
2-IPMA Ameliorates PM2.5-Induced Inflammation by Promoting Primary Ciliogenesis in RPE Cells

 ,
, 
 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Image-Based Fecal Metabolite Library Screening
2.3. Reagents
2.4. Gene Knockdown
2.5. Oxidative Stress Measurement
2.6. Western Blot Analysis
2.7. Cilia Staining and Counting
2.8. ELISA Assay
2.9. Statistical Analysis
3. Results and Discussion
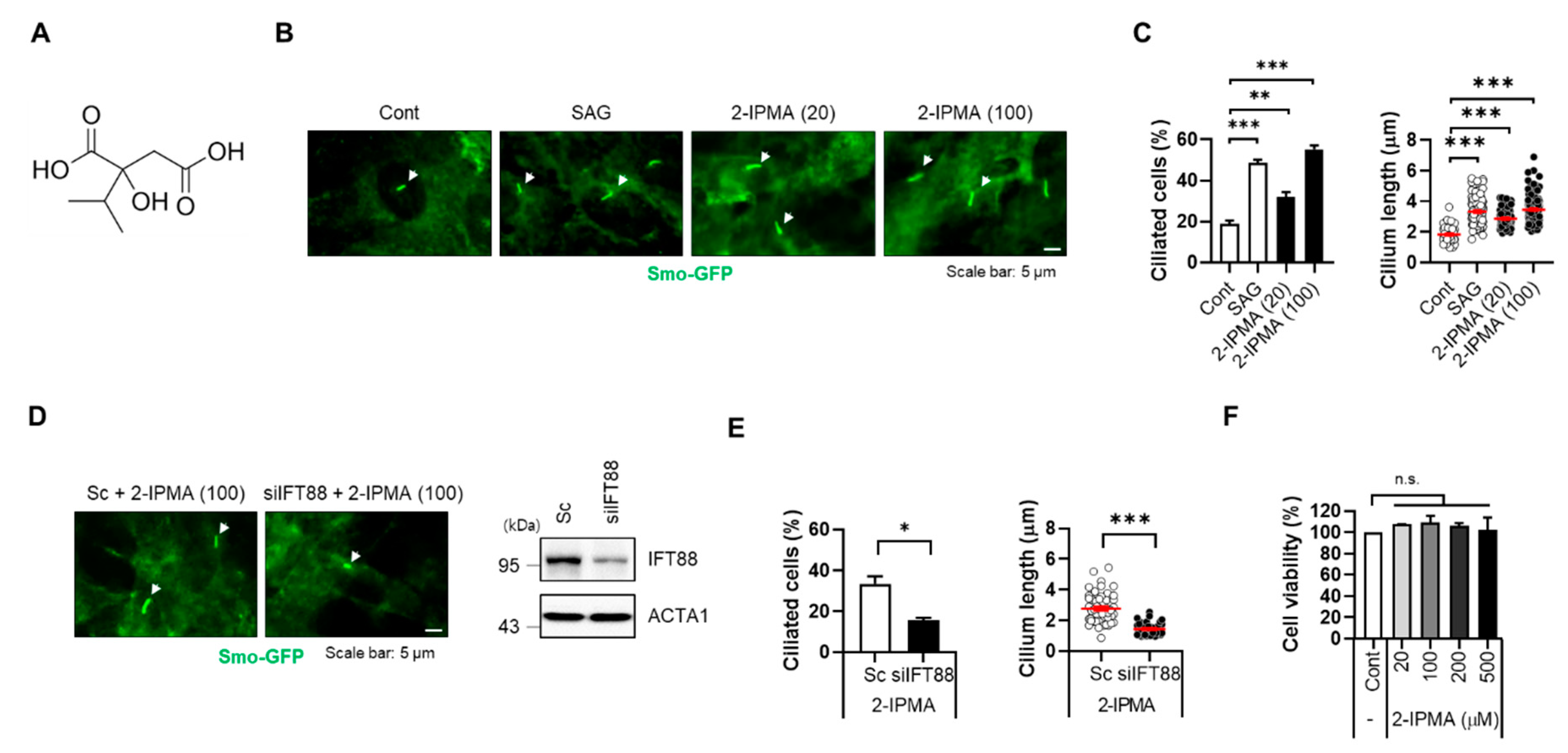
3.1. 2-IPMA Promotes Primary Ciliogenesis in RPE Cells
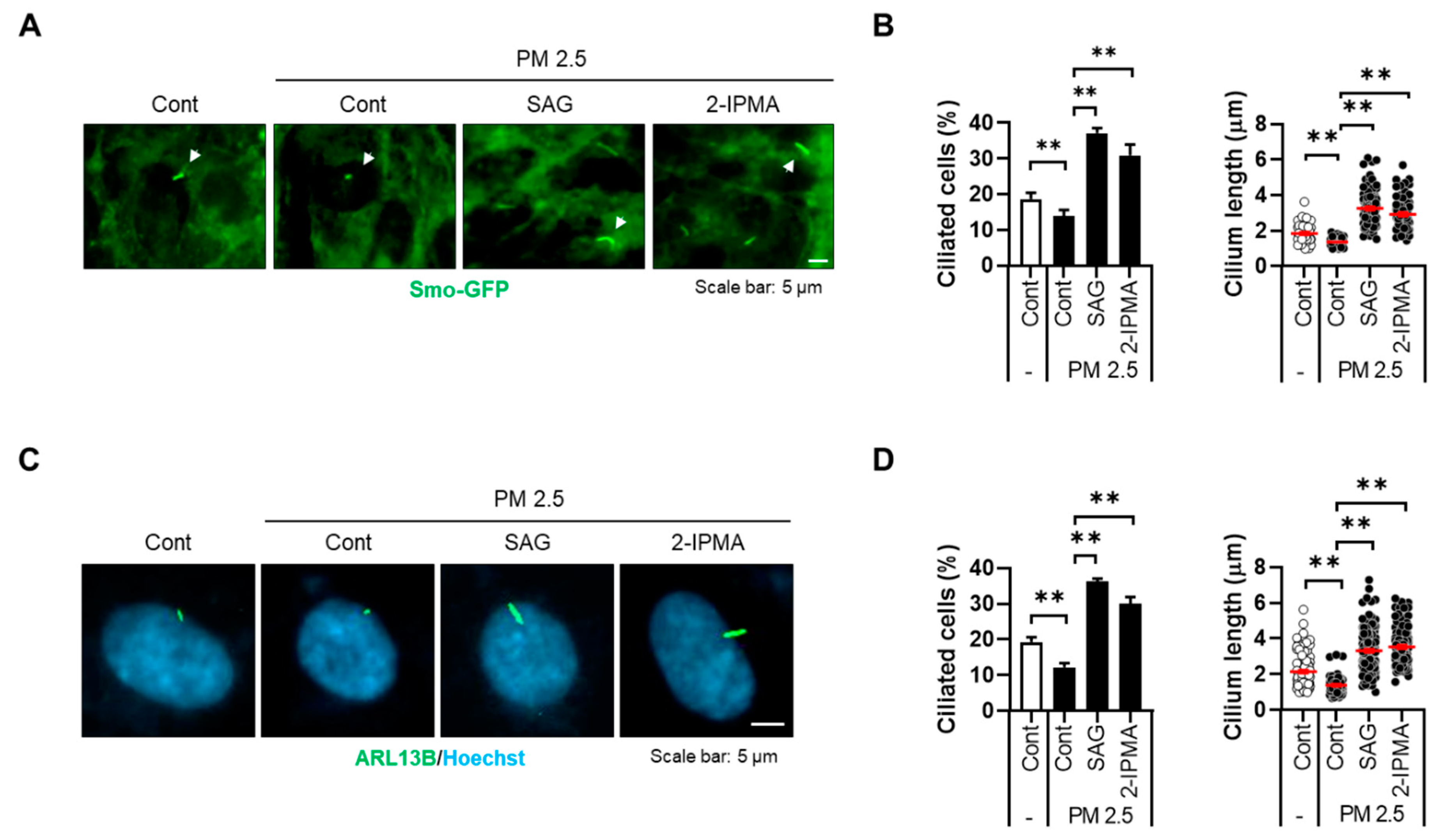
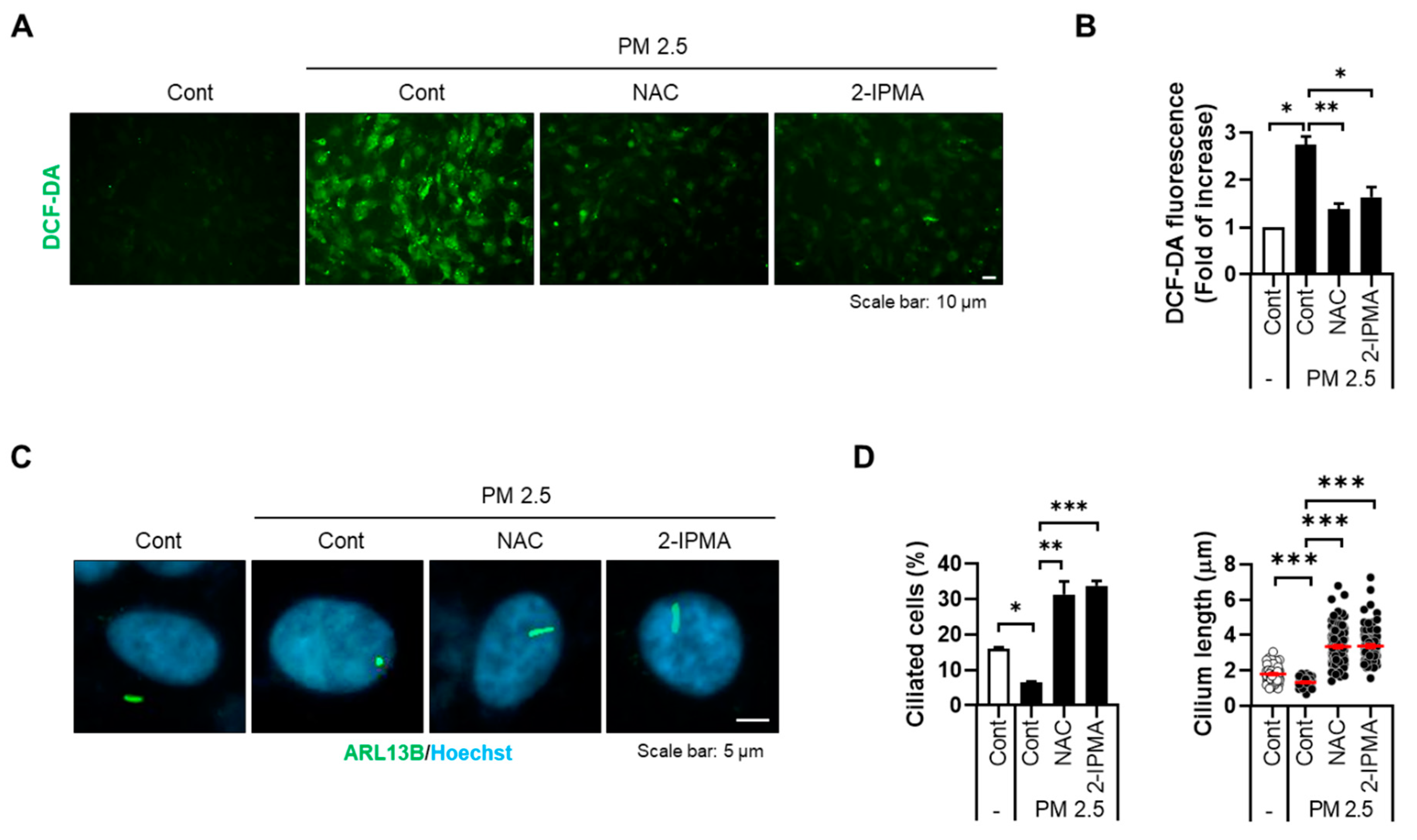
3.2. 2-IPMA Inhibits PM2.5-Mediated Ciliary Dysgenesis in RPE Cells
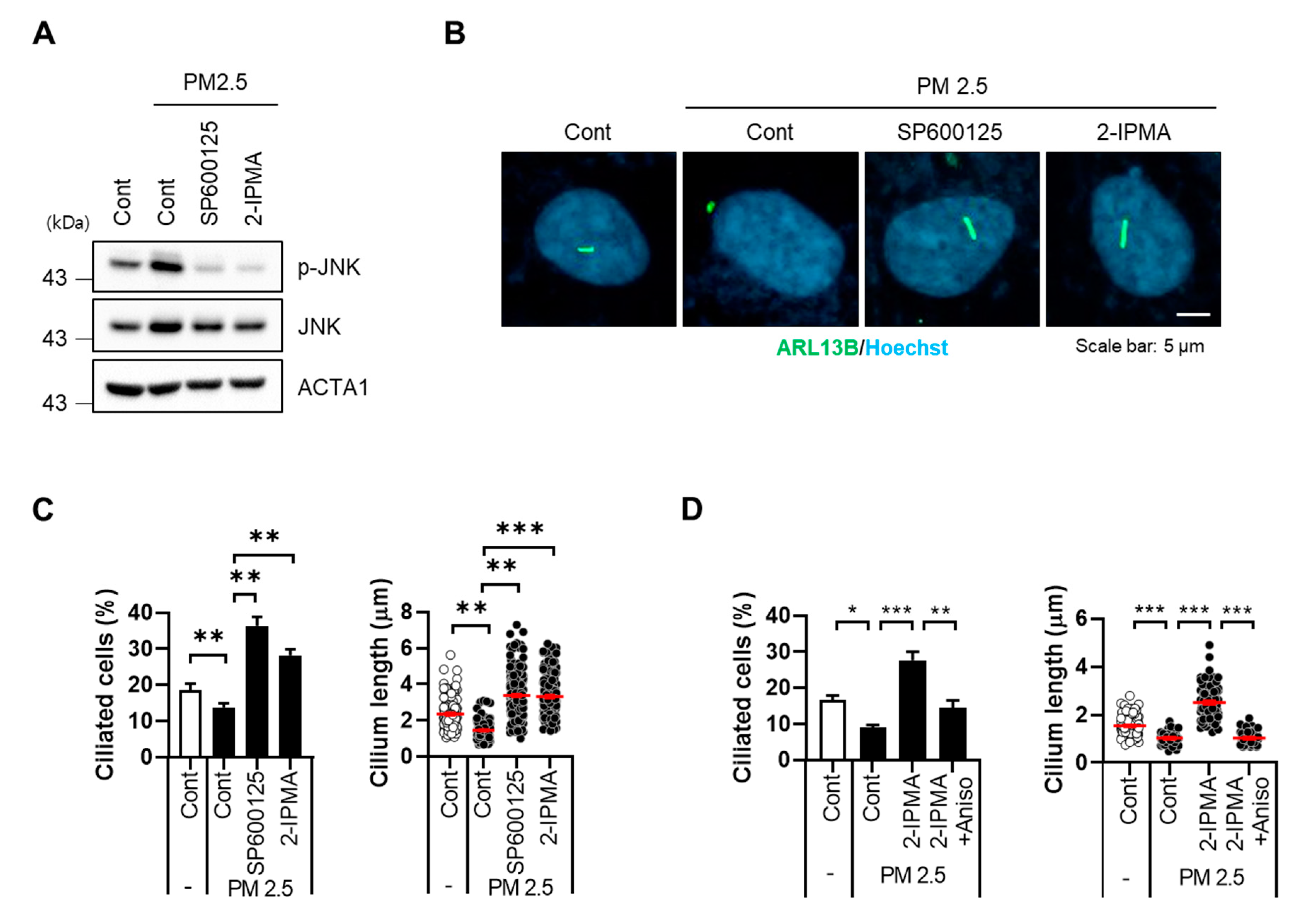
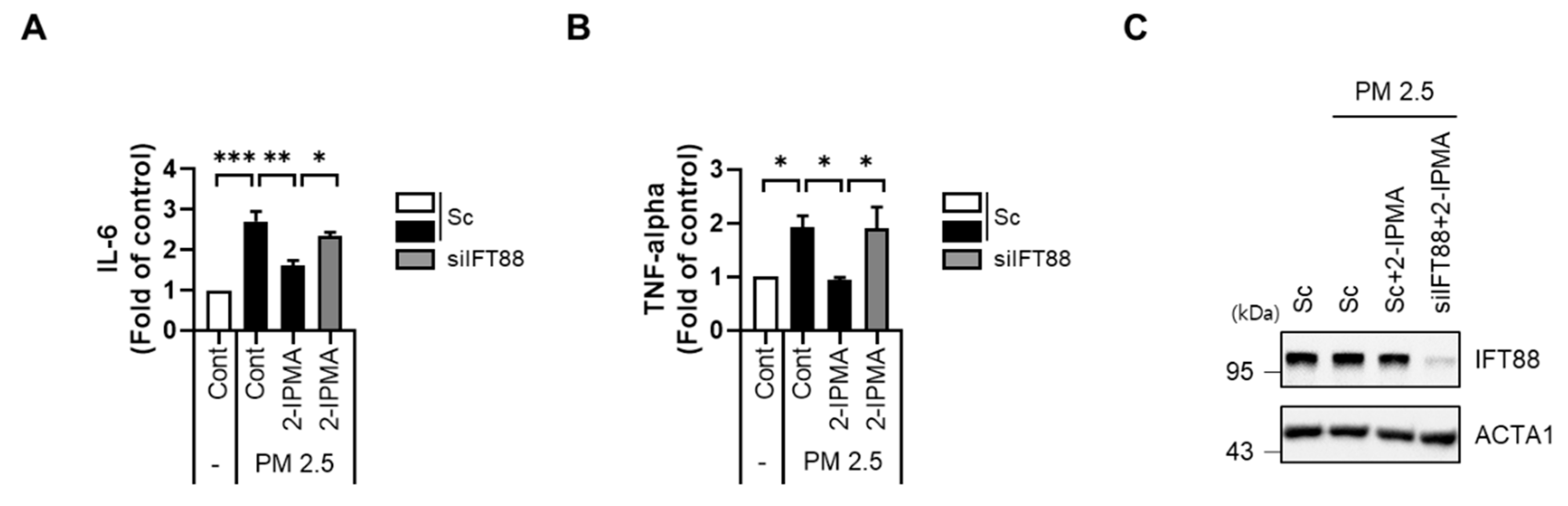
3.3. 2-IPMA Inhibits Activation of JNK and Production of Proinflammatory Cytokines in PM2.5-Treated RPE Cells
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Anderson, J.O.; Thundiyil, J.G.; Stolbach, A. Clearing the air: A review of the effects of particulate matter air pollution on human health. J. Med. Toxicol. 2012, 8, 166–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krutmann, J.; Liu, W.; Li, L.; Pan, X.; Crawford, M.; Sore, G.; Seite, S. Pollution and skin: From epidemiological and mechanistic studies to clinical implications. J. Dermatol. Sci. 2014, 76, 163–168. [Google Scholar] [CrossRef]
- Chuang, K.J.; Chan, C.C.; Su, T.C.; Lee, C.T.; Tang, C.S. The effect of urban air pollution on inflammation, oxidative stress, coagulation, and autonomic dysfunction in young adults. Am. J. Respir. Crit. Care Med. 2007, 176, 370–376. [Google Scholar] [CrossRef]
- Filep, Á.; Fodor, G.H.; Kun-Szabó, F.; Tiszlavicz, L.; Rázga, Z.; Bozsó, G.; Bozóki, Z.; Szabó, G.; Peták, F. Exposure to urban PM1 in rats: Development of bronchial inflammation and airway hyperresponsiveness. Respir. Res. 2016, 17, 26. [Google Scholar] [CrossRef] [Green Version]
