
Proteomic Research on the Antitumor Properties of Medicinal Mushrooms



Abstract
:1. Introduction
2. Genus Ganoderma
2.1. Ganoderma spp. Polysaccharides and Polysaccharopeptides
2.2. Ganoderma lucidum Immunomodulatory Proteins
2.3. Ganoderma spp. Triterpenes
3. Lentinus edodes
3.1. Various Lentinus edodes Polysaccharides
3.2. Lentinan
4. Genus Cordyceps
5. Genus Pleurotus
6. Trametes versicolor
7. Hericium erinaceus
8. Phellinus linteus
9. Various Mushroom Genera
9.1. Grifola frondosa
9.2. Antrodia cinnamomea
9.3. Auricularia auricula
9.4. Sporisorium reilianum
9.5. Agrocybe aegerita
9.6. Nectria haematococca
9.7. Agaricus bisporus
10. Ergosterol
11. Medicinal Mushroom Mixtures
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Mattiuzzi, C.; Lippi, G. Current Cancer Epidemiology. J. Epidemiol. Glob. Health 2019, 9, 217–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, J.B. Mechanism-based target identification and drug discovery in cancer research. Science 2000, 287, 1969–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thatishetty, A.V.; Agresti, N.; O’Brien, C.B. Chemotherapy-induced hepatotoxicity. Clin. Liver Dis. 2013, 17, 671. [Google Scholar] [CrossRef]
- Naidu, M.U.; Ramana, G.V.; Rani, P.U.; Mohan, I.K.; Suman, A.; Roy, P. Chemotherapy-induced and/or radiation therapy-induced oral mucositis--complicating the treatment of cancer. Neoplasia 2004, 6, 423–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nurgali, K.; Jagoe, R.T.; Abalo, R. Editorial: Adverse Effects of Cancer Chemotherapy: Anything New to Improve Tolerance and Reduce Sequelae? Front. Pharmacol. 2018, 9, 245. [Google Scholar] [CrossRef]
- Vega-Stromberg, T. Chemotherapy-induced secondary malignancies. J. Infus. Nurs. Off. Publ. Infus. Nurses Soc. 2003, 26, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.T. Global impact of edible and medicinal mushrooms on human welfare in the 21st century: Nongreen revolution. Int. J. Med. Mushr. 1999, 1, 1–8. [Google Scholar] [CrossRef]
- Aras, A.; Khalid, S.; Jabeen, S.; Farooqi, A.A.; Xu, B. Regulation of cancer cell signaling pathways by mushrooms and their bioactive molecules: Overview of the journey from benchtop to clinical trials. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2018, 119, 206–214. [Google Scholar] [CrossRef]
- Badalyan, S.M.; Barkhudaryan, A.; Rapior, S. Recent Progress in Research on the Pharmacological Potential of Mushrooms and Prospects for Their Clinical Application. In Medicinal Mushrooms; Agrawal, D., Dhanasekaran, M., Eds.; Springer: Singapore, 2019. [Google Scholar] [CrossRef]
- Zaidman, B.Y.; Yassin, M.; Mahajna, J.; Wasser, S.P. Medicinal mushroom modulators of molecular targets as cancer therapeutics. Appl. Microbiol. Biotechnol. 2005, 67, 453–468. [Google Scholar] [CrossRef]
- Coussens, L.M.; Fingleton, B.; Matrisian, L.M. Matrix metalloproteinase inhibitors and cancer: Trials and tribulations. Science 2002, 295, 2387–2392. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.Y.; Yang, F.Q.; Gao, J.L. Differential proteomics for studying action mechanisms of traditional Chinese medicines. Chin. Med. 2019, 14, 1. [Google Scholar] [CrossRef] [Green Version]
- Srinivas, P.R.; Srivastava, S.; Hanash, S.; Wright, G.L., Jr. Proteomics in early detection of cancer. Clin. Chem. 2001, 47, 1901–1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuen, J.W.M.; Gohel, M.D.I. Anticancer effects of Ganoderma lucidum: A review of scientific evidence. Nutr. Cancer Int. J. 2005, 53, 11–17. [Google Scholar] [CrossRef]
- Cör, D.; Knez, Ž.; Knez Hrnčič, M. Antitumour, Antimicrobial, Antioxidant and Antiacetylcholinesterase Effect of Ganoderma Lucidum Terpenoids and Polysaccharides: A Review. Molecules 2018, 23, 649. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Guan, S.H.; Yang, M.; Liu, X.; Guo, D.A. Differential protein expression in mouse splenic mononuclear cells treated with polysaccharides from spores of Ganoderma lucidum. Phytomed. Int. J. Phytother. Phytopharm. 2008, 15, 268–276. [Google Scholar] [CrossRef]
- Gao, Y.; Chan, E.; Zhou, S. Immunomodulating activities of Ganoderma, a mushroom with medicinal properties. Food Rev. Int. 2004, 20, 123–161. [Google Scholar] [CrossRef]
- Guo, L.; Xie, J.; Ruan, Y.; Zhou, L.; Zhu, H.; Yun, X.; Jiang, Y.; Lü, L.; Chen, K.; Min, Z.; et al. Characterization and immunostimulatory activity of a polysaccharide from the spores of Ganoderma lucidum. Int. Immunopharmacol. 2009, 9, 1175–1182. [Google Scholar] [CrossRef]
- Brown, G.D.; Gordon, S. Immune recognition of fungal beta-glucans. Cell. Microbiol. 2005, 7, 471–479. [Google Scholar] [CrossRef]
- Hermeking, H.; Benzinger, A. 14-3-3 proteins in cell cycle regulation. Semin. Cancer Biol. 2006, 16, 183–192. [Google Scholar] [CrossRef]
- Ohtsuka, T.; Ryu, H.; Minamishima, Y.A.; Macip, S.; Sagara, J.; Nakayama, K.I.; Aaronson, S.A.; Lee, S.W. ASC is a Bax adaptor and regulates the p53-Bax mitochondrial apoptosis pathway. Nat. Cell Biol. 2004, 6, 121–128. [Google Scholar] [CrossRef]
- Lafuse, W.P.; Brown, D.; Castle, L.; Zwilling, B.S. Cloning and characterization of a novel cDNA that is IFN-gamma-induced in mouse peritoneal macrophages and encodes a putative GTP-binding protein. J. Leukoc. Biol. 1995, 57, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Tomsig, J.L.; Sohma, H.; Creutz, C.E. Calcium-dependent regulation of tumour necrosis factor-alpha receptor signalling by copine. Biochem. J. 2004, 378 Pt 3, 1089–1094. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.Y.; Kitchens, R.L.; Munford, R.S. Phosphatidylinositides bind to plasma membrane CD14 and can prevent monocyte activation by bacterial lipopolysaccharide. J. Biol. Chem. 1998, 273, 24309–24313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishizaki, H.; Togawa, A.; Tanaka-Okamoto, M.; Hori, K.; Nishimura, M.; Hamaguchi, A.; Imai, T.; Takai, Y.; Miyoshi, J. Defective chemokine-directed lymphocyte migration and development in the absence of Rho guanosine diphosphate-dissociation inhibitors alpha and beta. J. Immunol. 2006, 177, 8512–8521. [Google Scholar] [CrossRef] [Green Version]
- Miletic, A.V.; Swat, M.; Fujikawa, K.; Swat, W. Cytoskeletal remodeling in lymphocyte activation. Curr. Opin. Immunol. 2003, 15, 261–268. [Google Scholar] [CrossRef]
- Fraiser, L.H.; Kanekal, S.; Kehrer, J.P. Cyclophosphamide toxicity. Characterising and avoiding the problem. Drugs 1991, 42, 781–795. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, P.L.; Holl, V.; Coelho, D.; Dufour, P.; Weltin, D.; Luu, B. Apoptosis at the interface of immunosuppressive and anticancer activities: The examples of two classes of chemical inducers, oxysterols and alkylating agents. Curr. Med. Chem. 2000, 7, 693–713. [Google Scholar] [CrossRef]
- Zhu, X.L.; Chen, A.F.; Lin, Z.B. Ganoderma lucidum polysaccharides enhance the function of immunological effector cells in immunosuppressed mice. J. Ethnopharmacol. 2007, 111, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Yue, Q.X.; Guan, S.H.; Wu, W.Y.; Yang, M.; Jiang, B.H.; Liu, X.; Guo, D.A. Proteomic analysis of possible target-related proteins of cyclophosphamide in mice thymus. Food Chem. Toxicol. 2009, 47, 1841–1847. [Google Scholar] [CrossRef]
- Zhang, J.; Ma, K.; Wang, H. Cyclophosphamide suppresses thioredoxin reductase in bladder tissue and its adaptive response via inductions of thioredoxin reductase and glutathione peroxidase. Chem.-Biol. Interact. 2006, 162, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Prescott, S.M.; Zimmerman, G.A.; Stafforini, D.M.; McIntyre, T.M. Platelet-activating factor and related lipid mediators. Annu. Rev. Biochem. 2000, 69, 419–445. [Google Scholar] [CrossRef]
- Zickermann, V.; Dröse, S.; Tocilescu, M.A.; Zwicker, K.; Kerscher, S.; Brandt, U. Challenges in elucidating structure and mechanism of proton pumping NADH:ubiquinone oxidoreductase (complex I). J. Bioenerg. Biomembr. 2008, 40, 475–483. [Google Scholar] [CrossRef]
- Yakovlev, G.; Hirst, J. Transhydrogenation reactions catalyzed by mitochondrial NADH-ubiquinone oxidoreductase (Complex I). Biochemistry 2007, 46, 14250–14258. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, I.; Deshpande, N. Drug effects on certain enzymes of carbohydrate metabolism in MCF-7 cells in culture. Clin. Oncol. 1984, 10, 253–260. [Google Scholar]
- D’Acquisto, F.; Paschalidis, N.; Raza, K.; Buckley, C.D.; Flower, R.J.; Perretti, M. Glucocorticoid treatment inhibits annexin-1 expression in rheumatoid arthritis CD4+ T cells. Rheumatology 2008, 47, 636–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Q.Z.; Lin, Z.B. Antitumor and anti-angiogenic activity of Ganoderma lucidum polysaccharides peptide. Acta Pharmacol. Sin. 2004, 25, 833–838. [Google Scholar]
- Sliva, D.; Labarrere, C.; Slivova, V.; Sedlak, M.; Lloyd, F.P., Jr.; Ho, N.W. Ganoderma lucidum suppresses motility of highly invasive breast and prostate cancer cells. Biochem. Biophys. Res. Commun. 2002, 298, 603–612. [Google Scholar] [CrossRef]
- Müller, C.I.; Kumagai, T.; O’Kelly, J.; Seeram, N.P.; Heber, D.; Koeffler, H.P. Ganoderma lucidum causes apoptosis in leukemia, lymphoma and multiple myeloma cells. Leuk. Res. 2006, 30, 841–848. [Google Scholar] [CrossRef] [PubMed]
