
An Optimized SPME-GC-MS Method for Volatile Metabolite Profiling of Different Alfalfa (Medicago sativa L.) Tissues



Abstract
:
1. Introduction
2. Results and Discussion
2.1. SPME Extraction Parameters
2.2. Fiber Composition
2.3. Extraction Temperature
2.4. Extraction Time
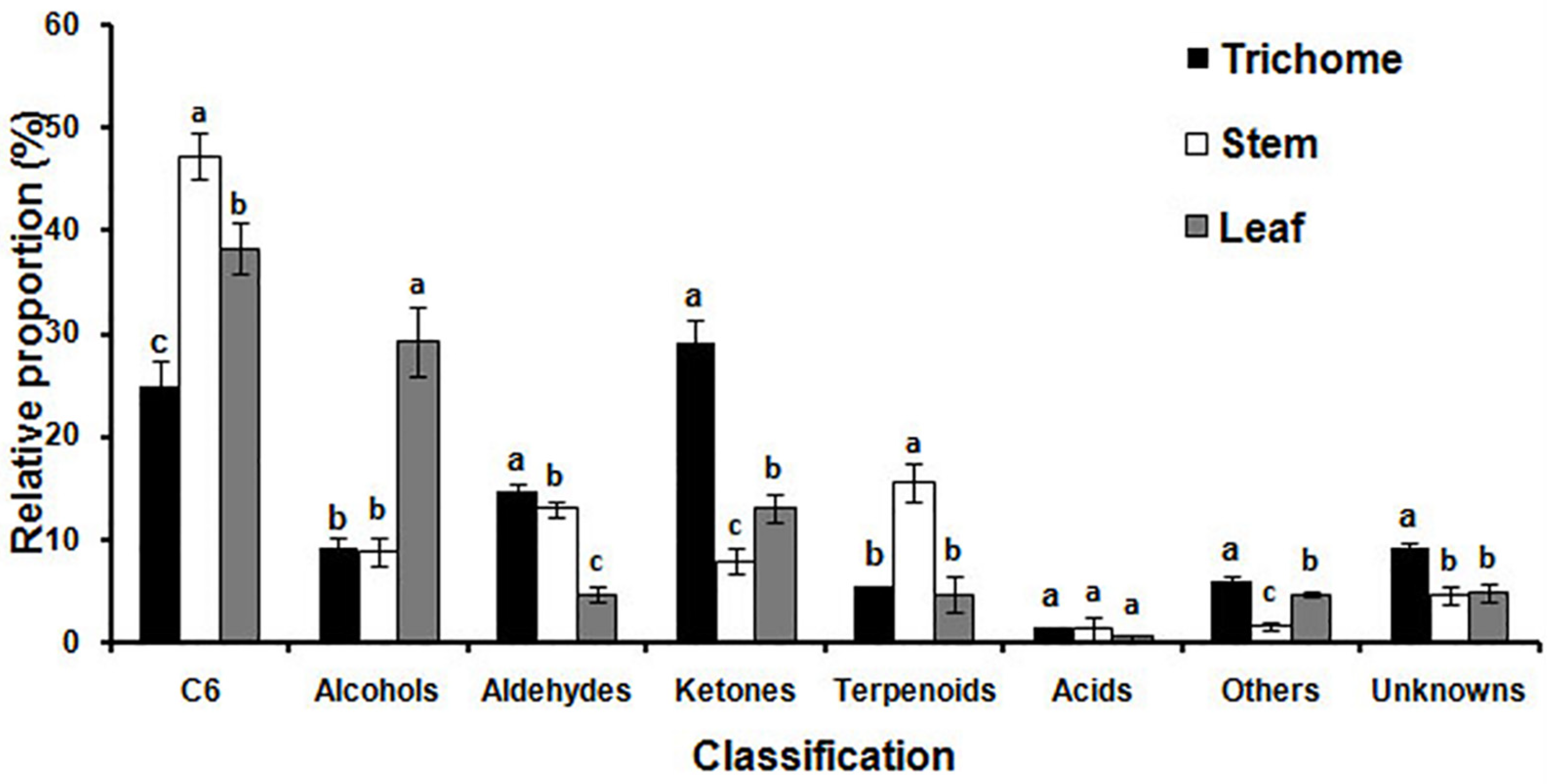
2.5. Large-Scale Profiling of Alfalfa Volatile Metabolites
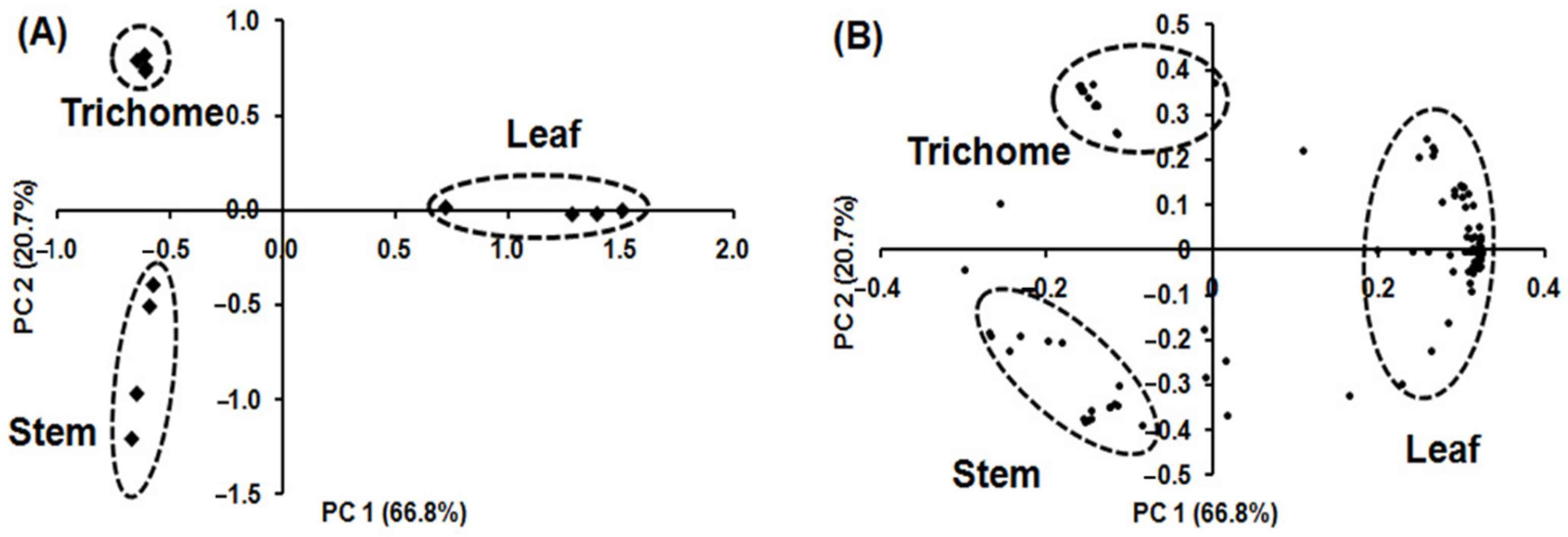
2.6. Multivariate Analysis
3. Materials and Methods
3.1. Chemicals
3.2. Plant Materials
3.3. SPME Procedure Optimization
3.4. Sample Preparation for Volatile Metabolic Profiling
3.5. GC-MS Analyses
3.6. Data Processing, Metabolite Identification and Semi-Quantification
3.7. Statistical Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- He, X.; Majid, B.; Zhang, H.; Liu, W.; Limmer, M.A.; Burken, J.G.; Shi, H. Green Analysis: Rapid-Throughput Analysis of Volatile Contaminants in Plants by Freeze-Thaw-Equilibration Sample Preparation and SPME-GC-MS Analysis. J. Agric. Food Chem. 2021, 69, 5428–5434. [Google Scholar] [CrossRef]
- Cincotta, F.; Verzera, A.; Tripodi, G.; Condurso, C. Volatile emerging contaminants in melon fruits, analysed by HS-SPME-GC-MS. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2018, 35, 512–518. [Google Scholar] [CrossRef]
- Li, C.; Xin, M.; Li, L.; He, X.; Yi, P.; Tang, Y.; Li, J.; Zheng, F.; Liu, G.; Sheng, J.; et al. Characterization of the aromatic profile of purple passion fruit (Passiflora edulis Sims) during ripening by HS-SPME-GC/MS and RNA sequencing. Food Chem. 2021, 355, 129685. [Google Scholar] [CrossRef] [PubMed]
- Charry-Parra, G.; Dejesus-Echevarria, M.; Perez, F.J. Beer volatile analysis: Optimization of HS/SPME coupled to GC/MS/FID. J. Food Sci. 2011, 76, C205–C211. [Google Scholar] [CrossRef] [PubMed]
- da Silva, G.C.; da Silva, A.A.; da Silva, L.S.; Godoy, R.L.; Nogueira, L.C.; Quiterio, S.L.; Raices, R.S. Method development by GC-ECD and HS-SPME-GC-MS for beer volatile analysis. Food Chem. 2015, 167, 71–77. [Google Scholar] [CrossRef]
- Hernandes, K.C.; Souza-Silva, E.A.; Assumpcao, C.F.; Zini, C.A.; Welke, J.E. Validation of an analytical method using HS-SPME-GC/MS-SIM to assess the exposure risk to carbonyl compounds and furan derivatives through beer consumption. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2019, 36, 1808–1821. [Google Scholar] [CrossRef]
