780203304945/ginkgo-biloba-teris-vanbeek?refId=506d1be2-44da-476c-8b76-7b85b6d5bfbf (accessed on 1 August 2021).
- Strømgaard, K.; Nakanishi, K. Chemistry and biology of terpene trilactones from Ginkgo biloba. Angew. Chem. Int. Edit. 2004, 43, 1640–1658. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, K. Terpene trilactones from Gingko biloba: From ancient times to the 21st century. Bioorgan. Med. Chem. 2005, 13, 4987–5000. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Kaur, P.; Singh, G.R.D.; Ahuja, P.S. Biology and chemistry of Ginkgo biloba. Fitoterapia 2008, 79, 401–418. [Google Scholar] [CrossRef]
- Van Beek, T.A.; Montoro, P. Chemical analysis and quality control of Ginkgo biloba leaves, extracts, and phytopharmaceuticals. J. Chromatogr. A 2009, 1216, 2002–2032. [Google Scholar] [CrossRef]
- Cheng, S.Y.; Xu, F.; Wang, Y. Advances in the study of flavonoids in Ginkgo biloba leaves. J. Med. Plants Res. 2009, 3, 1248–1252. [Google Scholar]
- Xu, M.; Dong, J.; Wang, H.; Huang, L. Complementary action of jasmonic acid on salicylic acid in mediating fungal elicitor-induced flavonol glycoside accumulation of Ginkgo biloba cells. Plant Cell Environ. 2009, 32, 960–967. [Google Scholar] [CrossRef]
- Ražná, K.; Sawinska, Z.; Ivanišová, E.; Vukovic, N.; Terentieva, M.; Stričík, M.; Kowalczewski, P.Ł.; Hlavačková, L.; Rovná, K.; Žiarovská, J.; et al. Properties of Ginkgo biloba L.: Antioxidant characterization, antimicrobial activities, and genomic microRNA based marker fingerprints. Int. J. Mol. Sci. 2020, 21, 3087. [Google Scholar] [CrossRef]
- Ellnain-Wojtaszek, M.; Kruczynski, Z.; Kasprzak, J. Analysis of the content of flavonoids, phenolic acids as well as free radicals from Ginkgo bioloba L. leaves during the vegetative cycle. Acta Pol. Pharm. 2001, 58, 205–209. [Google Scholar]
- Sabater-Jara, A.B.; Souliman-Youssef, S.; Novo-Uzal, E.; Almagro, L.; Belchi-Navarro, S.; Pedreno, M.A. Biotechnological approaches to enhance the biosynthesis of ginkgolides and bilobalide in Ginkgo biloba. Phytochem. Res. 2013, 12, 191–205. [Google Scholar] [CrossRef]
- Wang, L.-T.; Fan, X.-H.; Jian, Y.; Dong, M.-Z.; Yang, Q.; Meng, D.; Fu, Y.-J. A sensitive multiple reactions monitoring mass spectrometry method for simultaneous quantification of flavonol glycoside, terpene lactones, and biflavonoids in Ginkgo biloba leaves. J. Pharm. Biomed. Anal. 2019, 170, 335–340. [Google Scholar] [CrossRef]
- Liu, X.-G.; Yang, H.; Cheng, X.-L.; Liu, L.; Qin, Y.; Wang, Q. Direct analysis of 18 flavonol glycosides, aglycones and terpene trilactones in Ginkgo biloba tablets by matrix soild phase dispersion coupled with ultra-high performance liquid chromatography tandem triple quadrupole mass spectrometry. J. Pharm. Biomed. Anal. 2014, 97, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Lou, K.; Wu, G.; Wu, X.; Zhou, X.; Feng, Y.; Zhang, H.; Yu, P. Bioactive metabolites in Ginkgo biloba leaves: Variations by seasonal, meteorological and soil. Braz. J. Biol. 2020, 80, 790–797. [Google Scholar] [CrossRef]
- van Beek, T.A.; Lelyveld, G.P. Concentration of ginkgolides and bilobalide in Ginkgo biloba leaves in relation to the time of year. Plant Med. 1992, 58, 413–416. [Google Scholar] [CrossRef]
