
Bio-Mechanism of Catechin as Pheromone Signal Inhibitor: Prediction of Antibacterial Agent Action Mode by In Vitro and In Silico Study

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Chemical Composition of Extract Uncaria gambir Roxb.
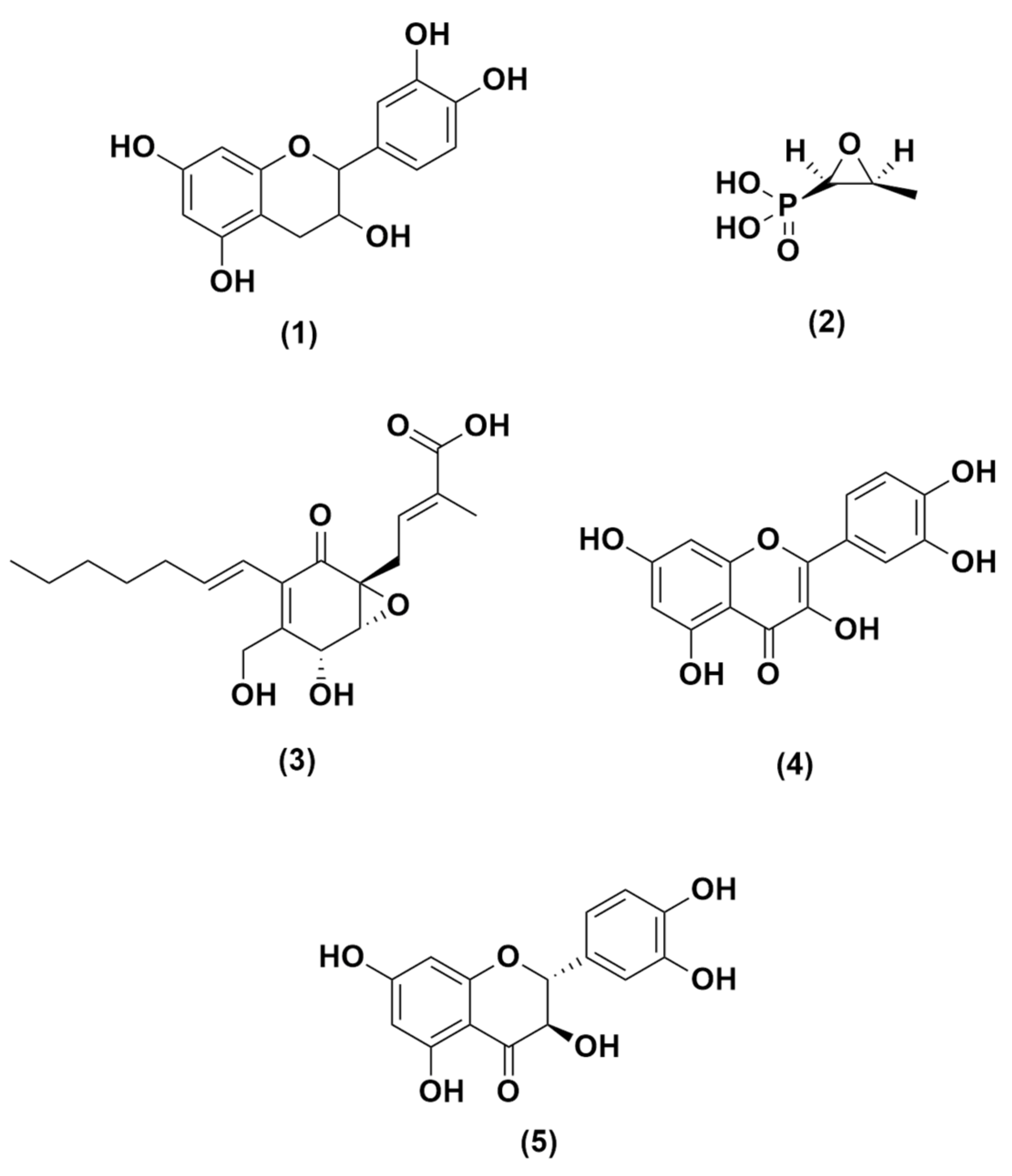
2.2. Structure Determination of Compound
2.3. Antibacterial Activity Assay of Extracts against E. faecalis ATCC 29212
2.4. Determining the Antibacterial Activity (MIC and MBC) of Compound
2.5. Antibacterial Activity Prediction from Uncaria gambir Roxb. through Molecular Docking
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Chemicals
4.2. Instruments
4.3. Materials for Antibacterial Activity Assay: In Vitro Study
4.4. Materials for Molecular Docking: In Silico Study
4.5. Experimental
4.5.1. Preparation of Extracts of Gambir (Uncaria gambir Roxb.) Fruit
4.5.2. Phytochemical Screening
4.5.3. Isolation of Compound from Ethyl Acetate Extract of Uncaria gambir Roxb. Fruit
4.5.4. Structure Determination of Active Compound 1
4.5.5. Evaluation of the Antibacterial Activity of the Extract and Active Compound of Uncaria gambir Roxb.
Microorganism Assay
Antibacterial Activity
4.5.6. In Silico Characterization of the Uncaria gambir Roxb. Compounds
4.5.7. Molecular Docking between MurA/GBAP/Gelatinase/Serine Protease and Uncaria gambir Roxb. Compound
4.5.8. Complex MurA/GBAP/Gelatinase/Serine Protease—Catechin Visualization and Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Araujo, M.W.B.; Charles, C.A.; Weinstein, R.B.; McGuire, J.A.; Parikh-Das, A.M.; Du, Q.; Zhang, J.; Berlin, J.A.; Gnsolley, J.C. Meta-analysis of the effect of an essential oil-containing mouthrinse on gingivitis and plaque. J. Am. Dent. Assoc. 2015, 146, 610–622. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Available online: https://www.who.int/oral_health/disease_burden/global/en/ (accessed on 1 October 2020).