- Chenxu, G.; Minxuan, X.; Yuting, Q.; Tingting, G.; Jinxiao, L.; Mingxing, W.; Sujun, W.; Yongjie, M.; Deshuai, L.; Qiang, L.; et al. iRhom2 loss alleviates renal injury in long-term PM2.5-exposed mice by suppression of inflammation and oxidative stress. Redox Biol. 2018, 19, 147–157. [Google Scholar] [CrossRef]
- Gillespie, P.; Tajuba, J.; Lippmann, M.; Chen, L.C.; Veronesi, B. Particulate matter neurotoxicity in culture is size-dependent. Neurotoxicology 2013, 36, 112–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, W.; You, J.; Zhi, C.; Li, L. The toxicity of ambient fine particulate matter (PM2.5) to vascular endothelial cells. J. Appl. Toxicol. 2021, 41, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, H.; Marshall, W.F. Ciliogenesis: Building the cell’s antenna. Nat. Rev. Mol. Cell Biol. 2011, 12, 222–234. [Google Scholar] [CrossRef] [PubMed]
- Anvarian, Z.; Mykytyn, K.; Mukhopadhyay, S.; Pedersen, L.B.; Christensen, S.T. Cellular signalling by primary cilia in development, organ function and disease. Nat. Rev. Nephrol. 2019, 15, 199–219. [Google Scholar] [CrossRef] [PubMed]
- Satir, P.; Pedersen, L.B.; Christensen, S.T. The primary cilium at a glance. J. Cell Sci. 2010, 123, 499–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang-Minh, L.B.; Deleyrolle, L.P.; Nakamura, N.S.; Parker, A.K.; Martuscello, R.T.; Reynolds, B.A.; Sarkisian, M.R. PCM1 depletion inhibits glioblastoma cell ciliogenesis and increases cell death and sensitivity to temozolomide. Transl. Oncol. 