
The Coffee Diterpene, Kahweol, Ameliorates Pancreatic β-Cell Function in Streptozotocin (STZ)-Treated Rat INS-1 Cells through NF-kB and p-AKT/Bcl-2 Pathways

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Kahweol Protects INS-1 Cells against STZ-Induced Damage
2.2. Kahweol Increases Insulin Secretion and Content in STZ-Treated Cells
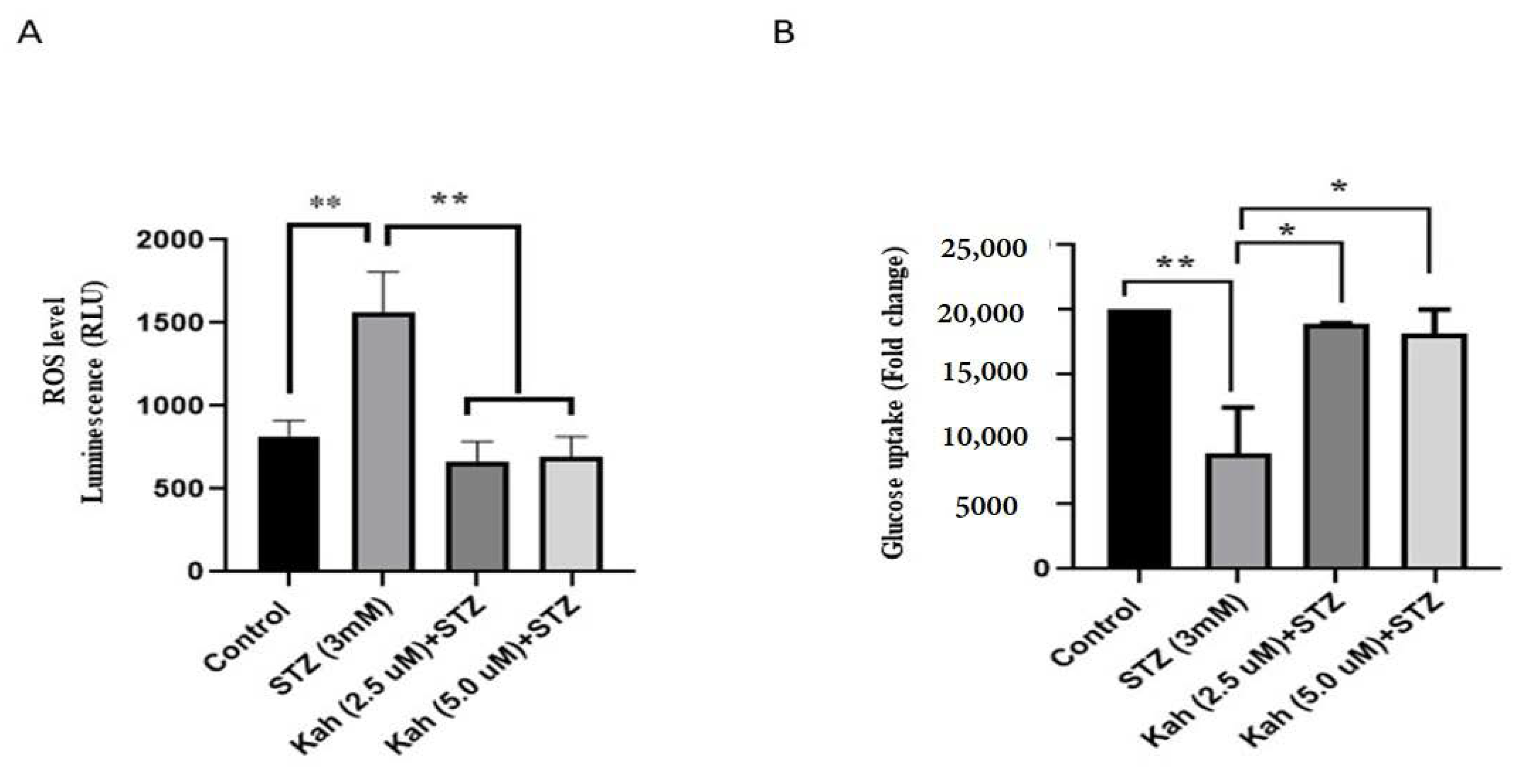
2.3. Kahweol Reduces Intracellular ROS and Increases Glucose Uptake in INS-1 Cells
2.4. Kahweol’s Effect on NF-κB Activation, p-AKT and Bcl-2, HMOX1, ID1 and ID3 Protein Expression
3. Discussion
4. Materials and Methods
4.1. Culturing of INS-1 Cell Line
4.2. Cell Viability Assay
4.3. Apoptosis Assay
4.4. Insulin Secretion Assays
4.5. Glucose Uptake Assay
4.6. Intracellular Reactive Oxygen Species (ROS) Measurement
4.7. Western Blot Analysis
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Sapra, A.; Bhandari, P. Diabetes Mellitus; StatPearls: Treasure Island, FL, USA, 2021. [Google Scholar]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martín, C. Pathophysiology of Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef]
- Anastasiou, I.A.; Eleftheriadou, I.; Tentolouris, A.; Koliaki, C.; Kosta, O.A.; Tentolouris, N. The Effect of Oxidative Stress and Antioxidant Therapies on Pancreatic B-cell Dysfunction: Results from in Vitro and in Vivo Studies. Curr. Med. Chem. 2020, 28, 1328–1346. [Google Scholar] [CrossRef]
- Ding, M.; Bhupathiraju, S.N.; Chen, M.; van Dam, R.M.; Hu, F.B. Caffeinated and decaffeinated coffee consumption and risk of type 2 diabetes: A systematic review and a dose-response meta-analysis. Diabetes Care 2014, 37, 569–586. [Google Scholar] [CrossRef] [Green Version]
- Huxley, R.; Lee, C.M.Y.; Barzi, F.; Timmermeister, L.; Czernichow, S.; Perkovic, V.; Grobbee, D.E.; Batty, D.; Woodward, M. Coffee, decaffeinated coffee, and tea consumption in relation to incident type 2 diabetes mellitus: A systematic review with meta-analysis. Arch. Intern. Med. 2009, 169, 2053–2063. [Google Scholar] [CrossRef]
- Shah, S.; Kumar, A. Coffee: Constituents and Health Benefits. Biotechnol. J. Int. 2020, 24, 22–38. [Google Scholar] [CrossRef]
- Campanha, F.G.; Dias, R.C.E.; Benassi, M.d.T. Discrimination of Coffee Species Using Kahweol and Cafestol: Effects of Roasting and of Defects; 2010. Available online: http://www.sbicafe.ufv.br:80/handle/123456789/3921 (accessed on 19 August 2021).
