
Overview of the Antioxidant and Anti-Inflammatory Activities of Selected Plant Compounds and Their Metal Ions Complexes


 ,
,
Abstract
:1. Introduction
2. Mechanism of Free Radicals
2.1. Exogenous ROS Production
2.2. Endogenous ROS/RNS Production
2.3. Biological Outcomes of Oxidation by ROS
2.3.1. ROS and Lipids
2.3.2. ROS and Proteins
2.3.3. ROS and DNA
3. Antioxidant Defense System
4. Biological Activities of Plant Compounds
4.1. Anticancer Activity
4.2. Anti-Inflammatory Activity
4.3. Antidiabetic Activity
4.4. Antibacterial Activity
5. Flavonoids, Coumarins and Chromones as Anti-Oxidative and Anti-Inflammatory Agents
5.1. Flavonoids as Antioxidants
5.2. Coumarins as Antioxidants
5.3. Chromones as Antioxidants
6. Antioxidant Complexes of Flavonoids, Coumarins, Chromones with Metal Ions
6.1. Complexes of Flavonoids with Metal Ions
6.2. Complexes of Coumarins with Metal Ions
6.3. Complexes of Chromones with Metal Ions
7. Binding Motifs and Supramolecular Architecture for Crystal Structure of Plant Compounds and Their Metal Complexes
7.1. Flavonoids (Flav)–Metal Complexes
- O-donors such as H2O, ClO4, NO3, dimethylsulfoxide (DMSO), 2-(2-hydroxyphenyl)-2-oxoacetic acid;
- P-donors such as PPh3;
- F-donors such as BF4;
- N-donors including pyridine(py);
- N,N-donors ((2,2′bipyridine (bipy), N,N,N′,N′-tetramethylenediamine (temed));
- N,N,N-donor (bis(pyridin-2-ylmethyl)-l2-azane (aza), 1,4,7-tribenzyl-1,4,7-triazonane (azon), N1-(3-(dimethylamino)propyl)-N3,N3-dimethylpropane-1,3-diamine (dpp), tri(1H-pyrazol-1-yl)borane (pyb), tris(1-ethyl-4-methyl-1H-imidazol-2-yl)phosphane (trim), tris(pyridin-2-ylmethyl)amine (tripy), N,N,N (1Z,3Z)-N,N′′′-di(pyridin-2-yl)-2l2-isoindoline-1,3-diimine (dipyim));
- N,N,N,N,-donor tris(pyridin-2-ylmethyl)amine (tripy);
- N,O-donors:(E)-1-(2-(l1-oxidaneyl)phenyl)-N-phenylmethanimine (oxphen);
- N,N,N,O-donor (2-((bis(pyridin-2-ylmethyl)amino)methyl)phenyl)(l1-oxidaneyl)methanone (azam);
- N,N,O,O,-donor (1E,1′E)-N,N′-(ethane-1,2-diyl)bis(1-(2-(l1-oxidaneyl)phenyl)methanimine) (etoxian) (Figure 12).
7.1.1. Cu(II)-Flav Complexes
7.1.2. Zn(II)-Flav Complexes
7.1.3. Co(II)-Flav Complexes
7.1.4. Fe(II/III)-Flav Complexes
7.1.5. Ni(II)-Flav Complexes
7.1.6. Al(III)-Flav Complexes
7.2. Chromones (Chrom)–Metal Ion Complexes
7.2.1. Cu(II)-Chromone Complexes
7.2.2. Zn(II)-Chromone Complexes
7.2.3. Ni(II)-Chromone Complexes
7.3. Coumarins (Coum)–Metal Complexes
7.3.1. Cu(II)-Coumarin Complexes
7.3.2. Zn(II)-Coumarin Complexes
7.3.3. Co(II)-Coumarin Complexes
7.3.4. Ni(II)-Coumarin Complexes
8. Application of Plant Compounds and Their Complexes in Medicinal Chemistry and Pharmacy
9. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Neha, K.; Haider, M.R.; Pathak, A.; Yar, M.S. Medicinal prospects of antioxidants: A review. Eur. J. Med. Chem. 2019, 178, 687–704. [Google Scholar] [CrossRef] [PubMed]
- Snezhkina, A.V.; Kudryavtseva, A.V.; Kardymon, O.L.; Savvateeva, M.V.; Melnikova, N.V.; Krasnov, G.S.; Dmitriev, A.A. ROS Generation and Antioxidant Defense Systems in Normal and Malignant Cells. Oxid. Med. Cell. Longev. 2019, 2019, 6175804. [Google Scholar] [CrossRef]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef]
- Moldogazieva, N.T.; Mokhosoev, I.M.; Feldman, N.B.; Lutsenko, S.V. ROS and RNS signalling: Adaptive redox switches through oxidative/nitrosative protein modifications. Free Radic. Res. 2018, 52, 507–543. [Google Scholar] [CrossRef]
- Ferreira, C.A.; Ni, D.; Rosenkrans, Z.T.; Cai, W. Scavenging of reactive oxygen and nitrogen species with nanomaterials. Nano Res. 2018, 11, 4955–4984. [Google Scholar] [CrossRef]
- Spitz, D.R.; Hauer-Jensen, M. Ionizing Radiation-Induced Responses: Where Free Radical Chemistry Meets Redox Biology and Medicine. Antioxid. Redox Signal. 2014, 20, 1407–1409. [Google Scholar] [CrossRef]
- Ramsay, R.R. Electron carriers and energy conservation in mitochondrial respiration. ChemTexts 2019, 5, 9. [Google Scholar] [CrossRef] [Green Version]
- Perillo, B.; Di Donato, M.; Pezone, A.; Di Zazzo, E.; Giovannelli, P.; Galasso, G.; Castoria, G.; Migliaccio, A. ROS in cancer therapy: The bright side of the moon. Exp. Mol. Med. 2020, 52, 192–203. [Google Scholar] [CrossRef]
- Sproll, C.; Ruge, W.; Andlauer, C.; Godelmann, R.; Lachenmeier, D.W. HPLC analysis and safety assessment of coumarin in foods. Food Chem. 2008, 109, 462–469. [Google Scholar] [CrossRef]
- Shen, Y.X.; Teng, H.L.; Yang, G.Z.; Mei, Z.N.; Chen, X.L. A new chromone derivative from Berchemia lineata. Yao Xue Xue Bao 2010, 45, 1139–1143. [Google Scholar]
- Tanaka, N.; Kashiwada, Y.; Nakano, T.; Shibata, H.; Higuchi, T.; Sekiya, M.; Ikeshiro, Y.; Takaishi, Y. Chromone and chromanone glucosides from Hypericumsikokumontanum and their anti-Helicobacter pylori activities. Phytochemistry 2009, 70, 141–146. [Google Scholar] [CrossRef]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric oxide AND Peroxynitrite in health and disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [Green Version]
- Oke, G.O.; Abiodun, A.A.; Imafidon, C.E.; Monsi, B.F. Zingiber officinale (Roscoe) mitigates CCl4-induced liver histopathology and biochemical derangements through antioxidant, membrane-stabilizing and tissue-regenerating potentials. Toxicol. Rep. 2019, 6, 416–425. [Google Scholar] [CrossRef]
- Antunes dos Santos, A.; Ferrer, B.; Marques Gonçalves, F.; Tsatsakis, A.; Renieri, E.; Skalny, A.; Farina, M.; Rocha, J.; Aschner, M. Oxidative Stress in Methylmercury-Induced Cell Toxicity. Toxics 2018, 6, 47. [Google Scholar] [CrossRef] [Green Version]
- Hamanaka, R.B.; Glasauer, A.; Hoover, P.; Yang, S.; Blatt, H.; Mullen, A.R.; Getsios, S.; Gottardi, C.J.; DeBerardinis, R.J.; Lavker, R.M.; et al. Mitochondrial reactive oxygen species promote epidermal differentiation and hair follicle development. Sci. Signal. 2013, 6, ra8. [Google Scholar] [CrossRef] [Green Version]
- Cheng, D.-B.; Zhang, X.-H.; Gao, Y.-J.; Ji, L.; Hou, D.; Wang, Z.; Xu, W.; Qiao, Z.-Y.; Wang, H. Endogenous Reactive Oxygen Species-Triggered Morphology Transformation for Enhanced Cooperative Interaction with Mitochondria. J. Am. Chem. Soc. 2019, 141, 7235–7239. [Google Scholar] [CrossRef]
- Papa, S.; Martino, P.L.; Capitanio, G.; Gaballo, A.; de Rasmo, D.; Signorile, A.; Petruzzella, V. The Oxidative Phosphorylation System in Mammalian Mitochondria. Adv. Exp. Med. Biol. 2012, 942, 3–37. [Google Scholar]
- Hu, N.; Ren, J. Reactive Oxygen Species Regulate Myocardial Mitochondria through Post-Translational Modification. React. Oxyg. Species 2016, 2, 264–271. [Google Scholar] [CrossRef]
- Peoples, J.N.; Saraf, A.; Ghazal, N.; Pham, T.T.; Kwong, J.Q. Mitochondrial dysfunction and oxidative stress in heart disease. Exp. Mol. Med. 2019, 51, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kaarniranta, K.; Pawlowska, E.; Szczepanska, J.; Jablkowska, A.; Blasiak, J. Role of Mitochondrial DNA Damage in ROS-Mediated Pathogenesis of Age-Related Macular Degeneration (AMD). Int. J. Mol. Sci. 2019, 20, 2374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rimessi, A.; Previati, M.; Nigro, F.; Wieckowski, M.R.; Pinton, P. Mitochondrial reactive oxygen species and inflammation: Molecular mechanisms, diseases and promising therapies. Int. J. Biochem. Cell Biol. 2016, 81, 281–293. [Google Scholar] [CrossRef]
- Van der Vliet, A.; Janssen-Heininger, Y.M.W.; Anathy, V. Oxidative stress in chronic lung disease: From mitochondrial dysfunction to dysregulated redox signaling. Mol. Asp. Med. 2018, 63, 59–69. [Google Scholar] [CrossRef]
- Dhawan, V. Reactive Oxygen and Nitrogen Species: General Considerations. In Studies on Respiratory Disorders. Oxidative Stress in Applied Basic Research and Clinical Practice; Ganguly, N., Jindal, S., Biswal, S., Barnes, P., Pawankar, R., Eds.; Humana Press: New York, NY, USA, 2014; pp. 27–47. [Google Scholar] [CrossRef]
- Martínez, M.C.; Andriantsitohaina, R. Reactive nitrogen species: Molecular mechanisms and potential significance in health and disease. Antioxid. Redox Signal. 2009, 11, 669–702. [Google Scholar] [CrossRef]
- Thomas, D.C. The phagocyte respiratory burst: Historical perspectives and recent advances. Immunol. Lett. 2017, 192, 88–96. [Google Scholar] [CrossRef]
- del Río, L.A.; López-Huertas, E. ROS Generation in Peroxisomes and its Role in Cell Signaling. Plant Cell Physiol. 2016, 57, 1364–1376. [Google Scholar] [CrossRef]
- Fransen, M.; Nordgren, M.; Wang, B.; Apanasets, O. Role of peroxisomes in ROS/RNS-metabolism: Implications for human disease. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2012, 1822, 1363–1373. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Lu, W.; Shi, B.; Klein, S.; Su, X. Peroxisomal regulation of redox homeostasis and adipocyte metabolism. Redox Biol. 2019, 24, 101167. [Google Scholar] [CrossRef]
- Schwarz, D.S.; Blower, M.D. The endoplasmic reticulum: Structure, function and response to cellular signaling. Cell. Mol. Life Sci. 2015, 73, 79–94. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; An, A.; Park, H.; Jang, K.; Moon, W.; Kang, M.; Lee, Y.; Ku, J.; Chung, M. Combined expression of protein disulfide isomerase and endoplasmic reticulum oxidoreductin 1-α is a poor prognostic marker for non-small cell lung cancer. Oncol. Lett. 2018, 16, 5753–5760. [Google Scholar] [CrossRef] [Green Version]
- Zito, E. ERO1: A protein disulfide oxidase and H2O2 producer. Free Radic. Biol. Med. 2015, 83, 299–304. [Google Scholar] [CrossRef]
- Zeeshan, H.; Lee, G.; Kim, H.-R.; Chae, H.-J. Endoplasmic Reticulum Stress and Associated ROS. Int. J. Mol. Sci. 2016, 17, 327. [Google Scholar] [CrossRef] [Green Version]
- Tabata, S.; Yamamoto, M.; Goto, H.; Hirayama, A.; Ohishi, M.; Kuramoto, T.; Mitsuhashi, A.; Ikeda, R.; Haraguchi, M.; Kawahara, K.; et al. Thymidine catabolism promotes NADPH oxidase-derived reactive oxygen species (ROS) signalling in KB and yumoto cells. Sci. Rep. 2018, 8, 6760. [Google Scholar] [CrossRef] [Green Version]
- Pegg, A.E. Functions of Polyamines in Mammals. J. Biol. Chem. 2016, 291, 14904–14912. [Google Scholar] [CrossRef] [Green Version]
- Gwozdzinski, K.; Pieniazek, A.; Gwozdzinski, L. Reactive Oxygen Species and Their Involvement in Red Blood Cell Damage in Chronic Kidney Disease. Oxid. Med. Cell. Longev. 2021, 2021, 6639199. [Google Scholar] [CrossRef]
- Burtenshaw, D.; Hakimjavadi, R.; Redmond, E.; Cahill, P. Nox, Reactive Oxygen Species and Regulation of Vascular Cell Fate. Antioxidants 2017, 6, 90. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Li, F.; Duan, Y.; Wen, C.; Wang, W.; Zhang, L.; Huang, R.; Yin, Y. Oxidative stress, nutritional antioxidants and beyond. Sci. China Life Sci. 2019, 63, 866–874. [Google Scholar] [CrossRef]
- Niki, E.; Yoshida, Y.; Saito, Y.; Noguchi, N. Lipid peroxidation: Mechanisms, inhibition, and biological effects. Biochem. Biophys. Res. Commun. 2005, 338, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Su, L.-J.; Zhang, J.-H.; Gomez, H.; Murugan, R.; Hong, X.; Xu, D.; Jiang, F.; Peng, Z.-Y. Reactive Oxygen Species-Induced Lipid Peroxidation in Apoptosis, Autophagy, and Ferroptosis. Oxid. Med. Cell. Longev. 2019, 2019, 5080843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willems, P.H.G.M.; Rossignol, R.; Dieteren, C.E.J.; Murphy, M.P.; Koopman, W.J.H. Redox Homeostasis and Mitochondrial Dynamics. Cell Metab. 2015, 22, 207–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, D.; Xiong, X.; Yang, H.; Wang, M.; He, Y.; Liu, Y.; Yin, Y. Effect of dietary soy oil, glucose, and glutamine on growth performance, amino acid profile, blood profile, immunity, and antioxidant capacity in weaned piglets. Sci. China Life Sci. 2018, 61, 1233–1242. [Google Scholar] [CrossRef] [PubMed]