- Bueno, M.; Resconi, V.C.; Campo, M.M.; Ferreira, V.; Escudero, A. Development of a robust HS-SPME-GC-MS method for the analysis of solid food samples. Analysis of volatile compounds in fresh raw beef of differing lipid oxidation degrees. Food Chem. 2019, 281, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Pavlidis, D.E.; Mallouchos, A.; Ercolini, D.; Panagou, E.Z.; Nychas, G.E. A volatilomics approach for off-line discrimination of minced beef and pork meat and their admixture using HS-SPME GC/MS in tandem with multivariate data analysis. Meat Sci. 2019, 151, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Shockley, F.W.; Backus, E.A.; Ellersieck, M.R.; Johnson, D.W.; McCaslin, M. Glandular-haired alfalfa resistance to potato leafhopper (Homoptera: Cicadellidae) and hopperburn: Development of resistance indices. J. Econ. Entomol. 2002, 95, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Sulc, R.M.; van Santen, E.; Johnson, K.D.; Sheaffer, C.C.; Undersander, D.J.; Bledsoe, L.W.; Hogg, D.B.; Willson, H.R. Glandular-haired cultivars reduce potato leafhopper damage in alfalfa. Agron. J. 2001, 93, 1287–1296. [Google Scholar] [CrossRef]
- Schwab, W.; Davidovich-Rikanati, R.; Lewinsohn, E. Biosynthesis of plant-derived flavor compounds. Plant. J. 2008, 54, 712–732. [Google Scholar] [CrossRef]
- Wang, F.; Park, Y.L.; Gutensohn, M. Glandular Trichome-Derived Mono- and Sesquiterpenes of Tomato Have Contrasting Roles in the Interaction with the Potato Aphid Macrosiphum euphorbiae. J. Chem. Ecol. 2021, 47, 204–214. [Google Scholar] [CrossRef]
- Elden, T.C.; McCaslin, M. Potato leafhopper (Homoptera: Cicadellidae) resistance in perennial glandular-haired alfalfa clones. J. Econ. Entomol. 1997, 90, 842–847. [Google Scholar] [CrossRef]
- Ranger, C.M.; Hower, A.A. Role of the glandular trichomes in resistance of perennial alfalfa to the potato leafhopper (Homoptera: Cicadellidae). J. Econ. Entomol. 2001, 94, 950–957. [Google Scholar] [CrossRef]
- Ranger, C.M.; Winter, R.E.K.; Backus, E.A.; Rottinghaus, G.E.; Ellersieck, M.R.; Johnson, D.W. Discrimination by the potato leafhopper (Hemiptera: Cicadellidae) of host volatiles from resistant and susceptible alfalfa, Medicago sativa L. Environ. Entomol. 2005, 34, 271–280. [Google Scholar] [CrossRef]
- Tava, A.; Pecetti, L. Volatiles from Medicago sativa complex flowers. Phytochemistry 1997, 45, 1145–1148. [Google Scholar] [CrossRef]