- Cahuana-Vasquez, R.A.; Ferreira dos Santos, S.S.; Koga-Ito, C.Y.; Cardoso Jorge, A.O. Antimicrobial activity of Uncaria tomentosa against oral human pathogens. Braz. Oral Res. 2007, 21, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Camere-Colarossi, R.; Ulloa-Urizar, G.; Medina-Flores, D.; Caballero-Garcia, S.; Mayta-Tovalino, F. Antibacterial activity of Myrciaria dubia (Camu camu) against Streptococcus mutans and Streptococcus sanguinis. Asian Pac. J. Trop. Biomed. 2016, 6, 740–744. [Google Scholar] [CrossRef] [Green Version]
- Benbelaïd, F.; Khadir, A.; Abdoune, M.A.; Bendahou, M.; Muselli, A. Antimicrobial activity of some essential oils against oral multidrug-resistant Enterococcus faecalis in both planktonic and biofilm state. Asian Pac. J. Trop. Biomed. 2014, 4, 463–472. [Google Scholar] [CrossRef] [Green Version]
- Walsh, C.T.; Wencewicz, T.A. Prospects for new antibiotics: A molecule-centered perspective. Antibiotics 2014, 67, 7–22. [Google Scholar] [CrossRef] [PubMed]
- Kurnia, D.; Apriyanti, E.; Soraya, C. Antibacterial flavonoids against oral bacteria of Enterococcus faecalis ATCC 29212 from Sarang Semut (Myrmecodia pendans) and its inhibitor activity against enzyme MurA. Curr. Drug Discov. Technol. 2019, 16, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.A. Structure, function and dynamics in the mur family of bacterial cell wall ligases. J. Mol. Biol. 2006, 4, 640–655. [Google Scholar] [CrossRef]
- Nikolaidis, I.; Dessen, A. Resistance to antibiotics targeted to the bacterial cell wall. Prot. Sci. 2014, 23, 243–259. [Google Scholar] [CrossRef]
- Bassler, B.L.; Losick, R. Bacterially Speaking. Cell 2006, 125, 237–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutherford, S.T.; Bassler, B.L. Bacterial quorum sensing: Its role in virulence and possibilities for its control. CSH Perspect. Med. 2012, 2, a012427. [Google Scholar] [CrossRef]
- Teixeira, N.; Varahan, S.; Gorman, M.J.; Palmer, K.L.; Zaidmann-Remy, A.; Yokohata, R.; Nakayama, J.; Hancock, L.E.; Jacinto, A.; Gilmore, M.S.; et al. Drosophila host model reveals new Enterococcus faecalis quorum-sensing associated virulence factors. PLoS ONE 2013, 8, e6470. [Google Scholar] [CrossRef] [Green Version]