- Cheung, W.M.; Hui, W.S.; Chu, P.W.; Chiu, S.W.; Ip, N.Y. Ganoderma extract activates MAP kinases and induces the neuronal differentiation of rat pheochromocytoma PC12 cells. FEBS Lett. 2000, 486, 291–296. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.B.; Wang, R.; Wu, H.L.; Li, Y.H.; Zhong, L.J.; Yu, H.M.; Li, X.J. Serum amyloid A mediates the inhibitory effect of Ganoderma lucidum polysaccharides on tumor cell adhesion to endothelial cells. Oncol. Rep. 2008, 20, 549–556. [Google Scholar] [PubMed]
- Gutfeld, O.; Prus, D.; Ackerman, Z.; Dishon, S.; Linke, R.P.; Levin, M.; Urieli-Shoval, S. Expression of serum amyloid A, in normal, dysplastic, and neoplastic human colonic mucosa: Implication for a role in colonic tumorigenesis. J. Histochem. Cytochem. Off. J. Histochem. Soc. 2006, 54, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Chan, D.C.; Chen, C.J.; Chu, H.C.; Chang, W.K.; Yu, J.C.; Chen, Y.J.; Wen, L.L.; Huang, S.C.; Ku, C.H.; Liu, Y.C.; et al. Evaluation of serum amyloid A as a biomarker for gastric cancer. Ann. Surg. Oncol. 2007, 14, 84–93. [Google Scholar] [CrossRef]
- Preciado-Patt, L.; Levartowsky, D.; Prass, M.; Hershkoviz, R.; Lider, O.; Fridkin, M. Inhibition of cell adhesion to glycoproteins of the extracellular matrix by peptides corresponding to serum amyloid A. Toward understanding the physiological role of an enigmatic protein. Eur. J. Biochem. 1994, 223, 35–42. [Google Scholar] [CrossRef]
- Brinckerhoff, C.E.; Mitchell, T.I.; Karmilowicz, M.J.; Kluve-Beckerman, B.; Benson, M.D. Autocrine induction of collagenase by serum amyloid A-like and beta 2-microglobulin-like proteins. Science 1989, 243, 655–657. [Google Scholar] [CrossRef]
- Mullan, R.H.; Bresnihan, B.; Golden-Mason, L.; Markham, T.; O’Hara, R.; FitzGerald, O.; Veale, D.J.; Fearon, U. Acute-phase serum amyloid A stimulation of angiogenesis, leukocyte recruitment, and matrix degradation in rheumatoid arthritis through an NF-kappaB-dependent signal transduction pathway. Arthritis Rheum. 2006, 54, 105–114. [Google Scholar] [CrossRef]
- Yokoi, K.; Shih, L.C.; Kobayashi, R.; Koomen, J.; Hawke, D.; Li, D.; Hamilton, S.R.; Abbruzzese, J.L.; Coombes, K.R.; Fidler, I.J. Serum amyloid A as a tumor marker in sera of nude mice with orthotopic human pancreatic cancer and in plasma of patients with pancreatic cancer. Int. J. Oncol. 2005, 27, 1361–1369. [Google Scholar] [CrossRef]
- Khan, N.; Cromer, C.J.; Campa, M.; Patz, E.F., Jr. Clinical utility of serum amyloid A and macrophage migration inhibitory factor as serum biomarkers for the detection of nonsmall cell lung carcinoma. Cancer 2004, 101, 379–384. [Google Scholar] [CrossRef]
- Jacob, L.; Scholten, P.C.; Kostev, K.; Kalder, M. Association between sleep disorders and the presence of breast cancer metastases in gynecological practices in Germany: A case-control study of 11,412 women. Breast Cancer Res. Treat. 2018, 171, 443–448. [Google Scholar] [CrossRef]
- Xian, H.; Li, J.; Zhang, Y.; Li, D.; Zhu, Y.; Li, S.; Tan, Z.; Lin, Z.; Li, X.; Pan, Y. Antimetastatic Effects of Ganoderma lucidum Polysaccharide Peptide on B16-F10-luc-G5 Melanoma Mice With Sleep Fragmentation. Front. Pharmacol. 2021, 12, 650216. [Google Scholar] [CrossRef]
- Wang, R.Q.; Lan, Y.L.; Lou, J.C.; Lyu, Y.Z.; Hao, Y.C.; Su, Q.F.; Ma, B.B.; Yuan, Z.B.; Yu, Z.K.; Zhang, H.Q.; et al. Expression and methylation status of LAMA2 are associated with the invasiveness of nonfunctioning PitNET. Ther. Adv. Endocrinol. Metab. 2019, 10, 2042018818821296. [Google Scholar] [CrossRef]
- Lin, T.Y.; Hsu, H.Y. Ling Zhi-8 reduces lung cancer mobility and metastasis through disruption of focal adhesion and induction of MDM2-mediated Slug degradation. Cancer Lett. 2016, 375, 340–348. [Google Scholar] [CrossRef]
- Levy, A.; Alhazzani, K.; Dondapati, P.; Alaseem, A.; Cheema, K.; Thallapureddy, K.; Kaur, P.; Alobid, S.; Rathinavelu, A. Focal Adhesion Kinase in Ovarian Cancer: A Potential Therapeutic Target for Platinum and Taxane-Resistant Tumors. Curr. Cancer Drug Targets 2019, 19, 179–188. [Google Scholar] [CrossRef]
- Ijaz, M.; Wang, F.; Shahbaz, M.; Jiang, W.; Fathy, A.H.; Nesa, E.U. The Role of Grb2 in Cancer and Peptides as Grb2 Antagonists. Protein Pept. Lett. 2018, 24, 1084–1095. [Google Scholar] [CrossRef] [PubMed]
- Zitvogel, L.; Galluzzi, L.; Viaud, S.; Vétizou, M.; Daillère, R.; Merad, M.; Kroemer, G. Cancer and the gut microbiota: An unexpected link. Sci. Transl. Med. 2015, 7, 271ps1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdallah Ismail, N.; Ragab, S.H.; Abd Elbaky, A.; Shoeib, A.R.; Alhosary, Y.; Fekry, D. Frequency of Firmicutes and Bacteroidetes in gut microbiota in obese and normal weight Egyptian children and adults. Arch. Med. Sci. AMS 2011, 7, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Spychala, M.S.; Venna, V.R.; Jandzinski, M.; Doran, S.J.; Durgan, D.J.; Ganesh, B.P.; Ajami, N.J.; Putluri, N.; Graf, J.; Bryan, R.M.; et al. Age-related changes in the gut microbiota influence systemic inflammation and stroke outcome. Ann. Neurol. 2018, 84, 23–36. [Google Scholar] [CrossRef]
- Davis, G.L.; Dempster, J.; Meler, J.D.; Orr, D.W.; Walberg, M.W.; Brown, B.; Berger, B.D.; O’Connor, J.K.; Goldstein, R.M. Hepatocellular carcinoma: Management of an increasingly common problem. Proceedings 2008, 21, 266–280. [Google Scholar] [CrossRef] [Green Version]
- Chai, Y.; Wang, G.; Fan, L.; Zhao, M. A proteomic analysis of mushroom polysaccharide-treated HepG2 cells. Sci. Rep. 2016, 6, 23565. [Google Scholar] [CrossRef]
- OuYang, F.; Wang, G.; Guo, W.; Zhang, Y.; Xiang, W.; Zhao, M. AKT signalling and mitochondrial pathways are involved in mushroom polysaccharide-induced apoptosis and G1 or S phase arrest in human hepatoma cells. Food Chem. 2013, 138, 2130–2139. [Google Scholar] [CrossRef]
- Liu, S.; Yang, Z.; Wei, H.; Shen, W.; Liu, J.; Yin, Q.; Li, X.; Yi, J. Increased DJ-1 and its prognostic significance in hepatocellular carcinoma. Hepato-Gastroenterology 2010, 57, 1247–1256. [Google Scholar]
- Fu, K.; Ren, H.; Wang, Y.; Fei, E.; Wang, H.; Wang, G. DJ-1 inhibits TRAIL-induced apoptosis by blocking pro-caspase-8 recruitment to FADD. Oncogene 2012, 31, 1311–1322. [Google Scholar] [CrossRef] [Green Version]
- De Silva, D.D.; Rapior, S.; Fons, F.; Bahkali, A.H.; Hyde, K.D. Medicinal mushrooms in supportive cancer therapies: An approach to anti-cancer effects and putative mechanisms of action. Fungal. Divers. 2012, 55, 1–35. [Google Scholar] [CrossRef]
- Kino, K.; Yamashita, A.; Yamaoka, K.; Watanabe, J.; Tanaka, S.; Ko, K.; Shimizu, K.; Tsunoo, H. Isolation and characterization of a new immunomodulatory protein, ling zhi-8 (LZ-8), from Ganoderma lucidium. J. Biol. Chem. 1989, 264, 472–478. [Google Scholar] [CrossRef]
- Lin, T.Y.; Hua, W.J.; Yeh, H.; Tseng, A.J. Functional proteomic analysis reveals that fungal immunomodulatory protein reduced expressions of heat shock proteins correlates to apoptosis in lung cancer cells. Phytomed. Int. J. Phytother. Phytopharm. 2021, 80, 153384. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.Y.; Hsu, H.Y.; Sun, W.H.; Wu, T.H.; Tsao, S.M. Induction of Cbl-dependent epidermal growth factor receptor degradation in Ling Zhi-8 suppressed lung cancer: LZ-8 induces EGFR degradation. Int. J. Cancer 2017, 140, 2596–2607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rong, B.; Yang, S. Molecular mechanism and targeted therapy of Hsp90 involved in lung cancer: New discoveries and developments (Review). Int. J. Oncol. 2018, 52, 321–336. [Google Scholar] [CrossRef]
- Vasaikar, S.V.; Ghosh, S.; Narain, P.; Basu, A.; Gomes, J. HSP70 mediates survival in apoptotic cells-Boolean network prediction and experimental validation. Front. Cell. Neurosci. 2015, 9, 319. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Wang, K.; Kuang, S.; Cao, R.; Bao, L.; Liu, R.; Liu, H.; Sun, C. The natural compound GL22, isolated from Ganoderma mushrooms, suppresses tumor growth by altering lipid metabolism and triggering cell death. Cell Death Dis. 2018, 9, 689. [Google Scholar] [CrossRef] [PubMed]
- Veerkamp, J.H.; van Moerkerk, H.T. Fatty acid-binding protein and its relation to fatty acid oxidation. Mol. Cell. Biochem. 1993, 123, 101–106. [Google Scholar] [CrossRef]
- DeBerardinis, R.J.; Lum, J.J.; Hatzivassiliou, G.; Thompson, C.B. The biology of cancer: Metabolic reprogramming fuels cell growth and proliferation. Cell Metab. 2008, 7, 11–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
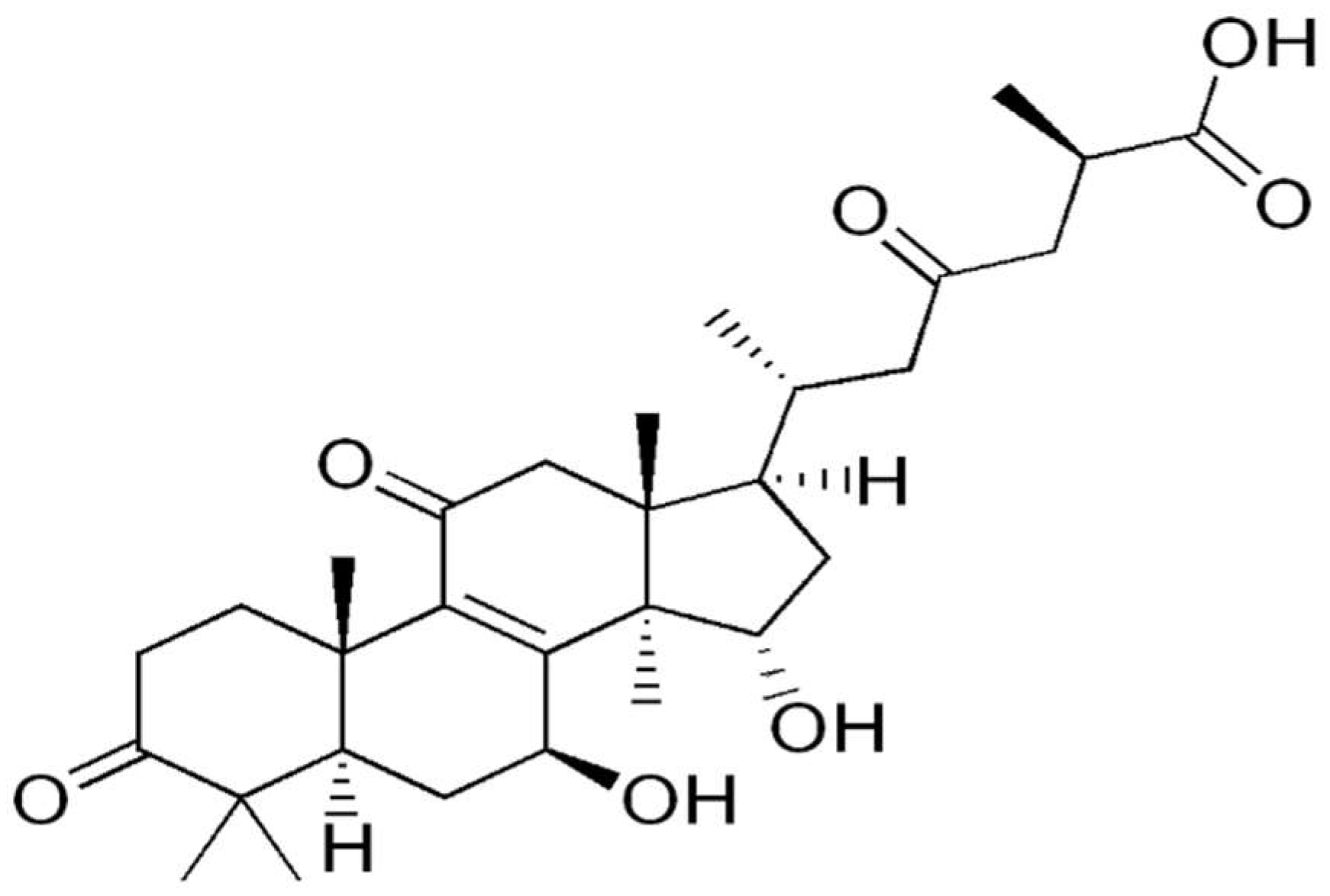
- Yue, Q.X.; Cao, Z.W.; Guan, S.H.; Liu, X.H.; Tao, L.; Wu, W.Y.; Li, Y.X.; Yang, P.Y.; Liu, X.; Guo, D.A. Proteomics characterization of the cytotoxicity mechanism of ganoderic acid D and computer-automated estimation of the possible drug target network. Mol. Cell. Proteom. MCP 2008, 7, 949–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karube, A.; Shidara, Y.; Hayasaka, K.; Maki, M.; Tanaka, T. Suppression of calphobindin I (CPB I) production in carcinoma of uterine cervix and endometrium. Gynecol. Oncol. 1995, 58, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Hayes, M.J.; Moss, S.E. Annexins and disease. Biochem. Biophys. Res. Commun. 2004, 322, 1166–1170. [Google Scholar] [CrossRef] [PubMed]
