
Rhodococcus as Biofactories for Microbial Oil Production

 ,
,
Abstract
:1. Introduction
2. Biological Properties of Oleaginous Rhodococci
3. Comparison of Rhodococci to Other Oleaginous Microorganisms
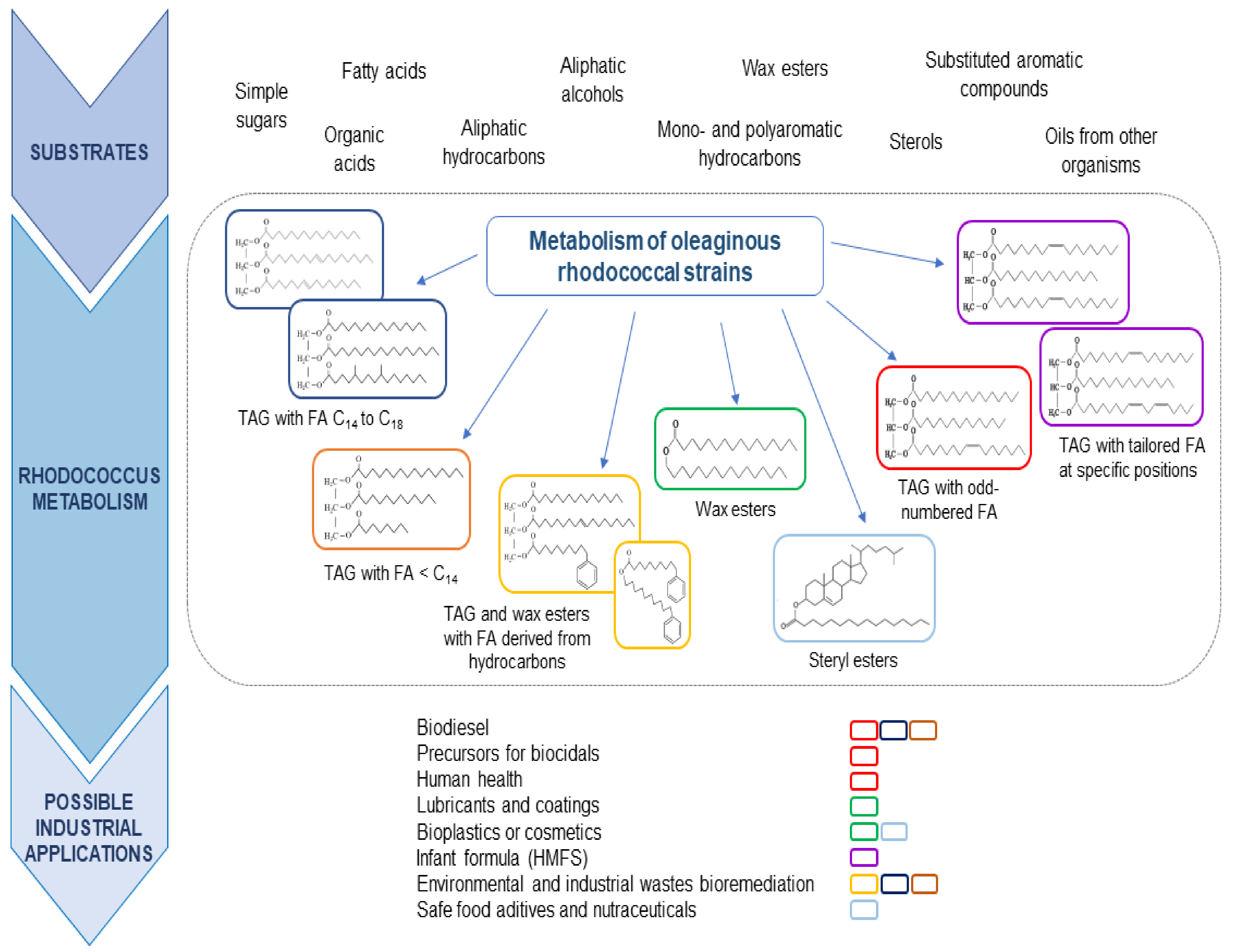
4. Catabolism of Different Carbon Sources and Lipogenesis Pathways Are Coupled in Oleaginous Rhodococci
4.1. Sugars and Organic Acids as Carbon Sources
4.2. Aliphatic Hydrocarbons
4.3. Alcohols
4.4. Aromatic Compounds
5. Conversion of Industrial Wastes into Microbial Oils by Rhodococci
6. Engineering Approaches to Improve Lipid Accumulation in Oleaginous Rhodococcus
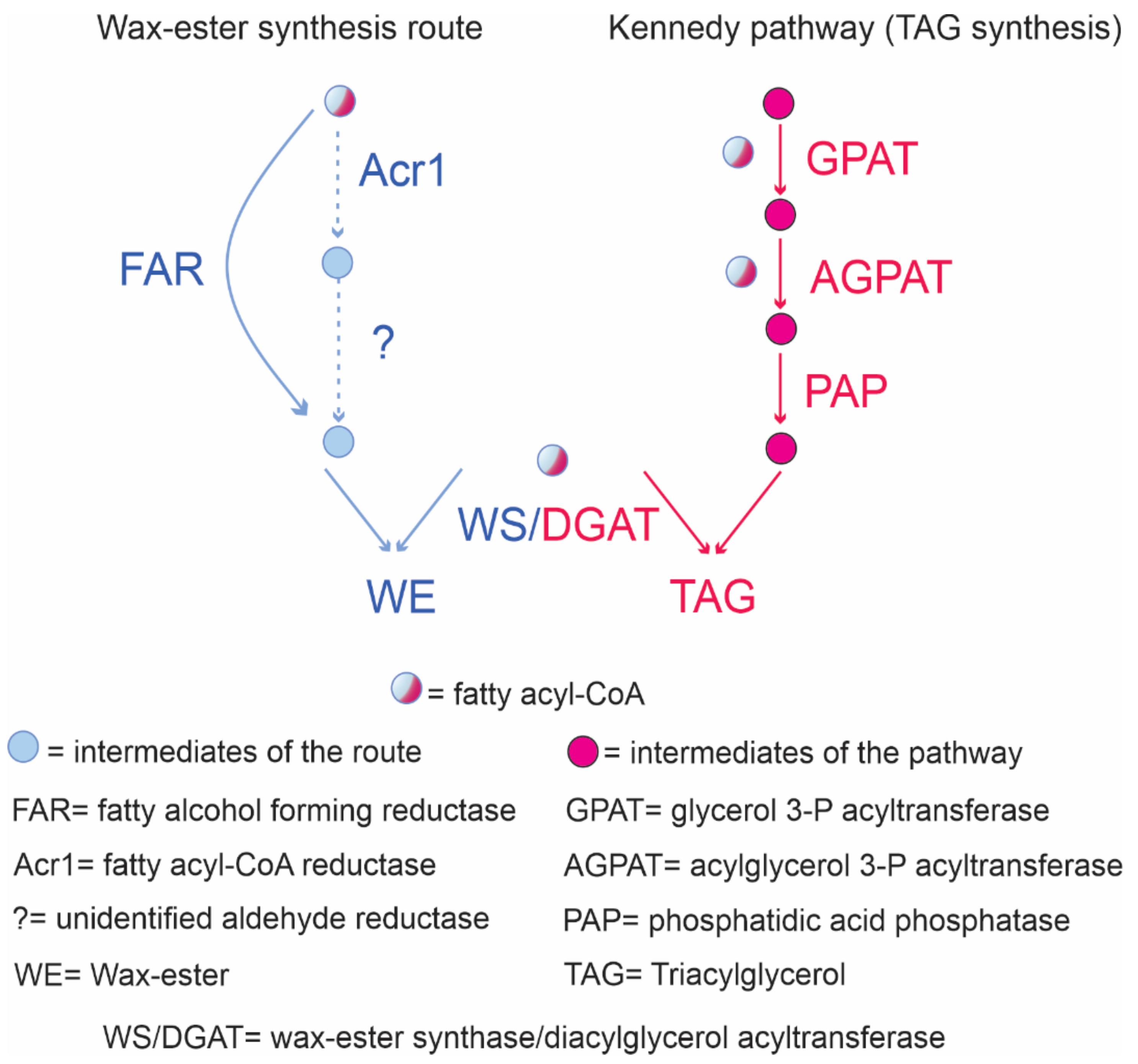
7. Wax Ester Biosynthesis in Oleaginous Rhodococci
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Ratledge, C.; Wynn, J.P. Oils from Microorganisms. In Bailey’s Industrial Oil and Fat Products; Shahidi, F., Ed.; John Wiley and Sons, Ltd.: Hoboken, NJ, USA, 2020; pp. 1–34. [Google Scholar] [CrossRef]
- Spagnuolo, M.; Yaguchi, A.; Blenner, M. Oleaginous yeast for biofuel and oleochemical production. Curr. Opin. Biotechnol. 2019, 57, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, H.M.; Herrero, O.M.; Lanfranconi, M.P.; Silva, R.A.; Villalba, M.S. Biotechnological production and significance of triacylglycerols and wax esters. In Consequences of Microbial Interactions with Hydrocarbons, Oils, and Lipids: Production of Fuels and Chemicals, Handbook of Hydrocarbon and Lipid Microbiology; Lee, S.Y., Ed.; Springer: Heidelberg, Germany, 2017. [Google Scholar] [CrossRef]
- Olukoshi, E.R.; Packter, N.M. Importance of stored triacylglycerols in Streptomyces: Possible carbon source for antibiotics. Microbiology 1994, 140, 931–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, H.M.; Steinbüchel, A. Triacylglycerols in prokaryotic microorganisms. Appl. Microbiol. Biotechnol. 2002, 60, 367–376. [Google Scholar] [CrossRef]
- Bredemeier, R.; Hulsch, R.; Metzger, J.O.; Berthe-Corti, L. Submersed culture production of extracellular wax esters by the marine bacterium Fundibacter jadensis. Mar. Biotechnol. 2003, 5, 579–583. [Google Scholar] [CrossRef]
- Röttig, A.; Hauschild, P.; Madkour, M.H.; Al-Ansari, A.M.; Almakishah, N.H.; Steinbüchel, A. Analysis and optimization of triacylglycerol synthesis in novel oleaginous Rhodococcus and Streptomyces strains isolated from desert soil. J. Biotechnol. 2016, 225, 48–56. [Google Scholar] [CrossRef]
- Alvarez, H.M.; Mayer, F.; Fabritius, D.; Steinbüchel, A. Formation of intracytoplasmic lipid inclusion by Rhodococcus opacus PD630. Arch. Microbiol. 1996, 165, 377–386. [Google Scholar] [CrossRef]
- Hernández, M.A.; Mohn, W.W.; Martínez, E.; Rost, E.; Alvarez, A.F.; Alvarez, H.M. Biosynthesis of storage compounds by Rhodococcus jostii RHA1 and global identification of genes involved in their metabolism. BMC Genom. 2008, 12, 600. [Google Scholar] [CrossRef] [Green Version]
- Sangal, V.; Goodfellow, M.; Jones, A.L.; Schwalbe, E.C.; Blom, J.; Hoskisson, P.A.; Sutcliffe, I.C. Next-generation systematics: An innovative approach to resolve the structure of complex prokaryotic taxa. Sci. Rep. 2016, 6, 38392. [Google Scholar] [CrossRef] [Green Version]
- Juarez, A.; Villa, J.A.; Lanza, V.F.; Lázaro, B.; de la Cruz, F.; Alvarez, H.M.; Moncalián, G. Nutrient starvation leading to triglyceride accumulation activates the Entner Doudoroff pathway in Rhodococcus jostii RHA1. Microb. Cell Factories 2017, 16, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Dávila Costa, J.S.; Herrero, O.M.; Alvarez, H.M.; Leichert, L. Label-free and redox proteomic analyses of the triacylglycerol-accumulating Rhodococcus jostii RHA1. Microbiology 2015, 161, 593–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacEachran, D.P.; Sinskey, A.J. The Rhodococcus opacus TadD protein mediates triacylglycerol metabolism by regulating intracellular NAD (P) H pools. Microb. Cell Factories 2013, 12, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández, M.A.; Alvarez, H.M. Increasing lipid production using an NADP+-dependent malic enzyme from Rhodococcus jostii. Microbiology 2019, 165, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Costa, J.S.D.; Silva, R.A.; Leichert, L.; Alvarez, H.M. Proteome analysis reveals differential expression of proteins involved in triacylglycerol accumulation by Rhodococcus jostii RHA1 after addition of methyl viologen. Microbiology 2017, 163, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Amara, S.; Seghezzi, N.; Otani, H.; Diaz-Salazar, C.; Liu, J.; Eltis, L.D. Characterization of key triacylglycerol biosynthesis processes in rhodococci. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Cereijo, A.E.; Asencion Diez, M.D.; Dávila Costa, J.S.; Alvarez, H.M.; Iglesias, A.A. On the Kinetic and Allosteric Regulatory Properties of the ADP-Glucose Pyrophosphorylase from Rhodococcus jostii: An Approach to Evaluate Glycogen Metabolism in Oleaginous Bacteria. Front. Microbiol. 2016, 7, 830. [Google Scholar] [CrossRef] [PubMed]
