
Metabolomic Characterization of Cerebrospinal Fluid from Intracranial Bacterial Infection Pediatric Patients: A Pilot Study




Abstract
:1. Introduction
2. Results and Discussions
2.1. Untargeted Analysis for Metabolic Profiling of CSF from Patients with Intracranial Bacterial Infection
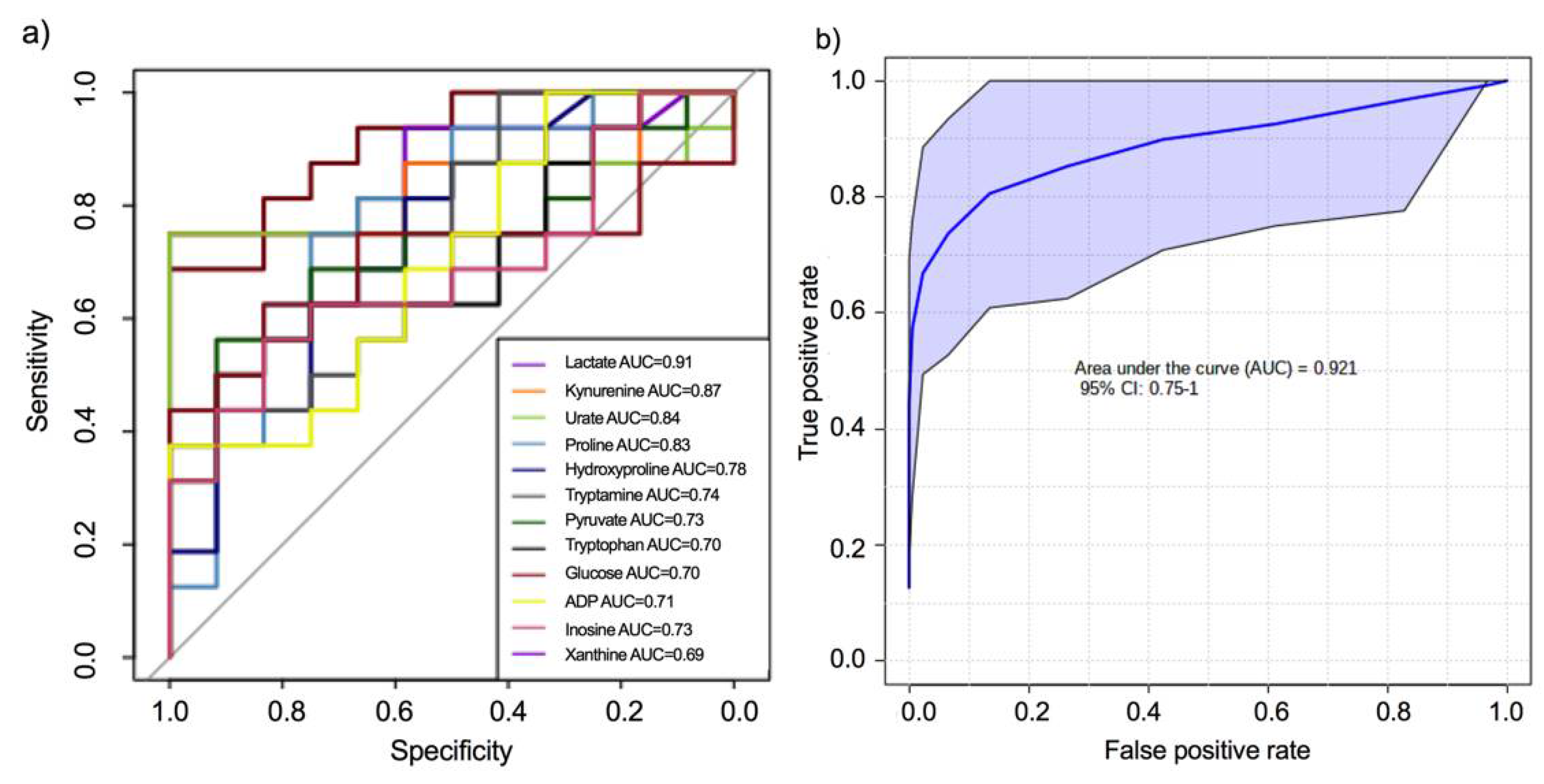
2.2. Targeted Metabolomic Analysis for Semi-Quantitative Verification and Pathway Analysis
3. Materials and Methods
3.1. CSF Sample Collection and Patient Information
3.2. Chemicals and Reagents
3.3. Metabolite Extraction
3.4. LC-MS/MS Analysis
3.5. Statistical Analysis
3.6. Identification and Pathway Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Spector, R.; Robert Snodgrass, S.; Johanson, C.E. A Balanced View of the Cerebrospinal Fluid Composition and Functions: Focus on Adult Humans. Exp. Neurol. 2015, 273, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Kaneko, N.; Sawamoto, K. Go with the Flow: Cerebrospinal Fluid Flow Regulates Neural Stem Cell Proliferation. Cell Stem Cell 2018, 22, 783–784. [Google Scholar] [CrossRef] [Green Version]
- Wishart, D.S.; Lewis, M.J.; Morrissey, J.A.; Flegel, M.D.; Jeroncic, K.; Xiong, Y.; Cheng, D.; Eisner, R.; Gautam, B.; Tzur, D.; et al. The Human Cerebrospinal Fluid Metabolome. J. Chromatogr. B 2008, 871, 164–173. [Google Scholar] [CrossRef]
- Gallego, J.A.; Gordon, M.L.; Claycomb, K.; Bhatt, M.; Lencz, T.; Malhotra, A.K. In Vivo MicroRNA Detection and Quantitation in Cerebrospinal Fluid. J. Mol. Neurosci. 2012, 47, 243–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Guo, Z.; Zou, L.; Yang, Y.; Zhang, L.; Ji, N.; Shao, C.; Sun, W.; Wang, Y. A Comprehensive Map and Functional Annotation of the Normal Human Cerebrospinal Fluid Proteome. J. Proteom. 2015, 119, 90–99. [Google Scholar] [CrossRef]
- Macron, C.; Lane, L.; Núñez Galindo, A.; Dayon, L. Deep Dive on the Proteome of Human Cerebrospinal Fluid: A Valuable Data Resource for Biomarker Discovery and Missing Protein Identification. J. Proteome Res. 2018, 17, 4113–4126. [Google Scholar] [CrossRef]
- Vitorino, R. Digging Deep into Peptidomics Applied to Body Fluids. Proteomics 2018, 18, 1700401. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vázquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. HMDB 4.0: The Human Metabolome Database for 2018. Nucleic Acids Res. 2018, 46, D608–D617. [Google Scholar] [CrossRef] [PubMed]