- Qin, X.; Singh, K.V.; Weinstock, G.M.; Murray, B.E. Characterization of fsr, a regulator controlling expression of gelatinase and serine protease in Enterococcus faecalis OG1RF. J. Bacteriol. 2001, 183, 3372–3382. [Google Scholar] [CrossRef] [Green Version]
- Ji, G.; Beavis, R.; Novick, R.P. Bacterial interference caused by autoinducing peptide variants. Science 1997, 276, 2027–2030. [Google Scholar] [CrossRef]
- Gaspar, F.; Teixeira, N.; Rigottier-Gois, L.; Marujo, P.; Nielsen-LeRoux, C.; Crespo, M.T.B.; Lopes, M.d.F.S.; Serror, P. Virulence of Enterococcus faecalis dairy strains in an insect model: The role of fsrB and gelE. Microbiology 2009, 155, 3564–3571. [Google Scholar] [CrossRef] [Green Version]
- Qin, X.; Singh, K.V.; Weinstock, G.M.; Murray, B.E. Effects of Enterococcus faecalis fsr genes on production of gelatinase and a serine protease and virulence. Infect. Immun. 2000, 68, 2579–2586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayama, J.; Chen, S.; Oyama, N.; Nishiguchi, K.; Azab, E.A.; Tanaka, E.; Kariyama, R.; Sonomoto, K. Revised model for Enterococcus faecalis fsr quorum-sensing system: The small open reading frame fsrD encodes the gelatinase biosynthesis-activating pheromone propeptide corresponding to staphylococcal AgrD. J. Bacteriol. 2006, 188, 8321–8326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, L.; Goraya, M.U.; Arafat, Y.; Ajmal, M.; Chen, J.; Yu, D. Molecular mechanism of quorum-sensing in Enterococcus faecalis: Its role in virulence and therapeutic approaches. Int. J. Mol. Sci. 2017, 18, 960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, J.; Xu, X.; Cheng, F.; Liu, H.; Luo, X.; Shen, J.; Chen, K.; Zhao, W.; Shen, X.; Jiang, H. Virtual Screening on natural products for discovering active compounds and target information. Curr. Med. Chem. 2005, 10, 2327–2342. [Google Scholar] [CrossRef] [PubMed]
- Lavecchia, A.; Giovanni, C.D. The biophilic quality index. A tool to improve a building from “Green” to restorative. Curr. Med. Chem. 2013, 20, 2839–2860. [Google Scholar] [CrossRef] [PubMed]
