
Stevioside Attenuates Insulin Resistance in Skeletal Muscle by Facilitating IR/IRS-1/Akt/GLUT 4 Signaling Pathways: An In Vivo and In Silico Approach

Abstract
:1. Introduction
2. Results
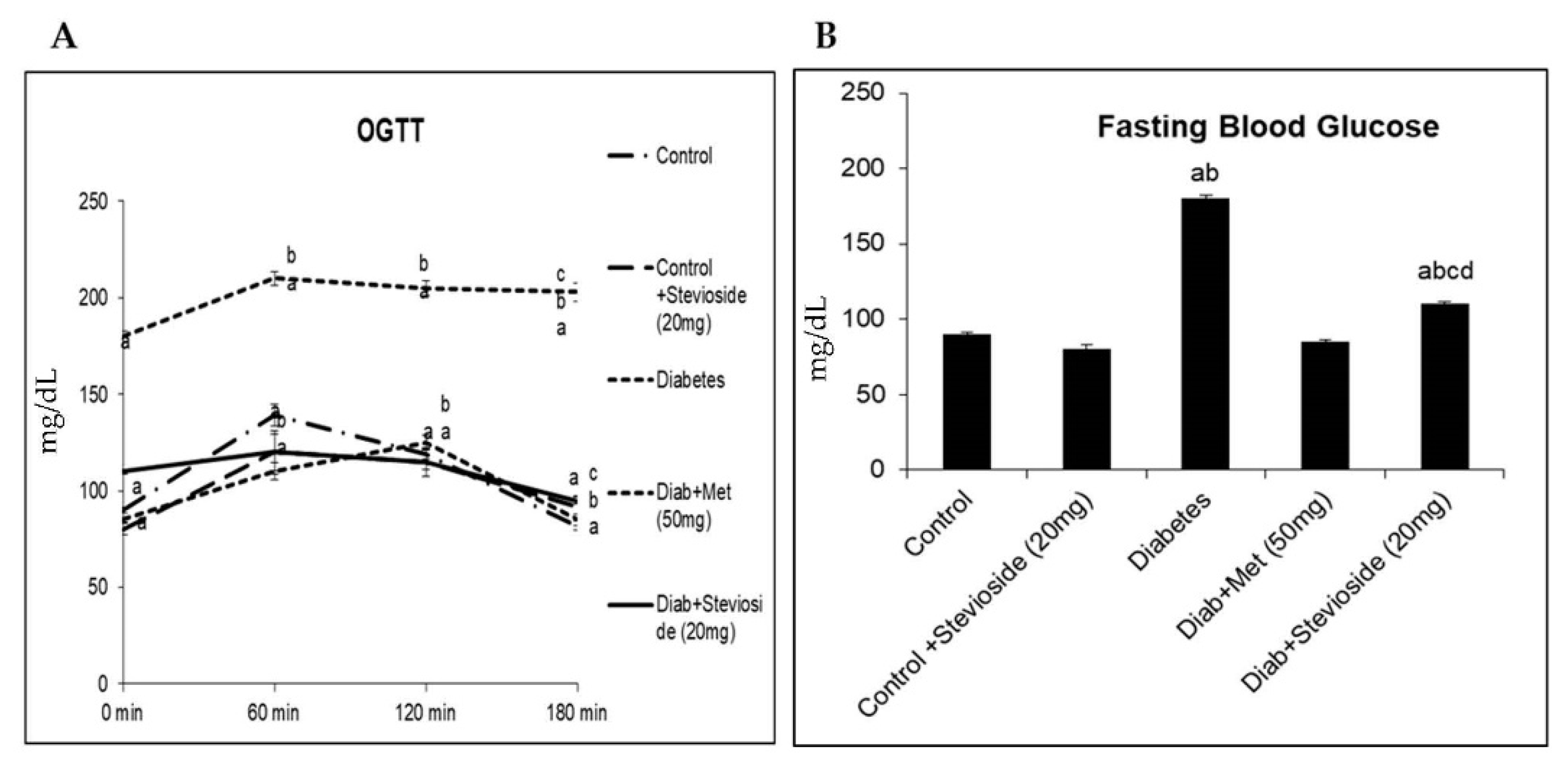
2.1. Stevioside Reduced FBG and Improved OGTT in T2DM Rats
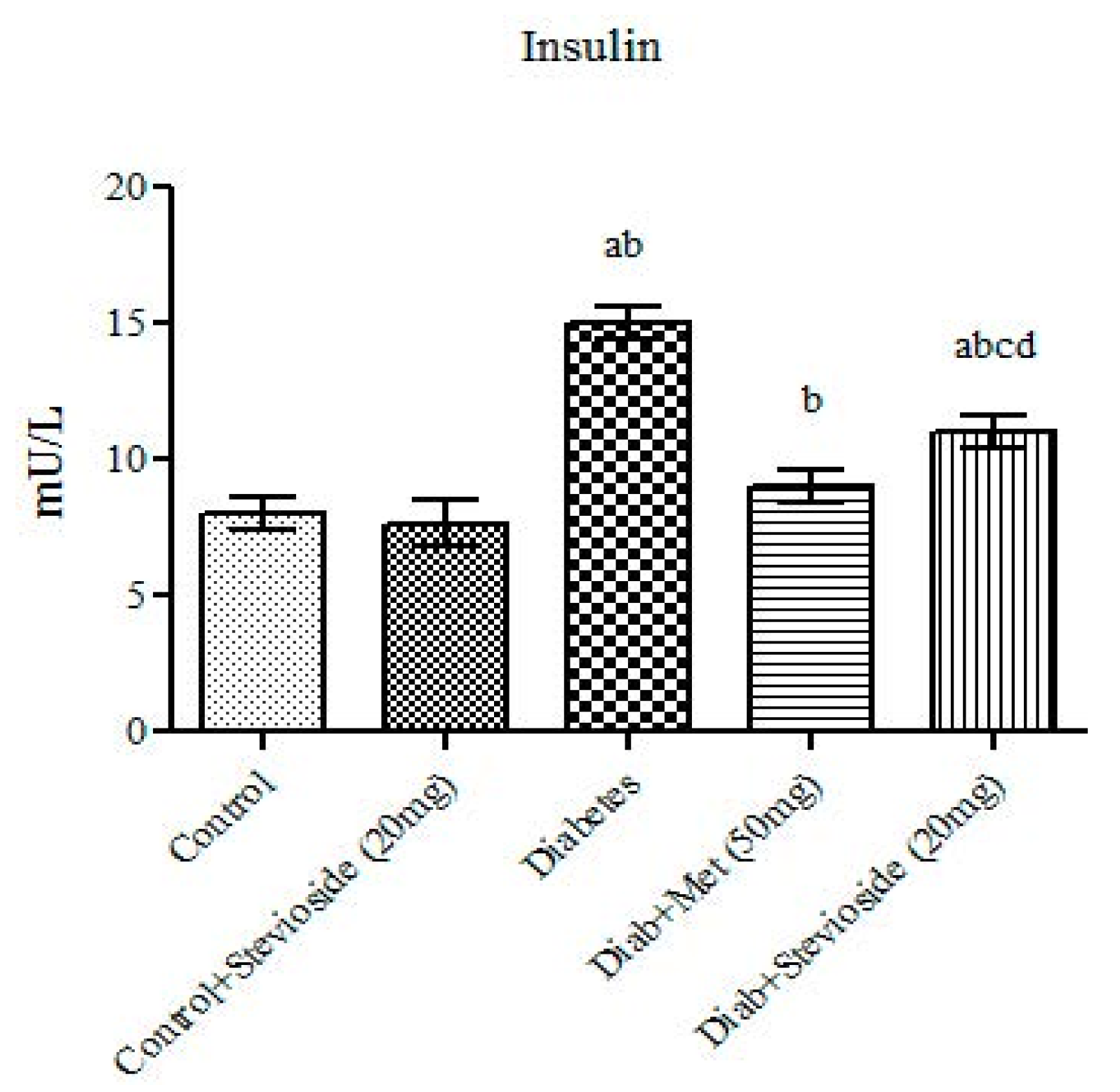
2.2. Effect of Stevioside on Fasting Serum Insulin T2DM Rats
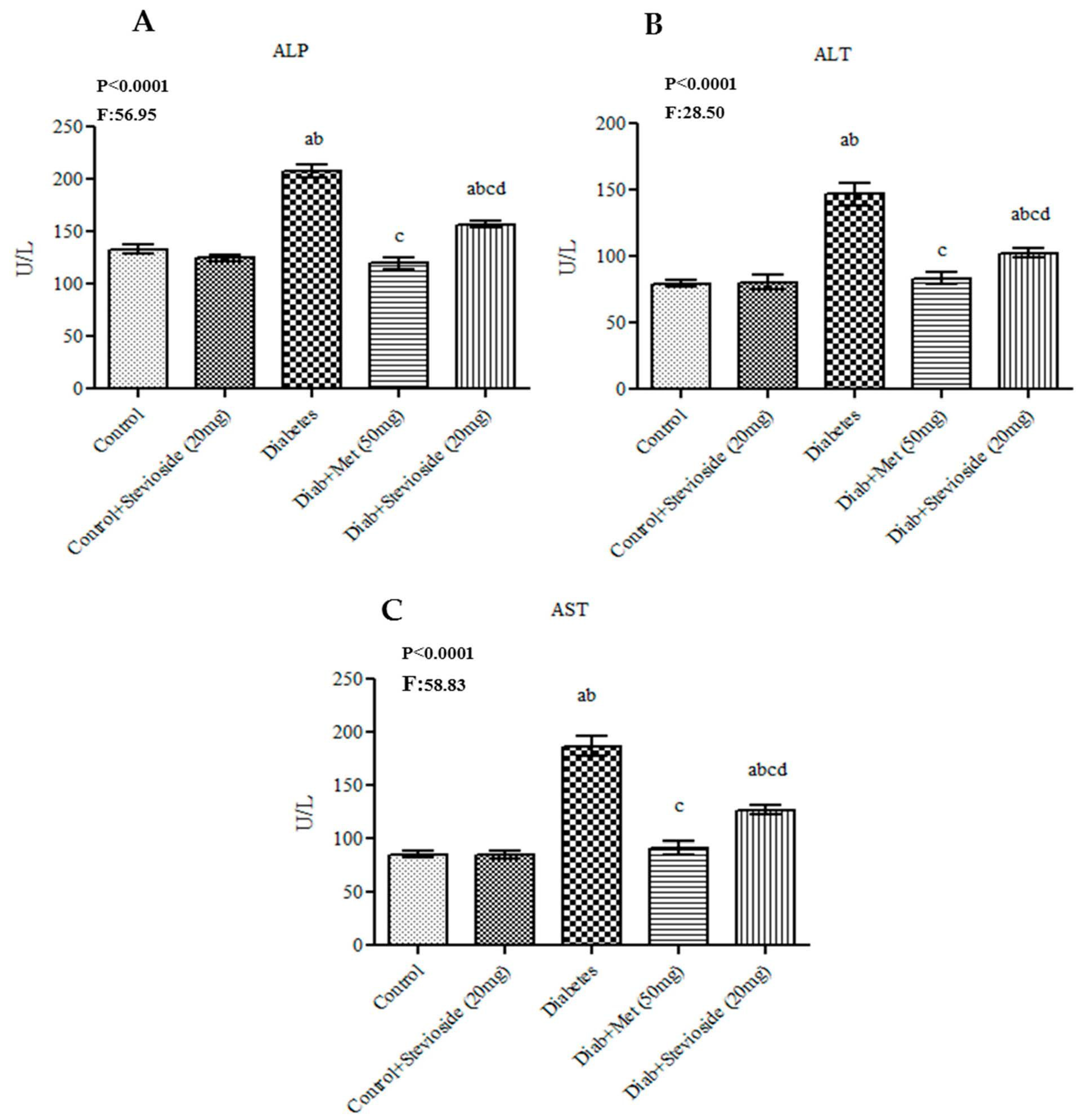
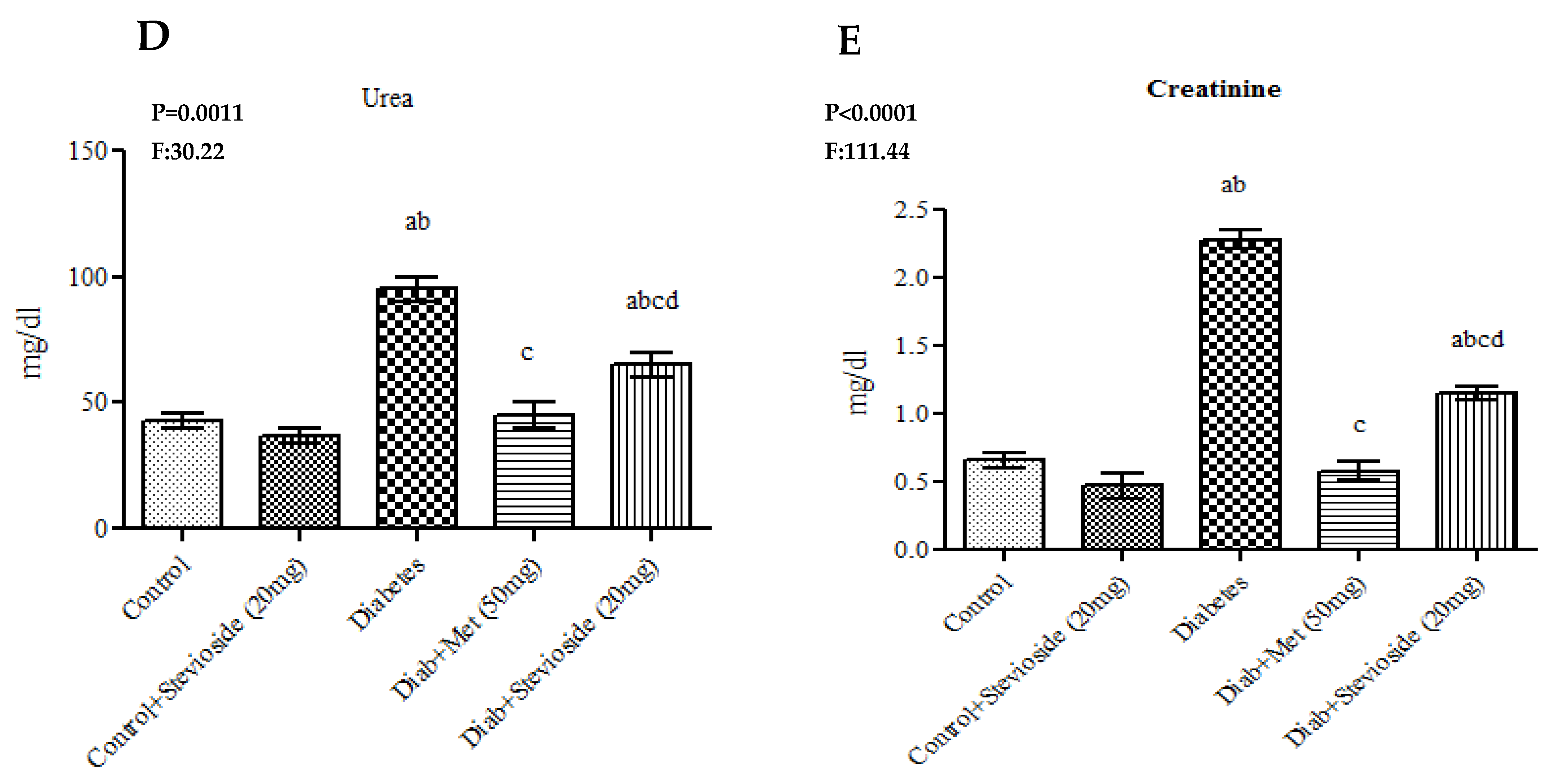
2.3. Effect of Stevioside on Liver and Kidney Function Markers in Diabetic Rats
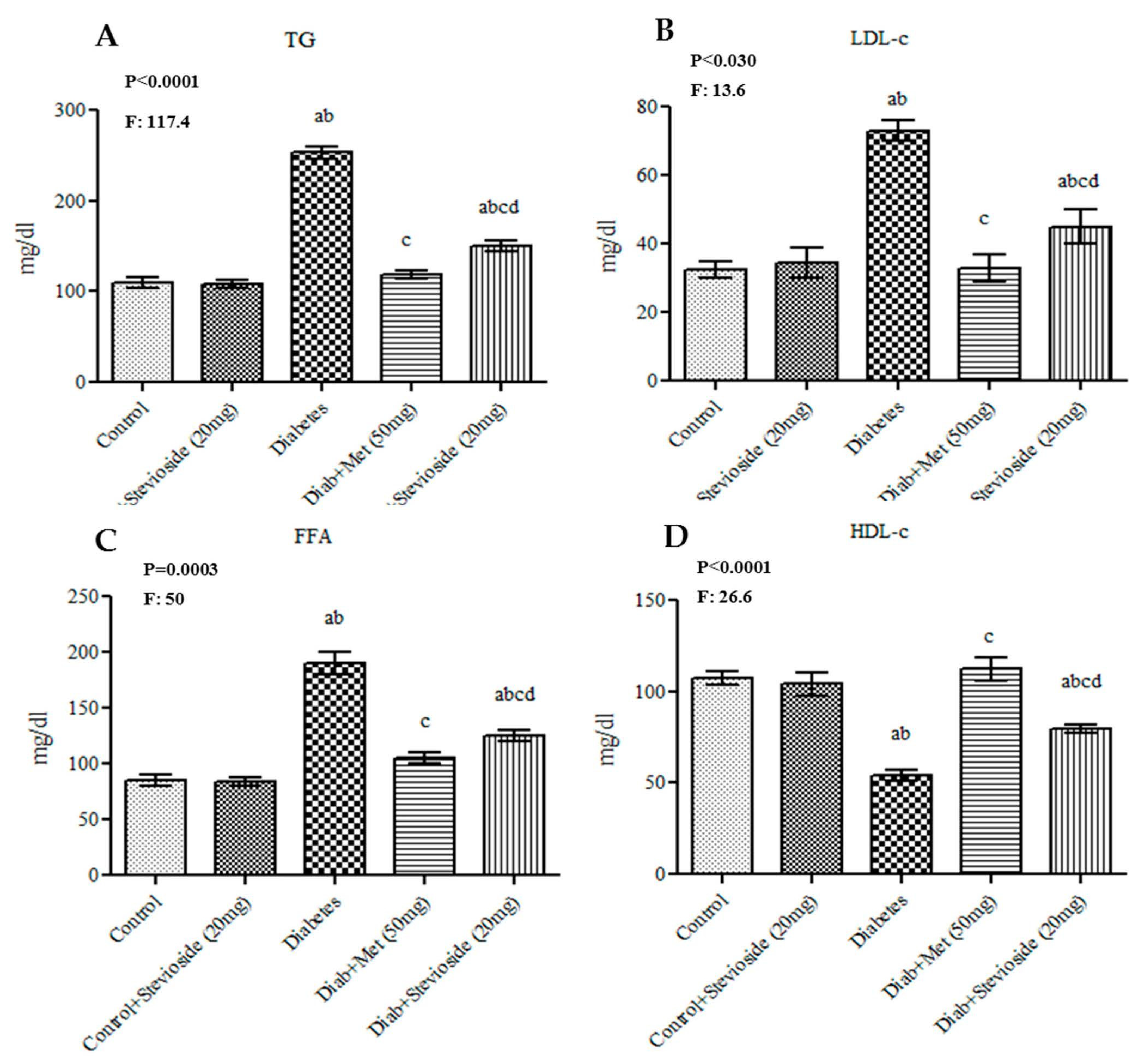
2.4. Stevioside Effect on Serum Lipid Profile
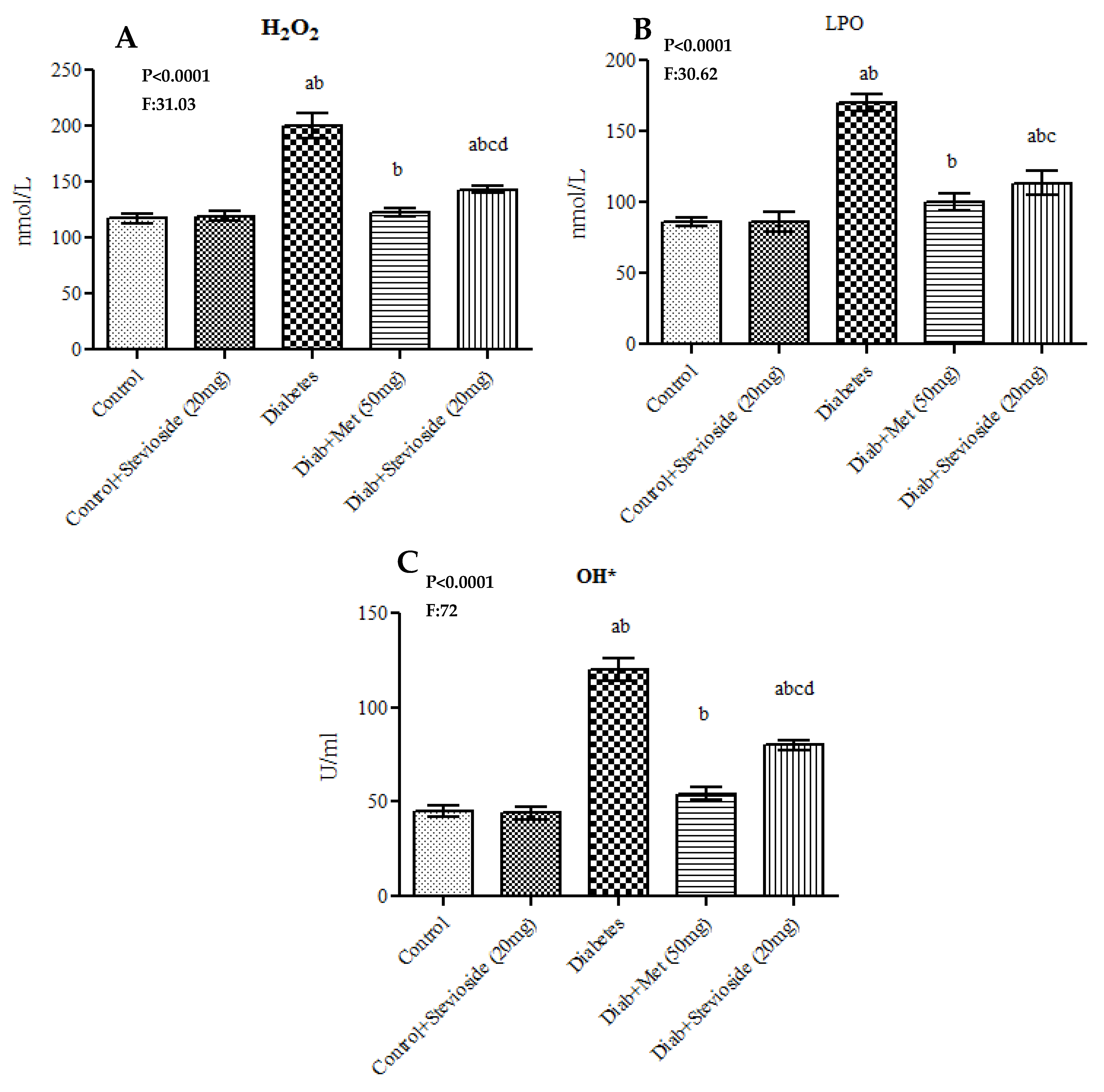
2.5. Stevioside Effect on Oxidative Stress Markers in the Gastrocnemius Muscle
2.6. Stevioside Elicits the Levels of Antioxidant Enzymes in the Gastrocnemius Muscles
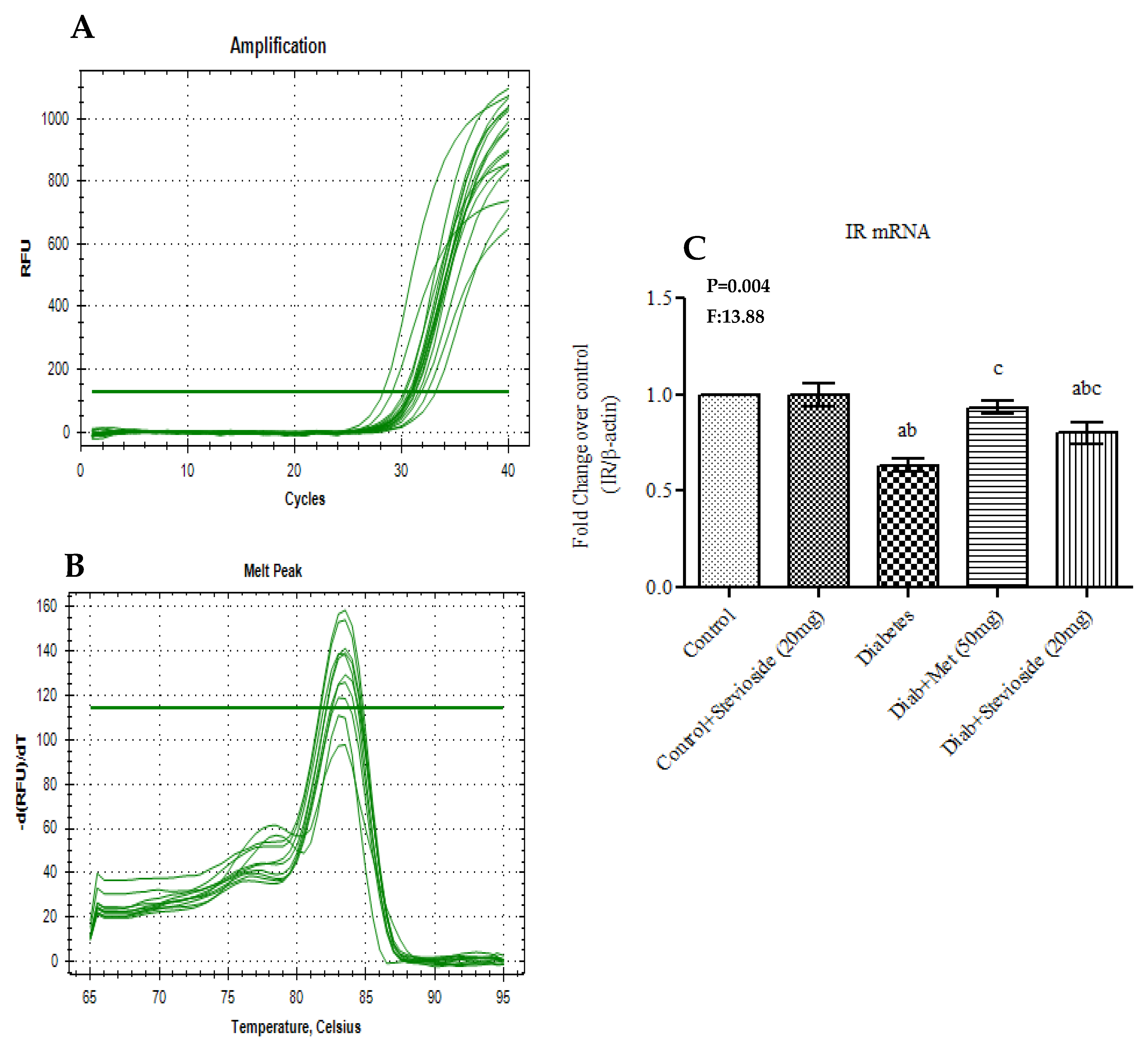
2.7. Stevioside Up-Regulated Muscular Gene Expression of Insulin Receptor (IR) in Type 2 Diabetic Rats
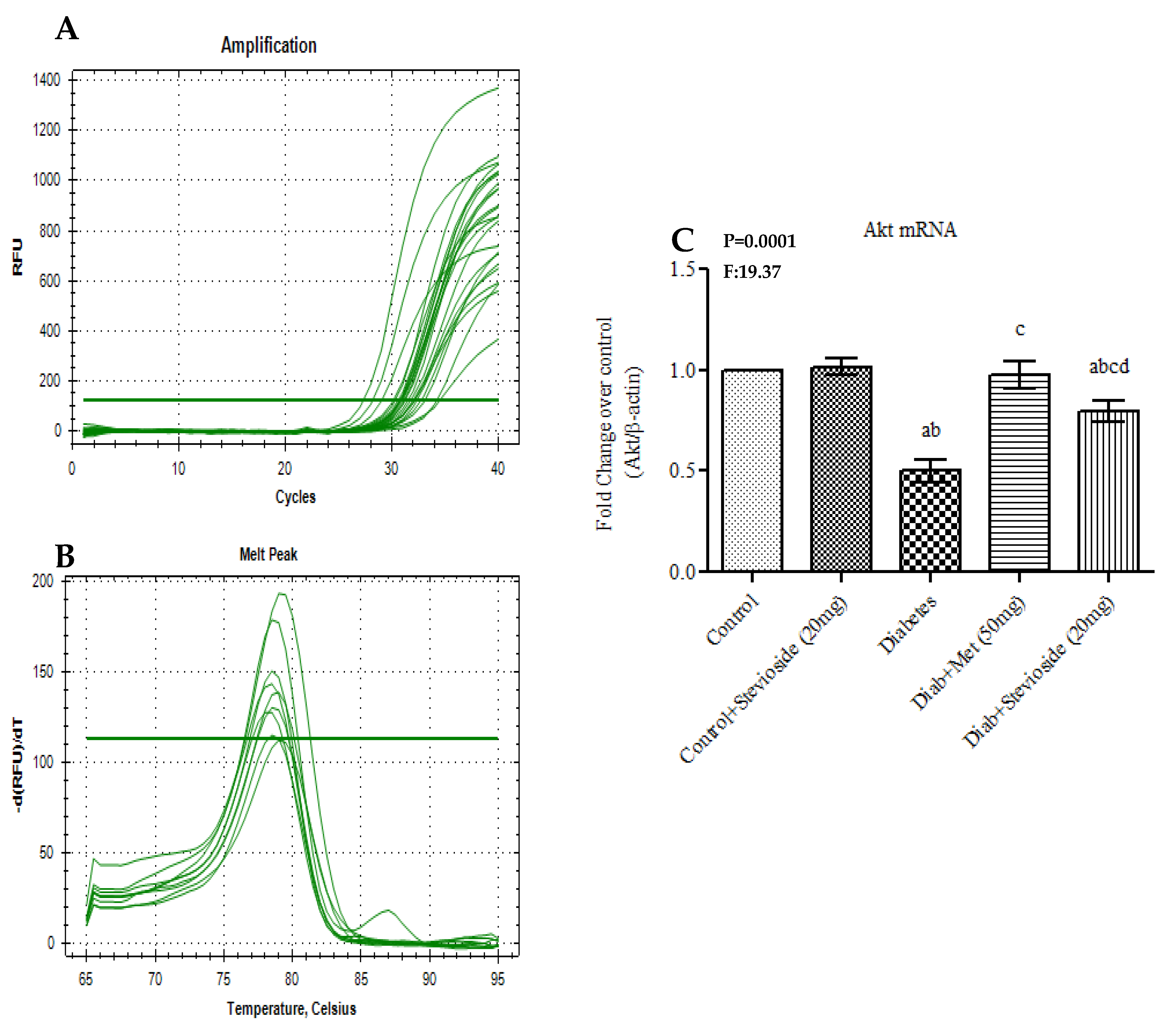
2.8. Stevioside Activated Akt Signaling Events in Diabetic Gastrocnemius Muscles
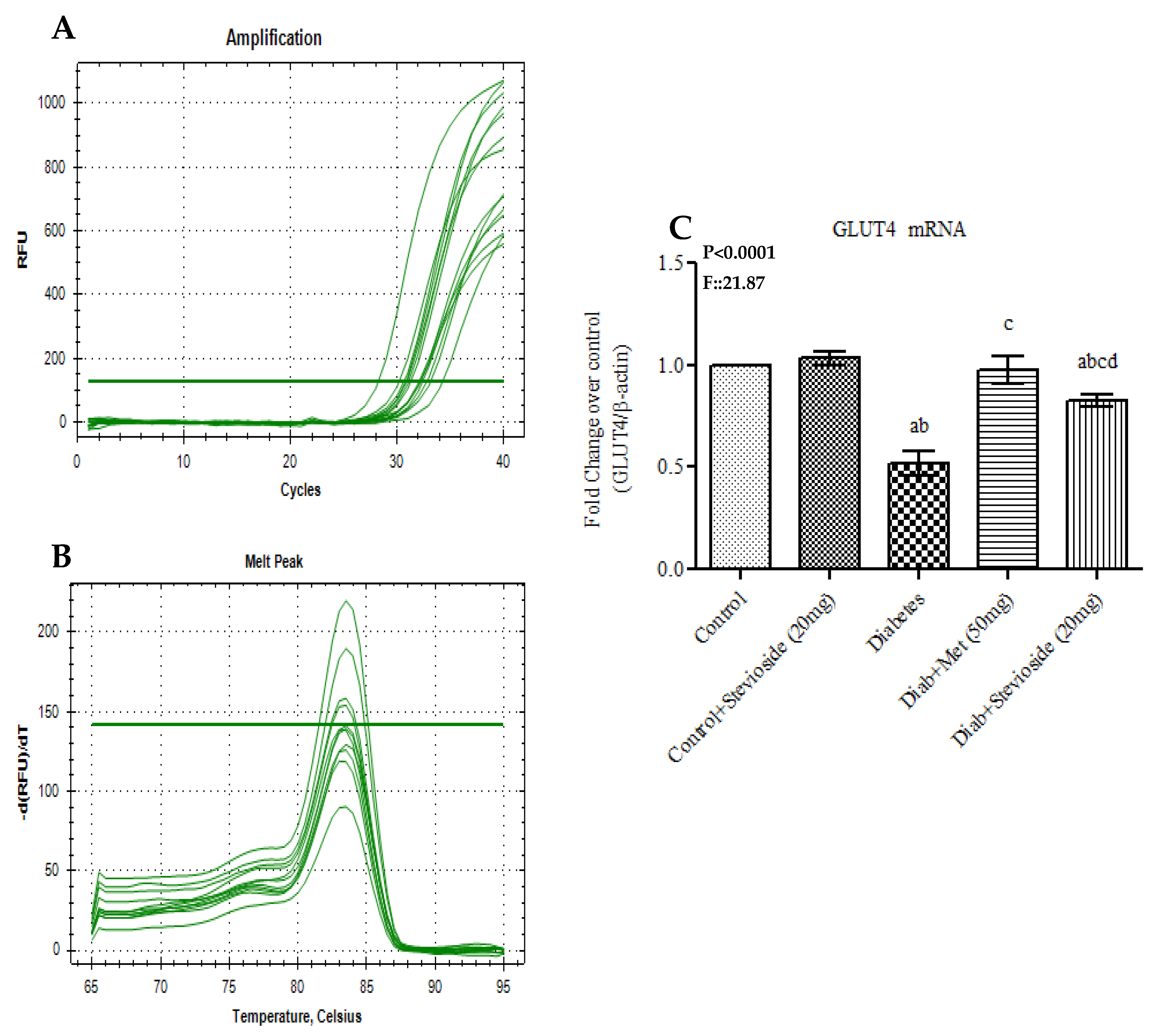
2.9. Stevioside Augmented GLUT 4 Trafficking in Gastrocnemius Muscles of T2DM Rats