- Blennow, K.; Zetterberg, H. Biomarkers for Alzheimer’s Disease: Current Status and Prospects for the Future. J. Intern. Med. 2018, 284, 643–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Q.; Liu, J.; Quan, J.; Liu, W.; Tan, H.; Li, W. MicroRNAs as Potential Biomarkers for the Diagnosis of Glioma: A Systematic Review and Meta-analysis. Cancer Sci. 2018, 109, 2651–2659. [Google Scholar] [CrossRef] [Green Version]
- Agoston, D.V.; Shutes-David, A.; Peskind, E.R. Biofluid Biomarkers of Traumatic Brain Injury. Brain Inj. 2017, 31, 1195–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Kuzhiumparambil, U.; Bandodkar, S.; Dale, R.C.; Fu, S. Cerebrospinal Fluid Metabolomics: Detection of Neuroinflammation in Human Central Nervous System Disease. Clin. Transl. Immunol. 2021, 10, e1318. [Google Scholar] [CrossRef]
- Forrester, J.V.; McMenamin, P.G.; Dando, S.J. CNS Infection and Immune Privilege. Nat. Rev. Neurosci. 2018, 19, 655–671. [Google Scholar] [CrossRef]
- Akdis, C.A. Does the Epithelial Barrier Hypothesis Explain the Increase in Allergy, Autoimmunity and Other Chronic Conditions? Nat. Rev. Immunol. 2021, 21, 739–751. [Google Scholar] [CrossRef]
- de Morais, S.D.; Kak, G.; Menousek, J.P.; Kielian, T. Immunopathogenesis of Craniotomy Infection and Niche-Specific Immune Responses to Biofilm. Front. Immunol. 2021, 12, 625467. [Google Scholar] [CrossRef] [PubMed]
- Imran, D.; Estiasari, R.; Maharani, K.; Sucipto; Lestari, D.C.; Yunus, R.E.; Yunihastuti, E.; Karyadi, T.H.; Oei, D.; Timan, I.S.; et al. Presentation, Etiology, and Outcome of Brain Infections in an Indonesian Hospital: A Cohort Study. Neurol. Clin. Pract. 2018, 8, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Letertre, M.P.M.; Dervilly, G.; Giraudeau, P. Combined Nuclear Magnetic Resonance Spectroscopy and Mass Spectrometry Approaches for Metabolomics. Anal. Chem. 2021, 93, 500–518. [Google Scholar] [CrossRef] [PubMed]
- Bakochi, A.; Mohanty, T.; Pyl, P.T.; Gueto-Tettay, C.A.; Malmström, L.; Linder, A.; Malmström, J. Cerebrospinal Fluid Proteome Maps Detect Pathogen-Specific Host Response Patterns in Meningitis. eLife 2021, 10, e64159. [Google Scholar] [CrossRef]
- van Laarhoven, A.; Dian, S.; Aguirre-Gamboa, R.; Avila-Pacheco, J.; Ricaño-Ponce, I.; Ruesen, C.; Annisa, J.; Koeken, V.A.C.M.; Chaidir, L.; Li, Y.; et al. Cerebral Tryptophan Metabolism and Outcome of Tuberculous Meningitis: An Observational Cohort Study. Lancet Infect. Dis. 2018, 18, 526–535. [Google Scholar] [CrossRef]
- Quist-Paulsen, E.; Aukrust, P.; Kran, A.-M.B.; Dunlop, O.; Ormaasen, V.; Stiksrud, B.; Midttun, Ø.; Ueland, T.; Ueland, P.M.; Mollnes, T.E.; et al. High Neopterin and IP-10 Levels in Cerebrospinal Fluid Are Associated with Neurotoxic Tryptophan Metabolites in Acute Central Nervous System Infections. J. Neuroinflamm. 2018, 15, 327. [Google Scholar] [CrossRef] [Green Version]