2016, 9, 392–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Park, K.C.; Sul, H.J.; Hong, H.J.; Kim, K.H.; Kero, J.; Shong, M. Loss of primary cilia promotes mitochondria-dependent apoptosis in thyroid cancer. Sci. Rep. 2021, 11, 4181. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Wang, L.; Tang, F.; Zeng, L.; Zhou, L.; Song, X.; Jia, W.; Chen, J.; Yang, Q. Resveratrol pretreatment decreases ischemic injury and improves neurological function via sonic hedgehog signaling after stroke in rats. Mol. Neurobiol. 2017, 54, 212–226. [Google Scholar] [CrossRef]
- Chen, H.Y.; Welby, E.; Li, T.; Swaroop, A. Retinal disease in ciliopathies: Recent advances with a focus on stem cell-based therapies. Transl. Sci. Rare Dis. 2019, 4, 97–115. [Google Scholar] [CrossRef] [Green Version]
- Larkins, C.E.; Aviles, G.D.G.; East, M.P.; Kahn, R.A.; Caspary, T. Arl13b regulates ciliogenesis and the dynamic localization of Shh signaling proteins. Mol. Biol. Cell 2011, 22, 4694–4703. [Google Scholar] [CrossRef]
- Pazour, G.J.; Baker, S.A.; Deane, J.A.; Cole, D.G.; Dickert, B.L.; Rosenbaum, J.L.; Witman, G.B.; Besharse, J.C. The intraflagellar transport protein, IFT88, is essential for vertebrate photoreceptor assembly and maintenance. J. Cell Biol. 2002, 157, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Bae, J.E.; Choi, H.; Shin, D.W.; Na, H.W.; Park, N.Y.; Kim, J.B.; Jo, D.S.; Cho, M.J.; Lyu, J.H.; Chang, J.H.; et al. Fine particulate matter (PM2.5) inhibits ciliogenesis by increasing SPRR3 expression via c-Jun activation in RPE cells and skin keratinocytes. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Singla, V.; Reiter, J.F. The primary cilium as the cell’s antenna: Signaling at a sensory organelle. Science 2006, 313, 629–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, G.; Jia, H.; Zhao, L.; Cho, D.H.; Fang, J. Small molecule fluorescent probes of protein vicinal dithiols. Chin. Chem. Lett. 2019, 30, 1704–1716. [Google Scholar] [CrossRef]