- Heitzman, M.E.; Neto, C.C.; Winiarz, E.; Vaisberg, A.J.; Hammond, G.B. Ethnobotany, phytochemistry and pharmacology of Uncaria (Rubiaceae). Phytochemistry 2005, 66, 5–29. [Google Scholar] [CrossRef]
- Grosvenor, P.W.; Supriono, A.; Gray, D.O. Medicinal plants from Riau Province, Sumatra, Indonesia. Part 2: Antibacterial and antifungal activity. J. Ethnopharmacol. 1995, 45, 97–111. [Google Scholar] [CrossRef]
- Rauf, A.; Rahmawaty; Siregar, A.Z. The Condition of Uncaria gambir Roxb. as one of important medicinal plants in North Sumatra Indonesia. Proc. Chem. 2015, 14, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Apea-Bah, F.B.; Hanafi, M.; Dewi, R.T.; Fajriah, S.; Darwaman, A.; Artanti, N.; Lotulung, P.; Ngadymang, P.; Minarti, B. Assessment of the DPPH and α-glucosidase inhibitory potential of gambier and qualitative identification of major bioactive compound. J. Med. Plants Res. 2009, 3, 736–757. [Google Scholar] [CrossRef]
- Chan, K.C. Gambirdine and isogambirdine, the alkaloids from Uncaria gambir (hunt) Roxb. Tetrahedron Lett. 1968, 9, 3403–3406. [Google Scholar] [CrossRef]
- Rismana, E.; Ningsih, S.; Fachrudin, F. In vitro study of xanthine oxidase inhibitory of Gambir (Uncaria gambir) hunter Roxb. extracts. Pharmacogn. J. 2014, 9, 862–865. [Google Scholar] [CrossRef] [Green Version]
- Merlini, L.; Mondelli, R.; Nasini, G.; Hesse, M. The structure of roxburghines A-E, new indole alkaloids from an Uncaria sp. Tetrahedron 1970, 26, 2259–2279. [Google Scholar] [CrossRef]
- Tsimogiannis, D.; Samiotaki, M.; Panayotou, G.; Oreopoulou, V. Characterization of flavonoid subgroups and hydroxy substitution by HPLC-MS/MS. Molecules 2007, 12, 593–606. [Google Scholar] [CrossRef] [PubMed]
- El-Razek, M.H. NMR Assignments of four catechin epimers. Asian J. Chem. 2007, 19, 4867–4872. [Google Scholar]