- Borah, J.; Dutta, P.; Khan, S.A.; Mahanta, J. A Comparison of Clinical Features of Japanese Encephalitis Virus Infection in the Adult and Pediatric Age Group with Acute Encephalitis Syndrome. J. Clin. Virol. 2011, 52, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.H.; Ivanisevic, J.; Siuzdak, G. Metabolomics: Beyond Biomarkers and towards Mechanisms. Nat. Rev. Mol. Cell Biol. 2016, 17, 451–459. [Google Scholar] [CrossRef] [Green Version]
- Wishart, D.S. Emerging Applications of Metabolomics in Drug Discovery and Precision Medicine. Nat. Rev. Drug Discov. 2016, 15, 473–484. [Google Scholar] [CrossRef]
- Rinschen, M.M.; Ivanisevic, J.; Giera, M.; Siuzdak, G. Identification of Bioactive Metabolites Using Activity Metabolomics. Nat. Rev. Mol. Cell Biol. 2019, 20, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Newgard, C.B. Metabolomics and Metabolic Diseases: Where Do We Stand? Cell Metab. 2017, 25, 43–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rampler, E.; Abiead, Y.E.; Schoeny, H.; Rusz, M.; Hildebrand, F.; Fitz, V.; Koellensperger, G. Recurrent Topics in Mass Spectrometry-Based Metabolomics and Lipidomics—Standardization, Coverage, and Throughput. Anal. Chem. 2021, 93, 519–545. [Google Scholar] [CrossRef] [PubMed]
- Cajka, T.; Fiehn, O. Toward Merging Untargeted and Targeted Methods in Mass Spectrometry-Based Metabolomics and Lipidomics. Anal. Chem. 2016, 88, 524–545. [Google Scholar] [CrossRef]
- Regenold, W.T.; Phatak, P.; Makley, M.J.; Stone, R.D.; Kling, M.A. Cerebrospinal Fluid Evidence of Increased Extra-Mitochondrial Glucose Metabolism Implicates Mitochondrial Dysfunction in Multiple Sclerosis Disease Progression. J. Neurol. Sci. 2008, 275, 106–112. [Google Scholar] [CrossRef] [Green Version]
- Mason, S.; Reinecke, C.J.; Solomons, R. Cerebrospinal Fluid Amino Acid Profiling of Pediatric Cases with Tuberculous Meningitis. Front. Neurosci. 2017, 11, 534. [Google Scholar] [CrossRef]
- Sühs, K.-W.; Novoselova, N.; Kuhn, M.; Seegers, L.; Kaever, V.; Müller-Vahl, K.; Trebst, C.; Skripuletz, T.; Stangel, M.; Pessler, F. Kynurenine Is a Cerebrospinal Fluid Biomarker for Bacterial and Viral Central Nervous System Infections. J. Infect. Dis. 2019, 220, 127–138. [Google Scholar] [CrossRef]
- van Zyl, C.D.W.; Loots, D.T.; Solomons, R.; van Reenen, M.; Mason, S. Metabolic Characterization of Tuberculous Meningitis in a South African Paediatric Population Using 1H NMR Metabolomics. J. Infect. 2020, 81, 743–752. [Google Scholar] [CrossRef]