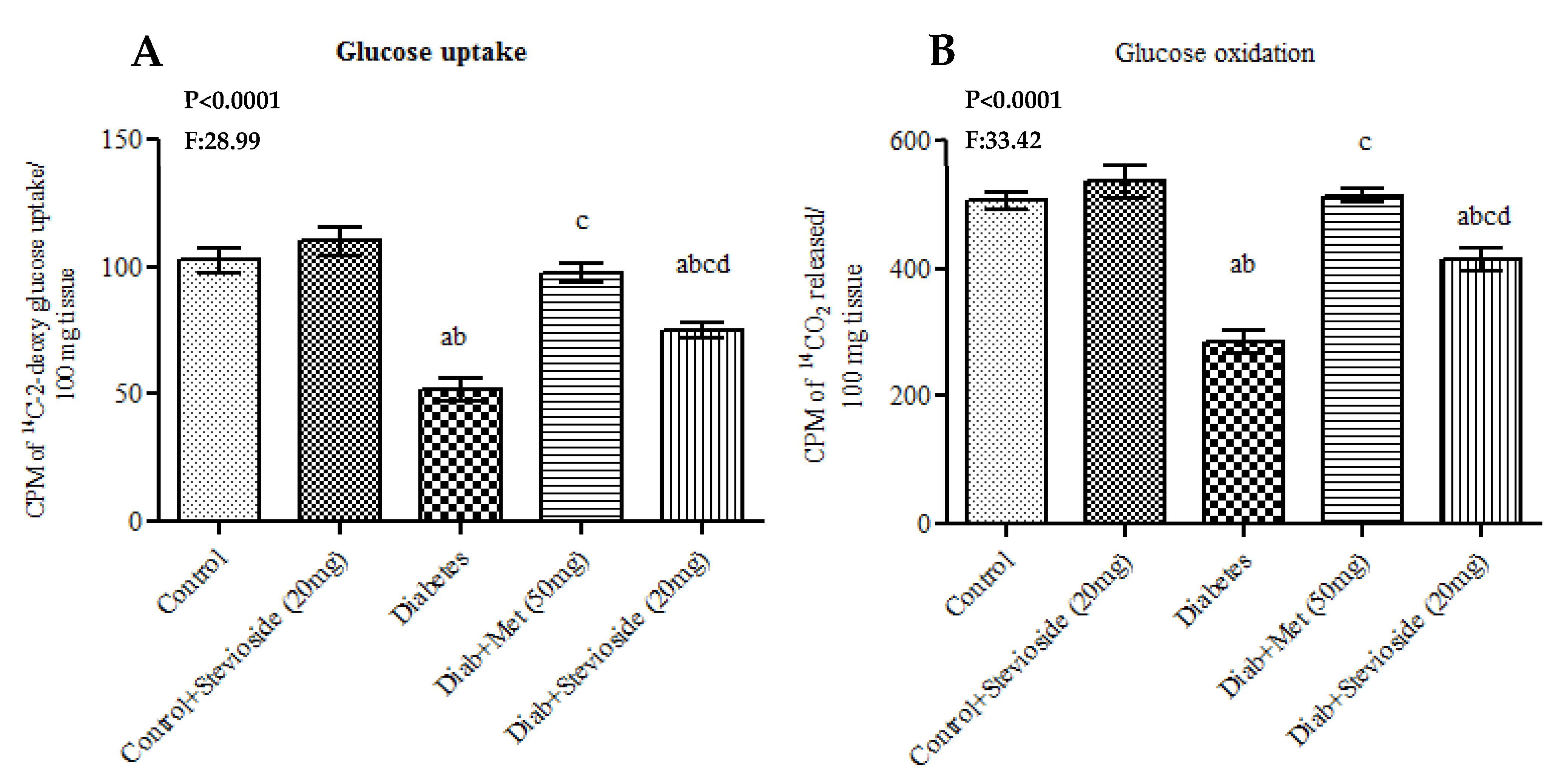
2.9.1. Effect of Stevioside on 14C-2-Deoxyglucose Uptake and 14C-Glucose Oxidation in the Gastrocnemius Muscles
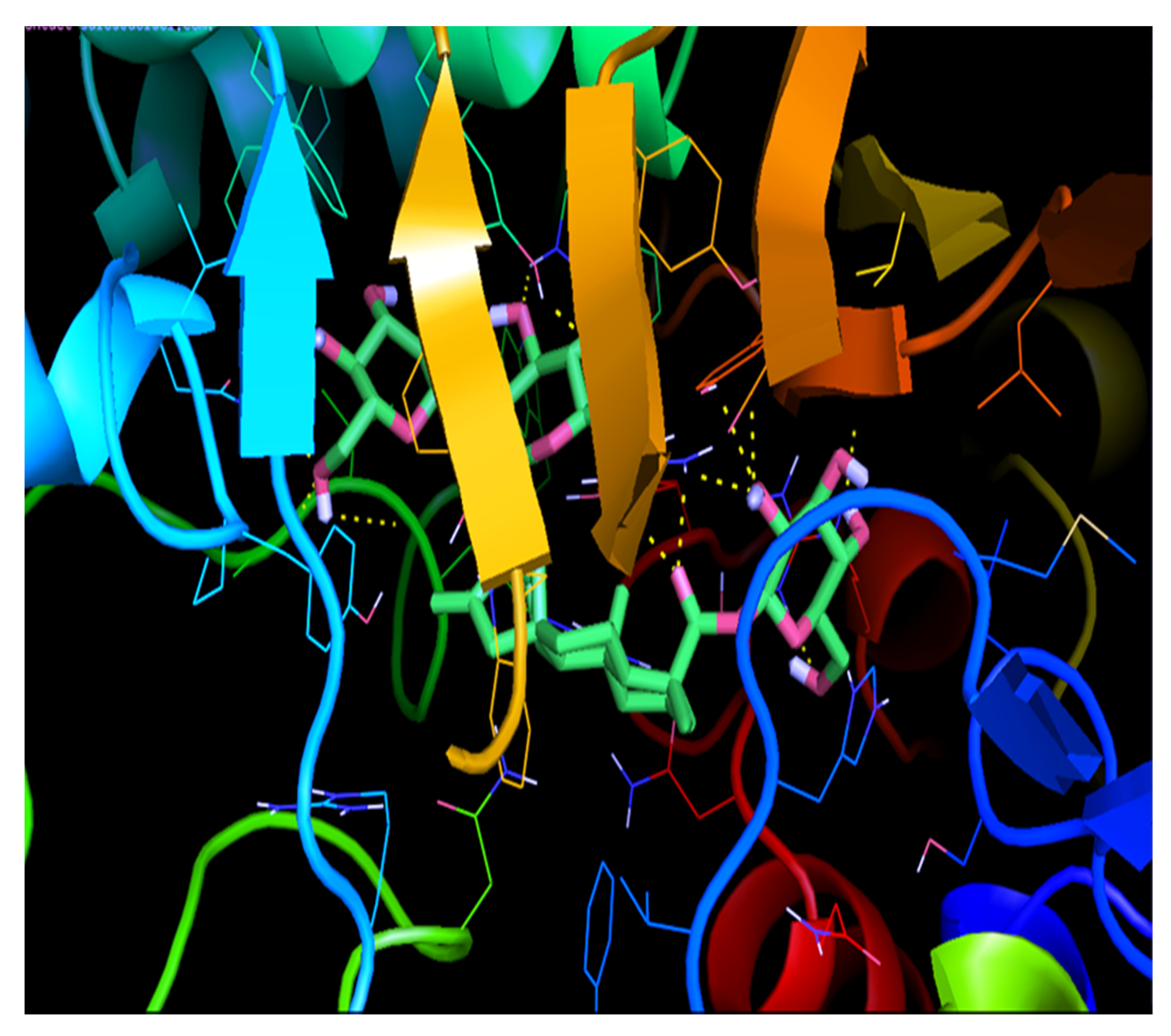
2.9.2. Molecular Docking Studies of IRS with Stevioside
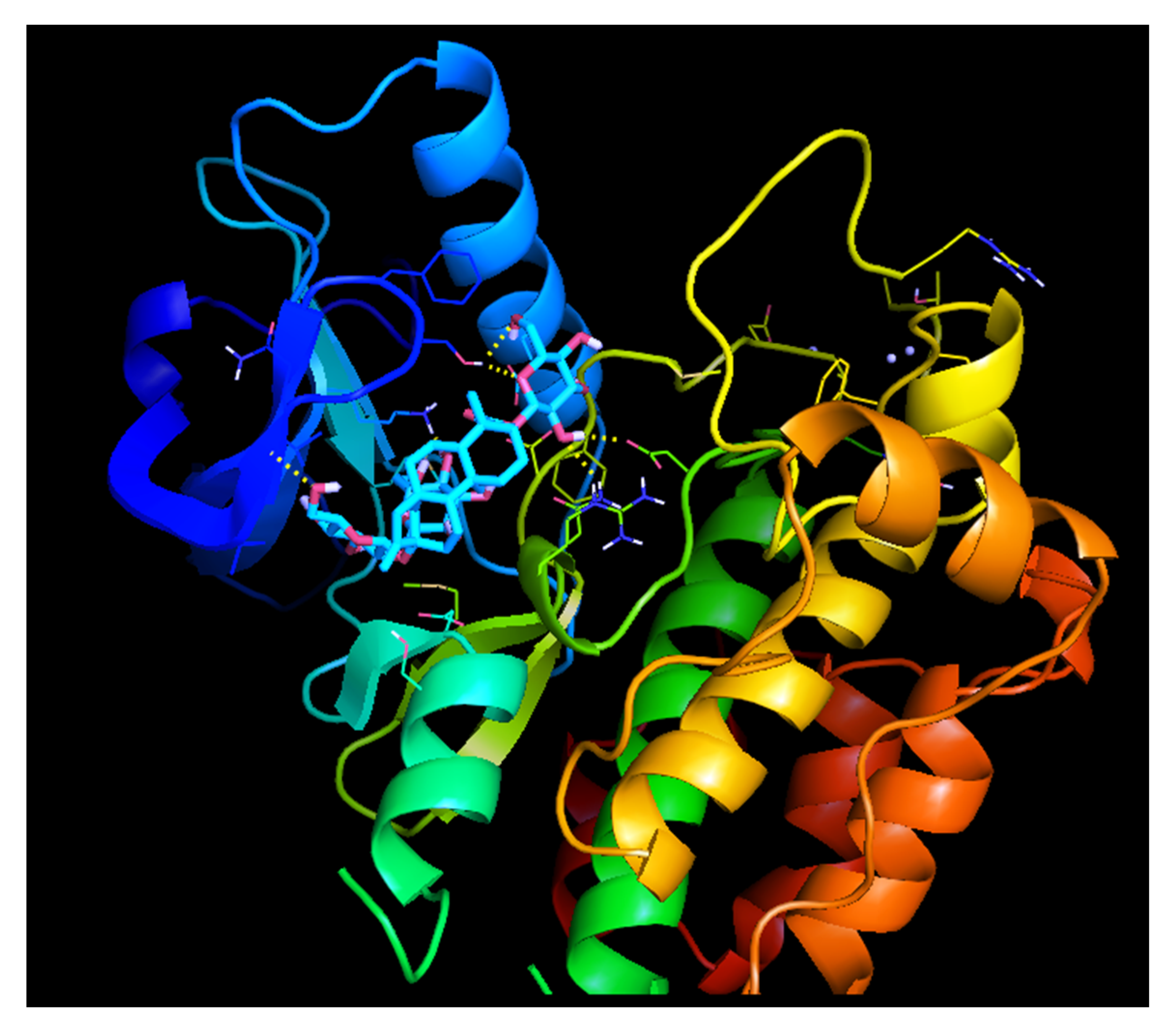
2.9.3. Molecular Docking Studies of GLUT 4 with Stevioside
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Animal
4.3. Induction of T2DM
4.4. Experimental Design
4.5. FBG Measurement
4.6. Oral Glucose Tolerance Test (OGTT)
4.7. Liver and Renal Function Markers
4.8. Estimation of Fasting Serum Insulin
4.9. Serum Lipid Profile Analysis
4.9.1. Estimation of ROS Levels in Gastrocnemius Muscles
4.9.2. Measurement of Antioxidant Enzymes in Gastrocnemius Muscles
4.9.3. Glucose Uptake by Gastrocnemius Muscles
4.9.4. Glucose Oxidation in Gastrocnemius Muscles
4.9.5. mRNA Expression Analysis
Total RNA Isolation, cDNA Conversion and Real-Time PCR
4.9.6. Statistical Analysis
4.9.7. In Silico Analysis
Protein Preparation
Homology Modelling of GLUT4 Protein
Protein Preparation
Ligand Preparation
Molecular Docking