- Mulla, A.; Christian, H.C.; Solito, E.; Mendoza, N.; Morris, J.F.; Buckingham, J.C. Expression, subcellular localization and phosphorylation status of annexins 1 and 5 in human pituitary adenomas and a growth hormone-secreting carcinoma. Clin. Endocrinol. 2004, 60, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Nonn, L.; Berggren, M.; Powis, G. Increased expression of mitochondrial peroxiredoxin-3 (thioredoxin peroxidase-2) protects cancer cells against hypoxia and drug-induced hydrogen peroxide-dependent apoptosis. Mol. Cancer Res. MCR 2003, 1, 682–689. [Google Scholar] [PubMed]
- Yuan, C.C.; Huang, T.S.; Ng, H.T.; Liu, R.S.; Hung, M.W.; Tsai, L.C. Elevated cytokeratin-19 expression associated with apoptotic resistance and malignant progression of human cervical carcinoma. Apoptosis Int. J. Program. Cell Death 1998, 3, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Yue, Q.X.; Song, X.Y.; Ma, C.; Feng, L.X.; Guan, S.H.; Wu, W.Y.; Yang, M.; Jiang, B.H.; Liu, X.; Cui, Y.J.; et al. Effects of triterpenes from Ganoderma lucidum on protein expression profile of HeLa cells. Phytomed. Int. J. Phytother. Phytopharm. 2010, 17, 606–613. [Google Scholar] [CrossRef] [PubMed]
- Rossi, D.; Kuroshu, R.; Zanelli, C.F.; Valentini, S.R. eIF5A and EF-P: Two unique translation factors are now traveling the same road. Wiley interdisciplinary reviews. RNA 2014, 5, 209–222. [Google Scholar] [CrossRef]
- Wägsäter, D.; Löfgren, S.; Hugander, A.; Dimberg, J. Expression of interleukin-17 in human colorectal cancer. Anticancer Res. 2006, 26, 4213–4216. [Google Scholar] [PubMed]
- Bomsztyk, K.; Denisenko, O.; Ostrowski, J. hnRNP K: One protein multiple processes. BioEssays News Rev. Mol. Cell. Dev. Biol. 2004, 26, 629–638. [Google Scholar] [CrossRef] [PubMed]
- Chevallet, M.; Wagner, E.; Luche, S.; van Dorsselaer, A.; Leize-Wagner, E.; Rabilloud, T. Regeneration of peroxiredoxins during recovery after oxidative stress: Only some overoxidized peroxiredoxins can be reduced during recovery after oxidative stress. J. Biol. Chem. 2003, 278, 37146–37153. [Google Scholar] [CrossRef] [Green Version]
- Andres-Mateos, E.; Perier, C.; Zhang, L.; Blanchard-Fillion, B.; Greco, T.M.; Thomas, B.; Ko, H.S.; Sasaki, M.; Ischiropoulos, H.; Przedborski, S.; et al. DJ-1 gene deletion reveals that DJ-1 is an atypical peroxiredoxin-like peroxidase. Proc. Natl. Acad. Sci. USA 2007, 104, 14807–14812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trachootham, D.; Alexandre, J.; Huang, P. Targeting cancer cells by ROS-mediated mechanisms: A radical therapeutic approach?. Nature reviews. Drug Discov. 2009, 8, 579–591. [Google Scholar] [CrossRef]
- Tsukumo, Y.; Tomida, A.; Kitahara, O.; Nakamura, Y.; Asada, S.; Mori, K.; Tsuruo, T. Nucleobindin 1 controls the unfolded protein response by inhibiting ATF6 activation. J. Biol. Chem. 2007, 282, 29264–29272. [Google Scholar] [CrossRef] [Green Version]
- Ozawa, M.; Muramatsu, T. Reticulocalbin, a novel endoplasmic reticulum resident Ca(2+)-binding protein with multiple EF-hand motifs and a carboxyl-terminal HDEL sequence. J. Biol. Chem. 1993, 268, 699–705. [Google Scholar] [CrossRef]
- Yue, Q.X.; Xie, F.B.; Guan, S.H.; Ma, C.; Yang, M.; Jiang, B.H.; Liu, X.; Guo, D.A. Interaction of Ganoderma triterpenes with doxorubicin and proteomic characterization of the possible molecular targets of Ganoderma triterpenes. Cancer Sci. 2008, 99, 1461–1470. [Google Scholar] [CrossRef] [PubMed]
- Guarguaglini, G.; Renzi, L.; D’Ottavio, F.; Di Fiore, B.; Casenghi, M.; Cundari, E.; Lavia, P. Regulated Ran-binding protein 1 activity is required for organization and function of the mitotic spindle in mammalian cells in vivo. Cell Growth Differ. Mol. Biol. J. Am. Assoc. Cancer Res. 2000, 11, 455–465. [Google Scholar]
- Adams, J.; Palombella, V.J.; Sausville, E.A.; Johnson, J.; Destree, A.; Lazarus, D.D.; Maas, J.; Pien, C.S.; Prakash, S.; Elliott, P.J. Proteasome inhibitors: A novel class of potent and effective antitumor agents. Cancer Res. 1999, 59, 2615–2622. [Google Scholar] [PubMed]
- Ayene, I.S.; Ford, L.P.; Koch, C.J. Ku protein targeting by Ku70 small interfering RNA enhances human cancer cell response to topoisomerase II inhibitor and gamma radiation. Mol. Cancer Ther. 2005, 4, 529–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Li, S.; Wang, X.; Zhang, L.; Cheung, P.C.K. Advances in lentinan: Isolation, structure, chain conformation and bioactivities. Food Hydrocolloid 2011, 25, 196–206. [Google Scholar] [CrossRef]
- Zhang, P.; Zhang, L.; Cheng, S. Chemical Structure and Molecular Weights of α-(1→3)-d-Glucan from Lentinus edodes. Biosci. Biotechnol. Biochem. 1999, 63, 1197–1202. [Google Scholar] [CrossRef]
- Zhang, T.; Chen, X.; Sun, L.; Guo, X.; Cai, T.; Wang, J.; Zeng, Y.; Ma, J.; Ding, X.; Xie, Z.; et al. Proteomics reveals the function reverse of MPSSS-treated prostate cancer-associated fibroblasts to suppress PC-3 cell viability via the FoxO pathway. Cancer Med. 2021, 10, 2509–2522. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Tao, N.; Liu, X.; Li, X.; Tang, J.; Ma, C.; Xu, X.; Shao, H.; Hou, B.; Wang, H.; et al. Polysaccharide from Lentinus edodes inhibits the immunosuppressive function of myeloid-derived suppressor cells. PLoS ONE 2012, 7, e51751. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Ma, J.; Zheng, Q.; Wang, Y.; Hu, M.; Ma, F.; Qin, Z.; Lei, N.; Tao, N. MPSSS impairs the immunosuppressive function of cancer-associated fibroblasts via the TLR4-NF-κB pathway. Biosci. Rep. 2019, 39, BSR20182171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erdogan, B.; Webb, D.J. Cancer-associated fibroblasts modulate growth factor signaling and extracellular matrix remodeling to regulate tumor metastasis. Biochem. Soc. Trans. 2017, 45, 229–236. [Google Scholar] [CrossRef] [Green Version]
- Puglisi, R.; Pastore, A. The role of chaperones in iron-sulfur cluster biogenesis. FEBS Lett. 2018, 592, 4011–4019. [Google Scholar] [CrossRef]
- Laverty, H.G.; Wakefield, L.M.; Occleston, N.L.; O’Kane, S.; Ferguson, M.W. TGF-beta3 and cancer: A review. Cytokine Growth Factor Rev. 2009, 20, 305–317. [Google Scholar] [CrossRef]
- Chung, K.S.; Cho, S.H.; Shin, J.S.; Kim, D.H.; Choi, J.H.; Choi, S.Y.; Rhee, Y.K.; Hong, H.D.; Lee, K.T. Ginsenoside Rh2 induces cell cycle arrest and differentiation in human leukemia cells by upregulating TGF-β expression. Carcinogenesis 2013, 34, 331–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chihara, G.; Hamuro, J.; Maeda, Y.Y.; Shiio, T.; Suga, T.; Takasuka, N.; Sasaki, T. Antitumor and metastasis-inhibitory activities of lentinan as an immunomodulator: An overview. Cancer detection and prevention. Suppl. Off. Publ. Int. Soc. Prev. Oncol. 1987, 1, 423–443. [Google Scholar]
- Chen, Y.W.; Hu, D.J.; Cheong, K.L.; Li, J.; Xie, J.; Zhao, J.; Li, S.P. Quality evaluation of lentinan injection produced in China. J. Pharm. Biomed. Anal. 2013, 78–79, 176–182. [Google Scholar] [CrossRef]
- Wang, Y.; Han, X.; Li, Y.D.; Wang, Y.; Zhao, S.Y.; Zhang, D.J.; Lu, Y. Lentinan dose dependence between immunoprophylaxis and promotion of the murine liver cancer. Oncotarget 2017, 8, 95152–95162. [Google Scholar] [CrossRef] [Green Version]
- Takai, E.; Tsukimoto, M.; Harada, H.; Sawada, K.; Moriyama, Y.; Kojima, S. Autocrine regulation of TGF-β1-induced cell migration by exocytosis of ATP and activation of P2 receptors in human lung cancer cells. J. Cell Sci. 2012, 125 Pt 21, 5051–5060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, W.; Fu, Z.; Wang, H.; Feng, J.; Wei, J.; Guo, J. Peroxiredoxin 2 is upregulated in colorectal cancer and contributes to colorectal cancer cells’ survival by protecting cells from oxidative stress. Mol. Cell. Biochem. 2014, 387, 261–270. [Google Scholar] [CrossRef]
- Xue, G.; Hao, L.Q.; Ding, F.X.; Mei, Q.; Huang, J.J.; Fu, C.G.; Yan, H.L.; Sun, S.H. Expression of annexin a5 is associated with higher tumor stage and poor prognosis in colorectal adenocarcinomas. J. Clin. Gastroenterol. 2009, 43, 831–837. [Google Scholar] [CrossRef]
- Liu, Z.; Zhan, Y.; Tu, Y.; Chen, K.; Liu, Z.; Wu, C. PDZ and LIM domain protein 1(PDLIM1)/CLP36 promotes breast cancer cell migration, invasion and metastasis through interaction with α-actinin. Oncogene 2015, 34, 1300–1311. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Reynolds, A.B.; Kanner, S.B.; Vines, R.R.; Parsons, J.T. Identification and characterization of a novel cytoskeleton-associated pp60src substrate. Mol. Cell. Biol. 1991, 11, 5113–5124. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Liu, D.Y.; Yuan, X.R.; Liu, Q.; Jiang, X.J.; Yuan, D.; Huang, J.; Li, X.J.; Yang, Z.Q. The expression of moesin in astrocytoma: Correlation with pathologic grade and poor clinical outcome. Med. Oncol. 2013, 30, 372. [Google Scholar] [CrossRef]
- Yang, X.; Li, Y.; Wang, W.; Li, C.; Zhao, D.; Hu, G.; Rao, H.; Hao, J.; Han, X. Antigens from H22 Cell Mutation Induced by Lentinan Are Correlated with HCC Immunoprophylaxis. Available online: https://www.researchsquare.com/article/rs-43542/v1 (accessed on 16 October 2021).