- Kumar, A.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, A162750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravipati, A.S.; Reddy, N.; Rao, S. Biologically active compounds from the Genus Uncaria (Rubiaceae). Stud. Nat. Prod. Chem. 2014, 43, 381–408. [Google Scholar] [CrossRef]
- Pan, X.; Chen, F.; Wu, T.; Tang, H.; Zhao, Z. The acid, bile tolerance and antimicrobial property of Lactobacillus acidophilus NIT. Food Control 2009, 20, 598–602. [Google Scholar] [CrossRef]
- Kuete, V. Potential of cameroonian plants and derived products against microbial infections: A review. Plant Med. 2010, 76, 1479–1491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Wass, T.W.P.D.; Astatkie, T.; Rupasinghe, H.P.V. Optimization of catechin and proanthocyanidin recovery from grape seeds using microwave-assisted extraction. Biomolecules 2020, 10, 243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, D.T.; Lavanya, P.; Doss, C.G.P.; Tayubi, I.A.; Kumar, D.R.N.; Yesurajan, I.F.; Siva, R.; Balaji, V. A Molecular docking and dynamics approach to screen potent inhibitors against fosfomycin resistant enzyme in clinical klebsiella pneumoniae. J. Cell Biochem. 2017, 7, 1–7. [Google Scholar] [CrossRef]
- Gupta, A.; Chaudhary, N.; Kakularam, K.R.; Pallu, R.; Polamarasetty, A. The augmenting effects of desolvation and conformational energy terms on the predictions of docking programs against mPGES-1. PLoS ONE. 2015, 1, e0134472. [Google Scholar] [CrossRef] [PubMed]
- Popovic, M.; Steinort, D.; Pillai, S.; Joukhadar, C. Fosfomycin: An old, new friend. Eur. J. Clin. Microbiol. Infect. Dis. 2010, 29, 127–142. [Google Scholar] [CrossRef] [PubMed]
- Kapitan, O.B.; Ambarsari, L.; Falah, S. Inhibition docking Simulation of Zerumbone, Gingerglycolipid B, and Curzerenone compound of Zingiber zerumbet from Timor Island against MurA enzyme. J. Appl. Chem. Sci. 2016, 3, 279–288. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.J.; Zheng, Q.C.; Zhang, J.L.; Chu, W.T.; Cui, Y.L.; Wang, Y.; Zhang, H.X. Fosfomycin induced structural change in fosfomycin resistance kinases FomA: Molecular dynamics and molecular docking studies. J. Mol. Model. 2014, 20, 2236. [Google Scholar] [CrossRef]