- Cereijo, A.E.; Alvarez, H.M.; Iglesias, A.A.; Asencion Diez, M.D. Glucosamine-P and rhodococcal ADP-glucose pyrophosphorylases: A hint to (re)discover (actino)bacterial amino sugar metabolism. Biochimie 2020, 176, 158–161. [Google Scholar] [CrossRef] [PubMed]
- Cereijo, A.E.; Kuhn, M.L.; Hernández, M.A.; Ballicora, M.A.; Iglesias, A.A.; Alvarez, H.M.; Asencion Diez, M.D. Study of duplicated galU genes in Rhodococcus jostii and a putative new metabolic node for glucosamine-1P in rhodococci. Biochim. Biophys. Acta Gen. Subj. 2021, 1865, 129727. [Google Scholar] [CrossRef]
- Hernández, M.A.; Lara, J.; Gago, G.; Gramajo, H.; Alvarez, H.M. The pleiotropic transcriptional regulator NlpR contributes to the modulation of nitrogen metabolism, lipogenesis and triacylglycerol accumulation in oleaginous rhodococci. Mol. Microbiol. 2017, 103, 366–385. [Google Scholar] [CrossRef]
- Hernández, M.A.; Gleixner, G.; Sachse, D.; Alvarez, H.M. Carbon Allocation in Rhodococcus jostii RHA1 in Response to disruption and overexpression of nlpR regulatory Gene, based on 13C-labeling analysis. Front. Microbiol. 2017, 8, 1992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antczak, M.; Płocińska, R.; Płociński, P.; Rumijowska-Galewicz, A.; Żaczek, A.; Strapagiel, D.; Dziadek, J. The NnaR orphan response regulator is essential for the utilization of nitrate and nitrite as sole nitrogen sources in mycobacteria. Sci. Rep. 2018, 8, 17552. [Google Scholar] [CrossRef]
- Kurosawa, K.; Bocazzi, P.; de Almeida, N.M.; Sinskey, A.J. High-cell-density batch fermentation of Rhodococcus opacus PD630 using a high glucose concentration for triacylglycerol production. J. Biotechnol. 2010, 147, 212–218. [Google Scholar] [CrossRef]
- Castro, A.R.; Rocha, I.; Alves, M.M.; Pereira, M.A. Rhodococcus opacus B4: A promising bacterium for production of biofuels and biobased chemicals. AMB Express 2016, 6, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, R.A.; Grossi, V.; Olivera, N.L.; Alvarez, H.M. Characterization of indigenous Rhodococcus sp. 602, a strain able to accumulate triacylglycerides from naphthyl compounds under nitrogen-starved conditions. Res. Microbiol. 2010, 161, 198–207. [Google Scholar] [CrossRef]
- Shields-Menard, S.A.; Amirsadeghi, M.; Sukhbaatar, B.; Revellame, E.; Hernandez, R.; Donaldson, J.R.; French, W.T. Lipid accumulation by Rhodococcus rhodochrous grown on glucose. J. Ind. Microbiol. Biotechnol. 2015, 42, 693–699. [Google Scholar] [CrossRef]
- Bhatia, S.K.; Gurav, R.; Choi, T.R.; Han, Y.H.; Park, Y.L.; Jung, H.R.; Yang, S.Y.; Song, H.S.; Yang, Y.H. A clean and green approach for odd chain fatty acids production in Rhodococcus sp. YHY01 by medium engineering. Bioresour. Technol. 2019, 286, 121383. [Google Scholar] [CrossRef] [PubMed]
- Bellou, S.; Triantaphyllidou, I.E.; Mizerakis, P.; Aggelis, G. High lipid accumulation in Yarrowia lipolytica cultivated under double limitation of nitrogen and magnesium. J. Biotechnol. 2016, 234, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Tchakouteu, S.S.; Kopsahelis, N.; Chatzifragkou, A.; Kalantzi, O.; Stoforos, N.G.; Koutinas, A.A.; Aggelis, G.; Papanikolaou, S. Rhodosporidium toruloides cultivated in NaCl-enriched glucose-based media: Adaptation dynamics and lipid production. Eng. Life Sci. 2016, 17, 237–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juanssilfero, A.B.; Kahar, P.; Amza, R.L.; Miyamoto, N.; Otsuka, H.; Matsumoto, H.; Kihira, C.; Thontowi, A.; Chiaki Ogino, Y.; Prasetya, B.; et al. Effect of inoculum size on single-cell oil production from glucose and xylose using oleaginous yeast Lipomyces starkeyi. J. Biosci. Bioeng. 2018, 6, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Kolouchová, I.; Maťátková, O.; Sigler, K.; Masák, J.; Řezanka, T. Lipid accumulation by oleaginous and non-oleaginous yeaststrains in nitrogen and phosphate limitation. Folia Microbiol. 2016, 61, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Sarkany, N.; Cui, Y. Biomass and lipid productivities of Chlorella vulgaris under autotrophic, heterotrophic and mixotrophic growth conditions. Biotechnol. Lett. 2009, 31, 1043–1049. [Google Scholar] [CrossRef]