- Shin, J.H.; Kim, P.S.; Kim, E.S.; Park, S.J.; Jo, Y.K.; Hwang, J.J.; Park, T.J.; Chang, J.W.; Seo, J.H.; Cho, D.H. BIX-01294-induced autophagy regulates elongation of primary cilia. Biochem. Biophys. Res. Commun. 2015, 460, 428–433. [Google Scholar] [CrossRef]
- Strassman, M.; Ceci, L.N. Enzymatic formation of α-isopropylmalic acid, an intermediate in leucine biosynthesis. J. Biol. Chem. 1963, 238, 2445–2452. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, N.; Mao, M.; Zhou, Y.; Wu, Y.; Li, J.; Zhang, W.; Peng, C.; Chen, X.; Li, J. Fine particulate matter (PM2.5) promotes IgE-mediated mast cell activation through ROS/Gadd45b/JNK axis. J. Dermatol. Sci. 2021, 102, 47–57. [Google Scholar] [CrossRef]
- Wang, X.; Chen, M.; Zhong, M.; Hu, Z.; Qiu, L.; Rajagopalan, S.; Fossett, N.G.; Chen, L.C.; Ying, Z. Exposure to concentrated ambient PM2.5 shortens lifespan and induces inflammation-associated signaling and oxidative stress in drosophila. Toxicol. Sci. 2017, 156, 199–207. [Google Scholar]
- Guan, L.; Geng, X.; Stone, C.; Cosky, E.E.; Ji, Y.; Du, H.; Zhang, K.; Sun, Q.; Ding, Y. PM2.5 exposure induces systemic inflammation and oxidative stress in an intracranial atherosclerosis rat model. Environ. Toxicol. 2019, 34, 530–538. [Google Scholar] [CrossRef]
- Chrzanowska, M.; Modrzejewska, A.; Modrzejewska, M. New insight into the role of the complement in the most common types of retinopathy-current literature review. Int. J. Ophthalmol. 2018, 11, 1856. [Google Scholar]
- Das, U.N. Diabetic macular edema, retinopathy and age-related macular degeneration as inflammatory conditions. Arch. Med. Sci. 2016, 12, 1142. [Google Scholar] [CrossRef] [Green Version]