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, V.; Najmi, A.K.; Akhtar, M.; Aqil, M.; Mujeeb, M.; Pillai, K.K. A pharmacological appraisal of medicinal plants with antidiabetic potential. J. Pharm. Bioallied Sci. 2012, 4, 27–42. [Google Scholar]
- Li, W.; Wan, H.; Yan, S.; Yan, Z.; Chen, Y.; Guo, P.; Ramesh, T.; Cui, Y.; Ning, L. Gold nanoparticles synthesized with Poria cocos modulates the anti-obesity parameters in high-fat diet and streptozotocin induced obese diabetes rat model. Arab J. Chem. 2020, 13, 5966–5977. [Google Scholar] [CrossRef]
- Lichtenstein, A.H.; Appel, L.J.; Brands, M.; Carnethon, M.; Daniels, S.; Franch, H.A.; Franklin, B.; Kris-Etherton, P.; Harris, W.S.; Howard, B.; et al. Diet and lifestyle recommendations revision 2006: A scientific statement from the American Heart Association Nutrition Committee. Circulation 2006, 114, 82–96. [Google Scholar] [CrossRef] [Green Version]
- Lozano, I.; Van der Werf, R.; Bietiger, W.; Seyfritz, E.; Peronet, C.; Pinget, M.; Jeandidier, N.; Maillard, E.; Marchioni, E.; Sigrist, S.; et al. High-fructose and high-fat diet-induced disorders in rats: Impact on diabetes risk, hepatic and vascular complications. JCNM 2016, 13, 15. [Google Scholar] [CrossRef] [Green Version]