- Ward, V.; Rehmann, L. Fast media optimization for mixotrophic cultivation of Chlorella vulgaris. Sci. Rep. 2019, 9, 19262. [Google Scholar] [CrossRef] [Green Version]
- Karpagam, R.; Preeti, R.; Ashokkumar, B.; Varalakshmi, P. Enhancement of lipid production and fatty acid profiling in Chlamydomonas reinhardtii, CC1010 for biodiesel production. Ecotoxicol. Environ. Saf. 2015, 121, 253–257. [Google Scholar] [CrossRef]
- Morales-Sánchez, D.; Tinoco-Valencia, R.; Kyndt, J.; Martinez, A. Heterotrophic growth of Neochloris oleoabundans using glucose as a carbon source. Biotechnol. Biofuels 2013, 6, 100. [Google Scholar] [CrossRef]
- Tang, X.; Chen, H.; Chen, Y.Q.; Chen, W.; Garre, V.; Song, Y.; Ratledge, C. Comparison of biochemical activities between high and low lipid-producing strains of Mucor circinelloides: An explanation for the high oleaginicity of strain WJ11. PLoS ONE 2015, 10, e0128396. [Google Scholar] [CrossRef] [Green Version]
- Bajpai, P.; Bajpai, P.K.; Ward, O.P. Eicosapentaenoic acid (EPA) production by Mortierella alpina ATCC 32222. Appl. Biochem. Biotechnol. 1991, 31, 267–272. [Google Scholar] [CrossRef]
- Chen, H.; Hao, G.; Wang, L.; Wang, H.; Gu, Z.; Liu, L.; Zhang, H.; Chen, W.; Chen, Y.Q. Identification of a critical determinant that enables efficient fatty acid synthesis in oleaginous fungi. Sci. Rep. 2015, 5, 11247. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, S.; Rontou, M.; Belka, A.; Athenaki, M.; Gardeli, C.; Mallouchos, A.; Kalantzi, O.; Koutinas, A.A.; Kookos, I.K.; Zeng, A.P.; et al. Conversion of biodiesel-derived glycerol into biotechnological products of industrial significance by yeast and fungal strains. Eng. Life Sci. 2016, 17, 262–281. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yue, Q.; Zhou, S.; Yan, J.; Zhang, X.; Ma, F. Trehalose Contributes to Gamma-Linolenic Acid Accumulation in Cunninghamella echinulata Based on de Novo Transcriptomic and Lipidomic Analyses. Front. Microbiol. 2018, 9, 1296. [Google Scholar] [CrossRef]
- Chutrakul, C.; Jeennor, S.; Panchanawaporn, S.; Cheawchanlertfa, P.; Suttiwattanakul, S.; Veerana, M.; Laoteng, K. Metabolic engineering of long chain-polyunsaturated fatty acid biosynthetic pathway in oleaginous fungus for dihomo-gamma linolenic acid production. J. Biotechnol. 2016, 218, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Jeennor, S.; Anantayanon, J.; Panchanawaporn, S.; Khoomrung, S.; Chutrakul, C.; Laoteng, K. Reengineering lipid biosynthetic pathways of Aspergillus oryzae for enhanced production of γ-linolenic acid and dihomo-γ-linolenic acid. Gene 2019, 706, 106–114. [Google Scholar] [CrossRef]
- Herrero, O.M.; Villalba, M.S.; Lanfranconi, M.P.; Alvarez, H.M. Rhodococcus bacteria as a promising source of oils from olive mill wastes. World J. Microbiol. Biotechnol. 2018, 34, 114. [Google Scholar] [CrossRef]
- Ochsenreither, K.; Glück, C.; Stressler, T.; Fischer, L.; Syldatk, C. Production Strategies and Applications of Microbial Single Cell Oils. Front. Microbiol. 2016, 7, 1539. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.U.; Park, J.M. Biodiesel production by various oleaginous microorganisms from organic wastes. Bioresour. Technol. 2018, 256, 502–508. [Google Scholar] [CrossRef]
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.H.; Jiang, J.G. Lipid Accumulation Mechanisms in Auto- and Heterotrophic Microalgae. J. Agric. Food Chem. 2017, 65, 8099–8110. [Google Scholar] [CrossRef] [PubMed]
- Chungjatupornchai, W.; Areerat, K.; Fa-Aroonsawat, S. Increased triacylglycerol production in oleaginous microalga Neochloris oleoabundans by overexpression of plastidial lysophosphatidic acid acyltransferase. Microb. Cell Factories 2019, 18, 53. [Google Scholar] [CrossRef] [PubMed]
- Archanaa, S.; Jose, S.; Mukherjee, A.; Suraishkumar, G.K. Sustainable Diesel Feedstock: A Comparison of Oleaginous Bacterial and Microalgal Model Systems. Bioenerg. Res. 2019, 12, 205–216. [Google Scholar] [CrossRef]
- Blakie, B. Production of Omega-3 Fatty Acids in Rhodococcus opacus PD630. Finished Dissertation, Master’s Thesis, University of Otago, Dunedin, New Zealand, 2015. Available online: https://ourarchive.otago.ac.nz/bitstream/handle/10523/5823/BlakieBoromDJ2015MSc.pdf?sequence=1 (accessed on 20 August 2020).