- Rimkiene, L.; Ivanauskas, L.; Zevzikovas, A.; Nenortiene, P.; Jakstas, V. Variation of terpene lactones composition in Ginkgo biloba (L.) leaves under the influence of harvesting time and growing location. Acta Physiol. Plant. 2021, 43, 46. [Google Scholar] [CrossRef]
- Zheng, J.; Zhang, X.; Fu, M.; Zeng, H.; Ye, J.; Zhang, W.; Liao, Y.; Xu, F. Effects of different stress treatments on the total terpene trilactone content and expression levels of key genes in Ginkgo biloba leaves. Plant Mol. Biol. Rep. 2020, 38, 521–530. [Google Scholar] [CrossRef]
- Inoue, H.; Kamoda, S.; Terada, T.; Saburi, Y. Ginkgolide production in Ginkgo biloba trees and cultured cells. Bull. Tokyo Univ. For. 1999, 101, 253–258. [Google Scholar]
- Ueda, J.; Kato, J. Isolation and identification of a senescence-promoting substance from wormwood (Artemisia absinthium L.). Plant Physiol. 1980, 66, 246–249. [Google Scholar] [CrossRef] [Green Version]
- Ueda, J.; Kato, J. Identification of jasmonic acid and abscisic acid as senescence-promoting substances from Cleyera ochnacea DC. Agric. Biol. Chem. 1982, 46, 1975–1976. [Google Scholar] [CrossRef]
- Saniewski, M.; Dziurka, M.; Dziurka, K.; Góraj-Koniarska, J.; Ueda, J.; Miyamoto, K. Methyl jasmonate induces leaf senescence of Ginkgo biloba L.: Relevance to endogenous levels of plant hormones. Plant Growth Regul. 2020, 91, 383–396. [Google Scholar] [CrossRef]
- Wasternack, C. Jasmonates: An update on biosynthesis, signal transduction and action in plant stress response, growth and development. Ann. Bot. 2007, 100, 681–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasternack, C.; Hause, B. Jasmonates: Biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef] [PubMed]
- Jibran, R.; Hunter, D.A.; Dijkwel, P.P. Hormonal regulation of leaf senescence through integration of developmental and stress signals. Plant Mol. Biol. 2013, 82, 547–561. [Google Scholar] [CrossRef]
- Zhang, H.; Zhou, C. Signal transduction in leaf senescence. Plant Mol. Biol. 2013, 82, 539–545. [Google Scholar] [CrossRef]
- Huang, H.; Liu, B.; Liu, L.; Song, S. Jasmonate action in plant growth and development. J. Exp. Bot. 2017, 68, 1349–1359. [Google Scholar] [CrossRef] [Green Version]
- Wojciechowska, N.; Sobieszczuk-Nowicka, E.; Bagniewska-Nadworna, A. Plant organ senescence—Regulation by manifold pathways. Plant Biol. 2018, 20, 167–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Zhang, W.; Zhang, Y.; Lang, D. Zhang, X. The role of methyl jasmonate to stress in plants. Funct. Plant Biol. 2019, 46, 197–212. [Google Scholar] [CrossRef] [PubMed]
- Saniewski, M.; Ueda, J.; Miyamoto, K. Relationships between jasmonates and auxin in regulation of some physiological processes in higher plants. Acta Physiol. Plant. 2002, 24, 211–220. [Google Scholar] [CrossRef]
- Yang, J.; Duan, G.; Li, C.; Liu, L.; Han, G.; Sahang, Y.; Wang, C. The crosstalk between jasmonic acid and other plant hormone signaling highlight the involvement of jasmonic acid as a core component in plant response to biotic and abiotic stresses. Front. Plant Sci. 2019, 10, 1349. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.Y.; Bowman, L.; Ding, M. Methyl jasmonate enhances antioxidant activity and flavonoid content in blackberries (Rubus sp.) and promotes antiproliferation of human cancer cells. Food Chem. 2008, 107, 1261–1269. [Google Scholar] [CrossRef]