- Rother, K.I. Diabetes treatment—bridging the divide. N. Engl. J. Med. 2007, 356, 1499–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Dellsperger, K.C.; Zhang, C. The link between metabolic abnormalities and endothelial dysfunction in type 2 diabetes: An update. Basic Res. Cardiol. 2012, 107, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagliassotti, M.J.; Horton, T.J.; Gayles, E.C.; Koppenhafer, T.A.; Rosenzweig, T.D.; Hill, J.O. Reduced insulin suppression of glucose appearance is related to susceptibility to dietary obesity in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1997, 272, 1264–1270. [Google Scholar] [CrossRef]
- Commerford, S.R.; Ferniza, J.B.; Bizeau, M.E.; Thresher, J.S.; Willis, W.T.; Pagliassotti, M.J. Diets enriched in sucrose or fat increase gluconeogenesis and G-6-Pase but not basal glucose production in rats. Am. J. Physiol. Endocrinol. Metab. 2002, 3, 545–555. [Google Scholar] [CrossRef] [Green Version]
- Podolin, D.A.; Wei, Y.; Pagliassotti, M.J. Effects of a high-fat diet and voluntary wheel running on gluconeogenesis and lipolysis in rats. J. Appl. Physiol. 1999, 86, 1374–1380. [Google Scholar] [CrossRef]
- Sifuentes-Franco, S.; Padilla-Tejeda, D.E.; Carrillo-Ibarra, S.; Miranda-Díaz, A.G. Oxidative stress, apoptosis, and mitochondrial function in diabetic nephropathy. Int. J. Endocrinol. 2018, 2018, 187587. [Google Scholar] [CrossRef]