- Chisti, Y. Constraints to commercialization of algal fuels. J. Biotechnol. 2013, 167, 201–214. [Google Scholar] [CrossRef]
- Sitepu, I.R.; Garay, L.A.; Sestric, R.; Levin, D.; Block, D.E.; German, J.B.; Boundy-Mills, K.L. Oleaginous yeasts for biodiesel: Current and future trends in biology and production. Biotechnol. Adv. 2014, 32, 1336–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albers, E.; Johansson, E.; Franzén, C.J.; Larsson, C. Selective suppression of bacterial contaminants by process conditions during lignocellulose based yeast fermentations. Biotechnol. Biofuels 2011, 4, 59. [Google Scholar] [CrossRef] [Green Version]
- Cappelletti, M.; Presentato, A.; Piacenza, E.; Firrincieli, A.; Turner, R.J.; Zannoni, D. Biotechnology of Rhodococcus for the production of valuable compounds. Appl. Microbiol. Biotechnol. 2020, 104, 8567–8594. [Google Scholar] [CrossRef]
- Kalscheuer, R.; Steinbüchel, A. A novel bifunctional wax ester synthase/acyl-CoA:diacylglycerol acyltransferase mediates wax ester and triacylglycerol biosynthesis in Acinetobacter calcoaceticus ADP1. J. Biol. Chem. 2003, 278, 8075–8082. [Google Scholar] [CrossRef] [Green Version]
- Anthony, W.E.; Carr, R.R.; DeLorenzo, D.M.; Campbell, T.P.; Shang, Z.; Foston, M.; Moon, T.S.; Dantas, G. Development of Rhodococcus opacus as a chassis for lignin valorization and bioproduction of high-value compounds. Biotechnol. Biofuels 2019, 12, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, A.; DeLorenzo, D.M.; Carr, R.; Moon, T.S. Bioconversion of renewable feedstocks by Rhodococcus opacus. Curr. Opin. Biotechnol. 2020, 64, 10–16. [Google Scholar] [CrossRef]
- Mahan, K.M.; Le, R.K.; Yuan, J.; Ragauskas, A.J. A review on the bioconversion of lignin to microbial lipid with oleaginous Rhodococcus opacus. J. Biotechnol. Biomater. 2017, 7, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Hollinshead, W.D.; Henson, W.R.; Abernathy, M.; Moon, T.S.; Tang, Y.J. Rapid metabolic analysis of Rhodococcus opacus PD630 via parallel 13C-metabolite fingerprinting. Biotechnol. Bioeng. 2016, 113, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Voss, I.; Steinbüchel, A. High cell density cultivation of Rhodococcus opacus for lipid production at a pilot-plant scale. Appl. Microbiol. Biotechnol. 2001, 55, 547–555. [Google Scholar] [CrossRef]
- Holder, J.W.; Ulrich, J.C.; DeBono, A.C.; Godfrey, P.A.; Desjardins, C.A.; Zucker, J.; Zeng, Q.; Leach, A.L.; Ghiviriga, I.; Dancel, C.; et al. Comparative and functional genomics of Rhodococcus opacus PD630 for biofuels development. PLoS Genet. 2011, 7, e1002219. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, H.M.; Kalscheuer, R.; Steinbüchel, A. Accumulation of storage lipids in species of Rhodococcus and Nocardia and effect of inhibitors and polyethylene glycol. Fett/Lipid 1997, 99, 239–246. [Google Scholar] [CrossRef]
- Zhang, L.S.; Xu, P.; Chu, M.Y.; Zong, M.H.; Yang, J.G.; Lou, W.Y. Using 1-propanol to significantly enhance the production of valuable odd-chain fatty acids by Rhodococcus opacus PD630. World J. Microbiol. Biotechnol. 2019, 35, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Fulco, A.J. Fatty acid metabolism in bacteria. Prog. Lipid Res. 1983, 22, 133–160. [Google Scholar] [CrossRef]
- Park, Y.K.; Dulermo, T.; Ledesma-Amaro, R.; Nicaud, J.M. Optimization of odd chain fatty acid production by Yarrowia lipolytica. Biotechnol. Biofuels 2018, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, B.; West, J.A.; Koulman, A. A Review of Odd-Chain Fatty Acid Metabolism and the Role of Pentadecanoic Acid (C15:0) and Heptadecanoic Acid (C17:0) in Health and Disease. Molecules 2015, 20, 2425–2444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, H.M. Relationship between β-oxidation pathway and the hydrocarbon-degrading profile in actinomycetes bacteria. Int. Biodeterior. Biodegrad. 2003, 52, 35–42. [Google Scholar] [CrossRef]
- Chu, M.Y.; Zhang, L.S.; Lou, W.Y.; Zong, M.H.; Tang, Y.Q.; Yang, J.G. Preparation and characterization of oil rich in odd chain fatty acids from Rhodococcus opacus PD630. J. Am. Oil Chem. Soc. 2019, 97, 25–33. [Google Scholar] [CrossRef]
- Kosa, M.; Ragauskas, A.J. Lipids from heterotrophic microbes: Advances in metabolism research. Trends Biotechnol. 2011, 29, 53–61. [Google Scholar] [CrossRef]
- Herrero, O.M.; Moncalián, G.; Alvarez, H.M. Physiological and genetic differences amongst Rhodococcus species for using glycerol as a source for growth and triacylglycerol production. Microbiology 2016, 162, 384–397. [Google Scholar] [CrossRef]
- Kurosawa, K.; Radek, A.; Plassmeier, J.K.; Sinskey, A.J. Improved glycerol utilization by a triacylglycerol producing Rhodococcus opacus strain for renewable fuels. Biotechnol. Biofuels 2015, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Holert, J.; Brown, K.; Hashimi, A.; Eltis, L.D.; Mohn, W.W. Steryl ester formation and accumulation in steroid-degrading bacteria. Appl. Environ. Microbiol. 2020, 86, e02353-19. [Google Scholar] [CrossRef] [PubMed]
- Ghosal, D.; Ghosh, S.; Dutta, T.K.; Ahn, Y. Current state of knowledge in microbial degradation of polycyclic aromatic hydrocarbons (PAHs): A review. Front. Microbiol. 2016, 7, 1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haritash, A.K. A comprehensive review of metabolic and genomic aspects of PAH-degradation. Arch. Microbiol. 2020, 202, 2033–2058. [Google Scholar] [CrossRef]