- Chartier, A.; Beaumesnil, M.; de Oliveira, A.L.; Elfakir, C.; Bostyn, S. Optimization of the isolation and quantitation of kahweol and cafestol in green coffee oil. Talanta 2013, 117, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Ismail, T.; Donati-Zeppa, S.; Akhtar, S.; Turrini, E.; Layla, A.; Sestili, P.; Fimognari, C. Coffee in cancer chemoprevention: An updated review. Expert Opin. Drug Metab. Toxicol. 2021, 17, 69–85. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Wang, C.; Xu, J.; Wang, S. Cafestol and Kahweol: A Review on Their Bioactivities and Pharmacological Properties. Int. J. Mol. Sci. 2019, 20, 4238. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.Y.; Kim, M.K.; Lee, S.H.; Hwang, J.S.; Park, K.G.; Jang, B.K. Kahweol Ameliorates the Liver Inflammation through the Inhibition of NF-kappaB and STAT3 Activation in Primary Kupffer Cells and Primary Hepatocytes. Nutrients 2018, 10, 863. [Google Scholar] [CrossRef] [Green Version]
- Alperet, D.J.; Rebello, S.A.; Khoo, E.Y.-H.; Tay, Z.; Seah, S.S.-Y.; Tai, B.-C.; Emady-Azar, S.; Chou, C.J.; Darimont, C.; van Dam, R.M. A randomized placebo-controlled trial of the effect of coffee consumption on insulin sensitivity: Design and baseline characteristics of the Coffee for METabolic Health (COMETH) study. Contem. Clin. Trials Commun. 2016, 4, 105–117. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, S.; Oksbjerg, N.; Young, J.; Jeppesen, P. Caffeic acid, naringenin and quercetin enhance glucose-stimulated insulin secretion and glucose sensitivity in INS-1E cells. Diabetes Obes. Metab. 2014, 16, 602–612. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Gomez, B.; Lezama, A.; Amigo-Benavent, M.; Ullate, M.; Herrero, M.; Martín, M.Á.; Mesa, M.D.; del Castillo, M.D. Insights on the health benefits of the bioactive compounds of coffee silverskin extract. J. Funct. Foods 2016, 25, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Mellbye, F.B.; Jeppesen, P.B.; Hermansen, K.; Gregersen, S. Cafestol, a bioactive substance in coffee, stimulates insulin secretion and increases glucose uptake in muscle cells: Studies in vitro. J. Nat. Prod. 2015, 78, 2447–2451. [Google Scholar] [CrossRef]
- Muley, A.; Muley, P.; Shah, M. Coffee to reduce risk of type 2 diabetes? A systematic review. Curr. Diabetes Rev. 2012, 8, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, G.D.; Zemdegs, J.C.; Theodoro, J.A.; Mota, J.F. Does long-term coffee intake reduce type 2 diabetes mellitus risk? Diabetol. Metab. Syndr. 2009, 1, 6. [Google Scholar] [CrossRef] [Green Version]
- Van Dam, R.M.; Willett, W.C.; Manson, J.E.; Hu, F.B. Coffee, caffeine, and risk of type 2 diabetes: A prospective cohort study in younger and middle-aged US women. Diabetes Care 2006, 29, 398–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Temple, J.L.; Bernard, C.; Lipshultz, S.E.; Czachor, J.D.; Westphal, J.A.; Mestre, M.A. The Safety of Ingested Caffeine: A Comprehensive Review. Front. Psychiatry 2017, 8, 80. [Google Scholar] [CrossRef] [Green Version]