- Maulucci, G.; Daniel, B.; Cohen, O.; Avrahami, Y.; Sasson, S. Hormetic and regulatory effects of lipid peroxidation mediators in pancreatic beta cells. Mol. Asp. Med. 2016, 49, 49–77. [Google Scholar] [CrossRef] [PubMed]
- Crescenzo, R.; Bianco, F.; Mazzoli, A.; Giacco, A.; Liverini, G.; Iossa, S. A possible link between hepatic mitochondrial dysfunction and diet-induced insulin resistance. Eur. J. Nutr. 2015, 55, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Hauck, A.K.; Bernlohr, D.A. Oxidative stress and lipotoxicity. J. Lipid Res. 2016, 57, 1976–1986. [Google Scholar] [CrossRef] [Green Version]
- Daiber, A.; Daub, S.; Bachschmid, M.; Schildknecht, S.; Oelze, M.; Steven, S.; Schmidt, P.; Megner, A.; Wada, M.; Tanabe, T.; et al. Protein Tyrosine Nitration and Thiol Oxidation by Peroxynitrite—Strategies to Prevent These Oxidative Modifications. Int. J. Mol. Sci. 2013, 14, 7542–7570. [Google Scholar] [CrossRef] [Green Version]
- Torres-Cuevas, I.; Kuligowski, J.; Cárcel, M.; Cháfer-Pericás, C.; Asensi, M.; Solberg, R.; Cubells, E.; Nuñez, A.; Saugstad, O.D.; Vento, M.; et al. Protein-bound tyrosine oxidation, nitration and chlorination by-products assessed by ultraperformance liquid chromatography coupled to tandem mass spectrometry. Anal. Chim. Acta 2016, 913, 104–110. [Google Scholar] [CrossRef]
- Saladino, J.; Liu, M.; Live, D.; Sharp, J.S. Aliphatic peptidyl hydroperoxides as a source of secondary oxidation in hydroxyl radical protein footprinting. J. Am. Soc. Mass Spectrom. 2009, 20, 1123–1126. [Google Scholar] [CrossRef] [Green Version]
- Davies, M.J. Protein and Peptide Alkoxyl Radicals Can Give Rise to C-Terminal Decarboxylation and Backbone Cleavage. Arch. Biochem. Biophys. 1996, 336, 163–172. [Google Scholar] [CrossRef]
- Reeg, S.; Grune, T. Protein Oxidation in Aging: Does It Play a Role in Aging Progression? Antioxid. Redox Signal. 2015, 23, 239–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estévez, M.; Luna, C. Dietary protein oxidation: A silent threat to human health? Crit. Rev. Food Sci. Nutr. 2017, 57, 3781–3793. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Bi, Y.; Sowers, J.R.; Hetz, C.; Zhang, Y. Endoplasmic reticulum stress and unfolded protein response in cardiovascular diseases. Nat. Rev. Cardiol. 2021, 18, 499–521. [Google Scholar] [CrossRef]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef] [Green Version]
- Cenini, G.; Lloret, A.; Cascella, R. Oxidative Stress in Neurodegenerative Diseases: From a Mitochondrial Point of View. Oxidative Med. Cell. Longev. 2019, 2019, 2105607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharifi-Rad, M.; Lankatillake, C.; Dias, D.A.; Docea, A.O.; Mahomoodally, M.F.; Lobine, D.; Chazot, P.L.; Kurt, B.; Boyunegmez Tumer, T.; Catarina Moreira, A.; et al. Impact of Natural Compounds on Neurodegenerative Disorders: From Preclinical to Pharmacotherapeutics. J. Clin. Med. 2020, 9, 1061. [Google Scholar] [CrossRef] [Green Version]
- Marnett, L.J.; Riggins, J.N.; West, J.D. Endogenous generation of reactive oxidants and electrophiles and their reactions with DNA and protein. J. Clin. Investig. 2003, 111, 583–593. [Google Scholar] [CrossRef]
- Ohshima, H. Genetic and epigenetic damage induced by reactive nitrogen species: Implications in carcinogenesis. Toxicol. Lett. 2003, 140–141, 99–104. [Google Scholar] [CrossRef]
- Srinivas, U.S.; Tan, B.W.Q.; Vellayappan, B.A.; Jeyasekharan, A.D. ROS and the DNA damage response in cancer. Redox Biol. 2019, 25, 101084. [Google Scholar] [CrossRef]
- Kowalska, M.; Piekut, T.; Prendecki, M.; Sodel, A.; Kozubski, W.; Dorszewska, J. Mitochondrial and Nuclear DNA Oxidative Damage in Physiological and Pathological Aging. DNA Cell Biol. 2020, 39, 1410–1420. [Google Scholar] [CrossRef]
- Nissanka, N.; Moraes, C.T. Mitochondrial DNA damage and reactive oxygen species in neurodegenerative disease. FEBS Lett. 2018, 592, 728–742. [Google Scholar] [CrossRef]
- Inupakutika, M.A.; Sengupta, S.; Devireddy, A.R.; Azad, R.K.; Mittler, R. The evolution of reactive oxygen species metabolism. J. Exp. Bot. 2016, 67, 5933–5943. [Google Scholar] [CrossRef]
- Wagner, B.A.; Venkataraman, S.; Buettner, G.R. The rate of oxygen utilization by cells. Free Radic. Biol. Med. 2011, 51, 700–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS Sources in Physiological and Pathological Conditions. Oxid. Med. Cell. Longev. 2016, 2016, 1245049. [Google Scholar] [CrossRef]
- Amir Aslani, B.; Ghobadi, S. Studies on oxidants and antioxidants with a brief glance at their relevance to the immune system. Life Sci. 2016, 146, 163–173. [Google Scholar] [CrossRef]
- Lazzarino, G.; Listorti, I.; Bilotta, G.; Capozzolo, T.; Amorini, A.; Longo, S.; Caruso, G.; Lazzarino, G.; Tavazzi, B.; Bilotta, P. Water- and Fat-Soluble Antioxidants in Human Seminal Plasma and Serum of Fertile Males. Antioxidants 2019, 8, 96. [Google Scholar] [CrossRef] [Green Version]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Flynn, J.M.; Melov, S. SOD2 in mitochondrial dysfunction and neurodegeneration. Free Radic. Biol. Med. 2013, 62, 4–12. [Google Scholar] [CrossRef] [Green Version]
- Younus, H. Therapeutic potentials of superoxide dismutase. Int. J. Health Sci. 2018, 12, 88–93. [Google Scholar]
- Saadat, M. Association between C-262T genetic polymorphism at the promoter region of the catalase gene (CAT) and the risk of inflammatory bowel diseases: Evidence from meta-analysis. Gene Rep. 2019, 14, 114–117. [Google Scholar] [CrossRef]
- Cardoso, B.R.; Hare, D.J.; Bush, A.I.; Roberts, B.R. Glutathione peroxidase 4: A new player in neurodegeneration? Mol. Psychiatry 2016, 22, 328–335. [Google Scholar] [CrossRef] [Green Version]
- Brigelius-Flohé, R.; Flohé, L. Regulatory Phenomena in the Glutathione Peroxidase Superfamily. Antioxid. Redox Signal. 2020, 33, 498–516. [Google Scholar] [CrossRef]
- Park, M.H.; Jo, M.R.; Kim, Y.R.; Lee, C.-K.; Hong, J.T. Roles of peroxiredoxins in cancer, neurodegenerative diseases and inflammatory diseases. Pharmacol. Ther. 2016, 163, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Ye, J.; Kamphorst, J.J.; Shlomi, T.; Thompson, C.B.; Rabinowitz, J.D. Quantitative flux analysis reveals folate-dependent NADPH production. Nature 2014, 510, 298–302. [Google Scholar] [CrossRef] [Green Version]
- Ulrich, K.; Jakob, U. The role of thiols in antioxidant systems. Free Radic. Biol. Med. 2019, 140, 14–27. [Google Scholar] [CrossRef]
- Lohan, S.B.; Vitt, K.; Scholz, P.; Keck, C.M.; Meinke, M.C. ROS production and glutathione response in keratinocytes after application of β-carotene and VIS/NIR irradiation. Chem.-Biol. Interact. 2018, 280, 1–7. [Google Scholar] [CrossRef]
- Ziller, A.; Fraissinet-Tachet, L. Metallothionein diversity and distribution in the tree of life: A multifunctional protein. Metallomics 2018, 10, 1549–1559. [Google Scholar] [CrossRef]
- Kumari, M.V.R.; Hiramatsu, M.; Ebadi, M. Free radical scavenging actions of metallothionein isoforms I and II. Free Radic. Res. 1998, 29, 93–101. [Google Scholar] [CrossRef] [PubMed]
- AL-Megrin, W.A.; Soliman, D.; Kassab, R.B.; Metwally, D.M.; Moneim, A.E.A.; El-Khadragy, M.F. Coenzyme Q10 Activates the Antioxidant Machinery and Inhibits the Inflammatory and Apoptotic Cascades Against Lead Acetate-Induced Renal Injury in Rats. Front. Physiol. 2020, 11, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arenas-Jal, M.; Suñé-Negre, J.M.; García-Montoya, E. Coenzyme Q10 supplementation: Efficacy, safety, and formulation challenges. Compr. Rev. Food Sci. Food Saf. 2020, 19, 574–594. [Google Scholar] [CrossRef] [Green Version]
- Kocot, J.; Luchowska-Kocot, D.; Kiełczykowska, M.; Musik, I.; Kurzepa, J. Does Vitamin C Influence Neurodegenerative Diseases and Psychiatric Disorders? Nutrients 2017, 9, 659. [Google Scholar] [CrossRef] [Green Version]
- Ballaz, S.J.; Rebec, G.V. Neurobiology of vitamin C: Expanding the focus from antioxidant to endogenous neuromodulator. Pharmacol. Res. 2019, 146, 104321. [Google Scholar] [CrossRef] [PubMed]
- Al-Niaimi, F.; Chiang, N.Y.Z. Topical Vitamin C and the Skin: Mechanisms of Action and Clinical Applications. J. Clin. Aesthet. Dermatol. 2017, 10, 14–17. [Google Scholar]
- Salehi, B.; Rescigno, A.; Dettori, T.; Calina, D.; Docea, A.O.; Singh, L.; Cebeci, F.; Özçelik, B.; Bhia, M.; Dowlati Beirami, A.; et al. Avocado–Soybean Unsaponifiables: A Panoply of Potentialities to Be Exploited. Biomolecules 2020, 10, 130. [Google Scholar] [CrossRef] [Green Version]
- Lloret, A.; Esteve, D.; Monllor, P.; Cervera-Ferri, A.; Lloret, A. The Effectiveness of Vitamin E Treatment in Alzheimer’s Disease. Int. J. Mol. Sci. 2019, 20, 879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salehi, B.; Capanoglu, E.; Adrar, N.; Catalkaya, G.; Shaheen, S.; Jaffer, M.; Giri, L.; Suyal, R.; Jugran, A.K.; Calina, D.; et al. Cucurbits Plants: A Key Emphasis to Its Pharmacological Potential. Molecules 2019, 24, 1854. [Google Scholar] [CrossRef] [Green Version]
- Abdulhadi, S.Y.; Gergees, R.N.; Hasan, G.Q. Molecular Identification, Antioxidant Efficacy of Phenolic Compounds, and Antimicrobial Activity of Beta-Carotene Isolated from Fruiting Bodies of Suillus sp. Karbala Int. J. Mod. Sci. 2020, 6, 365–374. [Google Scholar] [CrossRef]
- Wang, T.; Li, Q.; Bi, K. Bioactive flavonoids in medicinal plants: Structure, activity and biological fate. Asian J. Pharm. Sci. 2018, 13, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Ali, L.; Mabood, F.; Rizvi, T.S.; Rehman, N.U.; Arman, M.; Al-Shidani, S.; Al-Abri, Z.; Hussain, J.; Al-Harrasi, A. Total polyphenols quantification in Acridocarpus orientalis and Moringa peregrina by using NIR spectroscopy coupled with PLS regression. Chem. Data Collect. 2018, 13–14, 104–112. [Google Scholar] [CrossRef]
- Chen, X.-X.; Leung, G.P.-H.; Zhang, Z.-J.; Xiao, J.-B.; Lao, L.-X.; Feng, F.; Mak, J.C.-W.; Wang, Y.; Sze, S.C.-W.; Zhang, K.Y.-B. Proanthocyanidins from Uncaria rhynchophylla induced apoptosis in MDA-MB-231 breast cancer cells while enhancing cytotoxic effects of 5-fluorouracil. Food Chem. Toxicol. 2017, 107, 248–260. [Google Scholar] [CrossRef]
- Chen, X.-X.; Lam, K.H.; Chen, Q.-X.; Leung, G.P.-H.; Tang, S.C.; Sze, S.C.-W.; Xiao, J.-B.; Feng, F.; Wang, Y.; Zhang, K.Y.-B.; et al. Ficus virens proanthocyanidins induced apoptosis in breast cancer cells concomitantly ameliorated 5-fluorouracil induced intestinal mucositis in rats. Food Chem. Toxicol. 2017, 110, 49–61. [Google Scholar] [CrossRef]
- Chen, L.; Teng, H.; Jia, Z.; Battino, M.; Miron, A.; Yu, Z.; Cao, H.; Xiao, J. Intracellular signaling pathways of inflammation modulated by dietary flavonoids: The most recent evidence. Crit. Rev. Food Sci. Nutr. 2017, 58, 2908–2924. [Google Scholar] [CrossRef]
- Chen, L.; Teng, H.; Xie, Z.; Cao, H.; Cheang, W.S.; Skalicka-Woniak, K.; Georgiev, M.I.; Xiao, J. Modifications of dietary flavonoids towards improved bioactivity: An update on structure–activity relationship. Crit. Rev. Food Sci. Nutr. 2017, 58, 513–527. [Google Scholar] [CrossRef] [PubMed]