- Zampolli, J.; Zeaiter, Z.; Di Canito, A.; Di Gennaro, P. Genome analysis and-omics approaches provide new insights into the biodegradation potential of Rhodococcus. Appl. Microbiol. Biotechnol. 2019, 103, 1069–1080. [Google Scholar] [CrossRef]
- Roell, G.W.; Carr, R.R.; Campbell, T.; Shang, Z.; Henson, W.R.; Czajka, J.J.; Martín, H.G.; Zhang, F.; Fostona, M.; Gautam, D.; et al. A concerted systems biology analysis of phenol metabolism in Rhodococcus opacus PD630. Metab. Eng. 2019, 55, 120–130. [Google Scholar] [CrossRef] [Green Version]
- Henson, W.R.; Hsu, F.F.; Dantas, G.; Moon, T.S.; Foston, M. Lipid metabolism of phenol-tolerant Rhodococcus opacus strains for lignin bioconversion. Biotechnol. Biofuels 2018, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Pond, J.L.; Langworthy, T.A. Effect of growth temperature on the long-chain diols and fatty acids of Thermomicrobium roseum. J. Bacteriol. 1987, 169, 1328–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, H.M.; Luftmann, H.; Silva, R.A.; Cesari, A.C.; Viale, A.; Wältermann, M.; Steinbüchel, A. Identification of phenyldecanoic acid as constituent of triacylglycerols and wax ester produced by Rhodococcus opacus PD630. Microbiology 2002, 148, 1407–1412. [Google Scholar] [CrossRef] [Green Version]
- Goswami, L.; Kumar, R.V.; Manikandan, N.A.; Pakshirajan, K.; Pugazhenthi, G. Simultaneous polycyclic aromatic hydrocarbon degradation and lipid accumulation by Rhodococcus opacus for potential biodiesel production. J. Water Process Eng. 2017, 17, 1–10. [Google Scholar] [CrossRef]
- Herrero, O.M.; Alvarez, H.M. Whey as a renewable source for lipid production by Rhodococcus strains: Physiology and genomics of lactose and galactose utilization. Eur. J. Lipid Sci. Technol. 2016, 118, 262–272. [Google Scholar] [CrossRef]
- Gouda, M.K.; Omar, S.H.; Aouad, L.M. Single cell oil production by Gordonia sp. DG using agro-industrial wastes. World J. Microbiol. Biotechnol. 2008, 24, 1703–1711. [Google Scholar] [CrossRef]
- Saisriyoot, M.; Thanapimmetha, A.; Suwaleerat, T.; Chisti, Y.; Srinophakun, P. Biomass and lipid production by Rhodococcus opacus PD630 in molasses based media with and without osmotic-stress. J. Biotechnol. 2019, 297, 1–8. [Google Scholar] [CrossRef]
- Le, R.K.; Wells, T., Jr.; Das, P.; Meng, X.; Stoklosa, R.J.; Bhalla, A.; Hodge, D.B.; Yuan, J.S.; Ragauskas, A.J. Conversion of corn stover alkaline pre-treatment waste streams into biodiesel via Rhodococci. RSC Adv. 2017, 7, 4108–4115. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Li, X.; Ben, H.; Xue, X.; Yang, B. Lipid Production from Dilute Alkali Corn Stover Lignin by Rhodococcus Strains. ACS Sustain. Chem. Eng. 2017, 5, 2302–2311. [Google Scholar] [CrossRef]
- Paul, T.; Baskaran, D.; Pakshirajan, K.; Pugazhenti, G. Continuous bioreactor with cell recycle using tubular ceramic membrane for simultaneous wastewater treatment and bio-oil production by oleaginous Rhodococcus opacus. Chem. Eng. J. 2019, 367, 76–85. [Google Scholar] [CrossRef]
- Kumar, S.; Gupta, N.; Pakshirajan, K. Simultaneous lipid production and dairy wastewater treatment using Rhodococcus opacus in a batch bioreactor for potential biodiesel application. J. Environ. Chem. 2015, 3, 1630–1636. [Google Scholar] [CrossRef]
- Gupta, N.; Manikandan, N.A.; Pakshirajan, K. Real-time lipid production and dairy wastewater treatment using Rhodococcus opacus in a bioreactor under fed-batch, continuous and continuous cell recycling modes for potential biodiesel application. Biofuels 2017, 9, 239–245. [Google Scholar] [CrossRef]
- Goswami, L.; Namboodiri, M.T.; Kumar, R.V.; Pakshirajan, K.; Pugazhenthi, G. Biodiesel productionPotential oleaginous Rhodococcus opacus grown on biomass gasification wastewater. Renew. Energy 2017, 105, 400–406. [Google Scholar] [CrossRef]
- Wei, Z.; Zeng, G.; Kosa, M.; Huang, D.; Ragauskas, A.J. Pyrolysis Oil-Based Lipid Production as Biodiesel Feedstock by Rhodococcus opacus. Appl. Biochem. Biotechnol. 2015, 175, 1234–1246. [Google Scholar] [CrossRef] [PubMed]
- Dornau, A.; Robson, J.F.; Thomas, G.H.; McQueen Mason, S.J. Robust microorganisms for biofuel and chemical production from municipal solid waste. Microb. Cell Factories 2020, 19, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saisriyoot, M.; Sahaya, T.; Thanapimmetha, A.; Chisti, Y.; Srinophakun, P. Production of potential fuel oils by Rhodococcus opacus grown on petroleum processing wastewaters. J. Renew. Sustain. Energy 2016, 8, 063106. [Google Scholar] [CrossRef]
- McNamara, C.J.; Anastasiou, C.C.; O’ Flaherty, V.; Mitchell, R. Bioremediation of olive mill wastewater. Int. Biodeterior. Biodegrad. 2008, 61, 127–134. [Google Scholar] [CrossRef]
- Barbera, A.C.; Maucieri, C.; Cavallaro, V.; Ioppolo, A.; Spagna, G. Effects of spreading olive mill wastewater on soil properties and crops, a review. Agric. Water Manag. 2013, 119, 43–53. [Google Scholar] [CrossRef]
- Kurosawa, K.; Wewetzer, S.J.; Sinskey, A.J. Triacylglycerol Production from Corn Stover Using a Xylose-Fermenting Rhodococcus opacus Strain for Lignocellulosic Biofuels. J. Microb. Biochem. Technol. 2014, 6, 254–259. [Google Scholar] [CrossRef]
- Patel, A.; Mikes, F.; Matsakas, L. An overview of current pretreatment methods used to improve lipid extraction from oleaginous microorganisms. Molecules 2018, 23, 1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwangbo, M.; Chu, K.H. Recent advances in production and extraction of bacterial lipids for biofuel production. Sci. Total Environ. 2020, 734, 139420. [Google Scholar] [CrossRef] [PubMed]