- Hwang, Y.P.; Jeong, H.G. The coffee diterpene kahweol induces heme oxygenase-1 via the PI3K and p38/Nrf2 pathway to protect human dopaminergic neurons from 6-hydroxydopamine-derived oxidative stress. FEBS Lett. 2008, 582, 2655–2662. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Jung, K.S.; Lee, K.J.; Na, H.K.; Chun, H.-K.; Kho, Y.-H.; Jeong, H.G. The coffee diterpene kahweol suppress the inducible nitric oxide synthase expression in macrophages. Cancer Lett. 2004, 213, 147–154. [Google Scholar] [CrossRef]
- Islam, M.S. Experimental rodent models of type 2 diabetes: A review. Methods Find. Exp. Clin. Pharmacol. 2009, 31, 249–261. [Google Scholar] [CrossRef]
- Eguchi, N.; Vaziri, N.D.; Dafoe, D.C.; Ichii, H. The Role of Oxidative Stress in Pancreatic beta Cell Dysfunction in Diabetes. Int. J. Mol. Sci. 2021, 22, 1509. [Google Scholar] [CrossRef]
- Garcia-Sanchez, A.; Miranda-Diaz, A.G.; Cardona-Munoz, E.G. The Role of Oxidative Stress in Physiopathology and Pharmacological Treatment with Pro- and Antioxidant Properties in Chronic Diseases. Oxid. Med. Cell. Longev. 2020, 2020, 2082145. [Google Scholar] [CrossRef]
- Karin, M.; Yamamoto, Y.; Wang, Q.M. The IKK NF-kappa B system: A treasure trove for drug development. Nat. Rev. Drug Discov. 2004, 3, 17–26. [Google Scholar] [CrossRef]
- Glab, J.A.; Cao, Z.; Puthalakath, H. Bcl-2 family proteins, beyond the veil. Int. Rev. Cell Mol. Biol. 2020, 351, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Prasad, S.; Sitasawad, S.L. Multiple antioxidants improve cardiac complications and inhibit cardiac cell death in streptozotocin-induced diabetic rats. PLoS ONE 2013, 8, e67009. [Google Scholar] [CrossRef]
- Suanarunsawat, T.; Anantasomboon, G.; Piewbang, C. Anti-diabetic and anti-oxidative activity of fixed oil extracted from Ocimum sanctum L. leaves in diabetic rats. Exp. Ther. Med. 2016, 11, 832–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Wang, F.; Zhang, J.; Wang, K.; De, X.; Li, L.; Zhang, Y. Typical phthalic acid esters induce apoptosis by regulating the PI3K/Akt/Bcl-2 signaling pathway in rat insulinoma cells. Ecotoxicol. Environ. Saf. 2021, 208, 111461. [Google Scholar] [CrossRef] [PubMed]
- Lenzen, S.; Drinkgern, J.; Tiedge, M. Low antioxidant enzyme gene expression in pancreatic islets compared with various other mouse tissues. Free Radic. Biol. Med. 1996, 20, 463–466. [Google Scholar] [CrossRef]
- Bensellam, M.; Montgomery, M.K.; Luzuriaga, J.; Chan, J.Y.; Laybutt, D.R. Inhibitor of differentiation proteins protect against oxidative stress by regulating the antioxidant-mitochondrial response in mouse beta cells. Diabetologia 2015, 58, 758–770. [Google Scholar] [CrossRef]