- Zhang, G.; Feng, W.; Wu, J. Down-regulation of SEPT9 inhibits glioma progression through suppressing TGF-β-induced epithelial-mesenchymal transition (EMT). Biomed. Pharmacother. Biomed. Pharmacother. 2020, 125, 109768. [Google Scholar] [CrossRef]
- Zhou, Q.; Bauden, M.; Andersson, R.; Hu, D.; Marko-Varga, G.; Xu, J.; Sasor, A.; Dai, H.; Pawłowski, K.; Said Hilmersson, K.; et al. YAP1 is an independent prognostic marker in pancreatic cancer and associated with extracellular matrix remodeling. J. Transl. Med. 2020, 18, 77. [Google Scholar] [CrossRef]
- Weaver, A.M. Invadopodia: Specialized cell structures for cancer invasion. Clin. Exp. Metastasis 2006, 23, 97–105. [Google Scholar] [CrossRef]
- Wang, W.; Yang, X.; Li, C.; Li, Y.; Wang, H.; Han, X. Immunogenic Cell Death (ICD) of Murine H22 Cells Induced by Lentinan. Nutr. Cancer 2021, 1–10. [Google Scholar] [CrossRef]
- Krysko, D.V.; Garg, A.D.; Kaczmarek, A.; Krysko, O.; Agostinis, P.; Vandenabeele, P. Immunogenic cell death and DAMPs in cancer therapy. Nature reviews. Cancer 2012, 12, 860–875. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.D.; Martin, S.; Golab, J.; Agostinis, P. Danger signalling during cancer cell death: Origins, plasticity and regulation. Cell Death Differ. 2014, 21, 26–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casares, N.; Pequignot, M.O.; Tesniere, A.; Ghiringhelli, F.; Roux, S.; Chaput, N.; Schmitt, E.; Hamai, A.; Hervas-Stubbs, S.; Obeid, M.; et al. Caspase-dependent immunogenicity of doxorubicin-induced tumor cell death. J. Exp. Med. 2005, 202, 1691–1701. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.J.; Qin, Y.; Zhao, Z.H.; Zhang, Y.; Yang, J.H.; Zhai, D.H.; Cui, F.; Luo, C.; Lu, M.X.; Liu, P.P.; et al. Lentinan-functionalized Selenium Nanoparticles target Tumor Cell Mitochondria via TLR4/TRAF3/MFN1 pathway. Theranostics 2020, 10, 9083–9099. [Google Scholar] [CrossRef]
- Barni, S.; Cabiddu, M.; Ghilardi, M.; Petrelli, F. A novel perspective for an orphan problem: Old and new drugs for the medical management of malignant ascites. Crit. Rev. Oncol./Hematol. 2011, 79, 144–153. [Google Scholar] [CrossRef]
- Kampan, N.C.; Madondo, M.T.; McNally, O.M.; Stephens, A.N.; Quinn, M.A.; Plebanski, M. Interleukin 6 Present in Inflammatory Ascites from Advanced Epithelial Ovarian Cancer Patients Promotes Tumor Necrosis Factor Receptor 2-Expressing Regulatory T Cells. Front. Immunol. 2017, 8, 1482. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, H. Chemotherapy drugs induce pyroptosis through caspase-3-dependent cleavage of GSDME. Science China. Life Sci. 2018, 61, 739–740. [Google Scholar] [CrossRef]
- Davidovich, P.; Kearney, C.J.; Martin, S.J. Inflammatory outcomes of apoptosis, necrosis and necroptosis. Biol. Chem. 2014, 395, 1163–1171. [Google Scholar] [CrossRef]
- Fernandes, A.P.; Gandin, V. Selenium compounds as therapeutic agents in cancer. Biochim. Biophys. Acta 2015, 1850, 1642–1660. [Google Scholar] [CrossRef]
- Jeong, M.K.; Yoo, H.S.; Kang, I.C. The Extract of Cordyceps Militaris Inhibited the Proliferation of Cisplatin-Resistant A549 Lung Cancer Cells by Downregulation of H-Ras. J. Med. Food 2019, 22, 823–832. [Google Scholar] [CrossRef] [PubMed]
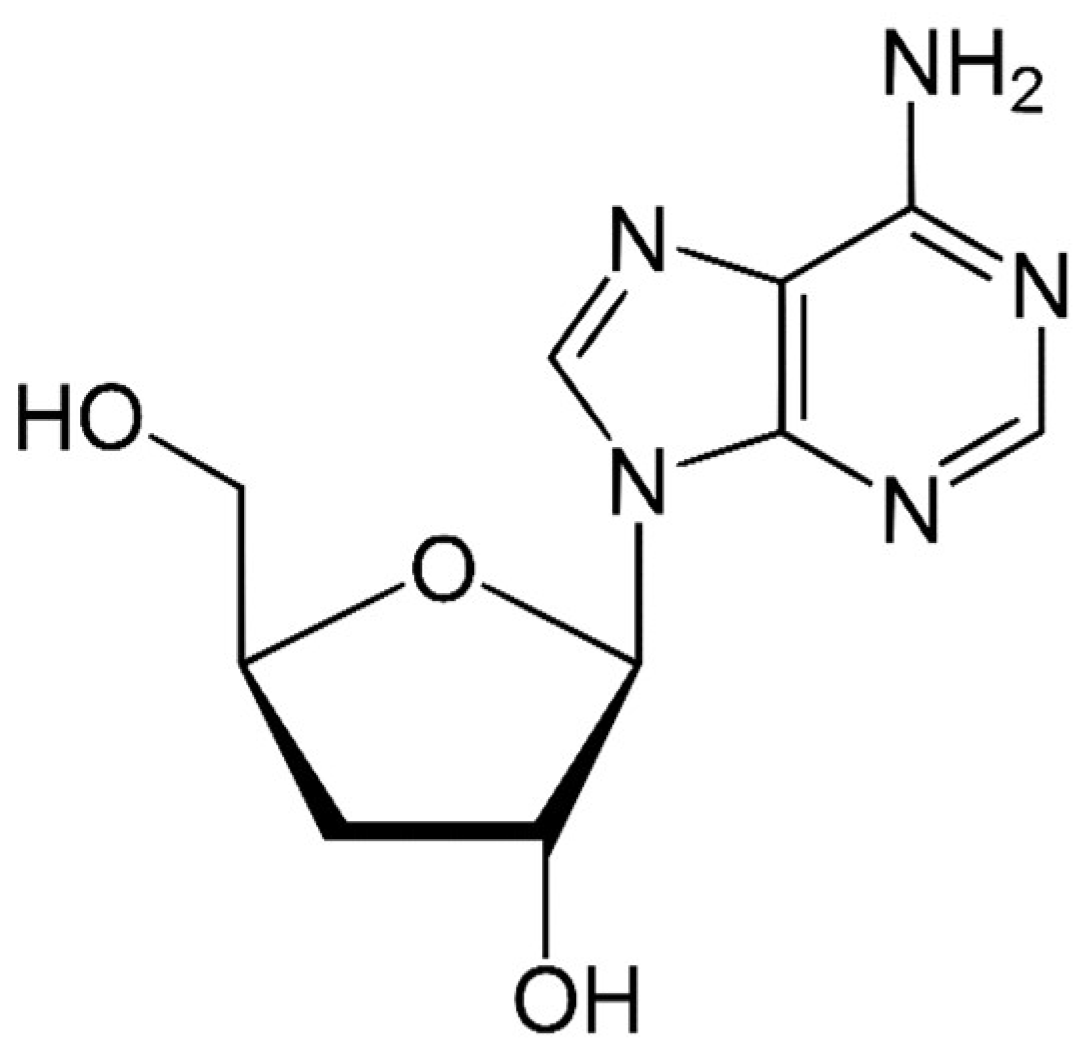
- Nakamura, K.; Konoha, K.; Yoshikawa, N.; Yamaguchi, Y.; Kagota, S.; Shinozuka, K.; Kunitomo, M. Effect of cordycepin (3′-deoxyadenosine) on hematogenic lung metastatic model mice. In Vivo 2005, 19, 137–141. [Google Scholar] [PubMed]
- Tian, X.; Li, Y.; Shen, Y.; Li, Q.; Wang, Q.; Feng, L. Apoptosis and inhibition of proliferation of cancer cells induced by cordycepin. Oncol. Lett. 2015, 10, 595–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, J.H.; Ng, T.B.; Cheung, R.C.; Ye, X.J.; Wang, H.X.; Lam, S.K.; Lin, P.; Chan, Y.S.; Fang, E.F.; Ngai, P.H.; et al. Proteins with antifungal properties and other medicinal applications from plants and mushrooms. Appl. Microbiol. Biotechnol. 2010, 87, 1221–1235. [Google Scholar] [CrossRef]
- Meng, F.; Cao, B.; Feng, Z.; Ma, S.; Wang, H.; Li, Y.; Li, H. Knockdown of mutated H-Ras V12 expression induces chemosensitivity of hepatocellular carcinoma cells to cisplatin treatment in vitro and in nude mouse xenografts. Oncol. Rep. 2014, 32, 2023–2030. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Li, J.; Gu, B.; Xiao, Y.; Chen, R.; Liu, X.; Xie, X.; Cao, L. Extracts of Cordyceps sinensis inhibit breast cancer cell metastasis via down-regulation of metastasis-related cytokines expression. J. Ethnopharmacol. 2018, 214, 106–112. [Google Scholar] [CrossRef]
- Joyce, J.A.; Pollard, J.W. Microenvironmental regulation of metastasis. Nat. Rev. Cancer 2009, 9, 239–252. [Google Scholar] [CrossRef]
- Colombo, M.P.; Chiodoni, C. Abstract A102: Osteopontin produced by myeloid cells determines the outcome of breast cancer metastases. Mol. Cancer Res. 2013, 11. [Google Scholar] [CrossRef]
- Liu, J.; Shen, J.X.; Hu, J.L.; Huang, W.H.; Zhang, G.J. Significance of interleukin-33 and its related cytokines in patients with breast cancers. Front. Immunol. 2014, 5, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iellem, A.; Mariani, M.; Lang, R.; Recalde, H.; Panina-Bordignon, P.; Sinigaglia, F.; D’Ambrosio, D. Unique chemotactic response profile and specific expression of chemokine receptors CCR4 and CCR8 by CD4(+)CD25(+) regulatory T cells. J. Exp. Med. 2001, 194, 847–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasiljevic, J.D.; Zivkovic, L.P.; Cabarkapa, A.M.; Bajic, V.P.; Djelic, N.J.; Spremo-Potparevic, B.M. Cordyceps sinensis: Genotoxic Potential in Human Peripheral Blood Cells and Antigenotoxic Properties Against Hydrogen Peroxide by Comet Assay. Altern. Ther. Health Med. 2016, 22 (Suppl. 2), 24–31. [Google Scholar] [PubMed]
- Wang, P.W.; Hung, Y.C.; Li, W.T.; Yeh, C.T.; Pan, T.L. Systematic revelation of the protective effect and mechanism of Cordycep sinensis on diethylnitrosamine-induced rat hepatocellular carcinoma with proteomics. Oncotarget 2016, 7, 60270–60289. [Google Scholar] [CrossRef] [Green Version]
- Paula Santos, N.; Colaço, A.; Gil da Costa, R.M.; Manuel Oliveira, M.; Peixoto, F.; Alexandra Oliveira, P. N-diethylnitrosamine mouse hepatotoxicity: Time-related effects on histology and oxidative stress. Exp. Toxicol. Pathol. Off. J. Ges. Fur Toxikol. Pathol. 2014, 66, 429–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.R.; Choi, H.N.; Lee, H.J.; Baek, H.A.; Park, H.S.; Jang, K.Y.; Chung, M.J.; Moon, W.S. A peroxisome proliferator-activated receptor gamma antagonist induces vimentin cleavage and inhibits invasion in high-grade hepatocellular carcinoma. Oncol. Rep. 2007, 18, 825–832. [Google Scholar]
- Wang, H.; Zhang, J.; Sit, W.H.; Lee, C.Y.; Wan, J.M. Cordyceps cicadae induces G2/M cell cycle arrest in MHCC97H human hepatocellular carcinoma cells: A proteomic study. Chin. Med. 2014, 9, 15. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.N.; Chen, C.H.; Sheu, J.C.; Lee, H.S.; Huang, G.T.; Yu, C.Y.; Lu, F.J.; Chow, L.P. Identification of human hepatocellular carcinoma-related biomarkers by two-dimensional difference gel electrophoresis and mass spectrometry. J. Proteome Res. 2005, 4, 2062–2069. [Google Scholar] [CrossRef]
- Hamaguchi, T.; Iizuka, N.; Tsunedomi, R.; Hamamoto, Y.; Miyamoto, T.; Iida, M.; Tokuhisa, Y.; Sakamoto, K.; Takashima, M.; Tamesa, T.; et al. Glycolysis module activated by hypoxia-inducible factor 1alpha is related to the aggressive phenotype of hepatocellular carcinoma. Int. J. Oncol. 2008, 33, 725–731. [Google Scholar]
- Wei, J.; Fang, D. Endoplasmic Reticulum Stress Signaling and the Pathogenesis of Hepatocarcinoma. Int. J. Mol. Sci. 2021, 22, 1799. [Google Scholar] [CrossRef] [PubMed]
- Halder, S.K.; Anumanthan, G.; Maddula, R.; Mann, J.; Chytil, A.; Gonzalez, A.L.; Washington, M.K.; Moses, H.L.; Beauchamp, R.D.; Datta, P.K. Oncogenic function of a novel WD-domain protein, STRAP, in human carcinogenesis. Cancer Res. 2006, 66, 6156–6166. [Google Scholar] [CrossRef] [Green Version]
- Noh, D.Y.; Ahn, S.J.; Lee, R.A.; Kim, S.W.; Park, I.A.; Chae, H.Z. Overexpression of peroxiredoxin in human breast cancer. Anticancer Res. 2001, 21, 2085–2090. [Google Scholar]
- Mishra, V.; Tomar, S.; Yadav, P.; Singh, M.P. Promising anticancer activity of polysaccharides and other macromolecules derived from oyster mushroom (Pleurotus sp.): An updated review. Int. J. Biol. Macromol. 2021, 182, 1628–1637. [Google Scholar] [CrossRef]
- Finimundy, T.C.; Abreu, R.; Bonetto, N.; Scariot, F.J.; Dillon, A.; Echeverrigaray, S.; Barros, L.; Ferreira, I.; Henriques, J.; Roesch-Ely, M. Apoptosis induction by Pleurotus sajorcaju (Fr.) Singer extracts on colorectal cancer cell lines. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2018, 112, 383–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berg, K.; Eide, P.W.; Eilertsen, I.A.; Johannessen, B.; Bruun, J.; Danielsen, S.A.; Bjørnslett, M.; Meza-Zepeda, L.A.; Eknæs, M.; Lind, G.E.; et al. Multi-omics of 34 colorectal cancer cell lines—A resource for biomedical studies. Mol. Cancer 2017, 16, 116. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Hong, S.H.; Park, C.; Kim, G.Y.; Leem, S.H.; Choi, S.H.; Keum, Y.S.; Hyun, J.W.; Kwon, T.K.; Hong, S.H.; et al. Hwang-Heuk-San induces apoptosis in HCT116 human colorectal cancer cells through the ROS-mediated activation of caspases and the inactivation of the PI3K/Akt signaling pathway. Oncol. Rep. 2016, 36, 205–214. [Google Scholar] [CrossRef]