- Yin, F.; Boveris, A.; Cadenas, E. Mitochondrial energy metabolism and redox signaling in brain aging and neurodegeneration. Antioxid. Redox Signal. 2014, 20, 353–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, F.; Siddaraju, N.S.; Urooj, A. In vitro hypoglycemic effects of Gymnema sylvestre, Tinospora cordifolia, Eugenia jambolana and Aegle marmelos. J. Nat. Med. 2011, 2, 52–55. [Google Scholar] [CrossRef]
- Aralelimath, V.R.; Bhise, S.B. Anti-diabetic effects of gymnema sylvester extract on streptozotocin induced diabetic rats and possible β-cell protective and regenerative evaluations. Dig. J. Nanomater. Biostruct. 2012, 7, 135–142. [Google Scholar]
- Carakostas, M.C.; Curry, L.L.; Boileau, A.C.; Brusick, D.J. Overview: The history, technical function and safety of rebaudioside A, a naturally occurring steviol glycoside, for use in food and beverages. Food Chem. Toxicol. 2008, 46, S1–S10. [Google Scholar] [CrossRef]
- Cantabella, D.; Piqueras, A.; Acosta-Motos, J.R.; Bernal-Vicente, A.; Hernández, J.A.; Díaz-Vivancos, P. Salt-tolerance mechanisms induced in Stevia rebaudiana Bertoni: Effects on mineral nutrition, antioxidative metabolism and steviol glycoside content. Plant Physiol. Biochem. 2017, 115, 484–496. [Google Scholar] [CrossRef]