- Herrera, D.R.; Tay, L.Y.; Rezende, E.C.; Kozlowski, V.A.; dos Santos, E.B. In vitro antimicrobial activity of phytotherapic Uncaria tomentosa against endodontic pathogens. J. Oral Sci. 2010, 52, 473–476. [Google Scholar] [CrossRef] [Green Version]
- Herli, M.A.; Riau, U.M.; Putra, A.; Andalas, U.; Bakhtiar, A. Molecular docking studies of flavonoid bioactive compound against rna polymerase mycobacterium tuberculosis research journal of pharmaceutical, biological and chemical sciences molecular docking studies of flavonoid bioactive compound against RNA polymerase Mycobacterium tuberculosis. Res. J. Pharm. Biol. Chem. Sci. 2016, 7, 512–523. [Google Scholar]
- Kurnia, D.; Hutabarat, G.S.; Windaryanti, D.; Herlina, T.; Herdiyati, Y.; Satari, M.H. Potential allylpyrocatechol derivatives as antibacterial agent against oral pathogen of S. sanguinis ATCC 10,556 and inhibitor of MurA enzymes: In vitro and in silico study. Dovepress 2020, 14, 2977–2985. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Z.; Wu, M.; Shang, Y.; Jiang, M.; Liu, J.; Qiao, C.; Ye, H.; Lin, Y.; Piao, M.; Sun, R.; et al. Taxifolin ameliorates high-fat diet feeding plus acute ethanol binge-induced steatohepatitis through inhibiting inflammatory caspase-1-dependent pyroptosis. Food Funct. 2020, 12, 362–372. [Google Scholar] [CrossRef]
- Starmans, D.A.J.; Nijhuis, H. Extraction of secondary metabolites from plant material: A review. Trend Food Sci. Technol. 1996, 7, 191–197. [Google Scholar] [CrossRef]
- Fekete, T.; Tumah, H.; Woodwell, J.; Truant, A.; Satischandran, V.; Axelrod, B.; Kreter, B. A comparison of serial plate agar dilution, bauer-kirbydisk diffusion, and the vitek automicrobic system for the determination of susceptibilities of Klebsiella spp., Enterobacter spp., and Pseudomonas aeruginosa to ten antimicrobial agent. Diagn. Microbiol. 1994, 18, 251–258. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute Document M7–A8. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 9th ed.; Approved Standard; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012. [Google Scholar]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An open chemical toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef] [Green Version]