- Liu, H.; Timko, M.P. Jasmonic acid signaling and molecular crosstalk with other phytohormones. Int. J. Mol. Sci. 2021, 22, 2914. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.-Y.; Li, J.-X.; Yang, C.-Q.; Hu, W.L.; Wang, L.-J.; Chen, X.-Y. The jasmonate-responsive AP2/ERF transcription factors AaERF1 and AaERF2 positively regulate artemisinin biosynthesis in Artemisia annua L. Mol. Plant 2021, 5, 353–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, L.; Zhu, S.; Zhao, T.; Zhang, M.; Liu, W.; Chen, M.; Lam, X.; Liao, Z. Enhancement of artemisinin content and relative expression of genes of artemisinin biosynthesis in Artemisia annua by exogenous MeJA treatment. Plant Growth Regul. 2014, 75, 435–441. [Google Scholar] [CrossRef]
- Vélez-Bermúdez, I.-C.; Salazar-Henao, J.E.; Fornalé, S.; López-Vidriero, I.; Franco-Zorrilla, J.-M.; Grotewold, E.; Gray, J.; Solano, R.; Schmidt, W.; Pagés, M.; et al. A MYB/ZML Complex Regulates Wound-Induced Lignin Genes in Maize. Plant Cell 2015, 27, 3245–3259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onohata, T.; Gomi, K. Overexpression of jasmonate-responsive OsbHLH034 in rice results in the induction of bacterial blight resistance via an increase in lignin biosynthesis. Plant Cell Rep. 2020, 39, 1175–1184. [Google Scholar] [CrossRef]
- Kim, H.-J.; Fonseca, J.M.; Choi, J.-H.; Kubota, C. Effect of methyl jasmonate on phenolic compounds and carotenoids of Romaine lettuce (Lactuca sativa L.). J. Agric. Food Chem. 2008, 55, 10366–10372. [Google Scholar] [CrossRef] [PubMed]
- Flores, G.; Blanch, G.P.; Ruiz de Castillo, M.L. Effect of postharvest methyl jasmonate treatment on fatty acid composition and phenolic acid content in olive fruits during storage. J. Sci. Food Agric. 2017, 97, 2767–2772. [Google Scholar] [CrossRef]
- Shi, D.; Wei, X.; Chen, G.; Hu, Y. Changes in photosynthetic characteristics and antioxidantive protection in male and female Ginkgo during natural senescence. J. Am. Soc. Hort. Sci. 2012, 137, 349–360. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Jiang, H.; Yu, S.Q.; Zhou, G.M. Sex-linked photosynthetic physiologic research and evolutionary ecological analysis in living fossil plant, Ginkgo biloba L. Acta Ecol. Sin. 2008, 28, 1128–1136. [Google Scholar] [CrossRef]
- Zhang, X.; Zhu, J.; Cheng, S.; Zhang, W.; Xu, F.; Liao, Y. Effect of exogenous gibberellins on endogenous hormone and ginkgolide content on Ginkgo biloba leaves. Not. Bot. Horti. Agrobo. 2020, 48, 140–149. [Google Scholar] [CrossRef] [Green Version]
- Gülz, P.-G.; Müller, E.; Schmitz, K.; Marner, F.-J.; Güth, S. Chemical composition and surface structures of epicuticular leaf waxes of Ginkgo biloba, Magnolia grandiflora and Liliodendron tulipifera. Z. Naturforsch. C 1992, 47, 516–526. [Google Scholar] [CrossRef]
- Cheng, S.; Zhang, W.; Sun, N.; Xu, F.; Li, L.; Liao, Y.; Cheng, H. Production of flavonoids and terpene lactones from optimized Ginkgo biloba tissue culture. Not. Bot. Horti. Agrobo. 2014, 42, 88–93. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.-M.; Min, J.-Y.; Kim, Y.-D.; Park, D.-J.; Jung, H.-N.; Kim, S.-W.; Choi, M.-S. Effects of methyl jasmonate and salicylic acid on the production of bilobalide and ginkgolides in cell cultures of Ginkgo biloba. In Vitro Cell. Dev. Biol. Plant 2006, 42, 44–49. [Google Scholar] [CrossRef]