- Gill, J.J.; Wang, B.; Sestak, E.; Young, R.; Chu, K.H. Characterization of a novel Tectivirus phage toil and its potential as an agent for biolipid extraction. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Liang, M.-H.; Jiang, J.-G. Advancing oleaginous microorganisms to produce lipid via metabolic engineering technology. Prog. Lipid Res. 2013, 52, 395–408. [Google Scholar] [CrossRef]
- Marella, E.R.; Holkenbrink, C.; Siewers, V.; Borodina, I. Engineering microbial fatty acid metabolism for biofuels and biochemicals. Curr. Opin. Biotech 2018, 50, 39–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, J.M.; Alper, H.S. Synthetic biology and molecular genetics in non-conventional yeasts: Current tools and future advances. Fungal. Genet. Biol. 2016, 89, 126–136. [Google Scholar] [CrossRef] [Green Version]
- Patra, P.; Das, M.; Kundu, P.; Ghosh, A. Recent advances in systems and synthetic biology approaches for developing novel cell-factories in non-conventional yeasts. Biotechnol. Adv. 2021, 47, 107695. [Google Scholar] [CrossRef] [PubMed]
- DeLorenzo, D.M.; Rottinghaus, A.G.; Henson, W.R.; Moon, T.S. Molecular Toolkit for Gene Expression Control and Genome Modification in Rhodococcus opacus PD630. ACS Synth. Biol. 2018, 7, 727–738. [Google Scholar] [CrossRef]
- Liang, Y.; Jiao, S.; Wang, M.; Yu, H.; Shen, Z. A CRISPR/Cas9-based genome editing system for Rhodococcus ruber TH. Metab. Eng. 2020, 57, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, A.F.; Alvarez, H.M.; Kalscheuer, R.; Wältermann, M.; Steinbüchel, A. Cloning and characterization of a gene involved in triacylglycerol biosynthesis and identification of additional homologous genes in the oleaginous bacterium Rhodococcus opacus PD630. Microbiology 2008, 154, 2327–2335. [Google Scholar] [CrossRef] [PubMed]
- Hernández, M.A.; Arabolaza, A.; Rodríguez, E.; Gramajo, H.; Alvarez, H.M. The atf2 gene is involved in triacylglycerol biosynthesis and accumulation in the oleaginous Rhodococcusopacus PD630. Appl. Microbiol. Biotechnol. 2013, 97, 2119–2130. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zhao, L.; Zan, X.; Song, Y.; Ratledge, C. Boosting fatty acid synthesis in Rhodococcus opacus PD630 by overexpression of autologous thioesterases. Biotechnol. Lett. 2016, 38, 999–1008. [Google Scholar] [CrossRef]
- MacEachran, D.P.; Prophete, M.E.; Sinskey, A.J. The Rhodococcus opacus PD630 heparin-binding hemagglutinin homolog TadA mediates lipid body formation. Appl. Environ. Microbiol. 2010, 76, 7217–7225. [Google Scholar] [CrossRef] [Green Version]
- Xie, S.; Sun, S.; Lin, F.; Li, M.; Pu, Y.; Cheng, Y.; Xu, B.; Liu, Z.; da Costa Sousa, L.; Dale, B.E.; et al. Mechanism-Guided Design of Highly Efficient Protein Secretion and Lipid Conversion for Biomanufacturing and Biorefining. Adv. Sci. 2019, 6, 1801980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.M.; Chae, T.U.; Choi, S.Y.; Kim, W.J.; Lee, S.Y. Engineering of an oleaginous bacterium for the production of fatty acids and fuels. Nat. Chem. Biol. 2019, 15, 721–729. [Google Scholar] [CrossRef]
- Hernández, M.A.; Comba, S.; Arabolaza, A.; Gramajo, H.; Alvarez, H.M. Overexpression of a phosphatidic acid phosphatase type 2 leads to an increase in triacylglycerol production in oleaginous Rhodococcus strains. Appl. Microbiol. Biotechnol. 2015, 99, 2191–2207. [Google Scholar] [CrossRef]
- Villalba, M.S.; Alvarez, H.M. Identification of a novel ATP-binding cassette transporter involved in long-chain fatty acid import and its role in triacylglycerol accumulation in Rhodococcus jostii RHA1. Microbiology 2014, 160, 1523–1532. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Yang, L.; Zhang, S.; Wang, Y.; Du, Y.; Pu, J.; Peng, G.; Chen, Y.; Zhang, H.; Yu, J.; et al. Identification of the major functional proteins of prokaryotic lipid droplets. J. Lipid Res. 2012, 53, 399–411. [Google Scholar] [CrossRef] [Green Version]
- Carman, G.M.; Han, G.-S. Roles of phosphatidate phosphatase enzymes in lipid metabolism. Trends Biochem. Sci. 2006, 31, 694–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Yang, Z.; Huang, X.; Peng, J.; Fei, X.; Gu, S.; Xie, Y.; Ji, C.; Mao, Y. Cloning; expression and characterization of a thermostable PAP2L2; a new member of the type-2 phosphatidic acid phosphatase family from Geobacillus toebii T-85. Biosci. Biotechnol. Biochem. 2008, 72, 3134–3141. [Google Scholar] [CrossRef] [PubMed]
- Comba, S.; Menendez-Bravo, S.; Arabolaza, A.; Gramajo, H. Identification and physiological characterization of phosphatidic acid phosphatase enzymes involved in triacylglycerol biosynthesis in Streptomyces coelicolor. Microb. Cell Factories 2013, 12, 9. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Ding, Y.; Yang, L.; Yu, J.; Liu, G.; Wang, X.; Zhang, S.; Yu, D.; Song, L.; Zhang, H.; et al. Integrated omics study delineates the dynamics of lipid droplets in Rhodococcus opacus PD630. Nucleic Acids Res. 2013, 42, 1052–1064. [Google Scholar] [CrossRef] [PubMed]
- Ishige, T.; Tani, A.; Sakai, Y.; Kato, N. Wax ester production by bacteria. Curr. Opin. Microbiol. 2003, 6, 244–250. [Google Scholar] [CrossRef]
- Rontani, J.-F. Production of wax esters by bacteria. In Handbook of Hydrocarbon and Lipid Microbiology; Timmis, K.N., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 459–470. [Google Scholar] [CrossRef]
- Wältermann, M.; Steinbüchel, A. Wax Ester and Triacylglycerol Inclusions. In Inclusions in Prokaryotes; Microbiology Monographs; Shively, J.M., Ed.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 1, pp. 138–160. [Google Scholar] [CrossRef]