- Ma, N.; Du, H.; Ma, G.; Yang, W.; Han, Y.; Hu, Q.; Xiao, H. Characterization of the Immunomodulatory Mechanism of a Pleurotus eryngii Protein by Isobaric Tags for Relative and Absolute Quantitation Proteomics. J. Agric. Food Chem. 2020, 68, 13189–13199. [Google Scholar] [CrossRef]
- Hu, Q.; Du, H.; Ma, G.; Pei, F.; Ma, N.; Yuan, B.; Nakata, P.A.; Yang, W. Purification, identification and functional characterization of an immunomodulatory protein from Pleurotus eryngii. Food Funct. 2018, 9, 3764–3775. [Google Scholar] [CrossRef]
- Zotti, T.; Scudiero, I.; Settembre, P.; Ferravante, A.; Mazzone, P.; D’Andrea, L.; Reale, C.; Vito, P.; Stilo, R. TRAF6-mediated ubiquitination of NEMO requires p62/sequestosome-1. Mol. Immunol. 2014, 58, 27–31. [Google Scholar] [CrossRef]
- Poligone, B.; Baldwin, A.S. Positive and negative regulation of NF-kappaB by COX-2: Roles of different prostaglandins. J. Biol. Chem. 2001, 276, 38658–38664. [Google Scholar] [CrossRef] [Green Version]
- Nobre, C.C.; de Araújo, J.M.; Fernandes, T.A.; Cobucci, R.N.; Lanza, D.C.; Andrade, V.S.; Fernandes, J.V. Macrophage Migration Inhibitory Factor (MIF): Biological Activities and Relation with Cancer. Pathol. Oncol. Res. POR 2017, 23, 235–244. [Google Scholar] [CrossRef]
- Zinatizadeh, M.R.; Momeni, S.A.; Zarandi, P.K.; Chalbatani, G.M.; Dana, H.; Mirzaei, H.R.; Akbari, M.E.; Miri, S.R. The Role and Function of Ras-association domain family in Cancer: A Review. Genes Dis. 2019, 6, 378–384. [Google Scholar] [CrossRef]
- Yang, C.S.; Lee, D.S.; Song, C.H.; An, S.J.; Li, S.; Kim, J.M.; Kim, C.S.; Yoo, D.G.; Jeon, B.H.; Yang, H.Y.; et al. Roles of peroxiredoxin II in the regulation of proinflammatory responses to LPS and protection against endotoxin-induced lethal shock. J. Exp. Med. 2007, 204, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Miyake, T.; Shirakawa, H.; Kusano, A.; Sakimoto, S.; Konno, M.; Nakagawa, T.; Mori, Y.; Kaneko, S. TRPM2 contributes to LPS/IFNγ-induced production of nitric oxide via the p38/JNK pathway in microglia. Biochem. Biophys. Res. Commun. 2014, 444, 212–217. [Google Scholar] [CrossRef]
- Katano, M.; Yamamoto, H.; Torisu, M. Gan to kagaku ryoho. Cancer Chemother. 1987, 14, 2321–2326. [Google Scholar]
- Kobayashi, H.; Matsunaga, K.; Oguchi, Y. Antimetastatic effects of PSK (Krestin), a protein-bound polysaccharide obtained from basidiomycetes: An overview. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 1995, 4, 275–281. [Google Scholar]
- Tanaka, K.; Wakasugi, J.; Seki, F.; Koizumi, Y.; Tsuchiya, S. Effect of PSK on cell growth, chemotaxis and platelet aggregating activity of colon 26 cell line. In Proceedings of the 50th Annual Japanese Cancer Association Meeting, Tokyo, Japan, 10–12 September 1991; p. 263. [Google Scholar]
- Hattori, T.S.; Komatsu, N.; Shichijo, S.; Itoh, K. Protein-bound polysaccharide K induced apoptosis of the human Burkitt lymphoma cell line, Namalwa. Biomed. Pharmacother. Biomed. Pharmacother. 2004, 58, 226–230. [Google Scholar] [CrossRef]
- Jiménez-Medina, E.; Berruguilla, E.; Romero, I.; Algarra, I.; Collado, A.; Garrido, F.; Garcia-Lora, A. The immunomodulator PSK induces in vitro cytotoxic activity in tumour cell lines via arrest of cell cycle and induction of apoptosis. BMC Cancer 2008, 8, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.L.; Jiang, P.P.; Sit, W.H.; Wan, J.M. Proteome of human T lymphocytes with treatment of cyclosporine and polysaccharopeptide: Analysis of significant proteins that manipulate T cells proliferation and immunosuppression. Int. Immunopharmacol. 2007, 7, 1311–1324. [Google Scholar] [CrossRef]
- Morris, P.J. Cyclosporine, FK-506 and other drugs in organ transplantation. Curr. Opin. Immunol. 1991, 3, 748–751. [Google Scholar] [CrossRef]
- Sheil, A.G. Cancer in immune-suppressed organ transplant recipients: Aetiology and evolution. Transplant. Proc. 1998, 30, 2055–2057. [Google Scholar] [CrossRef]
- Cui, J.; Chisti, Y. Polysaccharopeptides of Coriolus versicolor: Physiological activity, uses, and production. Biotechnol. Adv. 2003, 21, 109–122. [Google Scholar] [CrossRef]
- Ng, T.B. A review of research on the protein-bound polysaccharide (polysaccharopeptide, PSP) from the mushroom Coriolus versicolor (Basidiomycetes: Polyporaceae). Gen. Pharmacol. 1998, 30, 1–4. [Google Scholar] [CrossRef]
- Ooi, V.E.; Liu, F. Immunomodulation and anti-cancer activity of polysaccharide-protein complexes. Curr. Med. Chem. 2000, 7, 715–729. [Google Scholar] [CrossRef] [Green Version]
- Olofsson, B. Rho guanine dissociation inhibitors: Pivotal molecules in cellular signalling. Cell. Signal. 1999, 11, 545–554. [Google Scholar] [CrossRef]
- Plow, E.F.; Herren, T.; Redlitz, A.; Miles, L.A.; Hoover-Plow, J.L. The cell biology of the plasminogen system. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 1995, 9, 939–945. [Google Scholar] [CrossRef]
- Yu, C.L.; Burakoff, S.J. Involvement of proteasomes in regulating Jak-STAT pathways upon interleukin-2 stimulation. J. Biol. Chem. 1997, 272, 14017–14020. [Google Scholar] [CrossRef] [Green Version]
- Rabinovich, G.A.; Alonso, C.R.; Sotomayor, C.E.; Durand, S.; Bocco, J.L.; Riera, C.M. Molecular mechanisms implicated in galectin-1-induced apoptosis: Activation of the AP-1 transcription factor and downregulation of Bcl-2. Cell Death Differ. 2000, 7, 747–753. [Google Scholar] [CrossRef] [Green Version]
- Theil, E.C. Ferritin: At the crossroads of iron and oxygen metabolism. J. Nutr. 2003, 133 (Suppl. 1), 1549S–1553S. [Google Scholar] [CrossRef]
- Chang, T.S.; Jeong, W.; Choi, S.Y.; Yu, S.; Kang, S.W.; Rhee, S.G. Regulation of peroxiredoxin I activity by Cdc2-mediated phosphorylation. J. Biol. Chem. 2002, 277, 25370–25376. [Google Scholar] [CrossRef] [Green Version]
- Friedman, M. Chemistry, Nutrition, and Health-Promoting Properties of Hericium erinaceus (Lion’s Mane) Mushroom Fruiting Bodies and Mycelia and Their Bioactive Compounds. J. Agric. Food Chem. 2015, 63, 7108–7123. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Konishi, T.; Gao, Y.; Xu, D.; Gao, Q. Anti-Gastric Ulcer Activity of Polysaccharide Fraction Isolated from Mycelium Culture of Lion’s Mane Medicinal Mushroom, Hericium erinaceus (Higher Basidiomycetes). Int. J. Med. Mushrooms 2015, 17, 1055–1060. [Google Scholar] [CrossRef]
- Liu, J.Y.; Hou, X.X.; Li, Z.Y.; Shan, S.H.; Chang, M.C.; Feng, C.P.; Wei, Y. Isolation and structural characterization of a novel polysaccharide from Hericium erinaceus fruiting bodies and its arrest of cell cycle at S-phage in colon cancer cells. Int. J. Biol. Macromol. 2020, 157, 288–295. [Google Scholar] [CrossRef]
- Kim, S.P.; Nam, S.H.; Friedman, M. Hericium erinaceus (Lion’s Mane) mushroom extracts inhibit metastasis of cancer cells to the lung in CT-26 colon cancer-tansplanted mice. J. Agric. Food Chem. 2013, 61, 4898–4904. [Google Scholar] [CrossRef]
- Orditura, M.; Galizia, G.; Sforza, V.; Gambardella, V.; Fabozzi, A.; Laterza, M.M.; Andreozzi, F.; Ventriglia, J.; Savastano, B.; Mabilia, A.; et al. Treatment of gastric cancer. World J. Gastroenterol. 2014, 20, 1635–1649. [Google Scholar] [CrossRef]
- Kuo, H.C.; Kuo, Y.R.; Lee, K.F.; Hsieh, M.C.; Huang, C.Y.; Hsieh, Y.Y.; Lee, K.C.; Kuo, H.L.; Lee, L.Y.; Chen, W.P.; et al. A Comparative Proteomic Analysis of Erinacine A’s Inhibition of Gastric Cancer Cell Viability and Invasiveness. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 43, 195–208. [Google Scholar] [CrossRef]
- Mhawech, P. 14-3-3 proteins—An update. Cell Res. 2005, 15, 228–236. [Google Scholar] [CrossRef]
- Zhou, F.; Chen, E.; You, D.; Song, Y.; Sun, Z.; Yue, L. Both high expression of nucleophosmin/B23 and CRM1 predicts poorer prognosis in human gastric cancer. APMIS Acta Pathol. Microbiol. Immunol. Scand. 2016, 124, 1046–1053. [Google Scholar] [CrossRef] [PubMed]
- Lindström, M.S. NPM1/B23: A Multifunctional Chaperone in Ribosome Biogenesis and Chromatin Remodeling. Biochem. Res. Int. 2011, 2011, 195209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.C.; Kuo, H.C.; Shen, C.H.; Lu, C.C.; Huang, W.S.; Hsieh, M.C.; Huang, C.Y.; Kuo, Y.H.; Hsieh, Y.Y.; Teng, C.C.; et al. A proteomics approach to identifying novel protein targets involved in erinacine A-mediated inhibition of colorectal cancer cells’ aggressiveness. J. Cell. Mol. Med. 2017, 21, 588–599. [Google Scholar] [CrossRef] [Green Version]
- Kallergi, G.; Agelaki, S.; Markomanolaki, H.; Georgoulias, V.; Stournaras, C. Activation of FAK/PI3K/Rac1 signaling controls actin reorganization and inhibits cell motility in human cancer cells. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2007, 20, 977–986. [Google Scholar] [CrossRef] [PubMed]
- Lian, J.; Tang, J.; Shi, H.; Li, H.; Zhen, T.; Xie, W.; Zhang, F.; Yang, Y.; Han, A. Positive feedback loop of hepatoma-derived growth factor and β-catenin promotes carcinogenesis of colorectal cancer. Oncotarget 2015, 6, 29357–29374. [Google Scholar] [CrossRef] [Green Version]
- Maekawa, M.; Ishizaki, T.; Boku, S.; Watanabe, N.; Fujita, A.; Iwamatsu, A.; Obinata, T.; Ohashi, K.; Mizuno, K.; Narumiya, S. Signaling from Rho to the actin cytoskeleton through protein kinases ROCK and LIM-kinase. Science 1999, 285, 895–898. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Jaramillo, M.; Whaley, D.; Wells, A.; Panchapakesa, V.; Das, T.; Roy, P. Profilin-1 is a negative regulator of mammary carcinoma aggressiveness. Br. J. Cancer 2007, 97, 1361–1371. [Google Scholar] [CrossRef] [Green Version]