- Peddie, B.A.; Wong-She, J.; Randall, K.; Lever, M.; Chambers, S.T. Osmoprotective Properties and Accumulation of Betaine Analogues by Staphylococcus Aureus. FEMS Microbiol. Lett. 1998, 160, 25–30. [Google Scholar] [CrossRef]
- Christen, S.; Bifrare, Y.-D.; Siegenthaler, C.; Leib, S.L.; Täuber, M.G. Marked Elevation in Cortical Urate and Xanthine Oxidoreductase Activity in Experimental Bacterial Meningitis. Brain Res. 2001, 900, 244–251. [Google Scholar] [CrossRef]
- de Menezes, C.C.; Dorneles, A.G.; Sperotto, R.L.; Duarte, M.M.F.; Schetinger, M.R.C.; Loro, V.L. Oxidative Stress in Cerebrospinal Fluid of Patients with Aseptic and Bacterial Meningitis. Neurochem. Res. 2009, 34, 1255–1260. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.-P.; Chen, G.-Y.; Chuang, T.-Y.; Huang, Y.-T.; Chang, H.-T.; Chen, Y.-F.; Liu, W.-L.; Chen, Y.-J.; Hsu, C.-L.; Huang, M.-T.; et al. Increased Plasma Acetylcarnitine in Sepsis Is Associated With Multiple Organ Dysfunction and Mortality: A Multicenter Cohort Study. Crit. Care Med. 2019, 47, 210–218. [Google Scholar] [CrossRef]
- Pohanka, M. Role of Oxidative Stress in Infectious Diseases. A Review. Folia Microbiol. 2013, 58, 503–513. [Google Scholar] [CrossRef]
- Blaise, B.J.; Correia, G.D.S.; Haggart, G.A.; Surowiec, I.; Sands, C.; Lewis, M.R.; Pearce, J.T.M.; Trygg, J.; Nicholson, J.K.; Holmes, E.; et al. Statistical Analysis in Metabolic Phenotyping. Nat. Protoc. 2021, 16, 4299–4326. [Google Scholar] [CrossRef]
- Nazir, M.; Wani, W.A.; Malik, M.A.; Mir, M.R.; Ashraf, Y.; Kawoosa, K.; Ali, S.W. Cerebrospinal Fluid Lactate: A Differential Biomarker for Bacterial and Viral Meningitis in Children. J. Pediatr. 2018, 94, 88–92. [Google Scholar] [CrossRef]
- Mándi, Y.; Vécsei, L. The Kynurenine System and Immunoregulation. J. Neural. Transm. 2012, 119, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Warburg, O. On the Origin of Cancer Cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef]
- Nolt, B.; Tu, F.; Wang, X.; Ha, T.; Winter, R.; Williams, D.L.; Li, C. Lactate and Immunosuppression in Sepsis. Shock 2018, 49, 120–125. [Google Scholar] [CrossRef]
- Kealy, J.; Murray, C.; Griffin, E.W.; Lopez-Rodriguez, A.B.; Healy, D.; Tortorelli, L.S.; Lowry, J.P.; Watne, L.O.; Cunningham, C. Acute Inflammation Alters Brain Energy Metabolism in Mice and Humans: Role in Suppressed Spontaneous Activity, Impaired Cognition, and Delirium. J. Neurosci. 2020, 40, 5681–5696. [Google Scholar] [CrossRef] [PubMed]