- Modi, A.; Litoriya, N.; Prajapati, V.; Rafalia, R.; Narayanan, S. Transcriptional profiling of genes involved in steviol glycoside biosynthesis in Stevia rebaudiana bertoni during plant hardening. Dev. Dyn. 2014, 243, 1067–1073. [Google Scholar] [CrossRef]
- Yadav, A.K.; Singh, S.; Dhyani, D.; Ahuja, P.S. A review on the improvement of stevia [Stevia rebaudiana (Bertoni)]. Can. J. Plant Sci. 2011, 91, 1–27. [Google Scholar] [CrossRef]
- Ullah, A.; Munir, S.; Mabkhot, Y.; Badshah, S.L. Bioactivity profile of the diterpene isosteviol and its derivatives. Molecules 2019, 24, 678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbas Momtazi-Borojeni, A.; Esmaeili, S.A.; Abdollahi, E.; Sahebkar, A. A review on the pharmacology and toxicology of steviol glycosides extracted from Stevia rebaudiana. Curr. Pharm. Des. 2017, 23, 1616–1622. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ruiz, J.C.; Moguel-Ordoñez, Y.B.; Segura-Campos, M.R. Biological activity of Stevia rebaudiana Bertoni and their relationship to health. Crit. Rev. Food Sci. Nutr. 2017, 57, 2680–2690. [Google Scholar] [CrossRef] [PubMed]