- Forrester, J.V.; Kuffova, L.; Delibegovic, M. The Role of Inflammation in Diabetic Retinopathy. Front. Immunol. 2020, 11, e583687. [Google Scholar] [CrossRef]
- Zhang, J.; Duan, D.; Song, Z.; Liu, T.; Hou, Y.; Fang, J. Small molecules regulating reactive oxygen species homeostasis for cancer therapy. Med. Res. Rev. 2021, 41, 342–394. [Google Scholar] [CrossRef]
- Green, A.J.; McQuaid, S.; Hauser, S.L.; Allen, I.V.; Lyness, R. Ocular pathology in multiple sclerosis: Retinal atrophy and inflammation irrespective of disease duration. Brain 2010, 133, 1591–1601. [Google Scholar] [CrossRef] [Green Version]
- Mao, H.; Seo, S.J.; Biswal, M.R.; Li, H.; Conners, M.; Nandyala, A.; Jones, K.; Le, Y.Z.; Lewin, A.S. Mitochondrial oxidative stress in the retinal pigment epithelium leads to localized retinal degeneration. Invest. Ophthalmol. Vis. Sci. 2014, 55, 4613–4627. [Google Scholar] [CrossRef] [Green Version]
- Sim, H.; Lee, W.; Choo, S.; Park, E.K.; Baek, M.C.; Lee, I.K.; Park, D.H.; Bae, J.S. Sulforaphane Alleviates Particulate Matter-Induced Oxidative Stress in Human Retinal Pigment Epithelial Cells. Front. Med. 2021, 8, 906. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, G.; Lin, Z.; Wang, Y.; He, H.; Liu, T.; Kamp, D.W. Pro-inflammatory response and oxidative stress induced by specific components in ambient particulate matter in human bronchial epithelial cells. Environ. Toxicol. 2016, 31, 923–936. [Google Scholar] [CrossRef] [PubMed]
- Yue, W.; Tong, L.; Liu, X.; Weng, X.; Chen, X.; Wang, D.; Lu, Z.; Xu, Y.; Chen, Y. Short term Pm2.5 exposure caused a robust lung inflammation, vascular remodeling, and exacerbated transition from left ventricular failure to right ventricular hypertrophy. Redox Biol. 2019, 22, 101161. [Google Scholar] [CrossRef]