- Batlle, R.; Sanchez, C.; Nerin, C. A systematic approach to optimize solid-phase microextraction. Determination of pesticides in ethanol water mixtures used as food simulants. Anal. Chem. 1999, 71, 2417–2422. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.; Perestrelo, R.; Camara, J.S. Comparative analysis of the volatile fraction from Annona cherimola Mill. cultivars by solid-phase microextraction and gas chromatography-quadrupole mass spectrometry detection. Talanta 2009, 77, 1087–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García, Y.M.; Rufini, J.C.M.; Campos, M.P.; Guedes, M.N.S.; Augusti, R.; Melo, J.O.F. SPME Fiber Evaluation for Volatile Organic Compounds Extraction from Acerola. J. Braz. Chem. Soc. 2019, 30, 8. [Google Scholar] [CrossRef]
- Wei, S.; Xiao, X.; Wei, L.; Li, L.; Li, G.; Liu, F.; Xie, J.; Yu, J.; Zhong, Y. Development and comprehensive HS-SPME/GC-MS analysis optimization, comparison, and evaluation of different cabbage cultivars (Brassica oleracea L. var. capitata L.) volatile components. Food Chem. 2021, 340, 128166. [Google Scholar] [CrossRef]
- Hernandez, F.; Portoles, T.; Pitarch, E.; Lopez, F.J. Target and nontarget screening of organic micropollutants in water by solid-phase microextraction combined with gas chromatography/high-resolution time-of-flight mass spectrometry. Anal. Chem. 2007, 79, 9494–9504. [Google Scholar] [CrossRef]
- Lee, J.H.; Diono, R.; Kim, G.Y.; Min, D.B. Optimization of solid phase microextraction analysis for the headspace volatile compounds of Parmesan cheese. J. Agric. Food Chem. 2003, 51, 1136–1140. [Google Scholar] [CrossRef]
- Zhang, Y.; Gao, B.; Zhang, M.W.; Shi, J.; Xu, Y.J. Headspace solid-phase microextraction-gas chromatography-mass spectrometry analysis of the volatile components of longan (Dimocarpus longan Lour.). Eur. Food Res. Technol. 2009, 229, 457–465. [Google Scholar] [CrossRef]
- Ho, C.W.; Aida, W.M.W.; Maskat, M.Y.; Osman, H. Optimization of headspace solid phase microextraction (HS-SPME) for gas chromatography mass spectrometry (GC-MS) analysis of aroma compound in palm sugar (Arenga pinnata). J. Food Compos. Anal. 2006, 19, 822–830. [Google Scholar] [CrossRef]
- Mwatseteza, J.F.; Torto, N. Profiling volatile compounds from Mucuna beans by solid phase microextraction and gas chromatography-high resolution time of flight mass spectrometry. Food Chem. 2010, 119, 386–390. [Google Scholar] [CrossRef]