- Jeppesen, P.B.; Gregersen, S.; Rolfsen, S.E.; Jepsen, M.; Colombo, M.; Agger, A.; Xiao, J.; Kruhoffer, M.; Orntoft, T.; Hermansen, K. Antihyperglycemic and blood pressure-reducing effects of Stevioside in the diabetic Goto-Kakizaki rat. Metabolism 2003, 52, 372–378. [Google Scholar] [CrossRef] [Green Version]
- Merz, K.E.; Thurmond, D.C. Role of Skeletal Muscle in Insulin Resistance and Glucose Uptake. Compr. Physiol. 2020, 10, 785–809. [Google Scholar]
- Phielix, E.; Mensink, M. Type 2 diabetes mellitus and skeletal muscle metabolic function. Physiol. Behav. 2008, 94, 252–258. [Google Scholar] [CrossRef]
- Klip, A.; Ishiki, M. Recent developments in the regulation of glucose transporter-4 traffic: New signals, locations, and partners. Endocrinology 2005, 146, 5071–5078. [Google Scholar]
- DeFronzo, R.A.; Tripathy, D. Skeletal muscle insulin resistance is the primary defect in Type 2 diabetes. Diabetes Care 2009, 32, S157–S163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdul-Ghani, M.A.; DeFronzo, R.A. Pathogenesis of insulin resistance in skeletal muscle. J. Biomed. Biotechnol. 2010, 2010, 476279. [Google Scholar] [CrossRef] [Green Version]