- Chojnacka, K.; Lewandowska, U. Chemopreventive effects of polyphenol-rich extracts against cancer invasiveness and metastasis by inhibition of type IV collagenases expression and activity. J. Funct. Foods 2018, 46, 295–311. [Google Scholar] [CrossRef]
- Pan, P.; Huang, Y.-W.; Oshima, K.; Yearsley, M.; Zhang, J.; Yu, J.; Arnold, M.; Wang, L.-S. An immunological perspective for preventing cancer with berries. J. Berry Res. 2018, 8, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Reboredo-Rodríguez, P. Potential roles of berries in the prevention of breast cancer progression. J. Berry Res. 2018, 8, 307–323. [Google Scholar] [CrossRef]
- Rady, H.M.; Hemmaid, K.Z.; Esmaeil, N.N.; Eid, M.M.; Elshat, A.A. Sidr Kashmiry honey and its fractions induced apoptosis in hepatocellular carcinoma in vitro. Mediterr. J. Nutr. Metab. 2018, 11, 343–351. [Google Scholar] [CrossRef]
- Kaushik, S.; Shyam, H.; Agarwal, S.; Sharma, R.; Nag, T.C.; Dwivedi, A.K.; Balapure, A.K. Genistein potentiates Centchroman induced antineoplasticity in breast cancer via PI3K/Akt deactivation and ROS dependent induction of apoptosis. Life Sci. 2019, 239, 117073. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Jin, S.; Zhang, Q.Y.; Kang, X.M.; Wang, J.X.; Zhao, W.H. Daidzein induces MCF-7 breast cancer cell apoptosis via the mitochondrial pathway. Ann. Oncol. 2010, 21, 263–268. [Google Scholar] [CrossRef]
- Pandey, P.; Sayyed, U.; Tiwari, R.K.; Siddiqui, M.H.; Pathak, N.; Bajpai, P. Hesperidin Induces ROS-Mediated Apoptosis along with Cell Cycle Arrest at G2/M Phase in Human Gall Bladder Carcinoma. Nutr. Cancer 2018, 71, 676–687. [Google Scholar] [CrossRef]
- Palit, S.; Kar, S.; Sharma, G.; Das, P.K. Hesperetin Induces Apoptosis in Breast Carcinoma by Triggering Accumulation of ROS and Activation of ASK1/JNK Pathway. J. Cell. Physiol. 2015, 230, 1729–1739. [Google Scholar] [CrossRef]
- Park, S.; Lim, W.; Bazer, F.W.; Song, G. Naringenin suppresses growth of human placental choriocarcinoma via reactive oxygen species-mediated P38 and JNK MAPK pathways. Phytomedicine 2018, 50, 238–246. [Google Scholar] [CrossRef]
- Wu, D.; Zhang, J.; Wang, J.; Li, J.; Liao, F.; Dong, W. Hesperetin induces apoptosis of esophageal cancer cells via mitochondrial pathway mediated by the increased intracellular reactive oxygen species. Tumor Biol. 2015, 37, 3451–3459. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.-B.; Kim, J.-H.; Lee, H.; Pak, J.-N.; Shim, B.S.; Kim, S.-H. Reactive Oxygen Species and p53 Mediated Activation of p38 and Caspases is Critically Involved in Kaempferol Induced Apoptosis in Colorectal Cancer Cells. J. Agric. Food Chem. 2018, 66, 9960–9967. [Google Scholar] [CrossRef]
- Seydi, E.; Salimi, A.; Rasekh, H.R.; Mohsenifar, Z.; Pourahmad, J. Selective Cytotoxicity of Luteolin and Kaempferol on Cancerous Hepatocytes Obtained from Rat Model of Hepatocellular Carcinoma: Involvement of ROS-Mediated Mitochondrial Targeting. Nutr. Cancer 2018, 70, 594–604. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Zhang, T.; Ren, L.; Jing, S.; Li, Z.; Zuo, P.; Li, T.; Wang, Y.; Zhang, J.; Wei, Z. Cucurbitacin IIb induces apoptosis and cell cycle arrest through regulating EGFR/MAPK pathway. Environ. Toxicol. Pharmacol. 2021, 81, 103542. [Google Scholar] [CrossRef]
- Chae, H.-S.; Xu, R.; Won, J.-Y.; Chin, Y.-W.; Yim, H. Molecular Targets of Genistein and Its Related Flavonoids to Exert Anticancer Effects. Int. J. Mol. Sci. 2019, 20, 2420. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Cheng, X.; Chen, C.; Huijuan, W.; Zhao, H.; Liu, W.; Xiang, Z.; Wang, Q. Apigenin, a flavonoid constituent derived from P. villosa, inhibits hepatocellular carcinoma cell growth by CyclinD1/CDK4 regulation via p38 MAPK-p21 signaling. Pathol. Res. Pract. 2020, 216, 152701. [Google Scholar] [CrossRef]
- Zhang, H.-W.; Hu, J.-J.; Fu, R.-Q.; Liu, X.; Zhang, Y.-H.; Li, J.; Liu, L.; Li, Y.-N.; Deng, Q.; Luo, Q.-S.; et al. Flavonoids inhibit cell proliferation and induce apoptosis and autophagy through downregulation of PI3Kγ mediated PI3K/AKT/mTOR/p70S6K/ULK signaling pathway in human breast cancer cells. Sci. Rep. 2018, 8, 11255. [Google Scholar] [CrossRef]
- De Sousa Moraes, L.F.; Sun, X.; do Peluzio, M.; Zhu, M.-J. Anthocyanins/anthocyanidins and colorectal cancer: What is behind the scenes? Crit. Rev. Food Sci. Nutr. 2017, 59, 59–71. [Google Scholar] [CrossRef]
- Kashafi, E.; Moradzadeh, M.; Mohamadkhani, A.; Erfanian, S. Kaempferol increases apoptosis in human cervical cancer HeLa cells via PI3K/AKT and telomerase pathways. Biomed. Pharmacother. 2017, 89, 573–577. [Google Scholar] [CrossRef]
- Moradzadeh, M.; Hosseini, A.; Erfanian, S.; Rezaei, H. Epigallocatechin-3-gallate promotes apoptosis in human breast cancer T47D cells through down-regulation of PI3K/AKT and Telomerase. Pharmacol. Rep. 2017, 69, 924–928. [Google Scholar] [CrossRef]
- Elango, R.; Athinarayanan, J.; Subbarayan, V.P.; Lei, D.K.Y.; Alshatwi, A.A. Hesperetin induces an apoptosis-triggered extrinsic pathway and a p53-independent pathway in human lung cancer H522 cells. J. Asian Nat. Prod. Res. 2017, 20, 559–569. [Google Scholar] [CrossRef]
- Danciu, C.; Avram, S.; Pavel, I.Z.; Ghiulai, R.; Dehelean, C.A.; Ersilia, A.; Minda, D.; Petrescu, C.; Moaca, E.-A.; Soica, C. Main Isoflavones Found in Dietary Sources as Natural Anti-inflammatory Agents. Curr. Drug Targets 2018, 19, 841–853. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.; Reale, M.; Ullah, H.; Sureda, A.; Tejada, S.; Wang, Y.; Zhang, Z.-J.; Xiao, J. Anti-cancer effects of polyphenols via targeting p53 signaling pathway: Updates and future directions. Biotechnol. Adv. 2020, 38, 107385. [Google Scholar] [CrossRef]
- Khan, M.K.; Ahmad, K.; Hassan, S.; Imran, M.; Ahmad, N.; Xu, C. Effect of novel technologies on polyphenols during food processing. Innov. Food Sci. Emerg. Technol. 2018, 45, 361–381. [Google Scholar] [CrossRef]
- Majidinia, M.; Karimian, A.; Alemi, F.; Yousefi, B.; Safa, A. Targeting miRNAs by polyphenols: Novel therapeutic strategy for aging. Biochem. Pharmacol. 2020, 173, 113688. [Google Scholar] [CrossRef]
- Ding, S.; Jiang, H.; Fang, J. Regulation of Immune Function by Polyphenols. J. Immunol. Res. 2018, 2018, 1264074. [Google Scholar] [CrossRef] [Green Version]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The Immunomodulatory and Anti-Inflammatory Role of Polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef] [Green Version]
- Boshtam, M.; Asgary, S.; Kouhpayeh, S.; Shariati, L.; Khanahmad, H. Aptamers Against Pro- and Anti-Inflammatory Cytokines: A Review. Inflammation 2016, 40, 340–349. [Google Scholar] [CrossRef]
- Crouvezier, S.; Powell, B.; Keir, D.; Yaqoob, P. The effects of phenolic components of tea on the production of pro- and anti-inflammatory cytokines by human leukocytes in vitro. Cytokine 2001, 13, 280–286. [Google Scholar] [CrossRef]
- Essafi-Benkhadir, K.; Refai, A.; Riahi, I.; Fattouch, S.; Karoui, H.; Essafi, M. Quince (Cydonia oblonga Miller) peel polyphenols modulate LPS-induced inflammation in human THP-1-derived macrophages through NF-κB, p38MAPK and Akt inhibition. Biochem. Biophys. Res. Commun. 2012, 418, 180–185. [Google Scholar] [CrossRef]
- Fitó, M.; Cladellas, M.; de la Torre, R.; Martí, J.; Muñoz, D.; Schröder, H.; Alcántara, M.; Pujadas-Bastardes, M.; Marrugat, J.; López-Sabater, M.C.; et al. Anti-inflammatory effect of virgin olive oil in stable coronary disease patients: A randomized, crossover, controlled trial. Eur. J. Clin. Nutr. 2007, 62, 570–574. [Google Scholar] [CrossRef] [Green Version]
- Kolehmainen, M.; Mykkänen, O.; Kirjavainen, P.V.; Leppänen, T.; Moilanen, E.; Adriaens, M.; Laaksonen, D.E.; Hallikainen, M.; Puupponen-Pimiä, R.; Pulkkinen, L.; et al. Bilberries reduce low-grade inflammation in individuals with features of metabolic syndrome. Mol. Nutr. Food Res. 2012, 56, 1501–1510. [Google Scholar] [CrossRef] [Green Version]
- Sharma, V.; Mishra, M.; Ghosh, S.; Tewari, R.; Basu, A.; Seth, P.; Sen, E. Modulation of interleukin-1β mediated inflammatory response in human astrocytes by flavonoids: Implications in neuroprotection. Brain Res. Bull. 2007, 73, 55–63. [Google Scholar] [CrossRef]
- Ren, J.; Lu, Y.; Qian, Y.; Chen, B.; Wu, T.; Ji, G. Recent progress regarding kaempferol for the treatment of various diseases. Exp. Ther. Med. 2019, 18, 2759–2776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, H.; Hao, J.; Liu, T.; Zhang, D.; Lv, H.; Song, E.; Zhu, C. Hesperetin Suppresses Inflammatory Responses in Lipopolysaccharide-Induced RAW 264.7 Cells via the Inhibition of NF-κB and Activation of Nrf2/HO-1 Pathways. Inflammation 2016, 39, 964–973. [Google Scholar] [CrossRef] [PubMed]
- Lyu, S.-Y.; Park, W.-B. Production of Cytokine and NO by RAW 264.7 Macrophages and PBMC In Vitro Incubation with Flavonoids. Arch. Pharmacal Res. 2005, 28, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Maurya, A.K.; Vinayak, M. Quercetin Attenuates Cell Survival, Inflammation, and Angiogenesis via Modulation of AKT Signaling in Murine T-Cell Lymphoma. Nutr. Cancer 2017, 69, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, Y.; Kanatsu, J.; Toh, M.; Naka, A.; Kondo, K.; Iida, K. The Dietary Isoflavone Daidzein Reduces Expression of Pro-Inflammatory Genes through PPARα/γ and JNK Pathways in Adipocyte and Macrophage Co-Cultures. PLoS ONE 2016, 11, e0149676. [Google Scholar] [CrossRef]
- Venancio, V.P.; Cipriano, P.A.; Kim, H.; Antunes, L.M.; Talcott, S.T.; Mertens-Talcott, S.U. Cocoplum (Chrysobalanus icaco L.) anthocyanins exert anti-inflammatory activity in human colon cancer and non-malignant colon cells. Food Funct. 2017, 8, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Ai, X.-Y.; Qin, Y.; Liu, H.-J.; Cui, Z.-H.; Li, M.; Yang, J.-H.; Zhong, W.-L.; Liu, Y.-R.; Chen, S.; Sun, T.; et al. Apigenin inhibits colonic inflammation and tumorigenesis by suppressing STAT3-NF-κB signaling. Oncotarget 2017, 8, 100216–100226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xagorari, A.; Roussos, C.; Papapetropoulos, A. Inhibition of LPS-stimulated pathways in macrophages by the flavonoid luteolin. Br. J. Pharmacol. 2002, 136, 1058–1064. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.C.; Chow, M.-P.; Huang, W.-C.; Lin, Y.-C.; Chang, Y.-J. Flavonoids inhibit tumor necrosis factor-alpha-induced up-regulation of intercellular adhesion molecule-1 (ICAM-1) in respiratory epithelial cells through activator protein-1 and nuclear factor-kappaB: Structure-activity relationships. Mol. Pharm. 2004, 66, 683–693. [Google Scholar]
- Wadsworth, T.L.; McDonald, T.L.; Koop, D.R. Effects of Ginkgo biloba extract (EGb 761) and quercetin on lipopolysaccharide-induced signaling pathways involved in the release of tumor necrosis factor-α. Biochem. Pharmacol. 2001, 62, 963–974. [Google Scholar] [CrossRef]
- Sundaram, M.K.; Unni, S.; Somvanshi, P.; Bhardwaj, T.; Mandal, R.K.; Hussain, A.; Haque, S. Genistein Modulates Signaling Pathways and Targets Several Epigenetic Markers in HeLa Cells. Genes 2019, 10, 955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pons, D.G.; Vilanova-Llompart, J.; Gaya-Bover, A.; Alorda-Clara, M.; Oliver, J.; Roca, P.; Sastre-Serra, J. The phytoestrogen genistein affects inflammatory-related genes expression depending on the ERα/ERβ ratio in breast cancer cells. Int. J. Food Sci. Nutr. 2019, 70, 941–949. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Jin, G.; Ge, Y.; Guo, Z. Naringenin inhibits migration of breast cancer cells via inflammatory and apoptosis cell signaling pathways. Inflammopharmacology 2019, 27, 1021–1036. [Google Scholar] [CrossRef]