- Lacraz, G.; Figeac, F.; Movassat, J.; Kassis, N.; Coulaud, J.; Galinier, A.; Leloup, C.; Bailbe, D.; Homo-Delarche, F.; Portha, B. Diabetic beta-cells can achieve self-protection against oxidative stress through an adaptive up-regulation of their antioxidant defenses. PLoS ONE 2009, 4, e6500. [Google Scholar] [CrossRef] [Green Version]
- Taneera, J.; Dhaiban, S.; Hachim, M.; Mohammed, A.K.; Mukhopadhyay, D.; Bajbouj, K.; Hamoudi, R.; Salehi, A.; Hamad, M. Reduced expression of Chl1 gene impairs insulin secretion by down-regulating the expression of key molecules of β-cell function. Exp. Clin. Endocrinol. Diabetes 2019, 127, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Taneera, J.; Mohammed, I.; Mohammed, A.K.; Hachim, M.; Dhaiban, S.; Malek, A.; Duner, P.; Elemam, N.M.; Sulaiman, N.; Hamad, M.; et al. Orphan G-protein coupled receptor 183 (GPR183) potentiates insulin secretion and prevents glucotoxicity-induced beta-cell dysfunction. Mol. Cell. Endocrinol. 2020, 499, 110592. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Zhao, M.; Ren, Y.; Wu, Y.; Yang, J. Sesamin suppresses STZ induced INS-1 cell apoptosis through inhibition of NF-kappaB activation and regulation of Bcl-2 family protein expression. Eur. J. Pharmacol. 2015, 750, 52–58. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Huneidi, W.; Anjum, S.; Bajbouj, K.; Abu-Gharbieh, E.; Taneera, J. The Coffee Diterpene, Kahweol, Ameliorates Pancreatic β-Cell Function in Streptozotocin (STZ)-Treated Rat INS-1 Cells through NF-kB and p-AKT/Bcl-2 Pathways. Molecules 2021, 26, 5167. https://doi.org/10.3390/molecules26175167
El-Huneidi W, Anjum S, Bajbouj K, Abu-Gharbieh E, Taneera J. The Coffee Diterpene, Kahweol, Ameliorates Pancreatic β-Cell Function in Streptozotocin (STZ)-Treated Rat INS-1 Cells through NF-kB and p-AKT/Bcl-2 Pathways. Molecules. 2021; 26(17):5167. https://doi.org/10.3390/molecules26175167
Chicago/Turabian StyleEl-Huneidi, Waseem, Shabana Anjum, Khuloud Bajbouj, Eman Abu-Gharbieh, and Jalal Taneera. 2021. "The Coffee Diterpene, Kahweol, Ameliorates Pancreatic β-Cell Function in Streptozotocin (STZ)-Treated Rat INS-1 Cells through NF-kB and p-AKT/Bcl-2 Pathways" Molecules 26, no. 17: 5167. https://doi.org/10.3390/molecules26175167
APA StyleEl-Huneidi, W., Anjum, S., Bajbouj, K., Abu-Gharbieh, E., & Taneera, J. (2021). The Coffee Diterpene, Kahweol, Ameliorates Pancreatic β-Cell Function in Streptozotocin (STZ)-Treated Rat INS-1 Cells through NF-kB and p-AKT/Bcl-2 Pathways. Molecules, 26(17), 5167. https://doi.org/10.3390/molecules26175167