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Cassarino, T.G.; Bertoni, M.; Bordoli, L.; et al. SWISS-MODEL: Modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res. 2014, 42, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Suprijono, M.M.; Widjanarko, S.B.; Sujuti, H.; Kurnia, D. Computational study of antioxidant activity and bioavailability of Papua red fruit (Pandanus conoideus Lam.) flavonoids through docking toward human serum albumin. AIP Conf. Proc. 2018, 2018, 020020. [Google Scholar] [CrossRef]
- Rauf, M.A.; Zubair, S.; Azhar, A. Ligand docking and binding site analysis with PyMOL and Autodock/Vina. Int. J. Basic Appl. Sci. 2015, 4, 168–177. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Extracts | Concentration (%) | Inhibition Zone (mm) |
|---|---|---|---|
| 1 | n-Hexane | 10 | 0.00 |
| 2 | Ethyl acetate | 8.30 | |
| 3 | Methanol | 7.65 | |
| 4 | Water | 6.75 | |
| 5 | Chlorhexidine | 2 | 16.40 |
| No. | Compounds | Concentrations (%) | Inhibition Zones (mm) |
|---|---|---|---|
| 1 | Catechin (1) | 10 | 11.7 |
| 2 | Chlorhexidine | 2 | 26.5 |
| Compounds | Concentrations (%) | |
|---|---|---|
| MIC | MBC | |
| Catechin | 0.625 | 1.25 |
| Chlorhexidine | 3.12 | 6.25 |
| Fosfomycin | 62.5 | None |
| Ligand | Binding Affinity of Ligand-Protein Complex (kcal/mol) | |||
|---|---|---|---|---|
| MurA | GBAP | Gelatinase | Serine Protease | |
| Catechin | −8.5 | −5.2 | −7.8 | −7.0 |
| Fosfomycin | −4.6 | −3.1 | −4.6 | −3.8 |
| Ambuic acid | −7.8 | −4.5 | −6.6 | −6.5 |
| Quercetin | −8.5 | −5.2 | −8.3 | −6.9 |
| Taxifolin | −9.1 | −5.1 | −7.9 | −6.5 |
| Residues Binding at Ligand-Protein Complex | ||||
|---|---|---|---|---|
| Ligand | MurA | GBAP | Gelatinase | Serine Protease |
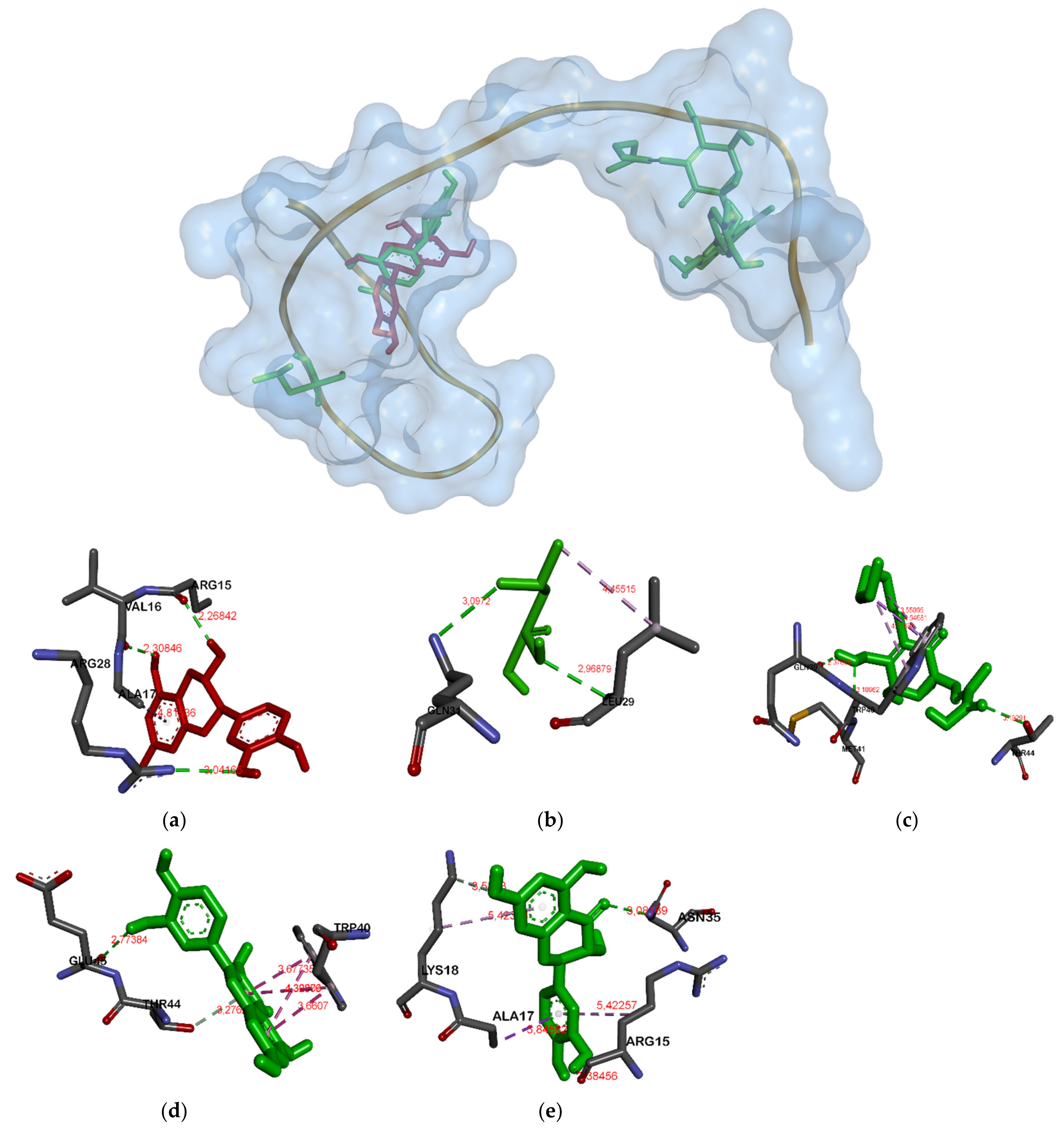