- Shirakami, Y.; Sakai, H.; Kochi, T.; Seishima, M.; Shimizu, M. Catechins and Its Role in Chronic Diseases. Adv. Exp. Med. Biol. 2016, 929, 67–90. [Google Scholar]
- Ginwala, R.; Bhavsar, R.; Chigbu, D.G.I.; Jain, P.; Khan, Z.K. Potential Role of Flavonoids in Treating Chronic Inflammatory Diseases with a Special Focus on the Anti-Inflammatory Activity of Apigenin. Antioxidants 2019, 8, 35. [Google Scholar] [CrossRef] [Green Version]
- Ghițu, A.; Schwiebs, A.; Radeke, H.H.; Avram, S.; Zupko, I.; Bor, A.; Pavel, I.Z.; Dehelean, C.A.; Oprean, C.; Bojin, F.; et al. A Comprehensive Assessment of Apigenin as an Antiproliferative, Proapoptotic, Antiangiogenic and Immunomodulatory Phytocompound. Nutrients 2019, 11, 858. [Google Scholar] [CrossRef] [Green Version]
- Ahangarpour, A.; Sayahi, M.; Sayahi, M. The antidiabetic and antioxidant properties of some phenolic phytochemicals: A review study. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 854–857. [Google Scholar] [CrossRef]
- Wang, Y.; Alkhalidy, H.; Liu, D. The Emerging Role of Polyphenols in the Management of Type 2 Diabetes. Molecules 2021, 26, 703. [Google Scholar] [CrossRef]
- Rouse, M.; Younès, A.; Egan, J.M. Resveratrol and curcumin enhance pancreatic β-cell function by inhibiting phosphodiesterase activity. J. Endocrinol. 2014, 223, 107–117. [Google Scholar] [CrossRef]
- Vetterli, L.; Brun, T.; Giovannoni, L.; Bosco, D.; Maechler, P. Resveratrol Potentiates Glucose-stimulated Insulin Secretion in INS-1E β-Cells and Human Islets through a SIRT1-dependent Mechanism. J. Biol. Chem. 2011, 286, 6049–6060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowley, T.J.; Bitner, B.F.; Ray, J.D.; Lathen, D.R.; Smithson, A.T.; Dallon, B.W.; Plowman, C.J.; Bikman, B.T.; Hansen, J.M.; Dorenkott, M.R.; et al. Monomeric cocoa catechins enhance β-cell function by increasing mitochondrial respiration. J. Nutr. Biochem. 2017, 49, 30–41. [Google Scholar] [CrossRef]
- Lee, H.; Im, S.W.; Jung, C.H.; Jang, Y.J.; Ha, T.Y.; Ahn, J. Tyrosol, an olive oil polyphenol, inhibits ER stress-induced apoptosis in pancreatic β-cell through JNK signaling. Biochem. Biophys. Res. Commun. 2016, 469, 748–752. [Google Scholar] [CrossRef]
- Belhadj, S.; Hentati, O.; Hamdaoui, G.; Fakhreddine, K.; Maillard, E.; Dal, S.; Sigrist, S. Beneficial Effect of Jojoba Seed Extracts on Hyperglycemia-Induced Oxidative Stress in RINm5f Beta Cells. Nutrients 2018, 10, 384. [Google Scholar] [CrossRef] [Green Version]
- Cai, S.; Sun, W.; Fan, Y.; Guo, X.; Xu, G.; Xu, T.; Hou, Y.; Zhao, B.; Feng, X.; Liu, T. Effect of mulberry leaf (Folium Mori) on insulin resistance via IRS-1/PI3K/Glut-4 signalling pathway in type 2 diabetes mellitus rats. Pharm. Biol. 2016, 54, 2685–2691. [Google Scholar] [CrossRef] [Green Version]
- Bryant, N.J.; Gould, G.W. Insulin stimulated GLUT4 translocation—Size is not everything! Curr. Opin. Cell Biol. 2020, 65, 28–34. [Google Scholar] [CrossRef]
- Ayeleso, T.; Ramachela, K.; Mukwevho, E. Aqueous-Methanol Extracts of Orange-Fleshed Sweet Potato (Ipomoea batatas) Ameliorate Oxidative Stress and Modulate Type 2 Diabetes Associated Genes in Insulin Resistant C2C12 Cells. Molecules 2018, 23, 2058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Modi, S.; Yaluri, N.; Kokkola, T.; Laakso, M. Plant-derived compounds strigolactone GR24 and pinosylvin activate SIRT1 and enhance glucose uptake in rat skeletal muscle cells. Sci. Rep. 2017, 7, 17606. [Google Scholar] [CrossRef] [Green Version]
- Ooi, D.J.; Adamu, H.A.; Imam, M.U.; Ithnin, H.; Ismail, M. Polyphenol-rich ethyl acetate fraction isolated from Molineria latifolia ameliorates insulin resistance in experimental diabetic rats via IRS1/AKT activation. Biomed. Pharmacother. 2018, 98, 125–133. [Google Scholar] [CrossRef]
- Kikuchi, A.; Takamura, T. Where does liver fat go? A possible molecular link between fatty liver and diabetes. J. Diabetes Investig. 2016, 8, 152–154. [Google Scholar] [CrossRef] [Green Version]
- Waltner-Law, M.E.; Wang, X.L.; Law, B.K.; Hall, R.K.; Nawano, M.; Granner, D.K. Epigallocatechin Gallate, a Constituent of Green Tea, Represses Hepatic Glucose Production. J. Biol. Chem. 2002, 277, 34933–34940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, D.M.; Kuhn, P.; Poulev, A.; Rojo, L.E.; Lila, M.A.; Raskin, I. In vivo and in vitro antidiabetic effects of aqueous cinnamon extract and cinnamon polyphenol-enhanced food matrix. Food Chem. 2012, 135, 2994–3002. [Google Scholar] [CrossRef] [Green Version]
- Imam, M.U.; Ismail, M. Nutrigenomic effects of germinated brown rice and its bioactives on hepatic gluconeogenic genes in type 2 diabetic rats and HEPG2 cells. Mol. Nutr. Food Res. 2013, 57, 401–411. [Google Scholar] [CrossRef]
- Do, G.-M.; Jung, U.J.; Park, H.-J.; Kwon, E.-Y.; Jeon, S.-M.; McGregor, R.A.; Choi, M.-S. Resveratrol ameliorates diabetes-related metabolic changes via activation of AMP-activated protein kinase and its downstream targets in db/db mice. Mol. Nutr. Food Res. 2012, 56, 1282–1291. [Google Scholar] [CrossRef] [PubMed]
- Jung, U.J.; Lee, M.-K.; Park, Y.B.; Kang, M.A.; Choi, M.-S. Effect of citrus flavonoids on lipid metabolism and glucose-regulating enzyme mRNA levels in type-2 diabetic mice. Int. J. Biochem. Cell Biol. 2006, 38, 1134–1145. [Google Scholar] [CrossRef]
- Wei, J.; Wu, H.; Zhang, H.; Li, F.; Chen, S.; Hou, B.; Shi, Y.; Zhao, L.; Duan, H. Anthocyanins inhibit high glucose-induced renal tubular cell apoptosis caused by oxidative stress in db/db mice. Int. J. Mol. Med. 2018, 41, 1608–1618. [Google Scholar] [CrossRef]
- Cordero Herrera, I.; Chen, X.; Devaraj, S. (–)-epicatechin attenuates high glucose-induced inflammation by epigenetic modulation in human monocytes. Atherosclerosis 2015, 241, 1369–1373. [Google Scholar] [CrossRef]
- Diarra, M.S.; Hassan, Y.I.; Block, G.S.; Drover, J.C.G.; Delaquis, P.; Oomah, B.D. Antibacterial activities of a polyphenolic-rich extract prepared from American cranberry (Vaccinium macrocarpon) fruit pomace against Listeria spp. LWT 2020, 123, 109056. [Google Scholar] [CrossRef]
- Guo, L.; Gong, S.; Wang, Y.; Sun, Q.; Duo, K.; Fei, P. Antibacterial Activity of Olive Oil Polyphenol Extract Against Salmonella Typhimurium and Staphylococcus aureus: Possible Mechanisms. Foodborne Pathog. Dis. 2020, 17, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.-L.; Chen, G.; Chen, S.-N.; Wang, Q.-R.; Wan, L.; Jian, S.-P. Characterization of polyphenolic constituents from Sanguisorba officinalis L. and its antibacterial activity. Eur. Food Res. Technol. 2019, 245, 1487–1498. [Google Scholar] [CrossRef]
- Venkatesh, T.; Bodke, Y.D. Synthesis, Antimicrobial and Antioxidant Activity of Chalcone Derivatives Containing Thiobarbitone Nucleus. Med. Chem. 2016, 6, 7. [Google Scholar] [CrossRef]
- Govindappa, M.; Hemashekhar, B.; Arthikala, M.-K.; Ravishankar, R.V.; Ramachandra, Y.L. Characterization, antibacterial, antioxidant, antidiabetic, anti-inflammatory and antityrosinase activity of green synthesized silver nanoparticles using Calophyllum tomentosum leaves extract. Results Phys. 2018, 9, 400–408. [Google Scholar] [CrossRef]
- Rosas-Burgos, E.C.; Burgos-Hernández, A.; Noguera-Artiaga, L.; Kačániová, M.; Hernández-García, F.; Cárdenas-López, J.L.; Carbonell-Barrachina, Á.A. Antimicrobial activity of pomegranate peel extracts as affected by cultivar. J. Sci. Food Agric. 2016, 97, 802–810. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.-S.; Oh, J.J.; Oh, K.-H. Antimicrobial activity and biofilm formation inhibition of green tea polyphenols on human teeth. Biotechnol. Bioprocess. Eng. 2010, 15, 359–364. [Google Scholar] [CrossRef]
- Pourhosseini, S.H.; Ahadi, H.; Aliahmadi, A.; Mirjalili, M.H. Chemical Composition and Antibacterial Activity of the Carvacrol-rich Essential Oils of Zataria multiflora Boiss. (Lamiaceae) from Southern Natural Habitats of Iran. J. Essent. Oil Bear. Plants 2020, 23, 779–787. [Google Scholar] [CrossRef]
- Bouarab-Chibane, L.; Forquet, V.; Lantéri, P.; Clément, Y.; Léonard-Akkari, L.; Oulahal, N.; Degraeve, P.; Bordes, C. Antibacterial Properties of Polyphenols: Characterization and QSAR (Quantitative Structure–Activity Relationship) Models. Front. Microbiol. 2019, 10, 829. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; He, M.; Zang, X.; Zhou, Y.; Qiu, T.; Pan, S.; Xu, X. A structure–activity relationship study of flavonoids as inhibitors of E. coli by membrane interaction effect. Biochim. Biophys. Acta BBA Biomembr. 2013, 1828, 2751–2756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konaté, K.; Hilou, A.; Mavoungou, J.; Lepengué, A.; Souza, A.; Barro, N.; Datté, J.Y.; M’Batchi, B.; Nacoulma, O. Antimicrobial activity of polyphenol-rich fractions from Sida alba L. (Malvaceae) against co-trimoxazol-resistant bacteria strains. Ann. Clin. Microbiol. Antimicrob. 2012, 11, 5. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, D.; Ghosh, D.; Bhattacharya, S.; Sarkar, S.; Karmakar, P.; Koley, H.; Gachhui, R. Antibacterial activity of polyphenolic fraction of Kombucha against Vibrio cholerae: Targeting cell membrane. Lett. Appl. Microbiol. 2018, 66, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.-D.; Qian, H.-L.; Feng, C.-M.; Wang, T.; Guo, Z.-Y.; Wu, X.-K.; Zhang, S.-H. Study on the Mechanism of Epigallocatechin Gallate (EGCG) to the Cell Membrane of Escherichia coli. Sci. Adv. Mater. 2019, 11, 262–268. [Google Scholar] [CrossRef]
- Antolak, H.; Czyzowska, A.; Sakač, M.; Mišan, A.; Đuragić, O.; Kregiel, D. Phenolic Compounds Contained in Little-known Wild Fruits as Antiadhesive Agents Against the Beverage-Spoiling Bacteria Asaia spp. Molecules 2017, 22, 1256. [Google Scholar] [CrossRef]
- Pellegrini, M.C.; Ponce, A.G. Beet (Beta vulgaris) and Leek (Allium porrum) Leaves as a Source of Bioactive Compounds with Anti-quorum Sensing and Anti-biofilm Activity. Waste Biomass Valoriz. 2019, 11, 4305–4313. [Google Scholar] [CrossRef]
- Xiong, L.-G.; Chen, Y.-J.; Tong, J.-W.; Huang, J.-A.; Li, J.; Gong, Y.-S.; Liu, Z.-H. Tea polyphenol epigallocatechin gallate inhibits Escherichia coli by increasing endogenous oxidative stress. Food Chem. 2017, 217, 196–204. [Google Scholar] [CrossRef]
- Tang, Q.-L.; Kang, A.-R.; Lu, C.-X. Phytochemical Analysis, Antibacterial Activity and Mode of Action of the Methanolic Extract of Scutellaria barbata Against Various Clinically Important Bacterial Pathogens. Int. J. Pharmacol. 2016, 12, 116–125. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, S.; Liang, W.; Mei, J.; Di, Y.; Lan, H.; Yang, Y.; Wang, W.; Luo, Y.; Wang, H. Antibacterial Activity and Mode of Action of Mentha arvensis Ethanol Extract against Multidrug-Resistant Acinetobacter baumannii. Trop. J. Pharm. Res. 2015, 14, 2099. [Google Scholar] [CrossRef] [Green Version]
- Ulrey, R.K.; Barksdale, S.M.; Zhou, W.; van Hoek, M.L. Cranberry proanthocyanidins have anti-biofilm properties against Pseudomonas aeruginosa. BMC Complement. Altern. Med. 2014, 14, 499. [Google Scholar] [CrossRef] [Green Version]
- Yi, S.; Zhu, J.; Fu, L.; Li, J. Tea polyphenols inhibit Pseudomonas aeruginosa through damage to the cell membrane. Int. J. Food Microbiol. 2010, 144, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Zang, X.; He, M.; Pan, S.; Xu, X. Structure–Activity Relationship of Flavonoids on Their Anti-Escherichia coli Activity and Inhibition of DNA Gyrase. J. Agric. Food Chem. 2013, 61, 8185–8190. [Google Scholar] [CrossRef] [PubMed]