- Kim, S.; Park, H.; Park, H.; Joung, B.; Kim, E. The acute respiratory exposure by intratracheal instillation of Sprague–Dawley rats with diesel particulate matter induces retinal thickening. Cutan. Ocul. Toxicol. 2016, 35, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Hwang-Bo, H.; Ji, S.Y.; Kim, M.Y.; Kim, S.Y.; Park, C.; Hong, S.H.; Kim, G.Y.; Song, K.S.; Hyun, J.W.; et al. Diesel particulate matter2.5 promotes epithelial-mesenchymal transition of human retinal pigment epithelial cells via generation of reactive oxygen species. Environ. Pollut. 2020, 262, 114301. [Google Scholar] [CrossRef]
- Kim, D.; Kim, Y.; Kim, Y. Effects of β-carotene on expression of selected microRNAs, histone acetylation, and DNA methylation in colon cancer stem cells. J. Cancer Prev. 2019, 24, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Fan, L.X.; Li, K.; Ramchandran, R.; Calvet, J.P.; Li, X. SIRT2 regulates ciliogenesis and contributes to abnormal centrosome amplification caused by loss of polycystin-1. Hum. Mol. Genet. 2014, 23, 1644–1655. [Google Scholar] [CrossRef]
- Tian, J.L.; Qin, H. O-GlcNAcylation regulates primary ciliary length by promoting microtubule disassembly. iScience 2019, 12, 379–391. [Google Scholar] [CrossRef] [Green Version]
- Skogerson, K.; Runnebaum, R.O.N.; Wohlgemuth, G.; De Ropp, J.; Heymann, H.; Fiehn, O. Comparison of gas chromatography-coupled time-of-flight mass spectrometry and 1H nuclear magnetic resonance spectroscopy metabolite identification in white wines from a sensory study investigating wine body. J. Agric. Food Chem. 2009, 57, 6899–6907. [Google Scholar] [CrossRef]