| Catechin | Ser162A, Gly164A, Asp305A, Ala297A | Arg15A, Ser33A, Val16A | His332A, Tyr343A, Glu329A, Asn298A | Asn145A, Asn262A, Asn268A, Leu212A |
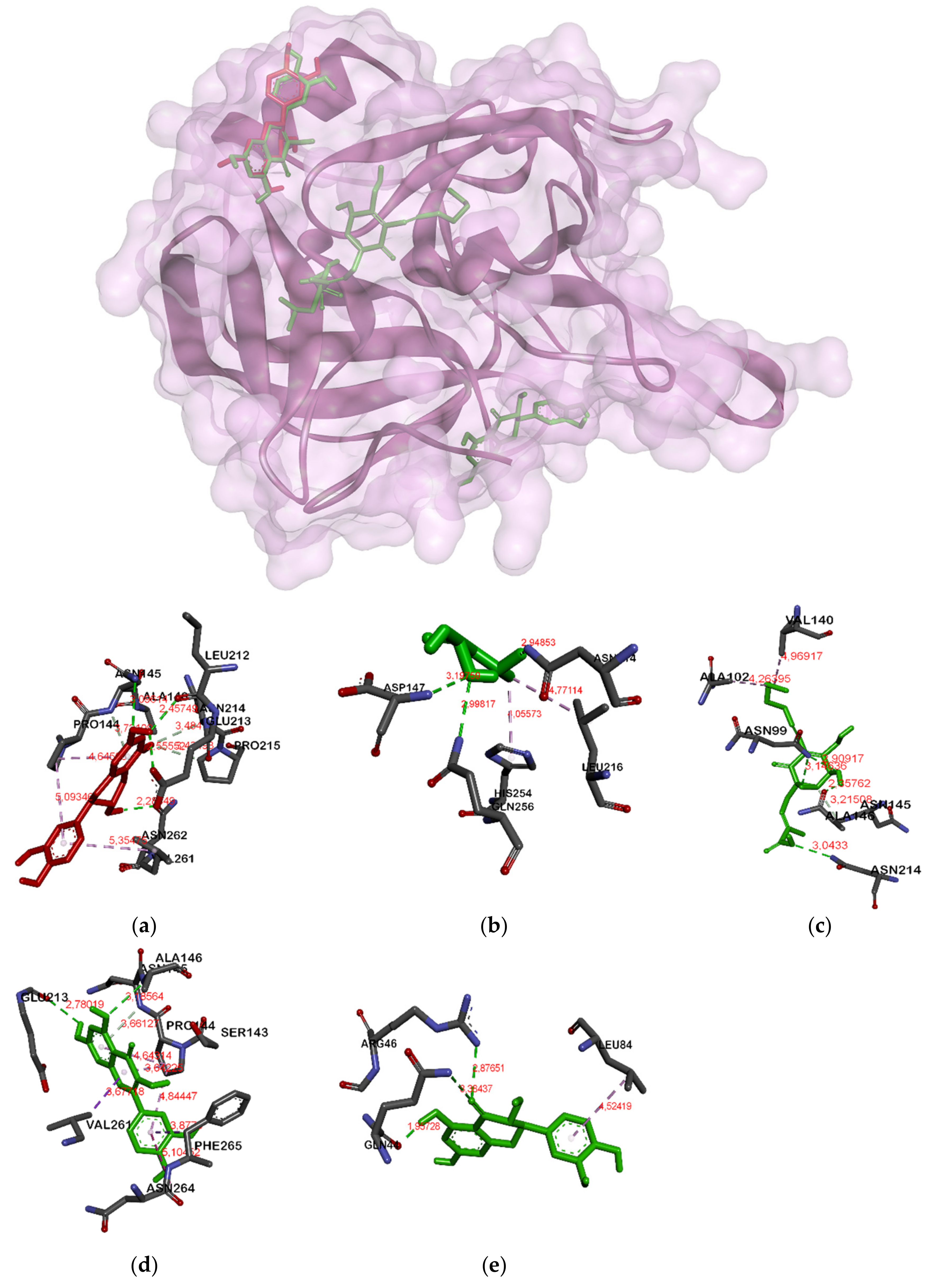
| Fosfomycin | Arg120A, Asn23A, Arg371A, Asp305A | Arg28A, Gln31A, Lys26A | Arg384A, Glu329A, Asn298A, His419A | Asn214A, Asp147A |
| Ambuic acid | Lys22A, Arg91A, Arg120A | Ile36A | Arg384A, Glu352A | Asn99A, Ala146A, Ser143A, Asn214A, Asp147A |
| Quercetin | Ser12A, Gly164A, Glu188A, Phe328A | Glu45A, Gln43A, Trp40A | Asn298A, His332A, Glu336A, Tyr343A, Glu329A | Asn145A, Asn262A, Asn268A, Glu213A |
| Taxifolin | Ser162A, Gly164A, Asp305A, Asn23A, Arg397A, Asp49A, Lys22A | Asn35A, Arg15A, Ser33A, Gln31A | Asn351A, His332A, Asn298A, Glu329A, Trp301A | Arg46A, Ser83A, His195A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurnia, D.; Ramadhanty, Z.F.; Ardani, A.M.; Zainuddin, A.; Dharsono, H.D.A.; Satari, M.H. Bio-Mechanism of Catechin as Pheromone Signal Inhibitor: Prediction of Antibacterial Agent Action Mode by In Vitro and In Silico Study. Molecules 2021, 26, 6381. https://doi.org/10.3390/molecules26216381
Kurnia D, Ramadhanty ZF, Ardani AM, Zainuddin A, Dharsono HDA, Satari MH. Bio-Mechanism of Catechin as Pheromone Signal Inhibitor: Prediction of Antibacterial Agent Action Mode by In Vitro and In Silico Study. Molecules. 2021; 26(21):6381. https://doi.org/10.3390/molecules26216381
Chicago/Turabian StyleKurnia, Dikdik, Zenika Febian Ramadhanty, Aprilina Mora Ardani, Achmad Zainuddin, Hendra Dian Adhita Dharsono, and Mieke Hemiawati Satari. 2021. "Bio-Mechanism of Catechin as Pheromone Signal Inhibitor: Prediction of Antibacterial Agent Action Mode by In Vitro and In Silico Study" Molecules 26, no. 21: 6381. https://doi.org/10.3390/molecules26216381
APA StyleKurnia, D., Ramadhanty, Z. F., Ardani, A. M., Zainuddin, A., Dharsono, H. D. A., & Satari, M. H. (2021). Bio-Mechanism of Catechin as Pheromone Signal Inhibitor: Prediction of Antibacterial Agent Action Mode by In Vitro and In Silico Study. Molecules, 26(21), 6381. https://doi.org/10.3390/molecules26216381