- Chinnam, N.; Dadi, P.K.; Sabri, S.A.; Ahmad, M.; Kabir, M.A.; Ahmad, Z. Dietary bioflavonoids inhibit Escherichia coli ATP synthase in a differential manner. Int. J. Biol. Macromol. 2010, 46, 478–486. [Google Scholar] [CrossRef] [Green Version]
- Dadi, P.K.; Ahmad, M.; Ahmad, Z. Inhibition of ATPase activity of Escherichia coli ATP synthase by polyphenols. Int. J. Biol. Macromol. 2009, 45, 72–79. [Google Scholar] [CrossRef]
- Małecka, M.; Skoczyńska, A.; Goodman, D.M.; Hartinger, C.G.; Budzisz, E. Biological properties of ruthenium(II)/(III) complexes with flavonoids as ligands. Coord. Chem. 2021, 436, 213849. [Google Scholar] [CrossRef]
- Chen, G.L.; Fan, M.X.; Wu, J.L.; Li, N.; Guo, M.Q. Antioxidant and antiinflammatory properties of flavonoids from lotus plumule. Food Chem. 2019, 277, 706–712. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jiang, Q.; Wang, T.; Liu, J.; Chen, D. Comparison of the antioxidant effects of quercitrin and isoquercitrin: Understanding the role of the 6″-OH group. Molecules 2016, 21, 1246. [Google Scholar] [CrossRef] [PubMed]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Zhao, B.; Li, M.; Shen, S.; Xinet, W. Studies on protective mechanism of four components of green tea polyphenols against lipid peroxidation in synaptosomes. Biochim. Biophys. Acta 1996, 1304, 210–222. [Google Scholar] [CrossRef]
- Abu-Amsha, R.; Croft, K.D.; Puddey, I.B.; Proudfoot, J.M.; Beilin, L.J. Phenolic contetnt of various beverages determines the extent of inhibition of human serum and LDL oxidation in vitro: Identification and mechanism of action of some cinnamic acid derivatives from red wine. Clin. Sci. 1996, 91, 449–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harborne, J.B.; Williams, C.A. Advances in flavonoid research since 1992. Phytochemistry 2000, 55, 481–504. [Google Scholar] [CrossRef]
- Nagao, A.; Seki, M.; Kobayashi, H. Inhibition of xanthine oxidase by flavonoids. Biosci. Biotechnol. Biochem. 1999, 63, 1787–1790. [Google Scholar] [CrossRef]
- Aucamp, J.; Gaspar, A.; Hara, Y.; Apostolides, Z. Inhibibition of xantine oxidase by catechins from tea (Camelia sinensis). Anticancer Res. 1997, 17, 4381–4385. [Google Scholar]
- Hodnick, W.F.; Duval, D.L.; Pardiniet, R.S. Inhibition of mitochondrial respiration and cyanide-stimulated generation of ROS by selective flavonoids. Biochem. Pharmacol. 1994, 47, 573–580. [Google Scholar] [CrossRef]
- Galleggiante, V.; de Santis, S.; Cavalcanti, E.; Scarano, A.; de Benedictis, M.; Serino, G.; Chieppa, M. Dendritic cells modulate iron homeostasis and inflammatory abilities following quercetin exposure. Curr. Pharm. Des. 2017, 23, 2139–2146. [Google Scholar] [CrossRef]
- Del Bo’, C.; Roursgaard, M.; Porrini, M.; Loft, S.; Møller, P.; Riso, P. Different effects of anthocyanins and phenolic acids from wild blueberry (Vaccinium angustifolium) on monocytes adhesion to endothelial cells in a TNF-α stimulated proinflammatory environment. Mol. Nutr. Food Res. 2016, 60, 2355–2366. [Google Scholar] [CrossRef]
- Gong, J.H.; Shin, D.; Han, S.Y.; Kim, J.L.; Kang, Y.H. Kaempferol suppresses eosionphil infiltration and airway inflammation in airway epithelial cells and in mice with allergic asthma, 2. J. Nutr. 2011, 142, 47–56. [Google Scholar] [CrossRef]
- Weng, Z.; Zhang, B.; Asadi, S.; Sismanopoulos, N.; Butcher, A.; Fu, X.; Theoharides, T.C. Quercetin is more effective than cromolyn in blocking human mast cell cytokine release and inhibits contact dermatitis and photosensitivity in humans. PLoS ONE 2012, 7, e33805. [Google Scholar] [CrossRef] [PubMed]
- Weng, Z.; Patel, A.B.; Panagiotidou, S.; Theoharides, T.C. The novel flavone tetramethoxyluteolin is a potent inhibitor of human mast cells. J. Allergy Clin. Immunol. 2015, 135, 1044–1052. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Yu, Q.; Zhao, W.; Zhang, J.; Liu, W.; Huang, M.; Zeng, X. Oligomeric proanthocyanidins attenuate airway inflammation in asthma by inhibiting dendritic cells maturation. Mol. Immunol. 2017, 91, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Kimata, M.; Shichijo, M.; Miura, T.; Serizawa, I.; Inagaki, N.; Nagai, H. Effects of luteolin, quercetin and baicalein on immunoglobulin E-mediated mediator release from human cultured mast cells. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 2000, 30, 501–508. [Google Scholar] [CrossRef]
- Rogerio, A.P.; Dora, C.L.; Andrade, E.L.; Chaves, J.S.; Silva, L.F.; Lemos-Senna, E.; Calixto, J.B. Anti-inflammatory effect of quercetin-loaded microemulsion in the airways allergic inflammatory model in mice. Pharmacol. Res. 2010, 61, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Wei, D.; Bian, T.; Xie, P.; Zou, J.; Mu, H.; Zhou, X. Genistein attenuated allergic airway inflammation by modulating the transcription factors T-bet, GATA-3 and STAT-6 in a murine model of asthma. Pharmacology 2012, 89, 229–236. [Google Scholar] [CrossRef]
- Liu, C.; Zhu, L.; Fukuda, K.; Ouyang, S.; Chen, X.; Wang, C.; Li, X. The flavonoid cyanidin blocks binding of the cytokine interleukin-17A to the IL-17RA subunit to alleviate inflammation in vivo. Sci. Signal. 2017, 10, eaaf8823. [Google Scholar] [CrossRef] [Green Version]
- Lacy, A.; O’Kennedy, R. Studies on Coumarins and Coumarin related compounds to determine their therapeutic role in the treatment of cancer. Curr. Pharm. Des. 2004, 10, 3797–3811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehlers, D.; Pfister, M.; Bork, W.; Nadolny, P. HPLC analysis of tonka bean extracts. Eur. Food Res. Technol. 1995, 201, 278–282. [Google Scholar] [CrossRef]
- Venugopala, K.N.; Rashmi, V.; Odhav, B. Review on natural coumarin lead compounds for their pharmacological activity. Biomed. Res. Int. 2013, 2013, 963248. [Google Scholar] [CrossRef] [Green Version]
- Skoczyńska, A.; Pastuszko, A.; Budzisz, E. Kumarynu—Właściwości i zastosowanie w kosmetologii i medycynie. Pol. J. Cosmetol. 2014, 17, 2–13. [Google Scholar]
- Lin, H.C.; Tsai, S.H.; Chen, C.S.; Chang, Y.C.; Lee, C.M.; Lai, Z.Y.; Lin, C.M. Structure-activity relationship of coumarin derivatives on xanthine oxidase-inhibiting and free radical-scavenging activities. Biochem. Pharmacol. 2008, 75, 1416–1425. [Google Scholar] [CrossRef] [PubMed]
- Costantino, L.; Rastelli, G.; Albasini, A. Natural polyhydroxylated compounds as inhibitors of xanthineoxidase. Pharmazie 1996, 51, 994–995. [Google Scholar] [PubMed]
- Lee, B.C.; Lee, S.Y.; Lee, H.J.; Sim, G.S.; Kim, J.H.; Cho, Y.H.; Lee, D.H.; Pyo, H.B.; Choe, T.B.; Moon, D.C.; et al. Anti-oxidative and Photo-protective Effects of Coumarins Isolated from Fraxinus chinensis. Arch. Pharm. Res. 2007, 30, 1293–1301. [Google Scholar] [CrossRef]
- Ferrari, A.M.; Sgobba, M.; Gamberini, M.C.; Rastelli, G. Relationship between quantum-chemical descriptors of proton dissociation and experimental acidity constants of various hydroxylated coumarins. Identification of the biologically active species for xanthine oxidase inhibition. Eur. J. Med. Chem. 2007, 42, 1028–1031. [Google Scholar] [CrossRef]
- Zhang, H.Y. Structure-Activity Relationships and Rational Design Strategies for Radical-Scavenging Antioxidants. Curr. Comp. Aid. Drug Des. 2005, 1, 257–273. [Google Scholar] [CrossRef]
- Rehakova, Z.; Koleckar, V.; Cervenka, F.; Jahodar, L.; Saso, L.; Opletal, L.; Jun, D.; Kuca, K. DPPH Radical Scavenging Activity of Several Naturally Occurring Coumarins and Their Synthesized Analogs Measured by the SIA Method. Toxicol. Mech. Methods 2008, 18, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Rajesh, P.M.; Natvar, P.J. In vitro antioxidantactivity of coumarin compounds by DPPH, Super oxide and nitric oxide free radical scavenging methods. J. Adv. Pharm. Edu. Res. 2011, 1, 52–68. [Google Scholar]
- Rodríguez, S.A.; Nazareno, M.A.; Baumgartner, M.T. Effect of different C3-aryl substituents on the antioxidantactivity of 4-hydroxycoumarin derivatives. Bioorg. Med. Chem. 2011, 19, 6233–6238. [Google Scholar] [CrossRef]
- Łuczaj, W.; Stankiewicz-Kranc, A.; Milewska, E.; Roszkowska-Jakimic, W.; Skrzydlewska, E. Effect of sweet grass extract against oxidative stress in rat liver and serum. Food Chem. Toxicol. 2012, 50, 135–140. [Google Scholar] [CrossRef]
- Martin-Aragon, S.; Benedi, J.M.; Villar, A.M. Effects of the Antioxidant (6,7-dihydroxycoumarin) Esculetin on the Glutathione System and Lipid Peroxidation in Mice. Gerontology 1998, 44, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Thuong, P.T.; Hung, T.M.; Ngoc, T.M.; Ha, D.T.; Min, B.S.; Kwack, S.J.; Kang, T.S.; Choi, J.S.; Bae, K.H. Antioxidant Activities of Coumarins from Korea Medicinal Plants and their Structure-Activity Relationships. Phytother. Res. 2010, 24, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, T.; Tahara, S.; Takabayashi, F. Inhibitory Effect of Natural Coumarin Compounds, Esculetin and Esculin, on Oxidative DNA Damage and Formation of Aberrant CryptFoci and TumorsInduced by 1,2-dimethylhydrazine in Rat Colons. Biol. Pharm. Bull. 2007, 30, 2052–2057. [Google Scholar] [CrossRef] [Green Version]
- Kleiner, H.E.; Vulimiri, S.V.; Starost, M.F.; Reed, M.J.; DiGiovanni, J. Oral administration of the citrus coumarin, isopimpinellin, blocks DNA adduct formation and skin tumor initiation by 7,12-dimethylbenz[a]anthracene in SENCAR mice. Carcinogenesis 2002, 23, 1667–1675. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.Q.; Yu, W.; Liu, Z.L. Antioxidative and prooxidativeeffects of coumarin derivatives on free initiated and photosensitized peroxidation of human low-denisty lipoproteid. Chem. Phys. Lipids 1999, 103, 125–135. [Google Scholar] [CrossRef]
- Singh, T.S.; Rao, B.S.M.; Mohan, H.; Mittal, J.P. A pulse radiolysis study of coumarin and its derivatives. J. Photochem. Photobiol. A Chem. 2002, 153, 163–171. [Google Scholar] [CrossRef]
- Fausta, N.; Lorrain, B.; Prasad, A.K.; Parmar, V.S.; Saso, L.; Scaccini, C. 4-methylcoumarins as antioxidants: Scavenging of peroxylradicals and inhibition of human low-density lipoprotein oxidation. Biochimie 2010, 92, 1147–1152. [Google Scholar] [CrossRef]
- Kumar, C.S.; El-Sheikh, M.A.; Alfarhan, A.F.; Kumar, R.S.; Idhayadhulla, A. Synthesis of novel coumarin analogues: Investigation of molecular docking interaction of SARS-CoV-2 proteins with natural and synthetic coumarin analogues and their pharmacokinetics studies. Saudi J. Biol. Sci. 2021, 28, 1100–1108. [Google Scholar] [CrossRef]
- Siriwach, R.; Kinoshita, H.; Kitani, S.; Igarashi, Y.; Pansuksan, K.; Panbangred, W.; Nihira, T. Mycoleptione, a new chromone derivative isolated from the endophytic fungus Mycoleptodiscu ssp. MU41. J. Antibiot. 2012, 65, 627–629. [Google Scholar] [CrossRef] [Green Version]
- Isaka, M.; Haritakun, R.; Supothina, S.; Choowong, W.; Mongkolsamrit, S. N-Hydroxypyridone alkaloids, chromone derivatives, and tetrahydroxanthones from the scale-insect pathogenic fungus Orbiocrella sp. BCC 33248. Tetrahedron 2014, 70, 9198–9203. [Google Scholar] [CrossRef]
- Khadem, S.; Marles, R.J. Chromone and flavonoid alkaloids: Occurrence and bioactivity. Molecules 2012, 17, 191–206. [Google Scholar] [CrossRef] [PubMed]