- Wlodarska, M.; Luo, C.; Kolde, R.; d’Hennezel, E.; Annand, J.W.; Heim, C.E.; Krastel, P.; Schmitt, E.K.; Omar, A.S.; Creasey, E.A.; et al. Indoleacrylic Acid Produced by Commensal Peptostreptococcus Species Suppresses Inflammation. Cell Host Microbe 2017, 22, 25–37.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovelace, M.D.; Varney, B.; Sundaram, G.; Lennon, M.J.; Lim, C.K.; Jacobs, K.; Guillemin, G.J.; Brew, B.J. Recent Evidence for an Expanded Role of the Kynurenine Pathway of Tryptophan Metabolism in Neurological Diseases. Neuropharmacology 2017, 112, 373–388. [Google Scholar] [CrossRef]
- Krishnan, N.; Dickman, M.B.; Becker, D.F. Proline Modulates the Intracellular Redox Environment and Protects Mammalian Cells against Oxidative Stress. Free Radic. Biol. Med. 2008, 44, 671–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgner, D.; Rockett, K.; Kwiatkowski, D. Nitric Oxide and Infectious Diseases. Arch. Dis. Child. 1999, 81, 185–188. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, M.; Wang, X.; Yi, H.; Xu, L.; Zhong, X.; Peng, F. Serum Uric Acid Levels in Patients with Infections of Central Nervous System. Acta Neurol. Belg. 2016, 116, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. Purinergic Signalling and Disorders of the Central Nervous System. Nat. Rev. Drug Discov. 2008, 7, 575–590. [Google Scholar] [CrossRef]
- Kuhn, M.; Sühs, K.-W.; Akmatov, M.K.; Klawonn, F.; Wang, J.; Skripuletz, T.; Kaever, V.; Stangel, M.; Pessler, F. Mass-Spectrometric Profiling of Cerebrospinal Fluid Reveals Metabolite Biomarkers for CNS Involvement in Varicella Zoster Virus Reactivation. J. Neuroinflamm. 2018, 15, 20. [Google Scholar] [CrossRef] [Green Version]
- Ratuszny, D.; Sühs, K.-W.; Novoselova, N.; Kuhn, M.; Kaever, V.; Skripuletz, T.; Pessler, F.; Stangel, M. Identification of Cerebrospinal Fluid Metabolites as Biomarkers for Enterovirus Meningitis. Int. J. Mol. Sci. 2019, 20, 337. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Möhn, N.; Al-Mekhlafi, A.; Schuchardt, S.; Skripuletz, T.; Sühs, W.; Pessler, F.; Stangel, M. Targeted Metabolomic Profiling of Cerebrospinal Fluid from Patients with Progressive Multifocal Leukoencephalopathy. PLoS ONE 2020, 15, e0242321. [Google Scholar] [CrossRef] [PubMed]
- Al-Mekhlafi, A.; Sühs, K.-W.; Schuchardt, S.; Kuhn, M.; Müller-Vahl, K.; Trebst, C.; Skripuletz, T.; Klawonn, F.; Stangel, M.; Pessler, F. Elevated Free Phosphatidylcholine Levels in Cerebrospinal Fluid Distinguish Bacterial from Viral CNS Infections. Cells 2021, 10, 1115. [Google Scholar] [CrossRef] [PubMed]