- Sukito, A.; Tachibana, S. Effect of methyl jasmonate and salicylic acid synergism on enhancement of bilobalide and ginkgolide by immobilized cell cultures of Ginkgo biloba. Bioresour. Bioprocess. 2016, 3, 24. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Xu, F.; Huang, X.; Zhang, W.; Cheng, H.; Li, L.; Cheng, S.; Shen, Y. Promoter analysis and transcriptional profiling of Ginkgo biloba 3-hydroxy-3-methylglutaryl coenzyme A reductase (GbHMGR) gene in abiotic stress responses. Not. Bot. Horti. Agrobo. 2015, 43, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Rao, S.; Meng, X.; Liao, Y.; Yu, T.; Cao, J.; Tan, J.; Xu, F.; Cheng, S. Characterization and functional analysis of two novel 3-hydroxy-3-methylglutaryl-coenzyme A reductase genes (GbHMGR2 and GbHMGR3) from Ginkgo biloba. Sci. Rep. 2019, 9, 14109. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Wang, L.; He, Z.; Lu, Z.; Cui, J.; Xu, N.; Jin, B.; Wang, L. Physiological and transcriptomic changes during autumn coloration and senescence in Ginkgo biloba leaves. Hortic. Plant J. 2020, 6, 396–408. [Google Scholar] [CrossRef]
- Ye, J.; Mao, D.; Cheng, S.; Zhang, X.; Tan, J.; Zheng, J.; Xu, F. Comparative transcriptome analysis reveals the potential stimulatory mechanism of terpene trilactone biosynthesis by exogenous salicylic acid in Ginkgo biloba. Ind. Crop Prod. 2020, 145, 112104. [Google Scholar] [CrossRef]
- Jung, J.H.G.; Himmelsbach, D.S. Isolation and characterization of wheat straw lignin. J. Agric. Food Chem. 1989, 37, 81–87. [Google Scholar] [CrossRef]
- Fry, S.C. Feruloylated pectins from the primary cell wall: Their structure and possible functions. Planta 1983, 157, 111–123. [Google Scholar] [CrossRef] [PubMed]
- de Buanafina, O.; Marcia, M. Feruloylation in grasses: Current and future perspectives. Mol. Plant 2009, 2, 861–872. [Google Scholar] [CrossRef]
- Mathew, S.; Abraham, T.E. Ferulic acid: An antioxidant found naturally in plant cell wall and feruloyl esterases involved in its release and their applications. Crit. Rev. Biotechnol. 2004, 24, 59–83. [Google Scholar] [CrossRef] [PubMed]
- Marchiosi, R.; dos Santos, W.D.; Constantin, R.P.; de Lima, R.B.; Soares, A.R.; Finger-Teixeira, A.; Mota, T.R.; de Oliveira, D.M.; Foletto-Felipe, M.P.; Abrahão, J.; et al. Biosynthesis and metabolic actions of simple phenolic acids in plants. Phytochem. Rev. 2020, 19, 865–906. [Google Scholar] [CrossRef]
- Kondo, T.; Mizuno, K.; Kato, T. Cell wall-bound p-coumaric acid and ferulic acids in Italian ryegrass. Can. J. Plant Sci. 1990, 71, 495–499. [Google Scholar] [CrossRef]
- Mnich, E.; Bjarnholt, N.; Eudes, A.; Harholt, J.; Holland, C.; Jørgensen, B.; Larsen, F.H.; Liu, M.; Manat, R.; Meyer, A.S.; et al. Phenolic cross-links: Building and de-constructing the plant cell wall. Nat. Prod. Rep. 2020, 37, 919–961. [Google Scholar] [CrossRef] [PubMed]
- Harris, P.J.; Trethewey, J.A.K. The distribution of ester-linked ferulic acid in the cell walls of angiosperms. Phytochem. Environ. 2010, 9, 19–33. [Google Scholar] [CrossRef]
- Wakabayashi, K.; Hoson, T.; Kamisaka, S. Osmotic stress suppresses cell wall stiffening and the increase in cell wall-bound ferulic and diferulic acids in wheat coleoptiles. Plant Physiol. 1997, 113, 967–973. [Google Scholar] [CrossRef] [Green Version]