- Ricciutelli, M.; Moretti, S.; Galarini, R.; Sagratini, G.; Mari, M.; Lucarini, S.; Vittori, S.; Caprioli, G. Identification and quantification of new isomers of isopropyl-malic acid in wine by LC-IT and LC-Q-Orbitrap. Food Chem. 2019, 294, 390–396. [Google Scholar] [CrossRef]
- Fliegauf, M.; Benzing, T.; Omran, H. Cilia: Hair-like organelles with many links to disease. Nat. Rev. Mol. Cell Biol. 2007, 8, 880–893. [Google Scholar] [CrossRef]
- Bae, J.E.; Kang, G.M.; Min, S.H.; Jo, D.S.; Jung, Y.K.; Kim, K.; Kim, M.S.; Cho, D.H. Primary cilia mediate mitochondrial stress responses to promote dopamine neuron survival in a Parkinson’s disease model. Cell Death Dis. 2019, 10, 952. [Google Scholar] [CrossRef]
- Choi, B.K.; D’Onofrio, P.M.; Shabanzadeh, A.P.; Koeberle, P.D. Stabilization of primary cilia reduces abortive cell cycle re-entry to protect injured adult CNS neurons from apoptosis. PLoS ONE 2019, 14, e0220056. [Google Scholar]
- Mehan, S.; Meena, H.; Sharma, D.; Sankhla, R. JNK: A stress-activated protein kinase therapeutic strategies and involvement in Alzheimer’s and various neurodegenerative abnormalities. J. Mol. Neurosci. 2011, 43, 376–390. [Google Scholar] [CrossRef]
- Schulz, E.; Dopheide, J.; Schuhmacher, S.; Thomas, S.R.; Chen, K.; Daiber, A.; Wenzel, P.; Münzel, T.; Keaney, J.F., Jr. Clinical perspective. Circulation 2008, 118, 1347–1357. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Li, X.; Nan, A.; Zhang, N.; Chen, L.; Zhou, H.; Zhang, H.; Qiu, M.; Zhu, J.; Ling, Y.; et al. Circular RNA 406961 interacts with ILF2 to regulate PM2.5-induced inflammatory responses in human bronchial epithelial cells via activation of STAT3/JNK pathways. Environ. Int. 2020, 141, 105755. [Google Scholar] [CrossRef] [PubMed]