- Sousa, E.T.; Rodrigues, F.D.; Martins, C.C.; de Oliveira, F.S.; Pereira, P.A.D.; de Andrade, J.B. Multivariate optimization and HS-SPME/GC-MS analysis of VOCs in red, yellow and purple varieties of Capsicum chinense sp. peppers. Microchem. J. 2006, 82, 142–149. [Google Scholar] [CrossRef]
- Bohlmann, J.; Keeling, C.I. Terpenoid biomaterials. Plant J. 2008, 54, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Schilmiller, A.L.; Last, R.L.; Pichersky, E. Harnessing plant trichome biochemistry for the production of useful compounds. Plant J. 2008, 54, 702–711. [Google Scholar] [CrossRef] [Green Version]
- Cuevas-Glory, L.F.; Pino, J.A.; Santiago, L.S.; Sauri-Duch, E. A review of volatile analytical methods for determining the botanical origin of honey. Food Chem. 2007, 103, 1032–1043. [Google Scholar] [CrossRef]
- Stein, S.E. An integrated method for spectrum extraction and compound identification from GCMS data. J. Am. Soc. Mass Spectrom. 1999, 10, 770–781. [Google Scholar] [CrossRef] [Green Version]
- Sumner, L.; Amberg, A.; Barrett, D.; Beale, M.; Beger, R.; Daykin, C.; Fan, T.; Fiehn, O.; Goodacre, R.; Griffin, J.; et al. Proposed minimum reporting standards for chemical analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broeckling, C.D.; Reddy, I.R.; Duran, A.L.; Zhao, X.C.; Sumner, L.W. MET-IDEA: Data extraction tool for mass spectrometry-based metabolomics. Anal. Chem. 2006, 78, 4334–4341. [Google Scholar] [CrossRef]
- Lei, Z.; Li, H.; Chang, J.; Zhao, P.; Sumner, L.W. MET-IDEA version 2.06: Improved efficiency and additional functions for mass spectrometry-based metabolomics data processing. Metabolomics 2012, 8, 105–110. [Google Scholar] [CrossRef]
- Holopainen, J.K. Multiple functions of inducible plant volatiles. Trends Plant Sci. 2004, 9, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Dudareva, N.; Negre, F.; Nagegowda, D.A.; Orlova, I. Plant volatiles: Recent advances and future perspectives. Crit. Rev. Plant Sci. 2006, 25, 417–440. [Google Scholar] [CrossRef]
- Farag, M.A.; Pare, P.W. C-6-green leaf volatiles trigger local and systemic VOC emissions in tomato. Phytochemistry 2002, 61, 545–554. [Google Scholar] [CrossRef]
- Kishimoto, K.; Matsui, K.; Wawa, R.; Takabayashi, J. Components of C6-aldehyde-induced resistance in Arabidopsis thaliana against a necrotrophic fungal pathogen, Botrytis cinerea. Plant Sci. 2006, 170, 715–723. [Google Scholar] [CrossRef]