- Langenhoven, J.H.; Breytenbach, J.C.; Gerritsma-van der Vijver, L.M. An antihypertensive chromone from Ptaeroxyl onobliquum. Planta Med. 1988, 54, 373. [Google Scholar] [CrossRef]
- Reynolds, J.E.F. Martindale—The Extra Pharmacopoeia, 30th ed.; The Pharmaceutical Press: London, UK, 1993; p. 1024. [Google Scholar]
- Yadav, P.; Parshad, B.; Manchanda, P.; Sharma, S.K. Chromones and their derivatives as radical scavengers: A remedy for cell impairment. Curr. Top. Med. Chem. 2014, 14, 2552–2575. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Tian, J.M.; Tang, H.Y.; Pan, S.-Y.; Zhang, A.-L.; Gao, J.-M. Chaetosemins A–E, new chromones isolated from an Ascomycete Chaetomium seminudum and their biological activities. RSC Adv. 2015, 5, 29185–29192. [Google Scholar] [CrossRef]
- Carola, C.; Pfluecker, F.; Graf, R.; Epstein, H.; Wirth, C. Tiliroside and dihydroxy methylchromone: From nature to cosmetic applications. J. Appl. Cosmetol. 2010, 28, 109–123. [Google Scholar]
- Phosrithong, N.; Samee, W.; Nunthanavanit, P.; Ungwitayatorn, J. In vitro antioxidant activity study of novel chromone derivatives. Chem. Biol. Drug Des. 2012, 79, 981–989. [Google Scholar] [CrossRef]
- Mazzei, M.; Sottofattori, E.; Dondero, R.; Ibrahim, M.; Melloni, E.; Michetti, M. N,N-Dialkylaminosubstituted chromones and isoxazoles as potential anti-inflammatory agents. Farmaco 1999, 54, 452–460. [Google Scholar] [CrossRef]
- Motai, T.; Kitanaka, S. Sesquiterpenechromones from Ferulafukanensisand theirnitricoxideproduction inhibitory effects. J. Nat. Prod. 2005, 68, 1732–1735. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Xu, Z.; Chai, X.; Zeng, K.; Jia, Y.; Bi, D.; Ma, Z.; Tu, P. Nine 2-(2-phenylethyl)-chromonederivatives from the resinouswood of Aquilaria sinensisand their inhibition of LPS-induced NO production in RAW 264.7 cells. Eur. J. Org. Chem. 2012, 27, 5389–5397. [Google Scholar] [CrossRef]
- Liu, S.Z.; Feng, J.Q.; Wu, J.; Zhao, W.-M. A new monoterpene coumarin and a new monoterpene-chromone from Gerbera delavayi. Helv. Chim. Acta 2010, 93, 2026–2029. [Google Scholar] [CrossRef]
- Semwal, R.B.; Semwal, D.K.; Combrinck, S.; Viljoen, A. Health benefits of chromones: Common ingredients of our daily diet. Phytochem. Rev. 2020, 19, 761–785. [Google Scholar] [CrossRef]
- Kasprzak, M.M.; Erxleben, A.; Ochocki, J. Properties and applications of flavonoid metal complexes. RSC Adv. 2015, 5, 45853–45877. [Google Scholar] [CrossRef]
- Grazul, M.; Budzisz, E. Biological activity of metal ions complexes of chromones, coumarins and flavones. Coord. Chem. Rev. 2009, 253, 2588–2598. [Google Scholar] [CrossRef]
- Souza, R.F.V.; Sussuchi, E.M.; de Giovani, W.F. Synthesis, electrochemical, spectral and antioxidant properties of complexes of flavonoids with metal ions. Synth. React. Inorg. Met.-Org. Chem. 2003, 33, 1125–1144. [Google Scholar] [CrossRef]
- Afanas’ev, I.A.B.; Ostrakhovitch, E.A.; Mikhal’chik, E.V.; Ibragimova, G.A.; Korkina, G.L. Enhancement of antioxidant and anti inflammatory activities of bioflavonoid rutin by complexation with transition metals. Biochem. Pharmacol. 2001, 61, 677–684. [Google Scholar] [CrossRef]
- Moridani, M.Y.; Pourahmad, J.; Bui, H.; Siraki, A.; O’Brien, P.J. Dietary flavonoid iron complexes as cytoprotective superoxide radical scavengers. Free Radic. Biol. Med. 2003, 34, 243–253. [Google Scholar] [CrossRef]
- Souza, R.F.V.; de Giovani, W.F. Antioxidant properties of complexes of flavonoids with metal ions. Redox Rep. 2004, 9, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Jabeen, E.; Naveed, K.J.; Safeer, A.; Murtazab, I.; Ali, T.; Hameed, S. Radical scavenging propensity of Cu2+, Fe3+ complexes of flavonoids and in-vivo radical scavenging by Fe(III) primuletin. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2017, 171, 432–438. [Google Scholar] [CrossRef]
- Jabbari, M.; Gharib, F. Solvent dependence on antioxidant activity of some water-insoluble flavonoids and their cerium(IV) complexes. J. Mol. Liq. 2012, 168, 36–41. [Google Scholar] [CrossRef]
- Wang, Q.; Zhao, H.; Zhu, M.; Gao, L.; Cheng, N.; Cao, W. Spectroscopy characterization, theoretical study andantioxidant activities of the flavonoids-Pb(II) complexes. J. Mol. Struct. 2020, 1209. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Takamura, H.; Matoba, T.C.; Terao, J. HPLC method for evaluation of the free radical-scavenging activity of foods by using 1,1-diphenyl-2-picrylhydrazyl. Biosci. Biotechnol. Biochem. 1998, 62, 1201–1204. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.Y.; Majumder, K.; Wu, J. Oxygen radical absorbance capacity of peptides from egg white protein ovotransferrin and their interaction with phytochemicals. Food Chem. 2010, 23, 635–641. [Google Scholar] [CrossRef]
- Medina, J.M.; Naso, L.G.; Pérez, A.L.; Rizzi, A.; Ferrer, E.G.; Williams, P.A. Antioxidant and anticancer effects and bioavailability studies of the flavonoid baicalin and its oxidovanadium(IV) complex. J. Inorg. Biochem. 2017, 166, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Alper, P.; Erkisa, M.; Genckal, H.M.; Sahin, S.; Ulukaya, E.; Ari, F. Synthesis, characterization, anticancer and antioxidant activity of new nickel(II) and copper(II) flavonoid complexes. J. Mol. Struct. 2019, 1196, 783–792. [Google Scholar] [CrossRef]
- Pereira, R.M.S.; Andrades, N.E.D.; Paulino, N.; Sawaya, A.C.H.F.; Eberlin, M.N.; Marcucci, M.C.; Favero, G.M.; Novak, E.M.; Bydlowski, S.P. Synthesis and characterization of a metal complex containing naringin and Cu, and its antioxidant, antimicrobial, antiinflammatory and tumor cell cytotoxicity. Molecules 2007, 12, 1352–1366. [Google Scholar] [CrossRef] [Green Version]
- Etcheverry, S.B.; Ferrer, E.G.; Naso, L.; Rivadeneira, J.; Salinas, V.; Williams, P.A. Antioxidant effects of the VO (IV) hesperidin complex and its role in cancer chemoprevention. J. Biol. Inorg. Chem. 2008, 13, 435–447. [Google Scholar] [CrossRef]
- Porfírio, D.A.; de Queiroz Ferreira, R.; Malagutti, A.R.; Agostini Valle, E.M. Electrochemical study of the increased antioxidant capacity offlavonoids through complexation with iron(II) ions. Electrochim. Acta 2014, 141, 33–38. [Google Scholar] [CrossRef]
- Miyashiro, C.A.H.V.; Diniz, S.N.; de Oliveira, D.A.F.; Gonçalves, I.D.; Pereira, R.M.S.; Silva, R.G.; Paulino, N.; Okuyama, C.E. The potentiation of anti-inflammatory effect and INOS and COX-2 gene expression inhibition by rut in when complexed with cooper. Br. J. Med. Med. Res. 2014, 4, 4289–4309. [Google Scholar] [CrossRef]
- Panhwar, Q.K.; Memon, S. Synthesis, characterization and antioxidant activity of rutin complexes. Pak. J. Anal. Environ. Chem. 2014, 15, 60–70. [Google Scholar]
- Panhwar, Q.; Memon, S. Synthesis, characterisation, and antioxidant study of Cr(III)-rutin complex. Chem. Pap. 2014, 68, 614–623. [Google Scholar] [CrossRef]
- Kostyuk, V.A.; Potapovich, A.I.; Vladykovskaya, E.N.; Korkina, L.G.; Afanas’ev, I.B.A. Influence of metal ions on flavonoid protection against asbestos-induced cell injury. Arch. Biochem. Biophys. 2001, 385, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Blois, M.S. Antioxidant Determinations by the Use of a Stable Free Radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Williams, W.B.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Molyneux, P. The Use of Stable Free Radical Diphenylpicrylhydrazyl (DPPH) for Estimating Antioxidant Activity. Songklanakarin J. Sci. Technol. 2004, 26, 211–219. [Google Scholar]
- Bejaoui, L.; Rohlicek, J.; Ben Hassen, R. New cobalt (II) complexes of ‘3-acetyl-4-hydroxy-2H-chromene-2-one’: Crystal structure and Hirshfeld surface analysis, fluorescence behaviour and antioxidant activity. J. Mol. Struct. 2018, 1173, 574–582. [Google Scholar] [CrossRef]
- Yuan, Y.V.; Bone, D.E.; Carrington, M.F. Antioxidant activity of dulse (Palmaria palmata) extract evaluated in vitro. Food Chem. 2005, 91, 485–494. [Google Scholar] [CrossRef]
- Nabavi, S.M.; Ebrahimzadeh, M.A.; Nabavi, S.F.; Hamidinia, A.; Bekhradnia, A.R. Determination of antioxidant activity, phenol and flavonoid content of Parrotia persica mey. Pharmacol. Online 2008, 2, 560–567. [Google Scholar]
- Kenchappa, R.; Bodke, Y.D.; Chandrashekar, A.; Aruna, S.M.A.; Peethamba, S.K. Synthesis of coumarin derivatives containing pyrazole and indenone rings as potent antioxidant and antihyperglycemic agents. Arab. J. Chem. 2017, 10, S3895–S3906. [Google Scholar] [CrossRef] [Green Version]
- Ronconi, L.; Sadler, P.J. Using coordination chemistry to design new medicines. Coord. Chem. Rev. 2007, 251, 1633–1648. [Google Scholar] [CrossRef]
- De Alcantara, F.C.; Lozano, V.F.; Velosa, S.V.; Machado dos Santos, M.R.; Silva Pereira, R.M. New coumarin complexes of Zn, Cu, Ni and Fe with antiparasiticactivity. Polyhedron 2015, 101, 165–170. [Google Scholar] [CrossRef]
- Kavitha, P.; Laxma Reddy, K. Synthesis, Structural Characterization, and Biological Activity Studies of Ni(II) and Zn(II) Complexes. Bioinorg. Chem. Appl. 2014, 2014, 568741. [Google Scholar] [CrossRef] [Green Version]
- Okabe, N.; Yamamoto, E.; Yasunori, M. trans-Bis(3-hydroxy-2-phenyl-4H-1-benzopyran-4-onato)bis(pyridine)copper(II). Acta Crystallogr. Sect. E Struct. Rep. Online 2003, 59, m715–m716. [Google Scholar] [CrossRef]
- Dai, F.; Yan, W.-J.; Du, Y.-T.; Bao, X.-Z.; Li, X.-Z.; Zhou, B. Structural basis, chemical driving forces and biological implications of flavones as Cu(II) ionophores. Free Radic. Biol. Med. 2017, 108, 554–563. [Google Scholar] [CrossRef]
- El Amrani, F.B.A.; Perello, L.; Real, J.A.; Gonzalez-Alvarez, M.; Alzuet, G.; Borras, J.; Garcia-Granda, S.; Montejo-Bernardo, J. Oxidative DNA cleavage induced by an iron(III) flavonoid complex: Synthesis, crystal structure and characterization of chlorobis(flavonolato)(methanol) iron(III) complex. J. Inorg. Biochem. 2006, 100, 1208–1218. [Google Scholar] [CrossRef]
- Farina, Y.; Yamin, B.M.; Fun, H.-K.; Yip, B.-C.; Teoh, S.-G. trans-Bis(3-hydroxy-2-phenyl-4H-1-benzopyran-4-onato)bis(pyridine)nickel(II). Acta Crystallogr. Sect. C Cryst. Struct. Commun. 1995, 51, 1537–1540. [Google Scholar] [CrossRef]
- Kaizer, J.; Barath, G.; Pap, J.; Speier, G.; Giorgi, M.; Reglier, M. Manganese and iron flavonolates as flavonol 2,4-dioxygenase mimics. Chem. Commun. 2007, 5235–5237. [Google Scholar] [CrossRef]
- Kahrovic, E.; Zahirovic, A.; Višnjevac, A.; Osmanković, I.; Turkušić, E.; Kurtagić, H. Chalcone and Flavonol Copper(II) Complexes Containing Schiff Base Co-Ligand: Synthesis, Crystal Structures and Catecholase-like Activity. Croat. Chem. Acta 2018, 91, 195–207. [Google Scholar] [CrossRef]
- Halevas, E.; Pekou, A.; Papi, R.; Mavroidi, B.; Hatzidimitriou, A.G.; Zahariou, G.; Litsardakis, G.; Sagnou, M.; Pelecanou, M.; Pantazaki, A.A. Synthesis, physicochemical characterization and biological properties of two novel Cu(II) complexes based on natural products curcumin and quercetin. J. Inorg. Biochem. 2020, 208, 111083–111085. [Google Scholar] [CrossRef] [PubMed]
- Balogh-Hergovich, E.; Kaizer, J.; Speier, G.; Fulop, V.; Parkanyi, L. Quercetin 2,3-Dioxygenase Mimicking Ring Cleavage of the Flavonolate Ligand Assisted by Copper. Synthesis and Characterization of Copper(I) Complexes [Cu(PPh3)2(fla)] (fla = Flavonolate) and [Cu(PPh3)2(O-bs)] (O-bs = O-Benzoylsalicylate). Inorg. Chem. 1999, 38, 3787–3795. [Google Scholar] [CrossRef]
- Kaizer, J.; Pap, J.; Speier, G.; Parkanyi, L. The Reaction of μ-η2:η2-Peroxo- and Bis(μ-oxo)dicopper Complexes with Flavonol. Eur. J. Inorg. Chem. 2004, 10, 2253–2259. [Google Scholar] [CrossRef]