- Zong, A.; Cao, H.; Wang, F. Anticancer polysaccharides from natural resources: A review of recent research. Carbohydr. Polym. 2012, 90, 1395–1410. [Google Scholar] [CrossRef]
- Li, Y.G.; Ji, D.F.; Zhong, S.; Zhu, J.X.; Chen, S.; Hu, G.Y. Anti-tumor effects of proteoglycan from Phellinus linteus by immunomodulating and inhibiting Reg IV/EGFR/Akt signaling pathway in colorectal carcinoma. Int. J. Biol. Macromol. 2011, 48, 511–517. [Google Scholar] [CrossRef]
- Li, Y.G.; Ji, D.F.; Zhong, S.; Liu, P.G.; Lv, Z.Q.; Zhu, J.X.; Chen, J.E.; Chen, H.P. Polysaccharide from Phellinus linteus induces S-phase arrest in HepG2 cells by decreasing calreticulin expression and activating the P27kip1-cyclin A/D1/E-CDK2 pathway. J. Ethnopharmacol. 2013, 150, 187–195. [Google Scholar] [CrossRef]
- Michalak, M.; Groenendyk, J.; Szabo, E.; Gold, L.I.; Opas, M. Calreticulin, a multi-process calcium-buffering chaperone of the endoplasmic reticulum. Biochem. J. 2009, 417, 651–666. [Google Scholar] [CrossRef] [PubMed]
- Erić, A.; Juranić, Z.; Milovanović, Z.; Marković, I.; Inić, M.; Stanojević-Bakić, N.; Vojinović-Golubović, V. Effects of humoral immunity and calreticulin overexpression on postoperative course in breast cancer. Pathol. Oncol. Res. 2009, 15, 89–90. [Google Scholar] [CrossRef]
- Chen, C.N.; Chang, C.C.; Su, T.E.; Hsu, W.M.; Jeng, Y.M.; Ho, M.C.; Hsieh, F.J.; Lee, P.H.; Kuo, M.L.; Lee, H.; et al. Identification of calreticulin as a prognosis marker and angiogenic regulator in human gastric cancer. Ann. Surg. Oncol. 2009, 16, 524–533. [Google Scholar] [CrossRef]
- Nanba, H.; Kubo, K. Effect of Maitake D-fraction on cancer prevention. Ann. N. Y. Acad. Sci. 1997, 833, 204–207. [Google Scholar] [CrossRef] [PubMed]
- Matsui, K.; Kodama, N.; Nanba, H. Effects of maitake (Grifola frondosa) D-Fraction on the carcinoma angiogenesis. Cancer Lett. 2001, 172, 193–198. [Google Scholar] [CrossRef]
- Shoji-Kawata, S.; Sumpter, R.; Leveno, M.; Campbell, G.R.; Zou, Z.; Kinch, L.; Wilkins, A.D.; Sun, Q.; Pallauf, K.; MacDuff, D.; et al. Identification of a candidate therapeutic autophagy-inducing peptide. Nature 2013, 494, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Cui, F.; Zan, X.; Li, Y.; Sun, W.; Yang, Y.; Ping, L. Grifola frondosa Glycoprotein GFG-3a Arrests S phase, Alters Proteome, and Induces Apoptosis in Human Gastric Cancer Cells. Nutr. Cancer 2016, 68, 267–279. [Google Scholar] [CrossRef] [PubMed]
- Gorynia, S.; Bandeiras, T.M.; Pinho, F.G.; McVey, C.E.; Vonrhein, C.; Round, A.; Svergun, D.I.; Donner, P.; Matias, P.M.; Carrondo, M.A. Structural and functional insights into a dodecameric molecular machine—The RuvBL1/RuvBL2 complex. J. Struct. Biol. 2011, 176, 279–291. [Google Scholar] [CrossRef]
- Gartner, W.; Rossbacher, J.; Zierhut, B.; Daneva, T.; Base, W.; Weissel, M.; Waldhäusl, W.; Pasternack, M.S.; Wagner, L. The ATP-dependent helicase RUVBL1/TIP49a associates with tubulin during mitosis. Cell Motil. Cytoskeleton. 2003, 56, 79–93. [Google Scholar] [CrossRef]
- Lin, M.; Hu, J.; Liu, T.; Li, J.; Chen, B.; Chen, X. Knockdown of nucleophosmin by RNA interference reverses multidrug resistance in resistant leukemic HL-60 cells. Immunobiology 2013, 218, 1147–1154. [Google Scholar] [CrossRef]
- Erlejman, A.G.; Lagadari, M.; Toneatto, J.; Piwien-Pilipuk, G.; Galigniana, M.D. Regulatory role of the 90-kDa-heat-shock protein (Hsp90) and associated factors on gene expression. Biochim. Biophys. Acta 2014, 1839, 71–87. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.F.; Tsai, Y.T.; Lai, Y.H.; Lin, C.C.; Chou, H.C.; Kuo, W.H.; Ko, M.L.; Wei, Y.S.; Wang, Y.S.; Lin, M.W.; et al. Proteomic analysis of Antrodia Cinnamomea-induced ER stress in liver cancer cells. J. Pharm. Biomed. Anal. 2020, 187, 113142. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Johnson, M.; Zhou, J.; Zhao, Y.; Kamal, M.A.; Qu, X. Antrodia cinnamomea Inhibits Growth and Migration of Lung Cancer Cells through Regulating p53-Bcl2 and MMPs Pathways. Am. J. Chin. Med. 2020, 48, 1941–1953. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.S.; Lin, Y.Y.; Yang, Y.H.; Lin, C.L.; Kuan, F.C.; Lu, C.N.; Chang, G.H.; Tsai, M.S.; Hsu, C.M.; Yeh, R.A.; et al. Antrodia cinnamomea extract inhibits the proliferation of tamoxifen-resistant breast cancer cells through apoptosis and skp2/microRNAs pathway. BMC Complement. Altern. Med. 2018, 18, 152. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Wang, J.; Li, L.; Hu, W.; Qu, Y.; Ding, Y.; Meng, L.; Teng, L.; Wang, D. Hepatoprotective Effects of Antrodia cinnamomea: The Modulation of Oxidative Stress Signaling in a Mouse Model of Alcohol-Induced Acute Liver Injury. Oxid. Med. Cell Longev. 2017, 2017, 7841823. [Google Scholar] [CrossRef] [Green Version]
- Timms, J.F.; Cramer, R. Difference gel electrophoresis. Proteomics 2008, 8, 4886–4897. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.C.; Chung, I.C.; Hsueh, C.; Tsang, N.M.; Chi, L.M.; Liang, Y.; Chen, C.C.; Wang, L.J.; Chang, Y.S. The antiapoptotic protein, FLIP, is regulated by heterogeneous nuclear ribonucleoprotein K and correlates with poor overall survival of nasopharyngeal carcinoma patients. Cell Death Differ. 2010, 17, 1463–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shenton, D.; Grant, C.M. Protein S-thiolation targets glycolysis and protein synthesis in response to oxidative stress in the yeast Saccharomyces cerevisiae. Biochem. J. 2003, 374 Pt 2, 513–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weeks, M.E.; Sinclair, J.; Butt, A.; Chung, Y.L.; Worthington, J.L.; Wilkinson, C.R.; Griffiths, J.; Jones, N.; Waterfield, M.D.; Timms, J.F. A parallel proteomic and metabolomic analysis of the hydrogen peroxide- and Sty1p-dependent stress response in Schizosaccharomyces pombe. Proteomics 2006, 6, 2772–2796. [Google Scholar] [CrossRef]
- Chen, J.N.; Wang, Y.T.; Wu, J.S. A glycoprotein extracted from golden oyster mushroom Pleurotus citrinopileatus exhibiting growth inhibitory effect against U937 leukemia cells. J. Agric. Food Chem. 2009, 57, 6706–6711. [Google Scholar] [CrossRef]
- Kang, M.A.; Jeon, Y.K.; Nam, M.J. Auricularia auricula increases an apoptosis in human hepatocellular carcinoma cells via a regulation of the peroxiredoxin1. J. Food Biochem. 2020, 44, e13373. [Google Scholar] [CrossRef]
- Cong, N.; Huang, W.; Yuan, J.P.; Li, G.Z.; Zhai, G.S.; Li, B.S. Peroxiredoxin1 promotes cell proliferation, migration and invasion of colorectal cancer via p38MAPK signaling. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 1922–1928. [Google Scholar] [CrossRef]
- Li, Y.; Maher, P.; Schubert, D. A role for 12-lipoxygenase in nerve cell death caused by glutathione depletion. Neuron 1997, 19, 453–463. [Google Scholar] [CrossRef] [Green Version]
- Kan, L.; Cui, D.; Chai, Y.; Ma, L.; Li, X.; Zhao, M. TMT-based quantitative proteomic analysis of antitumor mechanism of Sporisorium reilianum polysaccharide WM-NP-60 against HCT116 cells. Int. J. Biol. Macromol. 2020, 165 Pt B, 1755–1764. [Google Scholar] [CrossRef]
- Kan, L.; Chai, Y.; Li, X.; Zhao, M. Structural analysis and potential anti-tumor activity of Sporisorium reilianum (Fries) polysaccharide. Int. J. Biol. Macromol. 2020, 153, 986–994. [Google Scholar] [CrossRef] [PubMed]
- Isenberg, J.S.; Qin, Y.; Maxhimer, J.B.; Sipes, J.M.; Despres, D.; Schnermann, J.; Frazier, W.A.; Roberts, D.D. Thrombospondin-1 and CD47 regulate blood pressure and cardiac responses to vasoactive stress. Matrix Biol. J. Int. Soc. Matrix Biol. 2009, 28, 110–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Wang, J.; Zhang, C.; Lin, C.; Zhang, J.; Zhang, W.; Zhang, W.; Lu, Y.; Zheng, L.; Li, X. Circular RNA circITGA7 inhibits colorectal cancer growth and metastasis by modulating the Ras pathway and upregulating transcription of its host gene ITGA7. J. Pathol. 2018, 246, 166–179. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.T.; Huang, Z.J.; Chang, H.M. Proteomic analysis of human leukemic U937 cells incubated with conditioned medium of mononuclear cells stimulated by proteins from dietary mushroom of Agrocybe aegerita. J. Proteome Res. 2004, 3, 890–896. [Google Scholar] [CrossRef]
- Ou, H.T.; Shieh, C.J.; Chen, J.Y.; Chang, H.M. The antiproliferative and differentiating effects of human leukemic U937 cells are mediated by cytokines from activated mononuclear cells by dietary mushrooms. J. Agric. Food Chem. 2005, 53, 300–305. [Google Scholar] [CrossRef]
- Summerell, B.A.; Laurence, M.H.; Liew, E.C.Y.; Leslie, J.F. Biogeography and phylogeography of Fusarium: A review. Fungal Divers. 2010, 44, 3–13. [Google Scholar] [CrossRef]
- Xie, Y.; Li, S.; Sun, L.; Liu, S.; Wang, F.; Wen, B.; Sun, L.; Fang, X.; Chai, Y.; Cao, H.; et al. Fungal Immunomodulatory Protein from Nectria haematococca Suppresses Growth of Human Lung Adenocarcinoma by Inhibiting the PI3K/Akt Pathway. Int. J. Mol. Sci. 2018, 19, 3429. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.T.; Lin, T.Y.; Hsu, H.Y.; Sheu, F.; Ho, C.M.; Chen, E.I. Ling Zhi-8 mediates p53-dependent growth arrest of lung cancer cells proliferation via the ribosomal protein S7-MDM2-p53 pathway. Carcinogenesis 2011, 32, 1890–1896. [Google Scholar] [CrossRef] [Green Version]