- Mirabella, R.; Rauwerda, H.; Struys, E.A.; Jakobs, C.; Triantaphylides, C.; Haring, M.A.; Schuurink, R.C. The Arabidopsis her1 mutant implicates GABA in E-2-hexenal responsiveness. Plant J. 2008, 53, 197–213. [Google Scholar] [CrossRef]
- D’Auria, J.C.; Pichersky, E.; Schaub, A.; Hansel, A.; Gershenzon, J. Characterization of a BAHD acyltransferase responsible for producing the green leaf volatile (Z)-3-hexen-1-yl acetate in Arabidopsis thaliana. Plant J. 2007, 49, 194–207. [Google Scholar] [CrossRef] [Green Version]
- Khattree, R.; Naik, D.N. Multivariate Data Reduction and Discrimination with SAS® Software; SAS Institute Inc.: Cary, NC, USA, 2000. [Google Scholar]
- Gaquerel, E.; Weinhold, A.; Baldwin, I.T. Molecular interactions between the specialist herbivore Manduca sexta (Lepidoptera, Sphingidae) and its natural host Nicotiana attenuata. VIII. An unbiased GCxGC-ToFMS analysis of the plant’s elicited volatile emissions. Plant Physiol. 2009, 149, 1408–1423. [Google Scholar] [CrossRef] [Green Version]
- Tikunov, Y.; Lommen, A.; de Vos, C.H.R.; Verhoeven, H.A.; Bino, R.J.; Hall, R.D.; Bovy, A.G. A novel approach for nontargeted data analysis for metabolomics. Large-scale profiling of tomato fruit volatiles. Plant Physiol. 2005, 139, 1125–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plotto, A.; McDaniel, M.R.; Mattheis, J.P. Characterization of changes in ‘Gala’ apple aroma during storage using osme analysis, a gas chromatography-olfactometry technique. J. Am. Soc. Hort. Sci. 2000, 125, 714–722. [Google Scholar] [CrossRef] [Green Version]
- Genovese, A.; Ugliano, M.; Pessina, R.; Gambuti, A.; Piombino, P.; Moio, L. Comparison of the aroma compounds in apricot (Prunus armeniaca, L. cv Pellecchiella) and apple (Malus pumila, L. cv. Annurca) raw distillates. Ital. J. Food Sci. 2004, 16, 185–196. [Google Scholar]
- Frost, C.J.; Mescher, M.C.; Dervinis, C.; Davis, J.M.; Carlson, J.E.; De Moraes, C.M. Priming defense genes and metabolites in hybrid poplar by the green leaf volatile cis-3-hexenyl acetate. New Phytol. 2008, 180, 722–733. [Google Scholar] [CrossRef] [PubMed]
- Adams, A.; Bouckaert, C.; Van Lancker, F.; De Meulenaer, B.; De Kimpe, N. Amino acid catalysis of 2-alkylfuran formation from lipid oxidation-derived α,β-unsaturated aldehydes. J. Agric. Food Chem. 2011, 59, 11058–11062. [Google Scholar] [CrossRef]