- Willis, R.M.; Wahlen, B.D.; Seefeldt, L.C.; Barney, B.M. Characterization of a fatty acyl-CoA reductase from Marinobacter aquaeolei VT8: A bacterial enzyme catalyzing the reduction of fatty acyl-CoA to fatty alcohol. Biochemistry 2011, 50, 10550–10558. [Google Scholar] [CrossRef] [PubMed]
- Silva, R.A.; Grossi, V.; Alvarez, H.M. Biodegradation of phytane (2,6,10,14-tetramethylhexadecane) and accumulation of related isoprenoid wax esters by Mycobacterium ratisbonense strain SD4 under nitrogen-starved conditions. FEMS Microbiol. Lett. 2007, 272, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Barney, B.M.; Wahlen, B.D.; Garner, E.; Wei, J.; Seefeldt, L.C. Differences in substrate specificities of five bacterial wax ester synthases. Appl. Environ. Microbiol. 2012, 78, 5734–5745. [Google Scholar] [CrossRef] [Green Version]
- Round, J.; Roccor, R.; Li, S.-N.; Eltis, L.D. A Fatty Acyl Coenzyme A Reductase Promotes Wax Ester Accumulation in Rhodococcus jostii RHA1. Appl. Environ. Microbiol. 2017, 83, e00902-17. [Google Scholar] [CrossRef] [Green Version]
- Round, J.W.; Roccor, R.; Eltis, L.D. A biocatalyst for sustainable wax ester production: Re-wiring lipid accumulation in Rhodococcus to yield high-value oleochemicals. Green Chem. 2019, 21, 6468–6482. [Google Scholar] [CrossRef]
- Lanfranconi, M.P.; Alvarez, H.M. Rewiring neutral lipids production for the de novo synthesis of wax esters in Rhodococcus opacus PD630. J. Biotechnol. 2017, 260, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Hofvander, P.; Doan, T.T.; Hamberg, M. A prokaryotic acyl-CoA reductase performing reduction of fatty acyl-CoA to fatty alcohol. FEBS Lett. 2011, 585, 3538–3543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
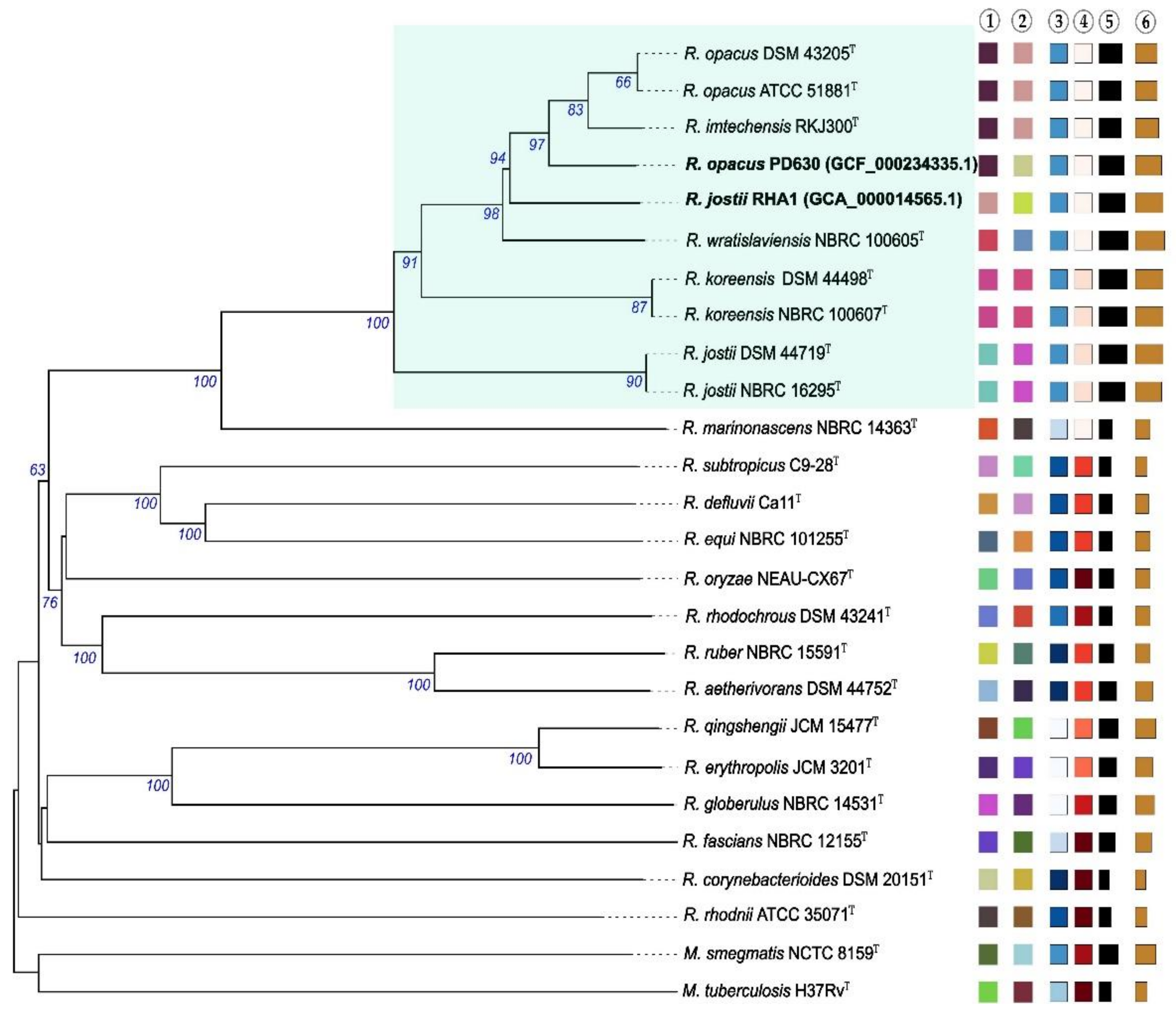
 , Max. 70.7%
, Max. 70.7%  ); ④ δ values (Min. 0.1
); ④ δ values (Min. 0.1  , Max. 0.3
, Max. 0.3  ); ⑤ overall genome sequence length (Min. 3.9 Mb
); ⑤ overall genome sequence length (Min. 3.9 Mb  , Max. 10.4
, Max. 10.4  ); and ⑥ number of proteins (Min. 3611
); and ⑥ number of proteins (Min. 3611  , Max. 9472
, Max. 9472  ).
, Max. 70.7% ); ④ δ values (Min. 0.1 , Max. 0.3 ); ⑤ overall genome sequence length (Min. 3.9 Mb , Max. 10.4 ); and ⑥ number of proteins (Min. 3611 , Max. 9472 ).
).
, Max. 70.7% ); ④ δ values (Min. 0.1 , Max. 0.3 ); ⑤ overall genome sequence length (Min. 3.9 Mb , Max. 10.4 ); and ⑥ number of proteins (Min. 3611 , Max. 9472 ).



{kind=link}
{kind=link}
{kind=link}
| SCM Type | Microorganisms | a Cultivation Mode | Cellular Biomass | b Total Fatty Acid Content (% CDW) and Cell Culture Time | c Main Fatty Acid Composition | References |
|---|---|---|---|---|---|---|
| Actinobacteria (oleaginous rhodococci) | R. jostii RHA1 | Shake flask | - | 50–60%, 2 days | SFAs (C16:0, C17:0); MUFAs (C16:1, C17:1, C18:1) | [9] |
| R. opacus PD630 | -Shake flask -Batch bioreactor -Shake flask | - 77.6 g/L 3.9 g/L | 50–76%, 2 days 38%, 4 days 28.7%, 1 day | SFAs (C16:0, C17:0); MUFAs (C17:1, C18:1) | [8,23,24] | |
| R. jostii 602 | Shake flask | - | 60–70%, 2 days | SFAs (C16:0, C17:0, C18:0); MUFAs (C18:1) | [25] | |
| R. opacus B4 | Shake flask | 2.5 g/L | 23%, 1 day | SFAs (C16:0, C17:0); MUFAs (C17:1, C18:1) | [24] | |
| R. rhodochrous | Batch bioreactor | 7 g/L | 43%, 7 days | SFAs (C16:0); MUFAs (C18:1) | [26] | |
| Rhodococcus sp. YHY01(marine strain) | Shake Flask | 1.3 g/L | 56%, 3 days | SFAs (C14:0, C16:0, C18:0); MUFAs (C18:1) | [27] | |
| Yeast | Yarrowia lipolytica ACA-DC50109 | Continuous bioreactor | 12.2 g/L | 47.5%, 9 days | SFAs (C16:0); MUFAs (C16:1,C18:1, C18:2) | [28] |
| Rhodosporidium toruloides Y4 | Fed-Batch bioreactor | 37.2 g/L | 64.5%, 6 days | SFAs (C16:0); MUFAs (C18:1, C18:2) | [29] | |
| Lipomyces starkeyiNBRC10381 | Shake flask | 21.6 g/L | 71.5%, 6 days | SFAs (C16:0); MUFAs (C18:1) | [30] | |
| Rhodotorula glutinis | Shake flask | 6.8 g/L | 43.0%, 4 days hs | SFAs (C16:0); MUFAs (C18:1) | [31] | |
| Microalgae (heterotrophic or mixotrophic conditions) | Chlorella vulgaris UTEX 259; UTEX 2714 | Shake flask Shake flask | 1.7g/L 6.1 g/L | 21%, 6 days 37.6%, 6 days | SFAs (C16:0); PUFAs (C18:2, GLA) | [32,33] |
| ChlamydomonareinhardtiiCC1010 | Shake flask | 0.37 g/L | 47% | SFAs (C16:0); MUFAs (C18:1) | [34] | |
| Neochloris oleoabundans UTEX 1185 | Batch bioreactor | 9.2 g/L | 52%, 9 days | SFAs (C16:0, C18:0); MUFAs (C18:1, C18:2) | [35] | |
| Fungi | Mucor circinelloidesWJ11 | Batch bioreactor | 14 g/L | 36%,3.5 days | SFAs (C16:0); MUFAs (C18:1); PUFAs (C18:2, GLA) | [36] |
| Mortierella alpina ATCC 32222 | -Shake flask -Batch bioreactor | 18 g/L 5 g/L | 18%, 6 days 20%, 3 days | SFAs (C16:0, C18:0); MUFAs (C18:1); PUFAs (ARA) | [37,38] | |
| Mortierella isabellina ATHUM 2935 | Shake flask | 10.2 g/L | 83%, 4 days | SFAs (C16:0); MUFAs (C18:1); PUFAs (C18:2) | [39] | |
| Cunningha mellaechinulata FR3 | Shake flask | 8.6 g/L | 34%, 8 days | SFAs (C16:0); MUFAs (C18:1); PUFAs (C18:2, GLA) | [40] | |
| Aspergillus oryzae BCC14614 | Shake flask | 16 g/L | 12–20%, 3–5 days | SFAs (C16:0; C18:0); MUFAs (C18:1); PUFAs (C18:2) | [41,42] |
| Strains | Wastes | Lipid Content (% w/w of CDW) | Biomass | References |
|---|---|---|---|---|
| R. opacus PD630 | Whey permeate | 45.1 | 6.1 a | [81] |
| R. opacus MR22 | Whey permeate | 46.1 | 6.3 a | [81] |
| R. opacus PD630 | Sugar cane molasses | 96 | NR | [82] |
| R. opacus PD630 | Orange waste | 83 | NR | [82] |
| R. opacus PD630 | Sweet whey | 82 | NR | [82] |
| R. opacus PD630 | Beet molasses and sucrose (Fed-batch biorreactor) | 52 | 37.4 a | [60] |