- Lynch, W.E.; Nivens, D.; Quillian, B.; Padgett, C.W.; Petrillo, A.; Peek, N.; Stone, J. A Copper(II) tris-imidazolylphosphine complex as a functional model of flavonol 2,4-dioxygenase. J. Mol. Struct. 2019, 1185, 99–109. [Google Scholar] [CrossRef]
- Grubel, K.; Rudzka, K.; Arif, A.M.; Klotz, K.L.; Halfen, J.A.; Berreau, L.M. Synthesis, Characterization, and Ligand Exchange Reactivity of a Series of First Row Divalent Metal 3-Hydroxyflavonolate Complexes. Inorg. Chem. 2010, 49, 82–96. [Google Scholar] [CrossRef] [PubMed]
- Balogh-Hergovich, E.; Kaizer, J.; Speier, G.; Huttner, G.; Zsolnai, L. Copper-mediated oxygenation of flavonolate in the presence of a tridentate N-ligand. Synthesis and crystal structures of [Cu(fla)(idpaH)]ClO4 and [Cu(idpaH)(O-bs)]ClO4, [fla=flavonolate, idpaH=3,3′-iminobis(N,N-dimethylpropylamine), O-bs=O-benzoylsalicylate]. Inorg. Chim. Acta 2000, 304, 72–77. [Google Scholar]
- Balogh-Hergovich, E.; Kaizer, J.; Speier, G.; Huttner, G.; Jacobi, A. Reparation and Oxygenation of (Flavonolato)copper Isoindoline Complexes with Relevance to Quercetin Dioxygenase. Inorg. Chem. 2000, 39, 4224–4229. [Google Scholar] [CrossRef] [PubMed]
- Lippai, I.; Speier, G.; Huttner, G.; Zsolnai, L. (2,2′-Bipyridine)(flavonolato)copper(II) Perchlorate, [Cu(bpy)(fla)]ClO4. Acta Crystallogr. Sect. C Cryst. Struct. Commun. 1997, 53, 1547–1549. [Google Scholar] [CrossRef]
- Lippai, I.; Speier, G.; Huttner, G.; Zsolnai, L. Crystal and molecular structure of a ketocarboxylatocopper(ii) intermediate in the oxygenation of a copper(i) flavonolate complex. Chem. Commun. 1997, 8, 741–742. [Google Scholar] [CrossRef]
- Zhang, Z.-T.; Shi, J. Self-assembly and crystal structure of a three-dimensional copper(II) complex. J. Coord. Chem. 2007, 60, 1485–1491. [Google Scholar] [CrossRef]
- Rybarczyk-Pirek, A.J.; Malecka, M.; Glinka, L.; Ochocki, J. Trans-Bis(3-aminoflavone-2N,O)bis(perchlorato-O)copper(II), a new potential antitumour agent. Acta Crystallogr. Sect. C Cryst. Struct. Commun. 2007, 63, 410–412. [Google Scholar] [CrossRef]
- Zurowska, B.; Erxleben, A.; Glinka, L.; Leczycka, M.; Zyner, E.; Ochocki, J. Synthesis, spectroscopy and magnetism of novel metal complexes of 3-aminoflavone (3-af). X-ray crystal structure of 3-af and [Cu(3-af)2(NO3)2]. Inorg. Chim. Acta 2009, 362, 739–744. [Google Scholar] [CrossRef]
- Zurowska, B.; Slepokura, K.; Kasprzak, M.; Fabijanska, M.; Szmigiero, L.; Ochocki, J. Synthesis, spectroscopy and magnetism of fluoridoborate transition metal complexes with aminoflavone ligand (3-af). X-ray crystal structure of [Cu(BF4)2(3-af)2] and [Zn(BF4)(3-af)2](BF4)·CH3C(O)OEt. Inorg. Chim. Acta 2013, 407, 19–24. [Google Scholar] [CrossRef]
- Huang, Q.-Q.; Sun, Y.-J.; Wu, H.-W.; Wang, Y.-L. A structural and functional model of copper(II)-flavonolate ES complex of flavonol 2,4-dioxygenase. Inorg. Chim. Acta 2017, 467, 7–10. [Google Scholar] [CrossRef]
- Li, X.; Li, J.; Dong, X.; Gao, X.; Zhang, D.; Liu, C. A novel 3-Hydroxychromone fluorescence sensor for intracellular Zn2+ and its application in the recognition of prostate cancer cells. Sens. Actuators B 2017, 245, 129–136. [Google Scholar] [CrossRef]
- Balogh-Hergovich, E.; Kaizer, J.; Speier, G.; Huttner, G.; Rutsch, P. [3,3′-Iminobis(N,N-dimethylpropylamine)](4′-methoxyflavonolato)zinc(II) perchlorate, [Zn(4′-MeOfla)(idpaH)]ClO4. Acta Crystallogr. Sect. C Cryst. Struct. Commun. 1999, 55, 557–558. [Google Scholar] [CrossRef]
- Kaizer, J.; Kupan, A.; Pap, J.; Speier, G.; Reglier, M.; Michel, G. Crystal structure of [3,3′-iminobis(N,N-dimethylpropylamine)] (flavonolato)zinc(II) Perchlorate, [Zn(fla)(idpaH)]ClO4. Z. Krist. N. Cryst. Struct. 2014, 215, 571–572. [Google Scholar] [CrossRef] [Green Version]
- Hoof, S.; Limberg, C. The Behavior of Trispyrazolylborato-Metal(II)-Flavonolate Complexes as Functional Models for Bacterial Quercetinase—Assessment of the Metal Impact. Inorg. Chem. 2019, 58, 12843–12853. [Google Scholar] [CrossRef]
- Anderson, S.N.; Noble, M.; Grubel, K.; Marshall, B.; Arif, A.M.; Berreau, L.M. Influence of supporting ligand microenvironment on the aqueous stability and visible light-induced CO-release reactivity of zinc flavonolato species. J. Coord. Chem. 2014, 67, 4061–4075. [Google Scholar] [CrossRef]
- Pieniazek, E.; Kalembkiewicz, J.; Dranka, M.; Woznicka, E. Syntheses, crystal structures and antioxidant study of Zn(II) complexes with morin-5′-sulfonic acid (MSA). J. Inorg. Biochem. 2014, 141, 180–187. [Google Scholar] [CrossRef]
- Sorenson, S.; Popova, M.; Arif, A.M.; Berreau, L.M. A bipyridine-ligated zinc(II) complex with bridging flavonolate ligation: Synthesis, characterization, and visible-light-induced CO release reactivity. Acta Crystallogr. Sect. C Cryst. Struct. Chem. 2017, 73, 703–709. [Google Scholar] [CrossRef] [Green Version]
- Annan, T.A.; Peppe, C.; Tuck, D.G. The direct electrochemical synthesis of d lo metal ion derivatives of some anionic bidentate oxygen donors. Can. J. Chem. 1990, 68, 423–430. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.-J.; Huang, Q.-Q.; Tano, T.; Itoh, S. Flavonolate Complexes of MII (M = Mn, Fe, Co, Ni, Cu, and Zn). Structural and Functional Models for the ES (Enzyme–Substrate) Complex of Quercetin 2,3-Dioxygenase. Inorg. Chem. 2013, 52, 10936–10948. [Google Scholar] [CrossRef] [PubMed]
- Baráth, G.; Rácz, G.; Speier, G.; Pap, J.S.; Kaizer, J.; Giorgi, M. Crystal structure of [3-(N-methyl-2-pyridyl-N-hydroxymethyl-2-pyridyl)-aminopropionic acid-)k4N,N′,N″,O](flavonolato-) k2O,O′)cobalt(III) chloride—Water (1:2), [Co(C15H17N3O3)(C15H9O3)]Cl·2H2O. Z. Krist. N. Cryst. Struct. 2011, 226, 414–416. [Google Scholar] [CrossRef]
- Sun, Y.-J.; Huang, Q.-Q.; Zhang, J.-J. Series of Structural and Functional Models for the ES (Enzyme–Substrate) Complex of the Co(II)-Containing Quercetin 2,3-Dioxygenase. Inorg. Chem. 2014, 53, 2932–2942. [Google Scholar] [CrossRef] [PubMed]
- Hiller, W.; Nishinaga, A.; Rieker, A. A Simple Model for the Enzyme-Substrate-Complex of the Quercetinase Reaction. Crystal Structure of Flavonolatocobalt(III)(salen). Z. Naturforsch. B Chem. Sci. 1992, 47, 1185–1188. [Google Scholar] [CrossRef]
- Barath, G.; Kaizer, J.; Speier, G.; Parkanyi, L.; Kuzmann, E.; Vertes, A. One metal–two pathways to the carboxylate-enhanced, iron-containing quercetinase mimics. Chem. Commun. 2009, 3630–3632. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.-J.; Huang, Q.-Q.; Zhang, J.-J. A series of NiII-flavonolate complexes as structural and functional ES (enzyme-substrate) models of the NiII-containing quercetin 2,3-dioxygenase. Dalton Trans. 2014, 43, 6480–6489. [Google Scholar] [CrossRef]
- Yun, H. Tetraaqua(7-hydroxy-5-oxidoflavone-6-sulfonato-kappa2O4,O5)nickel(II) dimethylformamide solvate monohydrate. Acta Crystallogr. Sect. C Cryst. Struct. Commun. 2006, 62, m469–m471. [Google Scholar]
- Hoof, S.; Limberg, C. Bioinspired Trispyrazolylborato Nickel(II) Flavonolate Complexes and Their Reactivity Toward Dioxygen. Z. Anorg. Allg. Chem. 2019, 645, 170–174. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, B.; Zhu, Q.; Wu, Z.; Lin, Y.; Xing, Y.; Hua, R. Gaodeng Xuexiao Huaxue Xuebao. Chem. J. Chin. Univ. 1998, 19, 410. (In Chinese) [Google Scholar]
- Kalaiarasi, G.; Rex, S.; Rajkumar, J.; Dharani, S.; Rath, N.P.; Prabhakaran, R. In vitro cytotoxicity of new water soluble copper (II) metallates containing 7-hydroxy-4-oxo-4H-chromene thiosemicarbazones. Polyhedron 2019, 173, 114120. [Google Scholar] [CrossRef]
- Myannik, K.A.; Yarovenko, V.N.; Beloglazkina, E.K.; Moiseeva, A.A.; Krayushkin, M.M. Novel copper(II), cobalt(II) and nickel(II) complexes with 5-(4-oxo-4H-chromen-3-yl)-4,5-dihydro-1,3,4-thiadiazole-2-carboxamide: Synthesis, structure, spectroscopic studies. Polyhedron 2018, 139, 208–214. [Google Scholar] [CrossRef]
- Balakrishnan, N.; Haribabu, J.; Dhanabalan, A.K.; Swaminathan, S.; Sun, S.; Dibwe, D.F.; Bhuvanesh, N.; Awale, S.; Karvembu, R. Thiosemicarbazone(s)-anchored water soluble mono- and bimetallic Cu(ii) complexes: Enzyme-like activities, biomolecular interactions, anticancer property and real-time live cytotoxicity. Dalton Trans. 2020, 49, 9411–9424. [Google Scholar] [CrossRef]
- Mucha, P.; Malecka, M.; Kupcewicz, B.; Lux, K.; Dolega, A.; Jezierska, J.; Budzisz, E. Copper(II) complexes of 7-amino-2-methylchromone and 7-aminoflavone: Magneto-structural, spectroscopic and DFT characterization. Polyhedron 2018, 153, 181–196. [Google Scholar] [CrossRef]
- Ilies, D.-C.; Pahontu, E.; Shova, S.; Georgescu, R.; Stanica, N.; Olar, R.; Gulea, A.; Rosu, T. Synthesis, characterization, crystal structure and antimicrobial activity of copper(II) complexes with a thiosemicarbazone derived from 3-formyl-6-methylchromone. Polyhedron 2014, 81, 123–131. [Google Scholar] [CrossRef]
- Rahman, F.-U.; Yu, S.-B.; Khalil, S.K.; Wu, Y.-P.; Koppireddi, S.; Li, Z.-T.; Wang, H.; Zhang, D.-W. Chromone and benzyldithiocarbazate based probe: A highly selective and sensitive platform for colorimetric sensing of Cu2+, single crystal of the complex and DFT calculations. Sens. Actuators B 2018, 263, 594–603. [Google Scholar] [CrossRef]
- Kaizer, J.; Pap, J.; Speier, G.; Reglier, M.; Giorgi, M. Synthesis, properties, and crystal structure of a novel 3-hydroxy-(4H)-benzopyran-4-one containing copper(II) complex, and its oxygenation and relevance to quercetinase. Transit. Met. Chem. 2004, 29, 630–633. [Google Scholar] [CrossRef]
- Grazul, M.; Kufelnicki, A.; Wozniczka, M.; Lorenz, I.-P.; Mayer, P.; Jozwiak, A.; Czyz, M.; Budzisz, E. Synthesis, structure, electrochemical properties, cytotoxic effects and antioxidant activity of 5-amino-8-methyl-4H-benzopyran-4-one and its copper(II) complexes. Polyhedron 2012, 31, 150–158. [Google Scholar] [CrossRef]
- Kalaiarasi, G.; Rex, S.; Rajkumar, J.; Dharani, S.; Lynch, V.M.; Prabhakaran, R. Synthesis, spectral characterization and biological evaluation of some copper (II) complexes containing 4-oxo-4H-chromene-3-carbaldehyde-4 (N)-substituted thiosemicarbazones. Inorg. Chim. Acta 2018, 471, 759–776. [Google Scholar] [CrossRef]
- Li, Y.; Yang, Z.; Liao, Z.; Han, Z.; Liu, Z.-C. Synthesis, crystal structure, DNA binding properties and antioxidant activities of transition metal complexes with 3-carbaldehyde-chromone semicarbazone. Inorg. Chem. Commun. 2010, 13, 1213–1216. [Google Scholar] [CrossRef]
- Li, Y.; Yang, Z.W.; Wu, J.-C. Synthesis, crystal structures, biological activities and fluorescence studies of transition metal complexes with 3-carbaldehyde chromone thiosemicarbazone. Eur. J. Med. Chem. 2010, 45, 5692–5701. [Google Scholar] [CrossRef] [PubMed]
- Selvamurugan, S.; Ramachandran, R.; Vijayan, P.; Manikandan, R.; Prakash, G.; Viswanathamurthi, P.; Velmurugan, K.; Nandhakumar, R.; Endo, A. Synthesis, crystal structure and biological evaluation of Ni(II) complexes containing 4-chromone-N(4)-substituted thiosemicarbazone ligands. Polyhedron 2016, 107, 57–67. [Google Scholar] [CrossRef]
- Bullock, S.J.; Felton, C.E.; Fennessy, R.V.; Harding, L.P.; Andrews, M.; Pope, S.J.A.; Rice, C.R.; Riis-Johannessen, T. Coumarin-based luminescent ligand that forms helicates with dicationic metal ions. Dalton Trans. 2009, 47, 10570–10573. [Google Scholar] [CrossRef]