- Jimenez, E.; Lanza, B.; Antinolo, M.; Albaladejo, J. Photooxidation of leaf-wound oxygenated compounds, 1-penten-3-ol, (Z)-3-hexen-1-ol, and 1-penten-3-one, initiated by OH radicals and sunlight. Environ. Sci. Technol. 2009, 43, 1831–1837. [Google Scholar] [CrossRef]
- Core, R.J.; Henning, J.A.; Gardea-Torresdey, J. Volatile Compounds from Medicago Spp. as Potential Signals for Alfalfa Weevil Response. J. Agric. Food Chem. 1994, 42, 2932–2936. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RT a | RI b | Compound | PDMS | CAR/PDMS | DVB/CAR/PDMS |
|---|---|---|---|---|---|
| Alcohols | |||||
| 7.27 | 680 | 1-Penten-3-ol | 0.39 ± 0.05 c | N.D. | 6.09 ± 1.14 |
| 8.57 | 725 | 3-Methyl-1-butanol | N.D. | 1.12 ± 0.06 | N.D. |
| 8.84 | 734 | 2-Pentyn-1-ol | N.D. | 0.85 ± 0.18 | 0.22 ± 0.03 |
| 9.53 | 752 | (Z)-2-Penten-1-ol | N.D. | 8.26 ± 0.14 | 1.56 ± 0.63 |
| 12.02 | 827 | 2-Hexyn-1-ol | 0.49 ± 0.06 | N.D. | 7.51 ± 0.79 |
| 16.59 | 981 | 1-Octen-3-ol | 55.85 ± 9.49 | 386.22 ± 13.22 | 335.81 ± 26.62 |
| 19.43 | 1068 | (E)-2-Octen-1-ol | N.D. | 11.42 ± 8.32 | N.D. |
| Aldehydes | |||||
| 4.75 | 2-Propenal | N.D. | 0.45 ± 0.04 | 1.10 ± 0.19 | |
| 5.71 | 595 | Butanal | 0.04 ± 0.00 | 0.34 ± 0.11 | 0.28 ± 0.06 |
| 6.66 | 645 | 2-Butenal | N.D. | 0.92 ± 0.06 | 0.47 ± 0.01 |
| 7.62 | 700 | Pentanal | N.D. | N.D. | 0.26 ± 0.04 |
| 9.14 | 742 | 2-Pentenal | 0.19 ± 0.06 | 20.29 ± 4.31 | 8.31 ± 1.68 |
| 10.48 | 777 | 3-Hexenal | 3.14 ± 0.93 | 275.96 ± 42.84 | 270.77 ± 31.30 |
| 10.60 | 779 | Hexanal | 3.68 ± 0.86 | 192.41 ± 3.52 | 12.13 ± 1.27 |
| 12.40 | 837 | (E)-2-Hexenal | 25.72 ± 7.04 | 975.18 ± 40.25 | 332.79 ± 10.45 |
| 13.86 | 896 | Heptanal | N.D. | 1.13 ± 0.02 | 0.22 ± 0.11 |
| 14.14 | 908 | (E,E)-2,4-Hexadienal | N.D. | 48.24 ± 0.89 | 11.00 ± 1.45 |
| 16.08 | 967 | Benzaldehyde | N.D. | 0.65 ± 0.10 | 0.49 ± 0.05 |
| 17.64 | 1013 | (E,E)-2,4-Heptadienal | 0.09 ± 0.01 | 2.53 ± 0.26 | 1.32 ± 0.14 |
| 18.80 | 1049 | Benzeneacetaldehyde | N.D. | 0.77 ± 0.18 | 0.66 ± 0.06 |
| 20.62 | 1106 | Nonanal | 0.12 ± 0.02 | 0.42 ± 0.03 | 0.63 ± 0.04 |
| Ketones | |||||
| 4.78 | 2-Propanone | 0.22 ± 0.08 | 1.08 ± 0.08 | 5.60 ± 0.82 | |
| 5.65 | 587 | 2,3-Butanedione | N.D. | 0.66 ± 0.16 | 0.65 ± 0.10 |
| 7.32 | 683 | 1-Penten-3-one | 0.82 ± 0.04 | 85.48 ± 10.26 | 28.95 ± 6.68 |