| R. opacus PD630 | Beet molasses and sucrose (stirred tank reactor) | 38.4 | 18.4 a | [60] |
| R. opacus PD630 | Sugar cane molasses | 29.8 | 11.9 a | [83] |
| R. opacus PD630 | Corn stover waste streams | 42.1 | 2.13 a | [84] |
| R. opacuS DSM 1069 | Corn stover waste streams | 12.6 | 2.02 a | [84] |
| R. jostii DSM 44719 | Corn stover waste streams | 23.3 | 1.1 a | [84] |
| Co- fermentation R. opacus PD630 and R. jostii RHA1VanA- | Alkali corn stover lignin | 29 | 0.35 b | [85] |
| R. opacus PD630 | Raw refinery wastewater | 86 | 3.6 a | [86] |
| R. opacus DSM 43205 | Raw dairy wastewater (Batch flask) | 33.3 | 4.5 b | [87] |
| R. opacus DSM 43205 | Raw dairy wastewater (Batch bioreactor) | 52 | 2.7 b | [87] |
| R. opacus PD630 | Dairy wastewater | 79 | 4 a | [88] |
| R. opacus DSM 43205 | Biomass gasification wastewater | 62.8 | 0.7 b | [89] |
| R. opacus PD630 | Light oil from pyrolysis biomass | 25.8 | 0.82 a | [90] |
| R. opacus DSM 1069 | Light oil from pyrolysis biomass | 22 | 0.90 a | [90] |
| R. opacus PD630 | Glycerol | 38.4 | 3.8 a | [70] |
| R. opacus MR22 | Glycerol | 35.5 | 3.3 a | [70] |
| R. jostii RHA1 | Glycerol | 30.5 | 2.5 a | [70] |
| R. erythropolis DSM 43060 | Glycerol | 38.3 | 4.1 a | [70] |
| R. fascians F7 | Glycerol | 44.6 | 4.3 a | [70] |
| R. opacus PD630-pTip-QC2/glpK1D1F7 | Glycerol | 41 | 4.4 a | [70] |
| R. opacus MITXM-173 | Glycerol | 40.4 | 5.69 a | [71] |
| R. opacus PD630 | Olive oil mill | 68.2 | 2.2 a | [43] |
| R. opacus MR22 | Olive oil mill | 82.2 | 2.2 a | [43] |
| R. jostii RHA1 | Olive oil mill | 67.3 | 2.6 a | [43] |
| R. wratislaviensis V | Olive oil mill | 88.4 | 2.6 a | [43] |
| R. opacus MITXM-61 | Organic fraction municipal solid waste fiber hydrolysate | 48.9 | 32.7 a | [91] |
| R. opacus PD630 | Petroleum wastewater supplemented with molasses (Batch bioreactor) | 52.5 | 5.91 a | [92] |
| R. opacus PD630 | Petroleum wastewater supplemented with molasses (Fed-batch bioreactor) | 54.4 | 7.24 a | [92] |
| Rhodococcus Strain | Genetic Strategy | Goal of the Study | References |
|---|---|---|---|
| R. opacus PD630 | Disruption and overexpression of atf1, atf2 genes | Gene role evaluation in TAG biosynthesis. TAG accumulation improving | [105,106] |
| R. opacus PD630 | Disruption and overexpression of nlpR gene | Gene role evaluation in TAG biosynthesis. TAG accumulation improving, under nitrogen rich-conditions | [20] |
| R. opacus PD630 | Thioesterasesoverexpression | Fatty acid (FA) biosynthesis improving | [107] |
| R. opacus PD630 | tadA disruption and overexpression | Gene role evaluation in LDs ontogeny. Effect on TAG accumulation | [108] |
| R. opacus PD630 | tadD disruption and overexpression | Gene role evaluation in TAG biosynthesis. TAG accumulation improving from NADPH supply | [13] |
| R. opacus PD630 | Disruption and overexpression of fasI gene | Gene role evaluation in TAG biosynthesis. TAG accumulation improving, under nitrogen rich-conditions | [109] |
| R. opacus PD630 | Acyl-coenzyme A (CoA) synthetases deletion and overexpression of three lipases. | Free fatty acids (FFAs) production | [110] |
| R. opacus PD630 | Acyl-CoA dehydrogenases deletion, overexpression of lipases, foldase, acyl-CoA synthetase and heterologous expression of an aldehyde/alcohol dehydrogenase and wax ester synthase | Fatty acid ethyl esters (FAEEs) production | [110] |
| R. opacus PD630 | Acyl-CoA dehydrogenases and alkane-1 monooxygenase deletion, overexpression of lipases, foldase, acyl-CoA synthetase and heterologous expression of acyl-CoA reductase, acyl-ACP reductase, and aldehyde deformylating oxygenase | Long-chain hydrocarbons (LCHCs) production | [110] |
| R. jostii RHA1 | Disruption and overexpression of atf8 gene | Gene role evaluation in TAG biosynthesis | [16] |
| R. jostii RHA1 | Disruption and overexpression of nlpR gene | Gene role evaluation in TAG biosynthesis. TAG accumulation improving, under nitrogen rich-conditions | [20] |
| R. jostii RHA1 | Deletion and overexpression of pap2 gene | Gene role evaluation in TAG biosynthesis. TAG accumulation improving from DAG supply | [111] |
| R. jostii RHA1 | Overexpression of ME-NADP+ gene (malic enzyme) | Gene role evaluation in TAG biosynthesis. TAG accumulation improving from NADPH supply | [14] |
| R. jostii RHA1 | Overexpression of ltp1 gene | Gene role evaluation in TAG biosynthesis. TAG and cellular biomass improving from fatty acids | [112] |
| R. jostii RHA1 | mlds deletion | Gene role evaluation in LDs ontogeny. Effect on TAG accumulation | [113] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvarez, H.M.; Hernández, M.A.; Lanfranconi, M.P.; Silva, R.A.; Villalba, M.S. Rhodococcus as Biofactories for Microbial Oil Production. Molecules 2021, 26, 4871. https://doi.org/10.3390/molecules26164871
Alvarez HM, Hernández MA, Lanfranconi MP, Silva RA, Villalba MS. Rhodococcus as Biofactories for Microbial Oil Production. Molecules. 2021; 26(16):4871. https://doi.org/10.3390/molecules26164871
Chicago/Turabian StyleAlvarez, Héctor M., Martín A. Hernández, Mariana P. Lanfranconi, Roxana A. Silva, and María S. Villalba. 2021. "Rhodococcus as Biofactories for Microbial Oil Production" Molecules 26, no. 16: 4871. https://doi.org/10.3390/molecules26164871
APA StyleAlvarez, H. M., Hernández, M. A., Lanfranconi, M. P., Silva, R. A., & Villalba, M. S. (2021). Rhodococcus as Biofactories for Microbial Oil Production. Molecules, 26(16), 4871. https://doi.org/10.3390/molecules26164871