- Ko, J.L.; Hsu, C.I.; Lin, R.H.; Kao, C.L.; Lin, J.Y. A new fungal immunomodulatory protein, FIP-fve isolated from the edible mushroom, Flammulina velutipes and its complete amino acid sequence. Eur. J. Biochem. 1995, 228, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Nie, Y.; Ding, Y.; Shi, L.; Tang, X. Recombinant expression of a novel fungal immunomodulatory protein with human tumor cell antiproliferative activity from Nectria haematococca. Int. J. Mol. Sci. 2014, 15, 17751–17764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsin, I.L.; Ou, C.C.; Wu, T.C.; Jan, M.S.; Wu, M.F.; Chiu, L.Y.; Lue, K.H.; Ko, J.L. GMI, an immunomodulatory protein from Ganoderma microsporum, induces autophagy in non-small cell lung cancer cells. Autophagy 2011, 7, 873–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ou, C.C.; Hsiao, Y.M.; Wang, W.H.; Ko, J.L.; Lin, M.Y. Stability of fungal immunomodulatory protein, FIP-gts and FIP-fve, in IFN-g production. Food Agric. Immunol. 2009, 20, 319–332. [Google Scholar] [CrossRef] [Green Version]
- Mahmood, R.I.; Abbass, A.K.; Razali, N.; Al-Saffar, A.Z.; Al-Obaidi, J.R. Protein profile of MCF-7 breast cancer cell line treated with lectin delivered by CaCO3NPs revealed changes in molecular chaperones, cytoskeleton, and membrane-associated proteins. Int. J. Biol. Macromol. 2021, 184, 636–647. [Google Scholar] [CrossRef]
- Hassan, M.A.; Rouf, R.; Tiralongo, E.; May, T.W.; Tiralongo, J. Mushroom lectins: Specificity, structure and bioactivity relevant to human disease. Int. J. Mol. Sci. 2015, 16, 7802–7838. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Shen, J.; Wang, J.; Yang, X.; Dong, S.; Lu, S. Nanoparticle-Based Drug Delivery System: A Patient-Friendly Chemotherapy for Oncology. Dose-response: A publication of International Hormesis Society. Dose-Response 2020, 18, 1559325820936161. [Google Scholar] [CrossRef]
- Nakanishi, M.; Shimada, M.; Niida, H. Genetic instability in cancer cells by impaired cell cycle checkpoints. Cancer Sci. 2006, 97, 984–989. [Google Scholar] [CrossRef]
- McKiernan, E.; McDermott, E.W.; Evoy, D.; Crown, J.; Duffy, M.J. The role of S100 genes in breast cancer progression. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2011, 32, 441–450. [Google Scholar] [CrossRef]
- Choi, J.; Kim, D.I.; Kim, J.; Kim, B.H.; Kim, A. Hornerin Is Involved in Breast Cancer Progression. J. Breast Cancer 2016, 19, 142–147. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.; Lee, D.H. Emerging roles of protein disulfide isomerase in cancer. BMB Rep. 2017, 50, 401–410. [Google Scholar] [CrossRef] [Green Version]
- Thongwatchara, P.; Promwikorn, W.; Srisomsap, C.; Chokchaichamnankit, D.; Boonyaphiphat, P.; Thongsuksai, P. Differential protein expression in primary breast cancer and matched axillary node metastasis. Oncol. Rep. 2011, 26, 185–191. [Google Scholar] [CrossRef]
- Luo, B.; Lee, A.S. The critical roles of endoplasmic reticulum chaperones and unfolded protein response in tumorigenesis and anticancer therapies. Oncogene 2013, 32, 805–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.; Si, M.L.; Wu, H.; Mo, Y.Y. MicroRNA-21 targets the tumor suppressor gene tropomyosin 1 (TPM1). J. Biol. Chem. 2007, 282, 14328–14336. [Google Scholar] [CrossRef] [Green Version]
- Sharma, M.; Ownbey, R.T.; Sharma, M.C. Breast cancer cell surface annexin II induces cell migration and neoangiogenesis via tPA dependent plasmin generation. Exp. Mol. Pathol. 2010, 88, 278–286. [Google Scholar] [CrossRef]
- Stehle, G.; Sinn, H.; Wunder, A.; Schrenk, H.H.; Stewart, J.C.; Hartung, G.; Maier-Borst, W.; Heene, D.L. Plasma protein (albumin) catabolism by the tumor itself--implications for tumor metabolism and the genesis of cachexia. Crit. Rev. Oncol./Hematol. 1997, 26, 77–100. [Google Scholar] [CrossRef]
- Wang, L.; Rubinstein, R.; Lines, J.L.; Wasiuk, A.; Ahonen, C.; Guo, Y.; Lu, L.F.; Gondek, D.; Wang, Y.; Fava, R.A.; et al. VISTA, a novel mouse Ig superfamily ligand that negatively regulates T cell responses. J. Exp. Med. 2011, 208, 577–592. [Google Scholar] [CrossRef]
- Zhou, H.; Zhang, Y.; Fu, Y.; Chan, L.; Lee, A.S. Novel mechanism of anti-apoptotic function of 78-kDa glucose-regulated protein (GRP78): Endocrine resistance factor in breast cancer, through release of B-cell lymphoma 2 (BCL-2) from BCL-2-interacting killer (BIK). J. Biol. Chem. 2011, 286, 25687–25696. [Google Scholar] [CrossRef] [Green Version]
- Boye, K.; Maelandsmo, G.M. S100A4 and metastasis: A small actor playing many roles. Am. J. Pathol. 2010, 176, 528–535. [Google Scholar] [CrossRef]
- Weete, J.D.; Abril, M.; Blackwell, M. Phylogenetic distribution of fungal sterols. PLoS ONE 2010, 5, e10899. [Google Scholar] [CrossRef] [Green Version]
- Kuo, C.; Hsieh, C.; Lin, W. Proteomic response of LAP-activated RAW 264.7 macrophages to the anti-inflammatory property of fungal ergosterol. Food Chem. 2011, 126, 207–212. [Google Scholar] [CrossRef]
- Kobori, M.; Yoshida, M.; Ohnishi-Kameyama, M.; Shinmoto, H. Ergosterol peroxide from an edible mushroom suppresses inflammatory responses in RAW264.7 macrophages and growth of HT29 colon adenocarcinoma cells. Br. J. Pharmacol. 2007, 150, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Pratt, W.B.; Toft, D.O. Regulation of signaling protein function and trafficking by the hsp90/hsp70-based chaperone machinery. Exp. Biol. Med. 2003, 228, 111–133. [Google Scholar] [CrossRef] [PubMed]
- Kurashige, S.; Akuzawa, Y.; Endo, F. Effects of Lentinus edodes, Grifola frondosa and Pleurotus ostreatus administration on cancer outbreak, and activities of macrophages and lymphocytes in mice treated with a carcinogen, N-butyl-N-butanolnitrosoamine. Immunopharmacol. Immunotoxicol. 1997, 19, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Shamtsyan, M.; Konusova, V.; Maksimova, Y.; Goloshchev, A.; Panchenko, A.; Simbirtsev, A.; Petrishchev, N.; Denisova, N. Immunomodulating and anti-tumor action of extracts of several mushrooms. J. Biotechnol. 2004, 113, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Jakopovich, I. New dietary supplements from medicinal mushrooms: Dr Myko San—A registration report. Int. J. Med. Mushrooms 2011, 13, 307–313. [Google Scholar] [CrossRef]
- Ivanković, S.; Hiršl, N.; Jakopović, I.; Jurin, M. The Influence of Medicinal Mushroom Preparations on Mouse Tumors. Int. J. Med. Mushrooms 2004, 6, 107–116. [Google Scholar] [CrossRef]
- Jakopovic, B.; Horvatić, A.; Klobučar, M.; Gelemanović, A.; Grbčić, P.; Oršolić, N.; Jakopovich, I.; Kraljević Pavelić, S. Treatment With Medicinal Mushroom Extract Mixture Inhibits Translation and Reprograms Metabolism in Advanced Colorectal Cancer Animal Model as Evidenced by Tandem Mass Tags Proteomics Analysis. Front. Pharmacol. 2020, 11, 1202. [Google Scholar] [CrossRef]
- Nair, A.B.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Driscoll, J.J.; Rixe, O. Overall survival: Still the gold standard: Why overall survival remains the definitive end point in cancer clinical trials. Cancer J. 2009, 15, 401–405. [Google Scholar] [CrossRef]
- Wolmark, N.; Fisher, B.; Wieand, H.S. The prognostic value of the modifications of the Dukes’ C class of colorectal cancer. An analysis of the NSABP clinical trials. Ann. Surg. 1986, 203, 115–122. [Google Scholar] [CrossRef]
- Wu, Y.; Deng, Z.; Wang, H.; Ma, W.; Zhou, C.; Zhang, S. Repeated cycles of 5-fluorouracil chemotherapy impaired anti-tumor functions of cytotoxic T cells in a CT26 tumor-bearing mouse model. BMC Immunol. 2016, 17, 29. [Google Scholar] [CrossRef]
- Jakopovic, B.; Oršolić, N.; Kraljević Pavelić, S. Antitumor, Immunomodulatory and Antiangiogenic Efficacy of Medicinal Mushroom Extract Mixtures in Advanced Colorectal Cancer Animal Model. Molecules 2020, 25, 5005. [Google Scholar] [CrossRef] [PubMed]
- Provenzani, A.; Fronza, R.; Loreni, F.; Pascale, A.; Amadio, M.; Quattrone, A. Global alterations in mRNA polysomal recruitment in a cell model of colorectal cancer progression to metastasis. Carcinogenesis 2006, 27, 1323–1333. [Google Scholar] [CrossRef] [Green Version]
- Chauvin, A.; Wang, C.S.; Geha, S.; Garde-Granger, P.; Mathieu, A.A.; Lacasse, V.; Boisvert, F.M. The response to neoadjuvant chemoradiotherapy with 5-fluorouracil in locally advanced rectal cancer patients: A predictive proteomic signature. Clin. Proteom. 2018, 15, 16. [Google Scholar] [CrossRef] [PubMed]
- Peng, F.; Huang, Y.; Li, M.Y.; Li, G.Q.; Huang, H.C.; Guan, R.; Chen, Z.C.; Liang, S.P.; Chen, Y.H. Dissecting characteristics and dynamics of differentially expressed proteins during multistage carcinogenesis of human colorectal cancer. World J. Gastroenterol. 2016, 22, 4515–4528. [Google Scholar] [CrossRef]
- Wielenga, M.; Colak, S.; Heijmans, J.; van Lidth de Jeude, J.F.; Rodermond, H.M.; Paton, J.C.; Paton, A.W.; Vermeulen, L.; Medema, J.P.; van den Brink, G.R. ER-Stress-Induced Differentiation Sensitizes Colon Cancer Stem Cells to Chemotherapy. Cell Rep. 2015, 13, 489–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aiderus, A.; Black, M.A.; Dunbier, A.K. Fatty acid oxidation is associated with proliferation and prognosis in breast and other cancers. BMC Cancer 2018, 18, 805. [Google Scholar] [CrossRef]