| 7.55 | 696 | 3-Pentanone | N.D. | 128.27 ± 20.89 | 2.02 ± 0.44 |
| 16.42 | 978 | 1-Octen-3-one | 3.02 ± 1.22 | 84.21 ± 38.37 | 87.53 ± 10.67 |
| 20.29 | 3,5-Octadien-2-one | N.D. | 0.23 ± 0.05 | N.D. | |
| Terpenoids | |||||
| 5.02 | 508 | Isoprene | N.D. | 0.65 ± 0.15 | 0.34 ± 0.04 |
| 18.31 | 1034 | (+)-(R)-Limonene | N.D. | N.D. | 0.34 ± 0.01 |
| 19.66 | 1075 | cis-Linalool oxide | N.D. | N.D. | 0.47 ± 0.07 |
| 20.48 | 1100 | β-Linalool | N.D. | N.D. | 0.51 ± 0.01 |
| 3.22 | 1189 | p-Menth-1-en-4-ol | N.D. | N.D. | 0.66 ± 0.06 |
| 23.62 | 1203 | α-Terpinol | N.D. | N.D. | 0.13 ± 0.01 |
| 24.38 | 1229 | β-Cyclocitral | 0.13 ± 0.05 | 0.43 ± 0.07 | 0.20 ± 0.03 |
| 28.83 | 1388 | β-Damascenone | 0.11 ± 0.03 | N.D. | 0.29 ± 0.01 |
| Others | |||||
| 5.82 | 598 | 2-Methylfuran | N.D. | 0.45 ± 0.25 | N.D. |
| 7.66 | 701 | 2-Ethylfuran | 1.00 ± 0.44 | 109.48 ± 8.14 | 6.52 ± 0.59 |
| 12.76 | 893 | 3-Ethylthiophene | 0.28 ± 0.09 | 199.94 ± 25.38 | 4.12 ± 0.54 |
| 15.91 | 960 | 5-Ethyl-2(5H)-furanone | N.D. | 34.15 ± 6.35 | 25.34 ± 4.38 |
| 17.31 | 1003 | 3-Hexen-1-ol acetate | 0.36 ± 0.01 | 3.67 ± 0.15 | 1.96 ± 0.20 |
| 24.19 | 1222 | Ionene | N.D. | N.D. | 0.09 ± 0.01 |
| 31.42 | 1488 | β-Ionone | 1.51 ± 0.47 | 0.78 ± 0.17 | 1.14 ± 0.16 |
| Total known metabolites | 19 | 32 | 38 | ||
| Total volatile metabolites | 48 ± 3 | 99 ± 8 | 97 ± 5 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, D.-S.; Lei, Z.; Bedair, M.; Sumner, L.W. An Optimized SPME-GC-MS Method for Volatile Metabolite Profiling of Different Alfalfa (Medicago sativa L.) Tissues. Molecules 2021, 26, 6473. https://doi.org/10.3390/molecules26216473
Yang D-S, Lei Z, Bedair M, Sumner LW. An Optimized SPME-GC-MS Method for Volatile Metabolite Profiling of Different Alfalfa (Medicago sativa L.) Tissues. Molecules. 2021; 26(21):6473. https://doi.org/10.3390/molecules26216473
Chicago/Turabian StyleYang, Dong-Sik, Zhentian Lei, Mohamed Bedair, and Lloyd W. Sumner. 2021. "An Optimized SPME-GC-MS Method for Volatile Metabolite Profiling of Different Alfalfa (Medicago sativa L.) Tissues" Molecules 26, no. 21: 6473. https://doi.org/10.3390/molecules26216473
APA StyleYang, D. -S., Lei, Z., Bedair, M., & Sumner, L. W. (2021). An Optimized SPME-GC-MS Method for Volatile Metabolite Profiling of Different Alfalfa (Medicago sativa L.) Tissues. Molecules, 26(21), 6473. https://doi.org/10.3390/molecules26216473