- Kamisaka, S.; Takeda, S.; Takahashi, K.; Shibata, K. Diferulic and ferulic acid in the cell wall of Avena coleoptiles: Their relationships to mechanical properties of the cell wall. Physiol. Plant. 1990, 78, 1–7. [Google Scholar] [CrossRef]
- Hura, T.; Dziurka, M.; Hura, K.; Ostrowska, A.; Dziurka, K. Different allocation of carbohydrates and phenolics in dehydrated leaves of triticale. J. Plant Physiol. 2016, 202, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Jahangir, M.; Abdel-Farid, I.B.; Kim, H.K.; Choi, Y.H.; Verpoorte, R. Healthy and unhealthy plants: The effect of stress on the metabolism of Brassicaceae. Environ. Exp. Bot. 2009, 67, 23–33. [Google Scholar] [CrossRef]
- Sati, P.; Pandey, A. Influence of age on Ginkgo Biloba phytochemicals in antimicrobial activity perspective. J. Graph. Era Univ. 2016, 4, 57–65. [Google Scholar]
- Perez-Balibrea, S.; Moreno, D.A.; Garcia-Viguera, C. Genotypic effects on the phytochemical quality of seeds and sprouts from commercial broccoli cultivars. Food Chem. 2011, 125, 348–354. [Google Scholar] [CrossRef]
- Ross, K.A.; Beta, T.; Arntfield, S.D. A comparative study of phenolic acids identified and quantified in dry beans using HPLC as affected by different extraction and hydrolysis methods. Food Chem. 2009, 113, 336–344. [Google Scholar] [CrossRef]
- Kivilompolo, M.; Hyotylainen, T. On-line coupled dynamic sonication-assisted extraction-liquid chromatography for the determination of phenolic acids in Lamiaceae herbs. J. Chrom. A 2009, 1216, 892–896. [Google Scholar] [CrossRef] [PubMed]
- Płatosz, N.; Sawicki, T.; Wiczkowski, W. Profile of phenolic acids and flavonoids of red beet and its fermentation products. Does long-term consumption of fermented beetroot juice affect phenolics profile in human blood plasma and urine? Pol. J. Food Nutr. Sci. 2020, 70, 55–65. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horbowicz, M.; Wiczkowski, W.; Góraj-Koniarska, J.; Miyamoto, K.; Ueda, J.; Saniewski, M. Effect of Methyl Jasmonate on the Terpene Trilactones, Flavonoids, and Phenolic Acids in Ginkgo biloba L. Leaves: Relevance to Leaf Senescence. Molecules 2021, 26, 4682. https://doi.org/10.3390/molecules26154682
Horbowicz M, Wiczkowski W, Góraj-Koniarska J, Miyamoto K, Ueda J, Saniewski M. Effect of Methyl Jasmonate on the Terpene Trilactones, Flavonoids, and Phenolic Acids in Ginkgo biloba L. Leaves: Relevance to Leaf Senescence. Molecules. 2021; 26(15):4682. https://doi.org/10.3390/molecules26154682
Chicago/Turabian StyleHorbowicz, Marcin, Wiesław Wiczkowski, Justyna Góraj-Koniarska, Kensuke Miyamoto, Junichi Ueda, and Marian Saniewski. 2021. "Effect of Methyl Jasmonate on the Terpene Trilactones, Flavonoids, and Phenolic Acids in Ginkgo biloba L. Leaves: Relevance to Leaf Senescence" Molecules 26, no. 15: 4682. https://doi.org/10.3390/molecules26154682
APA StyleHorbowicz, M., Wiczkowski, W., Góraj-Koniarska, J., Miyamoto, K., Ueda, J., & Saniewski, M. (2021). Effect of Methyl Jasmonate on the Terpene Trilactones, Flavonoids, and Phenolic Acids in Ginkgo biloba L. Leaves: Relevance to Leaf Senescence. Molecules, 26(15), 4682. https://doi.org/10.3390/molecules26154682