- Schmidt, C.; Sciacovelli, M.; Frezza, C. Fumarate hydratase in cancer: A multifaceted tumour suppressor. Semin. Cell Dev. Biol. 2020, 98, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.L.; He, Y.M. Network pharmacology analysis of the anti-cancer pharmacological mechanisms of Ganoderma lucidum extract with experimental support using Hepa1-6-bearing C57 BL/6 mice. J. Ethnopharmacol. 2018, 210, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Lee, W.Y.; Jung, K.; Kwon, Y.S.; Kim, D.; Hwang, G.S.; Kim, C.E.; Lee, S.; Kang, K.S. The Inhibitory Effect of Cordycepin on the Proliferation of MCF-7 Breast Cancer Cells, and its Mechanism: An Investigation Using Network Pharmacology-Based Analysis. Biomolecules 2019, 9, 414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Y.; Qiu, P.; Zhu, R.; Zhao, L.; Zhang, P.; Wang, Y.; Li, C.; Chai, K.; Shou, D.; Zhao, H. A Combined Phytochemistry and Network Pharmacology Approach to Reveal the Potential Antitumor Effective Substances and Mechanism of Phellinus igniarius. Front. Pharmacol. 2019, 10, 266. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mushroom Species | Compound | Type of Experiment (Tumor Model) | Proteomic Methodology | Validation | Reference |
|---|---|---|---|---|---|
| Ganoderma lucidum | Characterized Ganoderma lucidum spores (GL-SP) | In vitro Splenic mononuclear cells (MNCs) | 2-DE followed by MALDI-MS/MS | No | Ma, C., et al., 2008 |
| Ganoderma lucidum | Characterized Ganoderma lucidum spores (GL-SP) | In vivo Thymus lymphoid cells | 2-DE followed by MALDI-TOF MS/MS | Western blotting | Ma, C., et al., 2009 |
| Ganoderma lucidum | Characterized Ganoderma lucidum polysaccharide peptide (GlPS) from the fruiting body | In vivo Murine sarcoma S180 model | 2-DE followed by ESI-Q-TOF-MS/MS | Western blotting and RT-PCR | Li, Y.B., et al., 2008 |
| Ganoderma lucidum | Characterized Ganoderma lucidum polysaccharide peptide (GL-pp) | In vivo Murine melanoma (B16-F10-luc-G5) model | LC-MS/MS followed by bioinformatic analysis | No | Xian, H., et al., 2021 |
| Ganoderma lucidum, Phellinus linteus, Auricularia auricula | Polysaccharides from Ganoderma lucidum (GL), Phellinus linteus (PL), Auricularia auricula (AA) studied separately | In vitro Human HepG2 (hepatocellular carcinoma) cell line | 2-DE followed by MALDI-TOF-MS and bioinformatic analysis | Western blotting and RT-PCR | Chai, Y., et al., 2016 |
| Ganoderma lucidum | Fungal immunomodulatory protein Ling Zhi-8 (LZ-8) | In vivo Murine Lewis lung carcinoma cell line (LLC1) | 2-DE followed by LC-MS/MS and bioinformatic analysis | Western blotting | Lin, T.Y., et al., 2021 |
| Ganoderma leucocontextum | GL22 triterpene | Huh7.5 liver cancer cell line in vitro and in vivo (xenograft) mouse model | LC-MS/MS followed by bioinformatic analysis | Western blotting | Liu, G., et al., 2018 |
| Ganoderma lucidum | Ganoderic acid D | In vitro HeLa human cervical carcinoma cells | 2-DE followed by MALDI-TOF MS/MS | Western blotting | Yue, Q.X., et al., 2008 |
| Ganoderma lucidum | ganoderic acid F (GAF), ganoderic acid K (GAK), ganoderic acid B (GAB), ganoderic acid D (GAD) and ganoderic acid AM1 (GAAM1) | In vitro HeLa human cervical carcinoma cells | 2-DE followed by MALDI-TOF MS/MS | Western blotting | Yue, Q.X., et al., 2010 |
| Ganoderma lucidum | Characterized mixture of Ganoderma lucidum triterpenes (GTS) | In vitro HeLa human cervical carcinoma cells | 2-DE followed by MALDI-TOF MS/MS | Western blotting | Yue, Q.X., et al., 2008 |
| Lentinus edodes | MPSSS polysaccharide | In vitro Prostate CAFs (cancer associated fibroblasts) and PC-3 (human prostate cell line) | Tandem mass tag (TMT) labeling followed by LC-MS/MS and bioinformatic analysis | Western blotting | Zhang, T., et al., 2021 |
| Lentinus edodes | Lentinan | In vivo Murine H22 hepatoma cells | LC-MS/MS | No | Wang, Y., et al., 2017 |
| Lentinus edodes | Lentinan | In vivo ascites and solid H22 liver cancer models | LC-MS/MS | No | Yang, X., et al., 2020 |
| Lentinus edodes | Lentinan | In vivo Murine H22 hepatoma cells | HPLC-MS/nMS | ELISA | Wang, W., et al., 2021 |
| Lentinus edodes | Lentinan (LNT)-functionalized selenium nanoparticles | In vivo Ovarian cancer (OVCAR-3) and Erlich ascites carcinoma (EAC) models | LC-MS/MS followed by bioinformatic analysis | Western blotting | Liu, H.J., et al., 2020 |
| Cordyceps militaris | Cordyceps militaris (CM) fresh body or mycelia extract | In vitro Cisplatin-resistant A549 human lung adenocarcinoma cells | Protein antibody microarray | Western blotting | Jeong, M.K., et al., 2019 |
| Cordyceps sinensis | Characterized Cordyceps sinensis water extract (WECS) | In vitro and in vivo 4T1 breast cancer model | Protein array | No | Cai, H., et al., 2018 |
| Cordyceps sinensis | Characterized Cordyceps sinensis ethanol extract | In vivo DEN (diethylnitrosamine)-induced hepatocellular carcinoma | 2-DE followed by MALDI-TOF MS and bioinformatic analysis | Western blotting | Wang, P.W., et al., 2016 |
| Cordyceps cicadae | Water extract of Cordyceps cicadae | In vitro Human MHCC97H hepatocellular carcinoma cells | 2-DE followed by MALDI-TOF/TOF MS | Western blotting | Wang, H., et al., 2014 |
| Pleurotus sajor-caju | n-hexane, chloroform, ethyl acetate, ethanol and ethanol/water Pleurotus sajor-caju fruiting body extract | In vitro Human colorectal adenocarcinoma HCT-116wt, -Bax, -p21 cells | Protein array | Western blotting | Finimundy, T.C., et al., 2018 |
| Pleurotus eryngii | PEP 1b, a novel immunoregulatory protein | Raw 265.7 macrophage cells | *iTRAQ labeling followed by LC-MS/MS and bioinformatic analysis | Western blotting | Ma, N., et al., 2020 |
| Trametes versicolor | Polysaccharopeptide (PSP) | Ex vivo Human T lymphocytes | 2-DE followed by MALDI-TOF MS | Western blotting | Lee, C.L., et al., 2007 |
| Hericium erinaceus | Erinacine A | In vitro Human MKN28 and TSGH9201 human gastric carcinoma | 2-DE followed by MALDI-TOF/TOF | Western blotting | Kuo, H.C., et al., 2017 |
| Hericium erinaceus | Erinacine A | In vitro (DLD-1 and HCT-116) and in vivo (HCT-116 xenograft) human colorectal carcinoma cells | 2-DE followed by MALDI-TOF/TOF | Western blotting | Lee, K.C., et al., 2017 |
| Phellinus linteus | Proteoglycan P1 | In vitro and in vivo (xenograft) HepG2 human hepatocellular carcinoma cells | 2-DE followed by MALDI-TOF/TOF | RT-PCR and Western blotting | Li, Y.G., et al., 2013 |
| Grifola frondosa | Glycoprotein GFG-3a | In vitro Human gastric cancer cell line SGC-7901 | 2-DE followed by MALDI-TOF-MS | qRT-PCR and Western blotting | Cui, F., et al., 2016 |
| Antrodia cinnamomea | Antrodia cinnamomea fruiting body ethanolic extract | In vitro Liver cancer (HepG2 and C3A) and normal liver (Chang’s) cell lines | 2D-DIGE followed by MALDI-TOF MS and bioinformatic analysis | Immunoblot analysis | Chen, J.F., et al., 2020 |
| Auricularia auricula | Three types of Auricularia auricula isolates (FD, BS, BT) | In vitro hepatocellular cancer cell lines (HCT-15, huh-7, SK-MEL-5, SNU-213, and SNU-484) | 2-DE-GE followed by MALDI-TOF-MS | RT-PCT and Western blotting | Kang, M.A., et al., 2020 |
| Sporisorium reilianum | Sporisorium reilianum polysaccharide WM-NP-60 | In vitro Human colorectal adenocarcinoma HCT-116 cells | Tandem mass tag (TMT) labeling followed by nano-HPLC-MS/MS and bioinformatic analysis | qRT-PCR and Western blotting | Kan, L., et al., 2020 |
| Agrocybe aegerita | Fraction 2 proteins of Agrocybe aegerita (AA-f2-MNC-CM) | In vitro Human leukemic U937 cells | 2-DE followed by MALDI-TOF-MS | No | Wang, Y.T., et al., 2004 |
| Nectria haematococca | Fungal immunomodulatory protein (FIP) from Nectria hematococca (FIP-nha) | Human lung adenocarcinoma (A549 and NCI-H2347) calls in vitro and A549 in vivo | iTRAQ (isobaric tag for relative and absolute quantitation) followed by quadrupole-orbitrap MS and bioinformatic analysis | Western blotting | Xie, Y., et al., 2018 |
| Agaricus bisporus | ABL-Agaricus bisporus lectin conjugated with CaCO3 nanoparticles (ABL- CaCO3NPs) | In vitro Human breast cancer cells (MCF-7) | 2-DE followed by LC-MS and bioinformatic analysis | No | Mahmood, R.I., et al., 2021 |
| *NA | Ergosterol | In vitro Murine macrophage cell line RAW 264.7 | 2-DE followed by MALDI-TOF/TOF-MS | Western blotting | Kuo, C., et al., 2011 |
| Lentinus edodes, Ganoderma lucidum, Agaricus brasiliensis (=blazei ss. Heinem.), Grifola frondosa, Pleurotus ostreatus, and Trametes versicolor combined in a commercial medicinal mushroom extract blend Agarikon.1 | Various | In vivo Murine colorectal carcinoma CT26.WT model | Tandem mass tag (TMT) labeling followed by LC-MS/MS and bioinformatic analysis | Western blotting | Jakopovic, B., et al., 2020 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jakopovic, B.; Oršolić, N.; Jakopovich, I. Proteomic Research on the Antitumor Properties of Medicinal Mushrooms. Molecules 2021, 26, 6708. https://doi.org/10.3390/molecules26216708
Jakopovic B, Oršolić N, Jakopovich I. Proteomic Research on the Antitumor Properties of Medicinal Mushrooms. Molecules. 2021; 26(21):6708. https://doi.org/10.3390/molecules26216708
Chicago/Turabian StyleJakopovic, Boris, Nada Oršolić, and Ivan Jakopovich. 2021. "Proteomic Research on the Antitumor Properties of Medicinal Mushrooms" Molecules 26, no. 21: 6708. https://doi.org/10.3390/molecules26216708
APA StyleJakopovic, B., Oršolić, N., & Jakopovich, I. (2021). Proteomic Research on the Antitumor Properties of Medicinal Mushrooms. Molecules, 26(21), 6708. https://doi.org/10.3390/molecules26216708