- Haughey, N.J.; Cutler, R.G.; Tamara, A.; McArthur, J.C.; Vargas, D.L.; Pardo, C.A.; Turchan, J.; Nath, A.; Mattson, M.P. Perturbation of Sphingolipid Metabolism and Ceramide Production in HIV-Dementia. Ann. Neurol. 2004, 55, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards More Transparent and Integrative Metabolomics Analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [Green Version]
- Robin, X.; Turck, N.; Hainard, A.; Tiberti, N.; Lisacek, F.; Sanchez, J.-C.; Müller, M. PROC: An Open-Source Package for R and S+ to Analyze and Compare ROC Curves. BMC Bioinform. 2011, 12, 77. [Google Scholar] [CrossRef]
- Shen, X.; Wang, R.; Xiong, X.; Yin, Y.; Cai, Y.; Ma, Z.; Liu, N.; Zhu, Z.-J. Metabolic Reaction Network-Based Recursive Metabolite Annotation for Untargeted Metabolomics. Nat. Commun. 2019, 10, 1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular Framework for Processing, Visualizing, and Analyzing Mass Spectrometry-Based Molecular Profile Data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Sato, Y. KEGG Mapper for Inferring Cellular Functions from Protein Sequences. Protein Sci. 2020, 29, 28–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Infection | Control |
|---|---|---|
| Age | 2.1 ± 4.4 years | 1.6 ± 2.2 years |
| Gender | 62.5% male | 66.7% male |
| CSF cell count (/L) | 274 ± 132 | 6.25 ± 8.07 |
| CSF glucose (mmol/L) | 1.96 ± 1.63 | 2.85 ± 0.32 |
| CSF protein (mg/L) | 997 ± 562 | 350 ± 37 |
| CSF lactate (mmol/L) | 3.45 ± 0.08 | 1.50 ± 0.33 |
| CSF bacterial isolates | 75% negative 25% S. epidermidis positive | 100% negative |
| Blood WBC (/L) | 17.06 ± 6.88 | 9.15 ± 3.01 |
| Blood N% | 51.0 ± 17.8% | 34.9 ± 14.0% |
| Blood CRP (mg/L) | 31.0 ± 37.8 | 5.75 ± 3.03 |
| Blood PCT (ng/mL) | 1.29 ± 0.79 | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Liu, Y.; Chen, R.; Qiao, L. Metabolomic Characterization of Cerebrospinal Fluid from Intracranial Bacterial Infection Pediatric Patients: A Pilot Study. Molecules 2021, 26, 6871. https://doi.org/10.3390/molecules26226871
Wang Y, Liu Y, Chen R, Qiao L. Metabolomic Characterization of Cerebrospinal Fluid from Intracranial Bacterial Infection Pediatric Patients: A Pilot Study. Molecules. 2021; 26(22):6871. https://doi.org/10.3390/molecules26226871
Chicago/Turabian StyleWang, Yiwen, Yu Liu, Ruoping Chen, and Liang Qiao. 2021. "Metabolomic Characterization of Cerebrospinal Fluid from Intracranial Bacterial Infection Pediatric Patients: A Pilot Study" Molecules 26, no. 22: 6871. https://doi.org/10.3390/molecules26226871
APA StyleWang, Y., Liu, Y., Chen, R., & Qiao, L. (2021). Metabolomic Characterization of Cerebrospinal Fluid from Intracranial Bacterial Infection Pediatric Patients: A Pilot Study. Molecules, 26(22), 6871. https://doi.org/10.3390/molecules26226871