- Kataev, E.; Arnold, R.; Ruffer, T.; Lang, H. Fluorescence Detection of Adenosine Triphosphate in an Aqueous Solution Using a Combination of Copper(II) Complexes. Inorg. Chem. 2012, 51, 7948–7950. [Google Scholar] [CrossRef]
- Cui, Y.; Gao, Q.; Wang, H.; Wang, L.; Xie, Y.-B. Diaqua-bis-(2-oxo-2H-chromene-3-carboxyl-ato)copper(II). Acta Crystallogr. Sect. E Struct. Rep. Online 2011, 67, m782. [Google Scholar] [CrossRef]
- Huang, L.; Cheng, J.; Xie, K.; Xi, P.; Hou, F.; Li, Z.; Xie, G.; Shi, Y.; Liu, H.; Bai, D.; et al. Cu2+-selective fluorescent chemosensor based on coumarin and its application in bioimaging. Dalton Trans. 2011, 40, 10815–10817. [Google Scholar] [CrossRef] [PubMed]
- Bikas, R.; Farzaneh-Bonab, H.; Noshiranzadeh, N.; Aygun, M.; Emami, M.; Lis, T. Coumarin-naphthohydrazone ligand with a rare coordination mode to form Mn(II) and Co(II) 1-D coordination polymers: Synthesis, characterization, and crystal structure. J. Coord. Chem. 2018, 71, 1127–1146. [Google Scholar] [CrossRef]
- Kumar, V.; Kumar, A.; Diwan, U.; Singh, M.K.; Upadhyay, K.K. Turn “Off–On” Fluorescent Recognition of Cu2+ and Cys in Aqueous Medium: Implementation of Molecular Logic Gate and Cell Imaging Studies. Bull. Chem. Soc. Jpn. 2016, 89, 754–761. [Google Scholar] [CrossRef]
- Qian, B.; Varadi, L.; Trinchi, A.; Reichman, S.; Bao, L.; Lan, M.; Wei, G.; Cole, I.S. The Design and Synthesis of Fluorescent Coumarin Derivatives and Their Study for Cu2+ Sensing with an Application for Aqueous Soil Extracts. Molecules 2019, 24, 3569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandrasekhar, V.; Bag, P.; Pandey, M.D. Phosphorus-Supported Multidentate Coumarin-Containing Fluorescence, Sensors for Cu2+. Tetrahedron 2019, 65, 9876–9883. [Google Scholar] [CrossRef]
- Kumar, V.; Kumar, A.; Diwan, U.; Upadhyay, K.K. A Zn2+-responsive highly sensitive Fluorescent probe and 1D coordination polymer based on a coumarin platform. Dalton Trans. 2013, 42, 13078–13083. [Google Scholar] [CrossRef]
- Verma, S.; Savani, C. Vinay Singh, Synthesis, Photophysical, Thermal and Crystallographic Studies of 3-Aminocoumarin Based Monobasic k3-O,N,Otridentate/k2-N,O-bidentate Schiff Base Divalent Complexes. Chem. Sel. 2019, 4, 14244–14252. [Google Scholar]
- Aslkhademi, S.; Noshiranzadeh, N.; Sadjadi, M.S.; Mehrani, K.; Farhadyar, N. Synthesis, crystal structure and investigation of the catalytic and spectroscopic properties of a Zn (II) complex with coumarin-hydrazone ligand. Polyhedron 2019, 160, 115–120. [Google Scholar] [CrossRef]
- Wu, J.; Sheng, R.; Liu, W.; Wang, P.; Zhang, H.; Ma, J. Fluorescent sensors based on controllable conformational change for discrimination of Zn2+ over Cd2+. Tetrahedron 2012, 68, 5458–5463. [Google Scholar] [CrossRef]
- Areas, E.S.; Bronsato, B.J.; Pereira, T.M.; Guedes, G.P.; da S.Miranda, F.; Kummerle, A.E.; da Cruz, A.G.B.; Neves, A.P. Novel CoIII complexes containing fluorescent coumarin-N-acylhydrazone hybrid ligands: Synthesis, crystal structures, solution studies and DFT calculations. Spectrochim. Acta Part A 2017, 187, 130–142. [Google Scholar] [CrossRef]
- Mishra, R.K.; Upadhyay, K.K. Coumarin-Based Chromogenic Receptor for Ni2+ in Aqueous Medium Exhibiting a Reconfigurable Logic Gate Pattern. Eur. J. Org. Chem. 2011, 25, 4799–4885. [Google Scholar] [CrossRef]
- Jagtap, U.B.; Bapat, V.A. Artocarpus: A review of its traditional uses, phytochemistry and pharmacology. J. Ethnopharmacol. 2010, 129, 142–166. [Google Scholar] [CrossRef]
- Yue, W.; Ming, Q.; Lin, B.; Rahman, K.; Zheng, C.; Han, T.; Qin, L. Medicinal plant cell suspension cultures: Pharmaceutical applications and high-yielding trategies for the desired secondary metabolites. Crit. Rev. Biotechnol. 2016, 36, 215–232. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, S.; Krishnaswamy, S.; Devashya, V.; Sethuraman, S.; Krishnan, U.M. Flavonoid–Metal Ion Complexes: A Novel Class of Therapeutic Agents. Med. Res. Rev. 2014, 34, 677–702. [Google Scholar] [CrossRef] [PubMed]
- Liskova, A.; Samec, M.; Koklesova, L.; Samuel, S.M.; Zhai, K.; Al-Ishaq, R.K.; Abotaleb, M.; Nosal, V.; Kajo, K.; Ashrafizadeh, M.; et al. Flavonoids against the SARS-CoV-2 induced inflammatory storm. Biomed. Pharmacother. 2021, 138, 111430. [Google Scholar] [CrossRef]
- Mondal, S.; Rahaman, S.T. Flavonoids: A vital resource in healthcare and medicine. Pharm. Pharm. Int. J. 2020, 8, 91–104. [Google Scholar]
- Goris, T.; Perez-Valero, A.; Martınez, I.; Dong, Y.; Fernandez-Calleja, L.; San Leon, D.; Bornscheuer, U.T.; Magadan-Corpas, P.; Lombo, F.; Nogales, J. Repositioning microbial biotechnology against COVID-19: The case of microbial production of flavonoids. Microb. Biotechnol. 2021, 14, 94–110. [Google Scholar] [CrossRef]
- Symonowicz, M.; Kolanek, M. Flavonoids and their properties to form chelate complexes. Biotechnol Food Sci. 2012, 76, 35–41. [Google Scholar]
- Florkowska, K.; Duchnik, W.; Muzykiewicz, A.; Zielonka-Brzezicka, J.; Klimowicz, A. Flawonoidy w profilaktyce i leczeniu miażdżycy. Probl. Hig. Epidemiol. 2017, 98, 217–225. [Google Scholar]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Kubina, R.; Kabała-Dzik, A.; Wojtyczka, R.D.; Szaflarska-Stojko, E. Przeciwbakteryjne działanie galanginy zawartej w propolisie w stosunku do bakterii Gram-dodatnich. Farm. Przegl. Nauk. 2009, 8, 24–26. [Google Scholar]
- Prithviraj, K. Biological activities of flavonoids: An overview. Int. J. Pharm. Sci. Res. 2019, 10, 1567–1574. [Google Scholar]
- Zandi, K.; Teoh, B.-T.; Sam, S.-S.; Wong, P.-F.; Mohd Rais, M.; AbuBakar, S. Antiviral activity of four types of bioflavonoid against dengue virus type-2. Virol. J. 2011, 8, 560. [Google Scholar] [CrossRef] [Green Version]
- Ngwa, W.; Kumar, R.; Thompson, D.; Lyerly, W.; Moore, R.; Reid, T.-E.; Lowe, H.; Toyang, N. Potential of Flavonoid-Inspired Phytomedicines against COVID-19. Molecules 2020, 25, 2707. [Google Scholar] [CrossRef]
- Muchtaridi, M.; Fauzi, M.; Khairul Ikram, N.K.; Gazzali, A.M.; Wahab, H.A. Natural Flavonoids as Potential Angiotensin-Converting Enzyme 2 Inhibitors for Anti-SARS-CoV-2. Molecules 2020, 25, 3980. [Google Scholar] [CrossRef] [PubMed]
- Pandey, P.; Khan, F.; Kumar Rana, A.; Srivastava, Y.; Kumar, S.J.; Kumar, N.J. A Drug Repurposing Approach Towards Elucidating the Potential of Flavonoids as COVID-19 Spike Protein Inhibitors. Biointerface Res. Appl. Chem. 2021, 11, 8482–8501. [Google Scholar]
- Valizadeh, H.; Abdolmohammadi-vahid, S.; Danshina, S.; Ziya Gencr, M.; Ammari, A.; Sadeghia, A.; Roshangar, L.; Aslani, S.; Esmaeilzadeh, A.; Ghaebi, M.; et al. Nano-curcumin therapy, a promising method in modulating inflammatory cytokines in COVID-19 patients. Int. Immunopharmacol. 2020, 89, 107088. [Google Scholar] [CrossRef]
- Conti, P.; Ronconi, G.; Caraffa, A.; Gallenga, C.E.; Ross, R.; Frydas, I.; Kritas, S.K. Induction of pro-inflamatory cytokines (IL-1 i IL-6) and lung inflammation by coronavirus-19 (COVI-19 lub SARS-CoV-2): Anti-infalamatory strategies. J. Biol. Regul. Homeost. Agents 2020, 34, 327–331. [Google Scholar] [PubMed]
- Fei, J.; Liang, B.; Jiang, C.; Ni, H.; Liming, W. Luteolin inhibits IL-1β-induced inflammation in rat chondrocytes and attenuates osteoarthritis progression in a rat model. Biomed. Pharmacother. 2019, 109, 1586–1592. [Google Scholar] [CrossRef] [PubMed]
- Hadrich, F.; Sayadi, S. Apigetrin inhibits adipogenesis in 3T3-L1 cells by downregulating PPARγ and CEBP-α. Lipids Health Dis. 2018, 17, 3–8. [Google Scholar] [CrossRef] [Green Version]
- Salaverry, L.S.; Parrado, A.C.; Mangone, F.M.; Dobrecky, C.B.; Flor, S.A.; Lombardo, T.; Sotelo, A.D.; Saccodossi, N.; Rugna, A.Z.; Blanco, G.; et al. In vitro anti-inflammatory properties of Smilax campestris aqueous extract in human macrophages, and characterization of its flavonoid profile. J. Ethnopharmacol. 2020, 247, 112282. [Google Scholar] [CrossRef]




















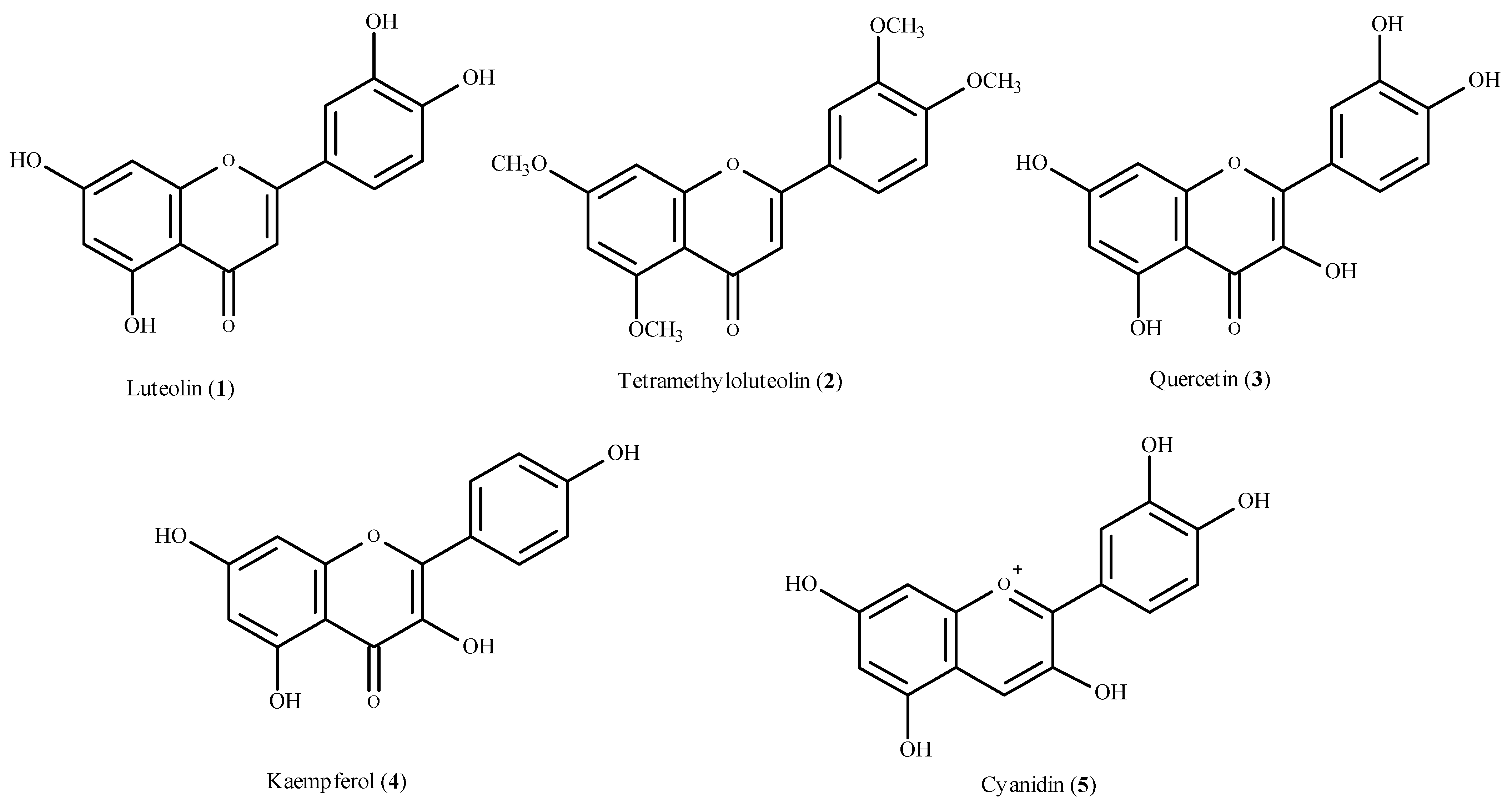{kind=link}
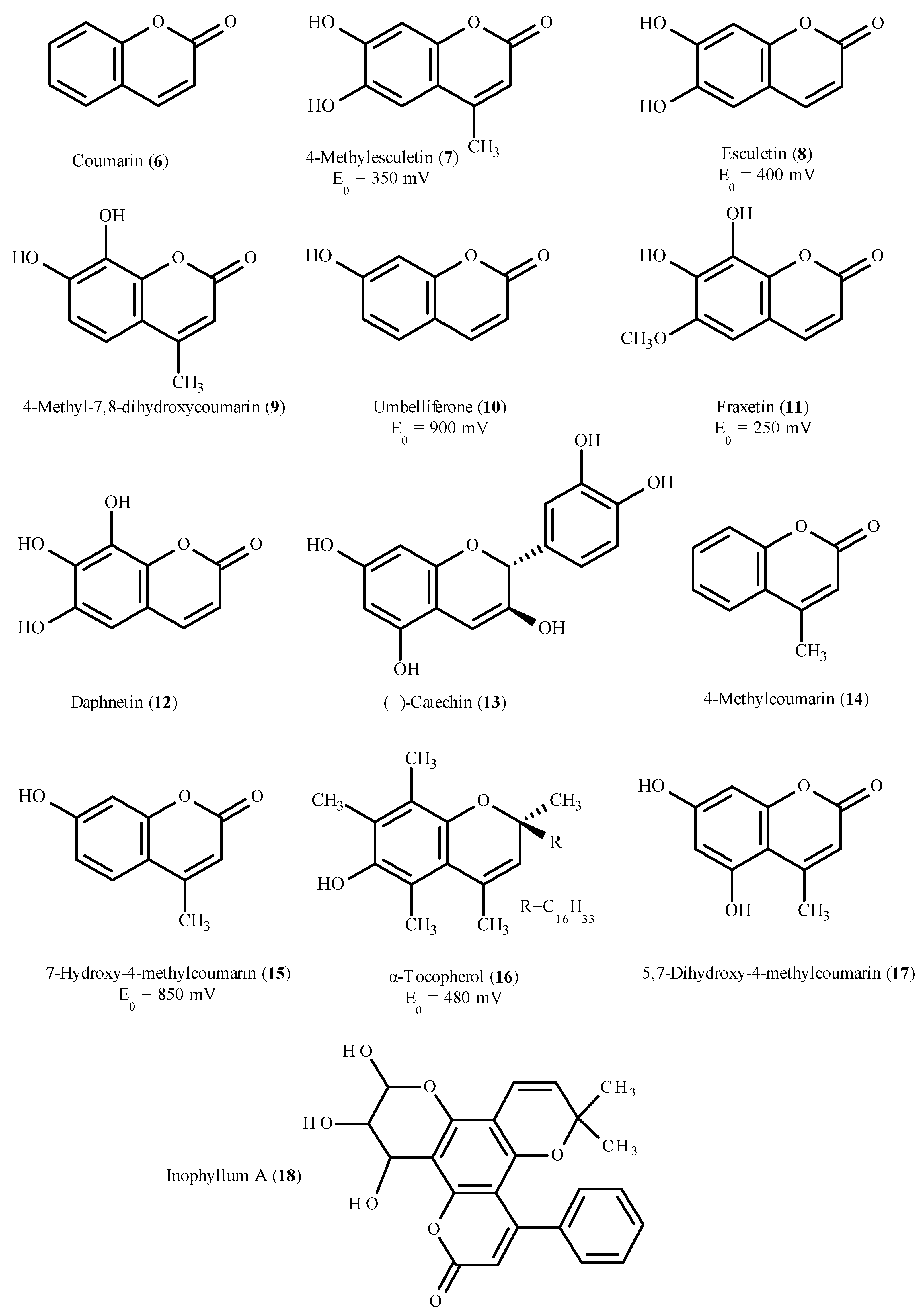{kind=link}
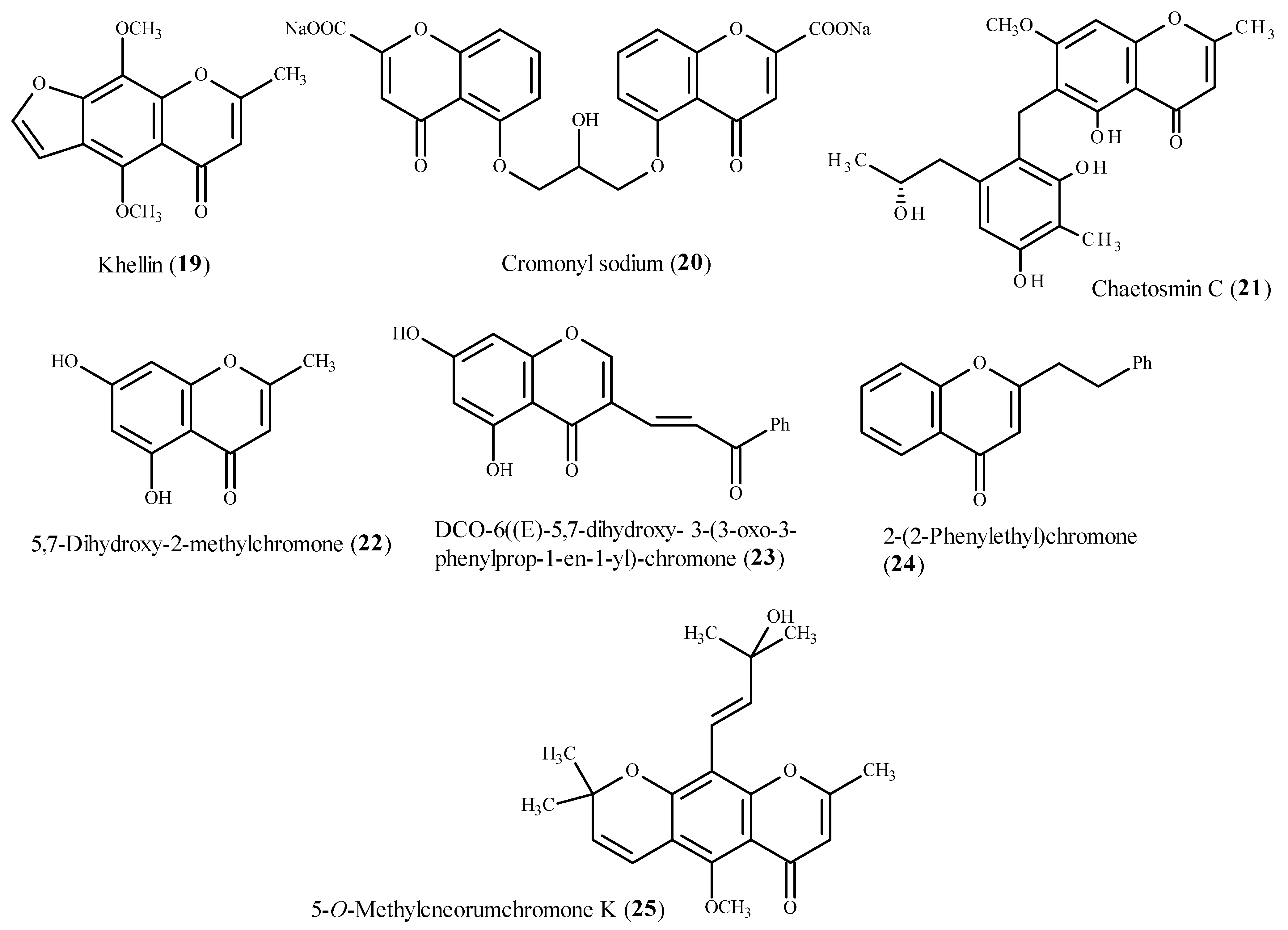{kind=link}
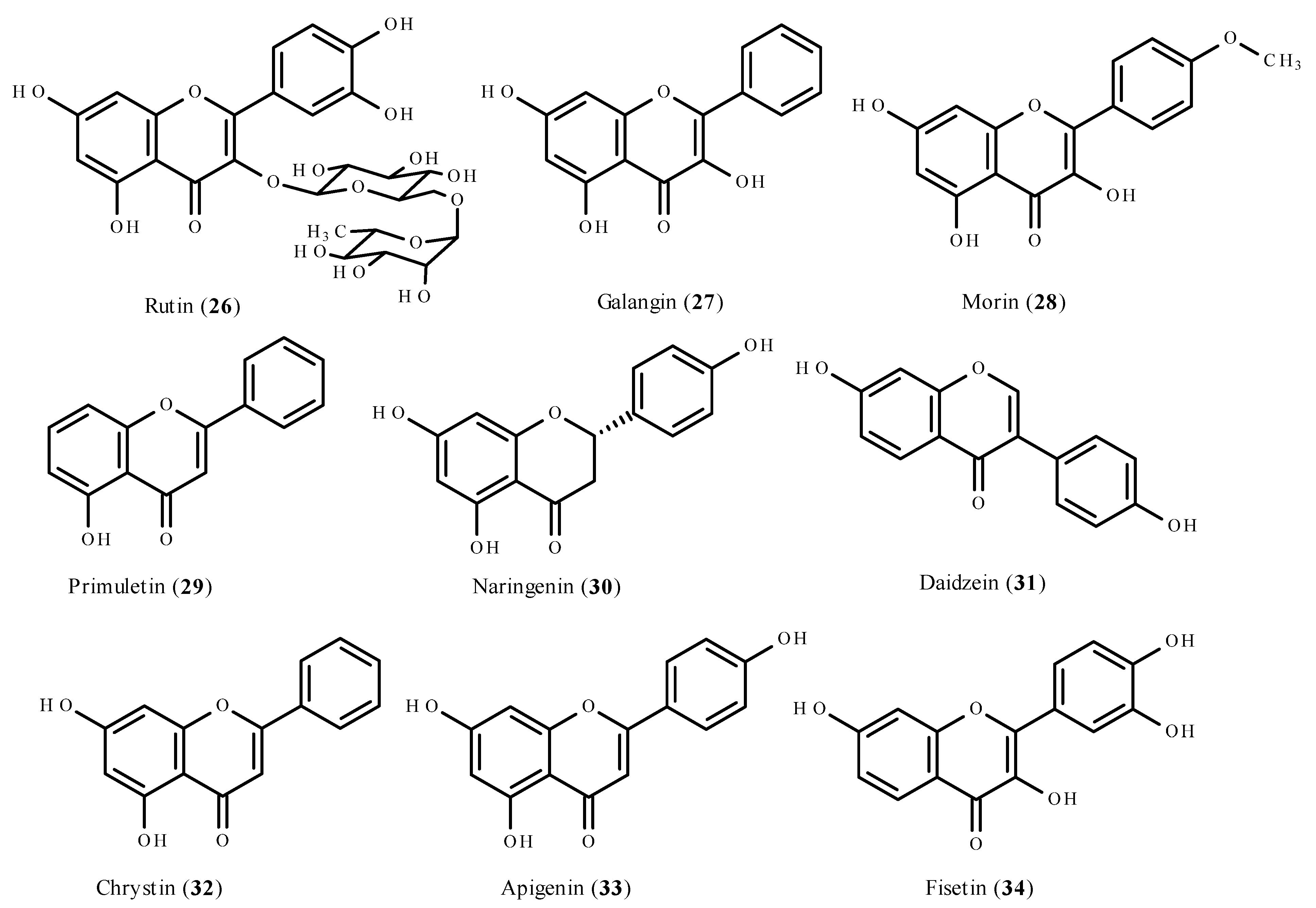{kind=link}
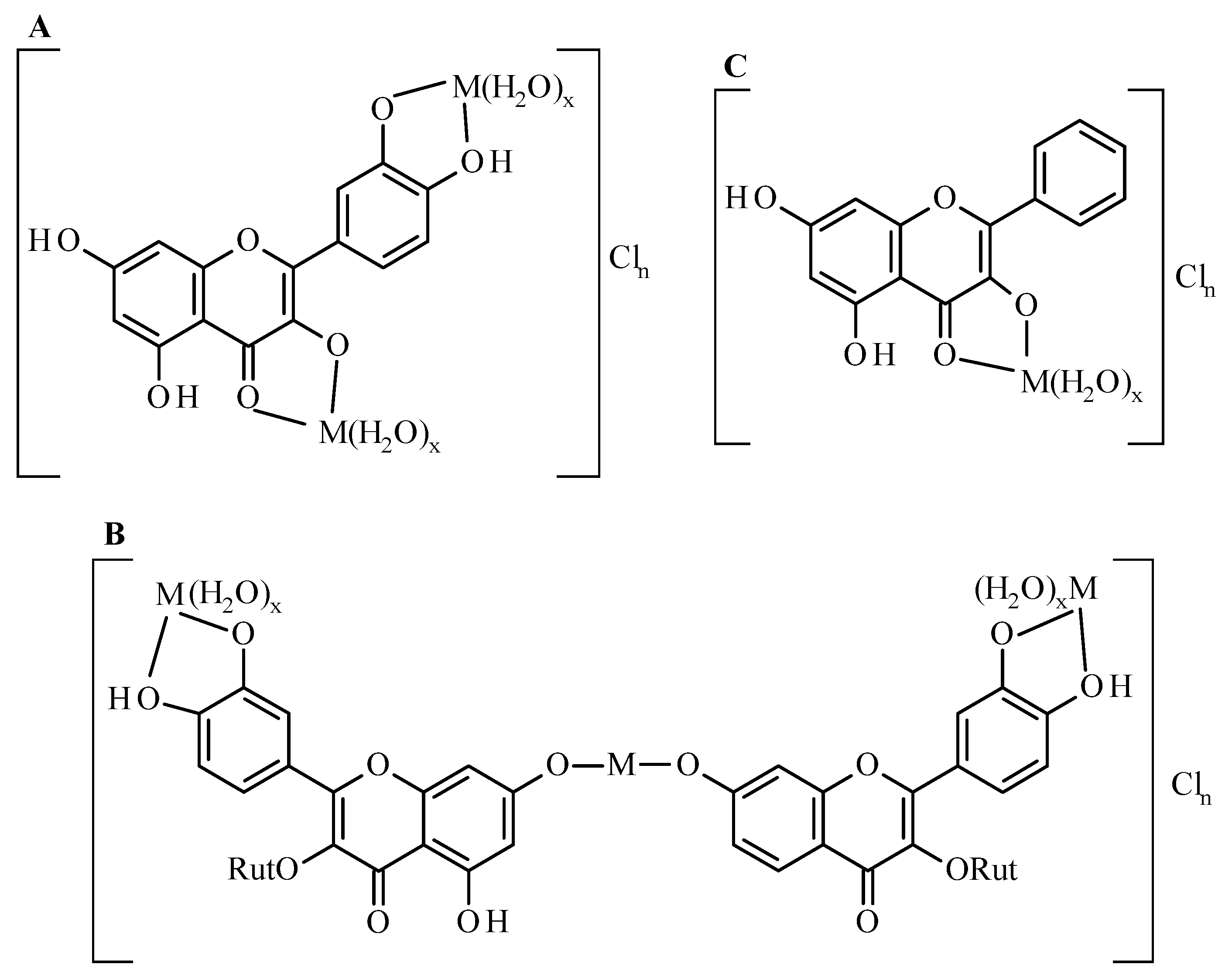{kind=link}
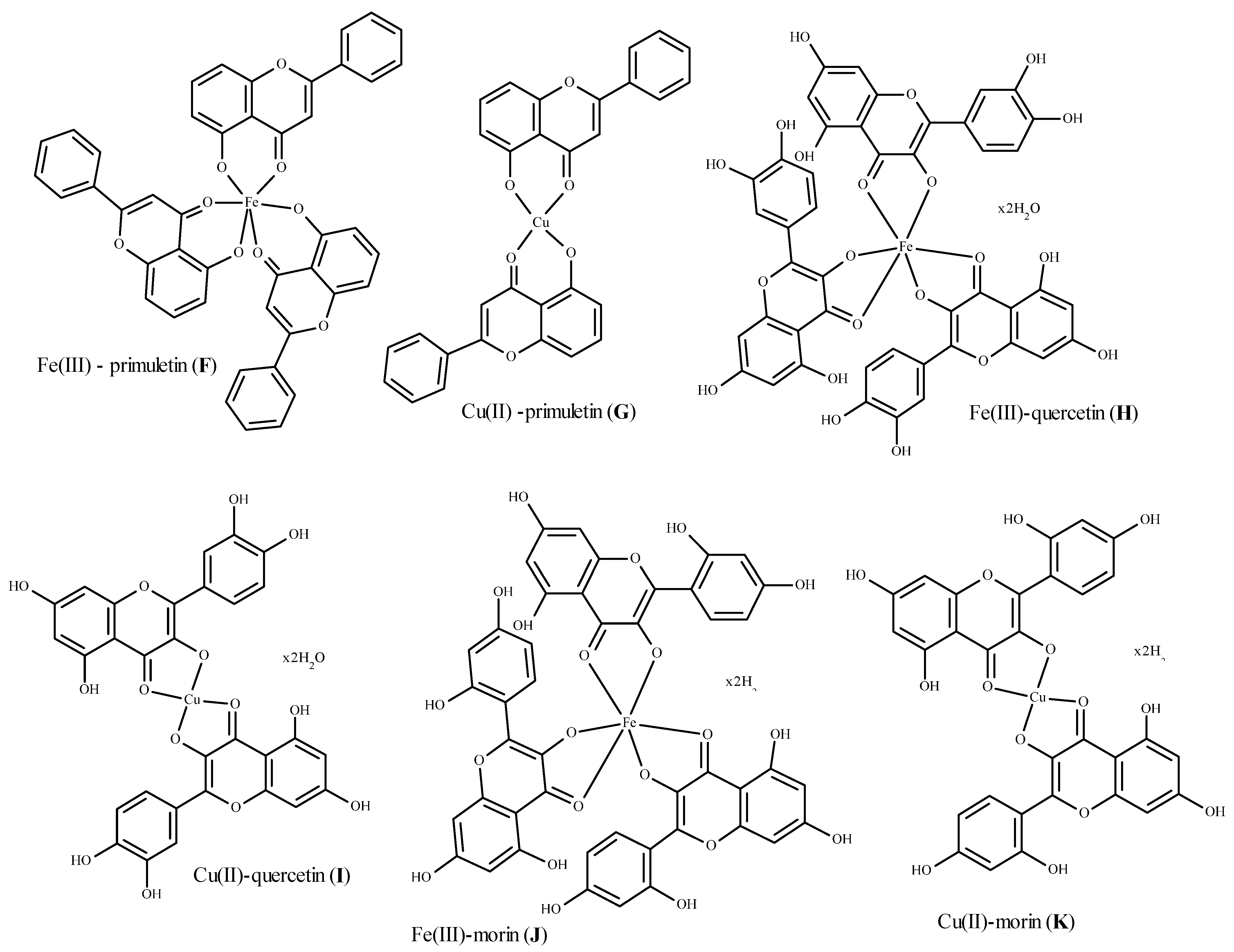{kind=link}
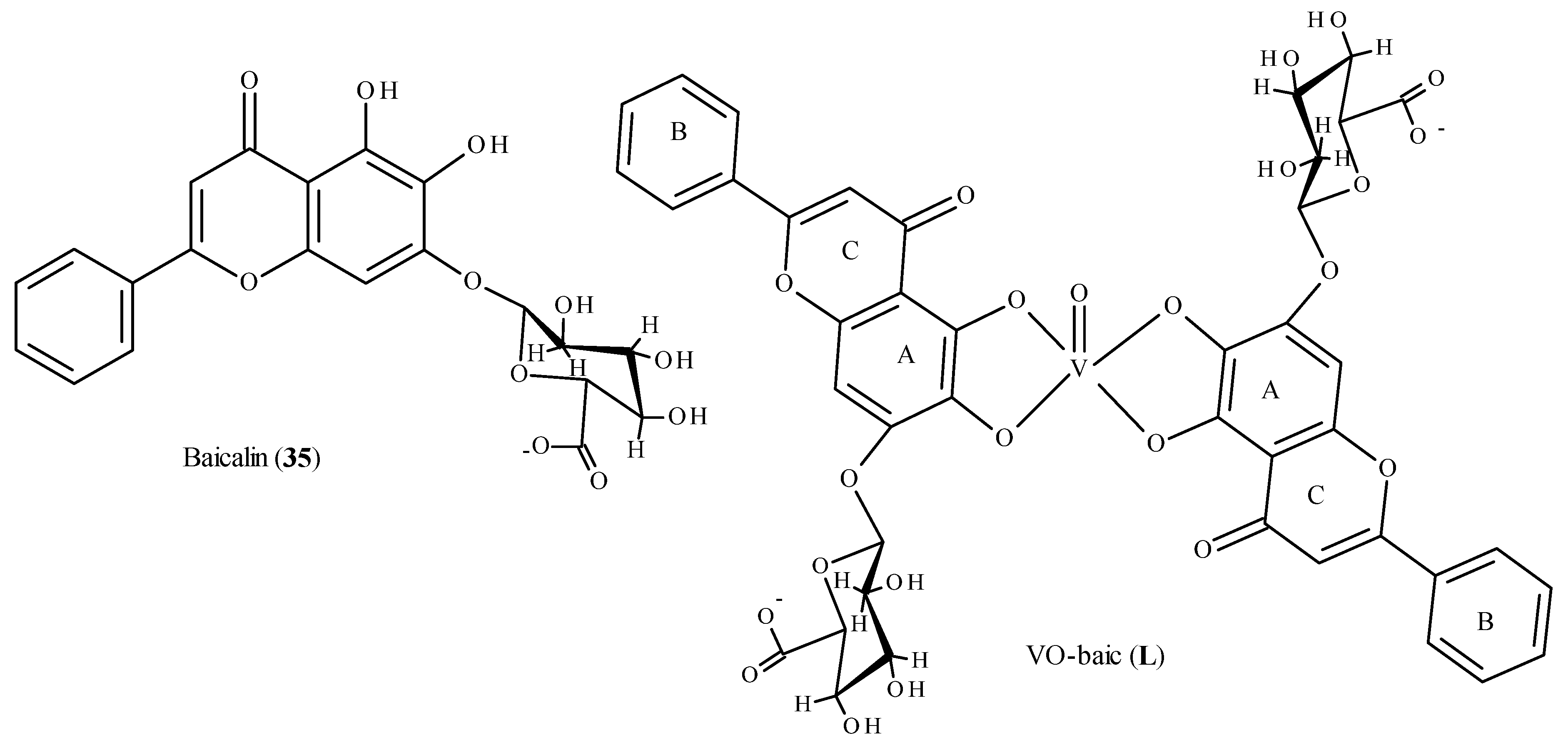{kind=link}
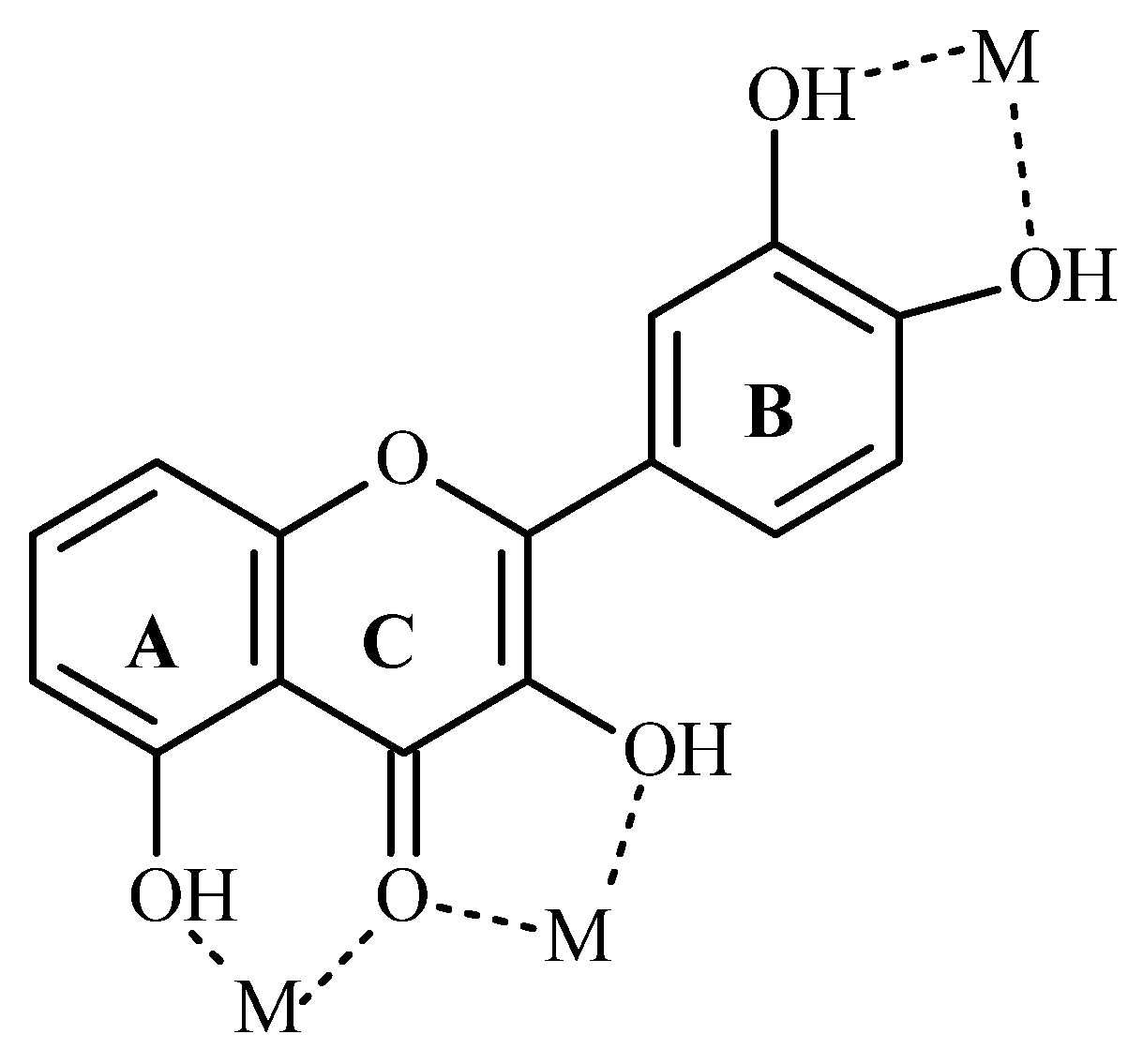{kind=link}
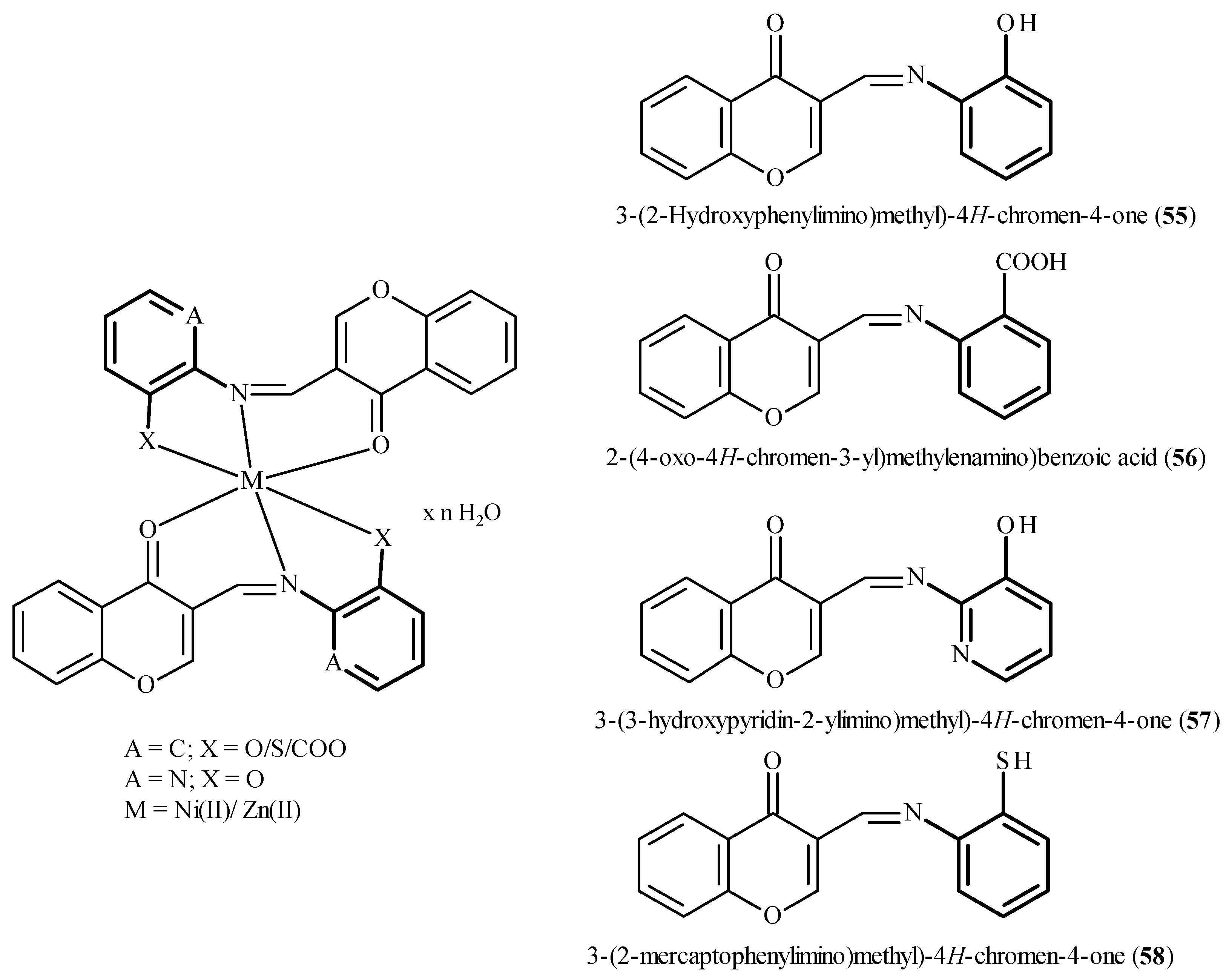{kind=link}
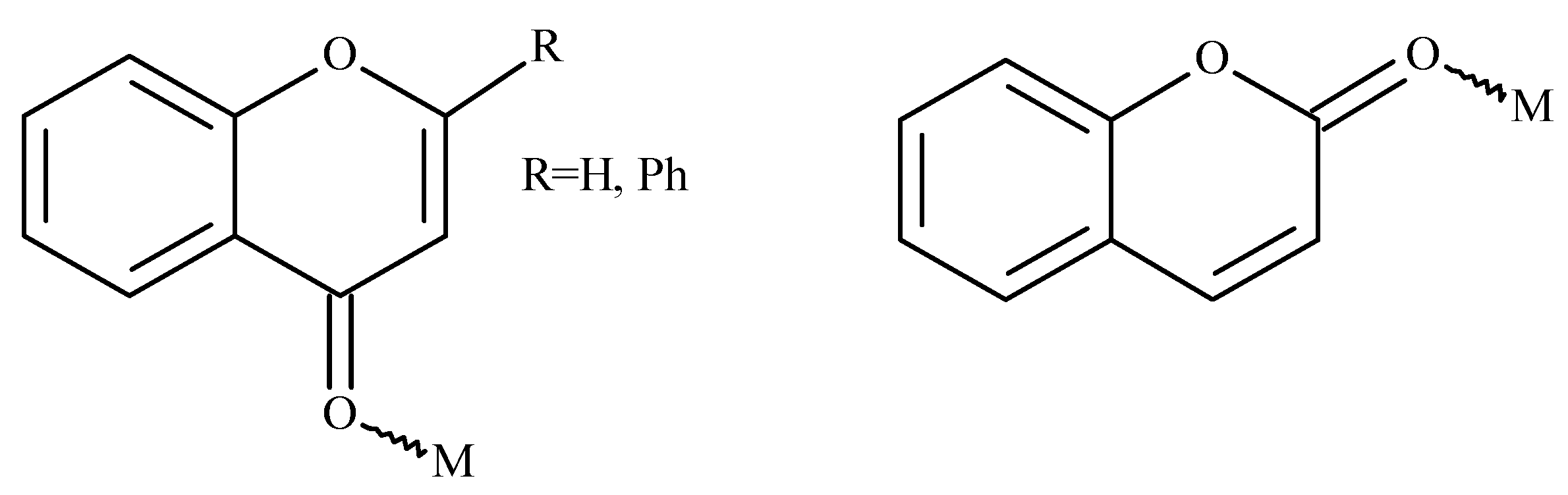{kind=link}
{kind=link}
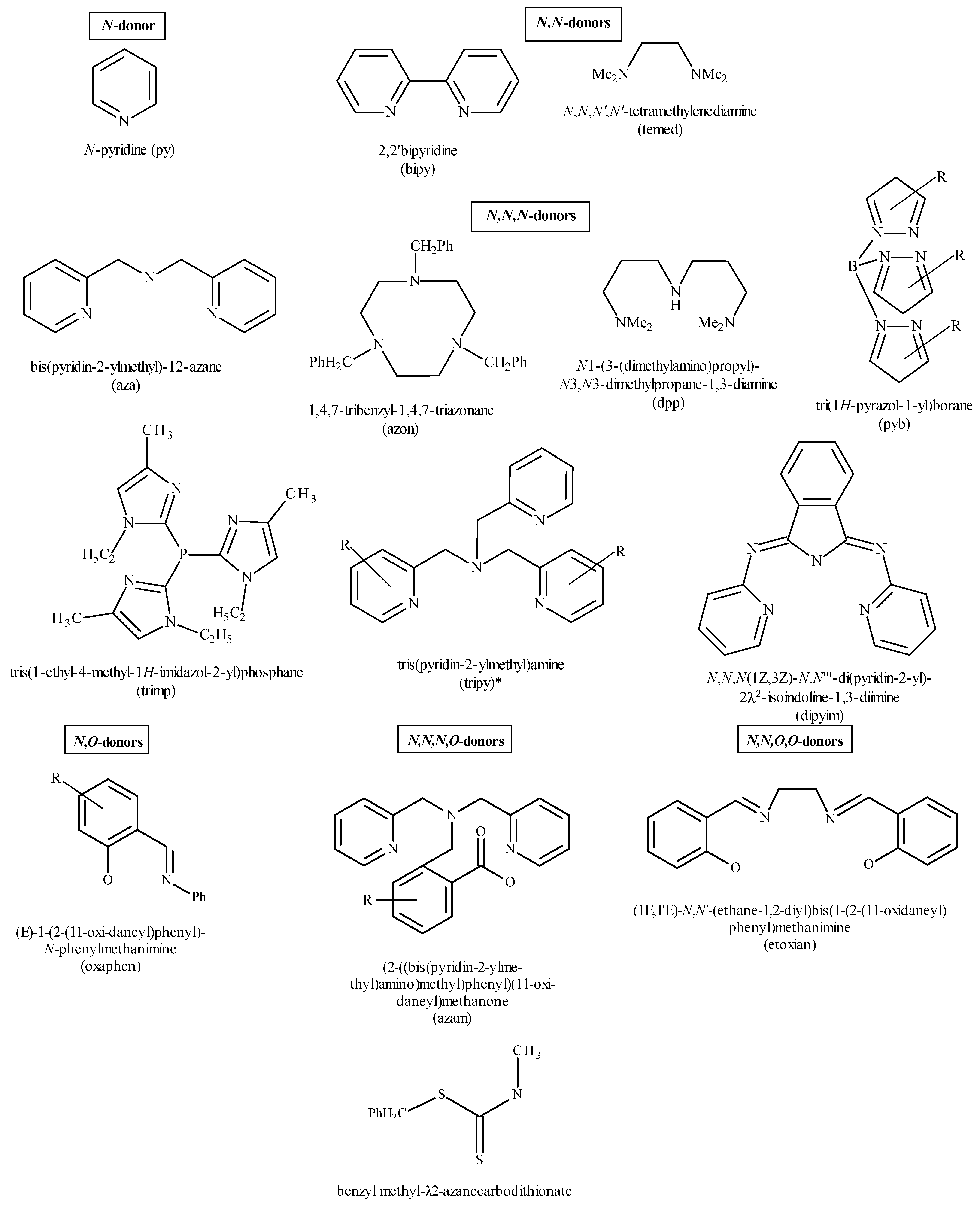{kind=link}
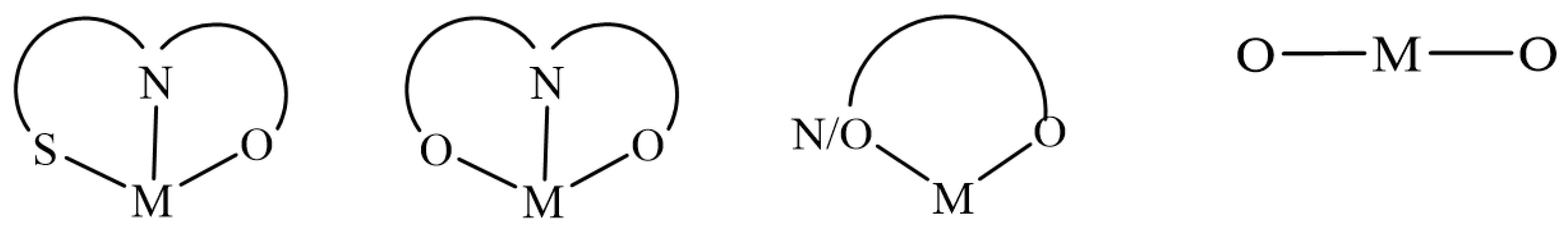{kind=link}
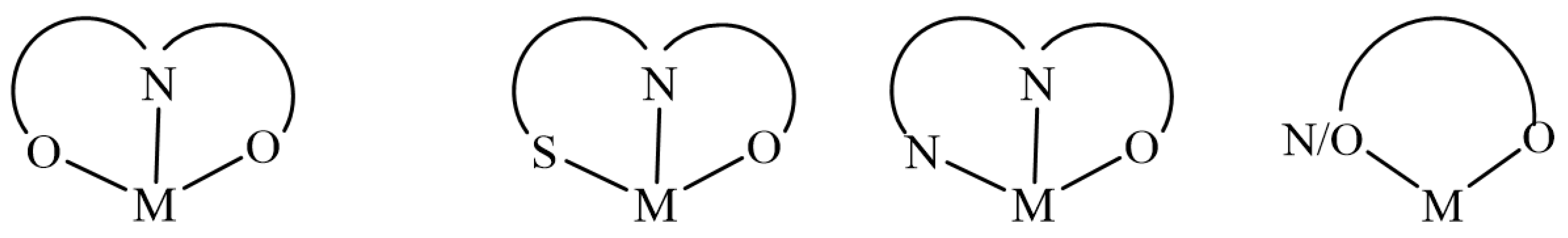{kind=link}
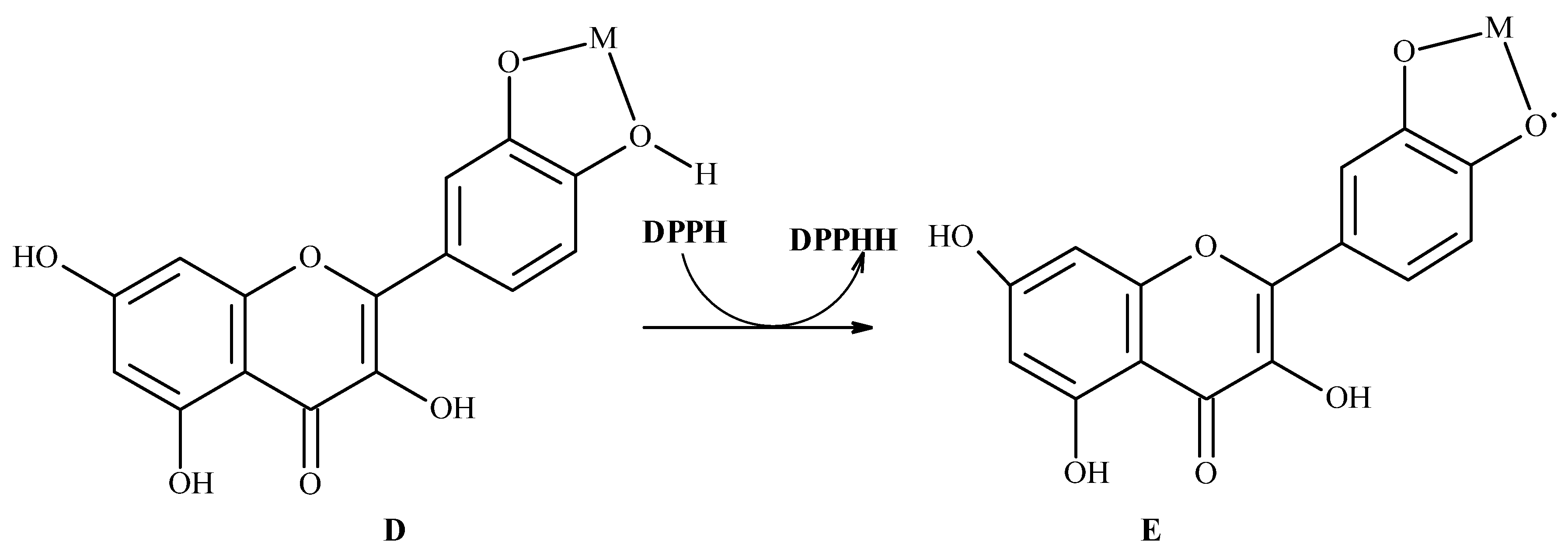{kind=link}
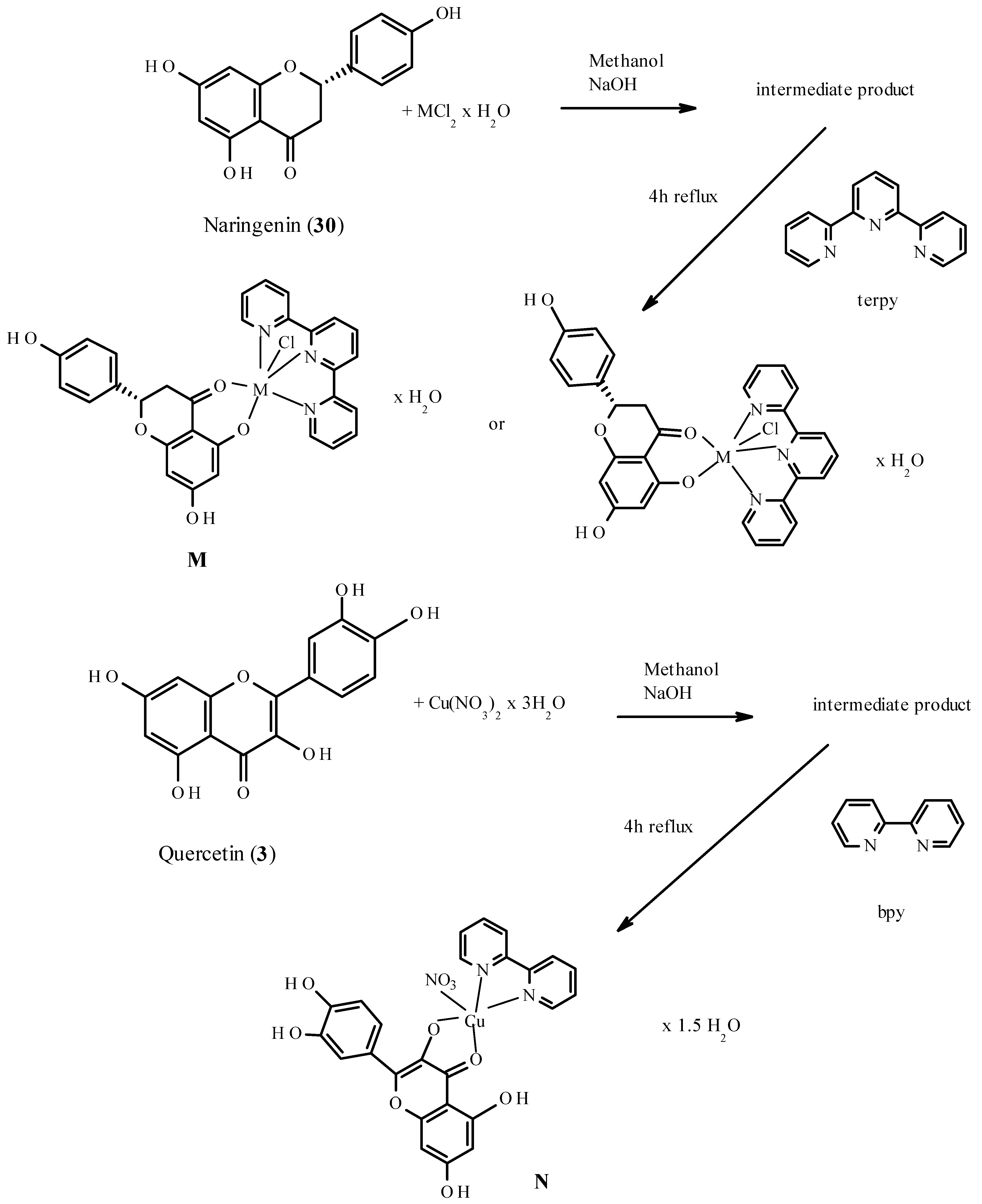{kind=link}
{kind=link}
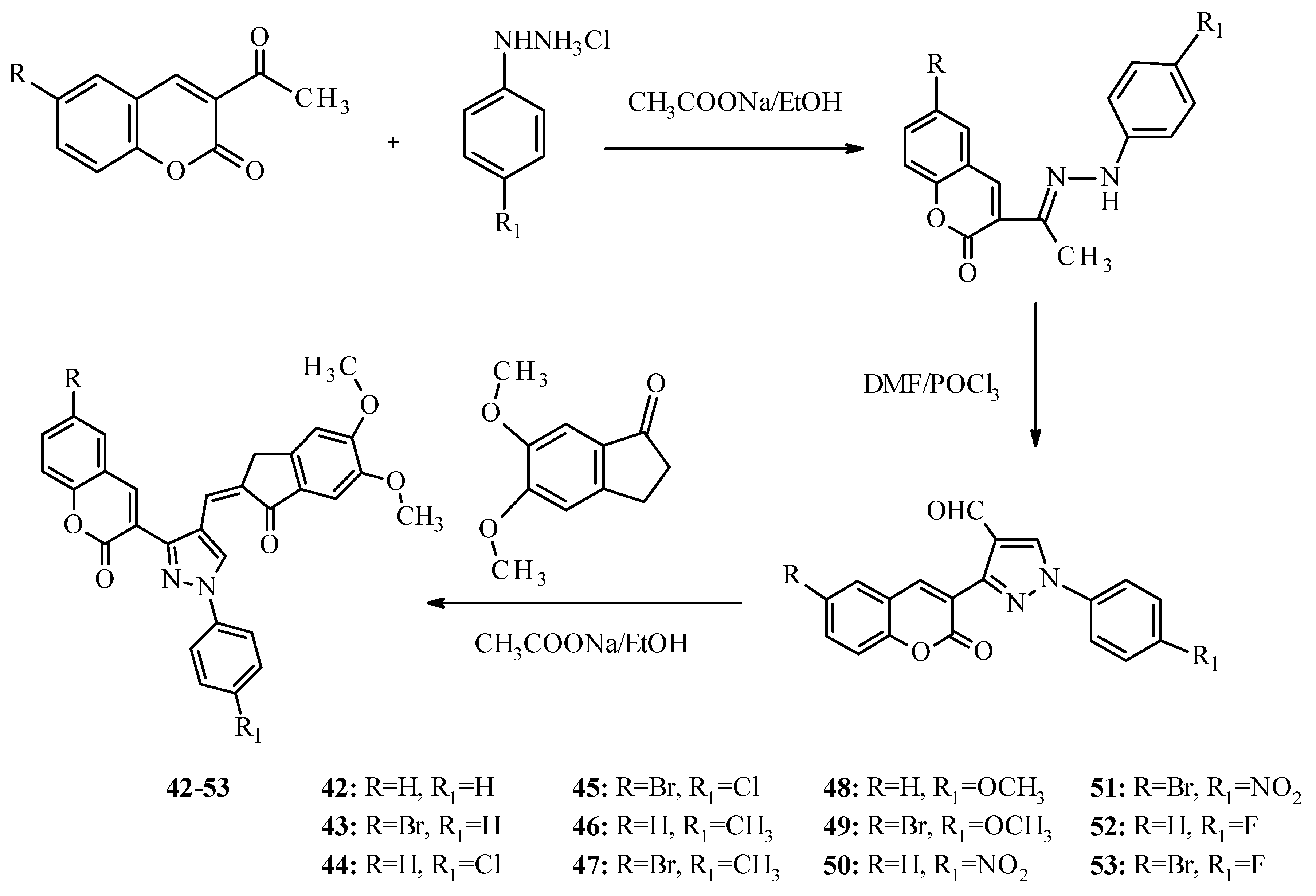{kind=link}
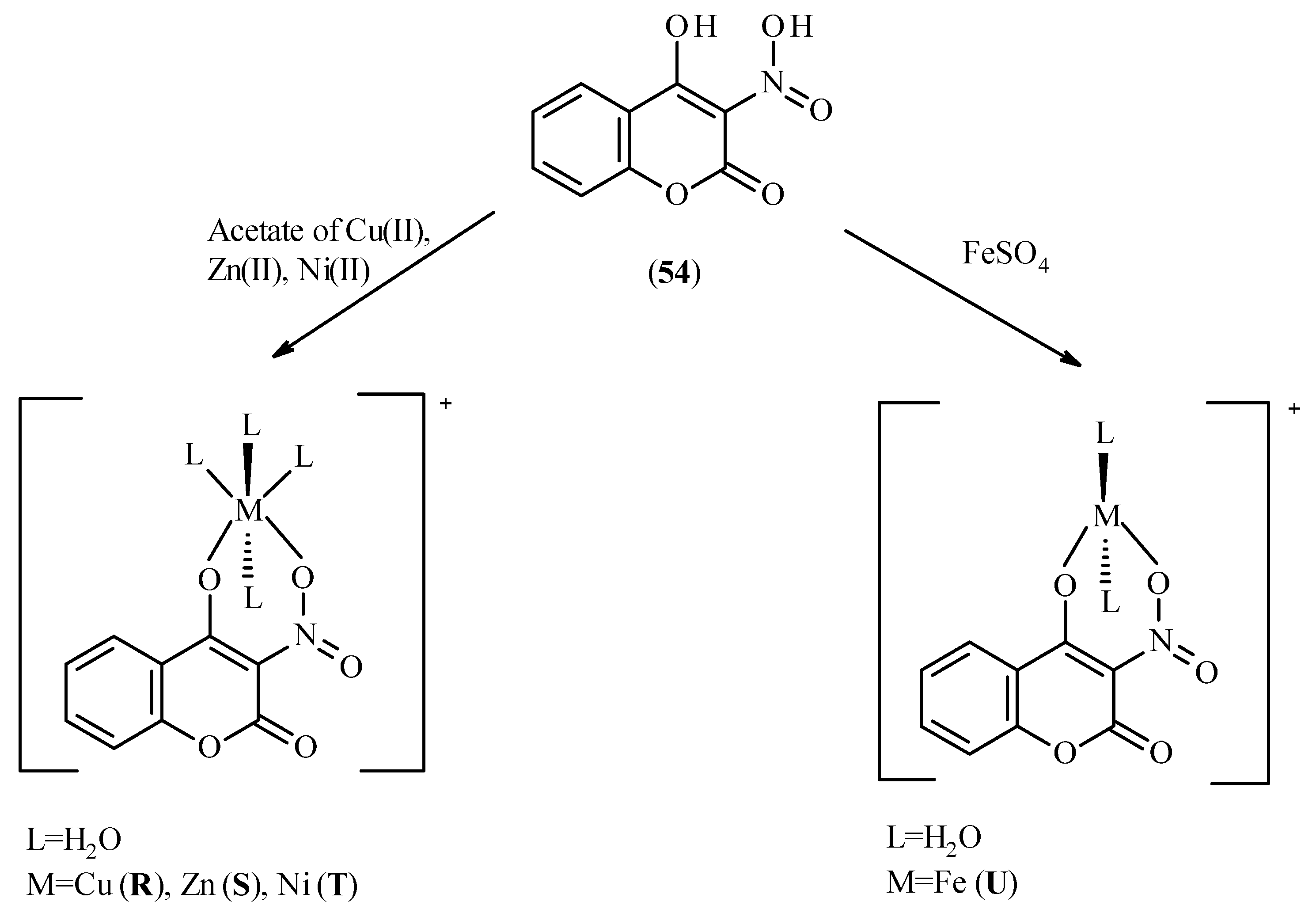{kind=link}
| Compound | TEC50 [min] | AE (104) | Classification According to AE | DPPH Radical Scavenging Activities [1 × 10−4 M] |
|---|---|---|---|---|
| Quercetin (3) | 4.52 | 7.95 | High | 61.83 |
| [Cu2(L1)(H2O)4]Cl2 | 3.78 | 11.56 | Very high | 63.15 |
| [Fe2(L1)(H2O)8]Cl2 | 3.24 | 14.67 | Very high | 63.94 |
| [Al2(L1)(H2O)8]Cl2 | 1.85 | 32.33 | Very high | 65.31 |
| [Zn2(L1)(H2O)4]Cl2 | 2.40 | 21.70 | Very high | 64.24 |
| Rutin (26) | 62.73 | 0.09 | Low | 14.66 |
| [Cu3(L2)(H2O)6]Cl2 | 18.04 | 1.10 | Medium | 24.70 |
| [Fe3(L2)(H2O)12]Cl2 | 13.85 | 1.54 | Medium | 25.54 |
| [Al3(L2)(H2O)12]Cl2 | 9.15 | 2.91 | Medium | 28.19 |
| [Zn3(L2)(H2O)6]Cl2 | 11.96 | 1.96 | Medium | 26.57 |
| Galangin (27) | 44.11 | 0.16 | Low | 29.17 |
| [Cu(L3)(H2O)2]Cl | 10.07 | 2.6 | Medium | 30.76 |
| [Fe(L3)(H2O)4]Cl | 9.10 | 3.06 | Medium | 30.82 |
| [Al(L3)(H2O)4]Cl2 | 5.20 | 6.48 | High | 33.49 |
| [Zn(L3)(H2O)2]Cl | 6.93 | 4.48 | Medium | 32.12 |
| Catechin (13) | 18.88 | 1.31 | Medium | 48.96 |
| [Cu(L4)(H2O)6]Cl | 15.59 | 1.81 | Medium | 51.01 |
| [Fe(L2)(H2O)12]Cl | 13.39 | 2.16 | Medium | 51.27 |
| [Al(L4)(H2O)12]Cl2 | 8.65 | 4.08 | Medium | 54.34 |
| [Zn (L4)(H2O)6]Cl | 10.92 | 2.93 | Medium | 52.19 |
| Compound | Substituent | ||
|---|---|---|---|
| R1 | R2 | ||
| 36 |  | 3′-(CF3)-Phenyl | 3″-(CF3)-benzoyl |
| 37 | 3′-(Cl)-Phenyl | 3″-(Cl)-benzoyl | |
| 38 | 3′-(OCH3)-Phenyl | 3″-(OCH3)-benzoyl | |
| 39 | 4′-(F)-Phenyl | 4′-(F)-benzoyl | |
| 40 | 4′-(NO2)-Phenyl | 4′-(NO2)-benzoyl | |
| 41 | 4′-(OCH3)-Phenyl | 4′-(OCH3)-benzoyl | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mucha, P.; Skoczyńska, A.; Małecka, M.; Hikisz, P.; Budzisz, E. Overview of the Antioxidant and Anti-Inflammatory Activities of Selected Plant Compounds and Their Metal Ions Complexes. Molecules 2021, 26, 4886. https://doi.org/10.3390/molecules26164886
Mucha P, Skoczyńska A, Małecka M, Hikisz P, Budzisz E. Overview of the Antioxidant and Anti-Inflammatory Activities of Selected Plant Compounds and Their Metal Ions Complexes. Molecules. 2021; 26(16):4886. https://doi.org/10.3390/molecules26164886
Chicago/Turabian StyleMucha, Paulina, Anna Skoczyńska, Magdalena Małecka, Paweł Hikisz, and Elzbieta Budzisz. 2021. "Overview of the Antioxidant and Anti-Inflammatory Activities of Selected Plant Compounds and Their Metal Ions Complexes" Molecules 26, no. 16: 4886. https://doi.org/10.3390/molecules26164886
APA StyleMucha, P., Skoczyńska, A., Małecka, M., Hikisz, P., & Budzisz, E. (2021). Overview of the Antioxidant and Anti-Inflammatory Activities of Selected Plant Compounds and Their Metal Ions Complexes. Molecules, 26(16), 4886. https://doi.org/10.3390/molecules26164886