- Semeraro, F.; Morescalchi, F.; Cancarini, A.; Russo, A.; Rezzola, S.; Costagliola, C. Diabetic retinopathy, a vascular and inflammatory disease: Therapeutic implications. Diabetes Metab. 2019, 45, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, J.; Ma, Q.; Tang, J.; Jiang, M.; Cao, X.; Lin, L.; Kong, N.; Yu, S.; Sood, A.; et al. Chronic exposure to diesel exhaust may cause small airway wall thickening without lumen narrowing: A quantitative computerized tomography study in Chinese diesel engine testers. Part. Fibre Toxicol. 2021, 18, 14. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, J.Y.; Bae, J.-E.; Kim, J.B.; Jo, D.S.; Park, N.Y.; Kim, Y.H.; Lee, H.J.; Kim, S.H.; Kim, S.H.; Jeon, H.B.; et al. 2-IPMA Ameliorates PM2.5-Induced Inflammation by Promoting Primary Ciliogenesis in RPE Cells. Molecules 2021, 26, 5409. https://doi.org/10.3390/molecules26175409
Choi JY, Bae J-E, Kim JB, Jo DS, Park NY, Kim YH, Lee HJ, Kim SH, Kim SH, Jeon HB, et al. 2-IPMA Ameliorates PM2.5-Induced Inflammation by Promoting Primary Ciliogenesis in RPE Cells. Molecules. 2021; 26(17):5409. https://doi.org/10.3390/molecules26175409
Chicago/Turabian StyleChoi, Ji Yeon, Ji-Eun Bae, Joon Bum Kim, Doo Sin Jo, Na Yeon Park, Yong Hwan Kim, Ha Jung Lee, Seong Hyun Kim, So Hyun Kim, Hong Bae Jeon, and et al. 2021. "2-IPMA Ameliorates PM2.5-Induced Inflammation by Promoting Primary Ciliogenesis in RPE Cells" Molecules 26, no. 17: 5409. https://doi.org/10.3390/molecules26175409
APA StyleChoi, J. Y., Bae, J. -E., Kim, J. B., Jo, D. S., Park, N. Y., Kim, Y. H., Lee, H. J., Kim, S. H., Kim, S. H., Jeon, H. B., Na, H. -W., Choi, H., Ryu, H. -Y., Ryoo, Z. Y., Lee, H. -S., & Cho, D. -H. (2021). 2-IPMA Ameliorates PM2.5-Induced Inflammation by Promoting Primary Ciliogenesis in RPE Cells. Molecules, 26(17), 5409. https://doi.org/10.3390/molecules26175409