- Di Meo, S.; Iossa, S.; Venditti, P. Skeletal muscle insulin resistance: Role of mitochondria and other ROS sources. J. Endocrinol. 2017, 233, R15–R42. [Google Scholar] [CrossRef] [Green Version]
- Zhao, P.; Tian, D.; Song, G.; Ming, Q.; Liu, J.; Shen, J.; Liu, Q.H.; Yang, X. Neferine Promotes GLUT 4 Expression and Fusion With the Plasma Membrane to Induce Glucose Uptake in L6 Cells. Front. Pharmacol. 2019, 10, 999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logan, I.E.; Bobe, G.; Miranda, C.L.; Vasquez-Perez, S.; Choi, J.; Lowry, M.B.; Sharpton, T.J.; Morgun, A.; Maier, C.S.; Stevens, J.F.; et al. Germ-Free Swiss Webster Mice on a High-Fat Diet Develop Obesity, Hyperglycemia, and Dyslipidemia. Microorganisms 2020, 8, 520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotimi, S.O.; Rotimi, O.A.; Adelani, I.B.; Onuzulu, C.; Obi, P.; Okungbaye, R. Stevioside modulates oxidative damage in the liver and kidney of high fat/low streptozocin diabetic rats. Heliyon 2018, 4, e00640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, R.; Jiang, S.; Zhang, L.; Yu, Z. Mitochondrial electron transport chain, ROS generation and uncoupling (Review). Int. J. Mol. Med. 2019, 44, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Schönfeld, P.; Wojtczak, L. Fatty acids as modulators of the cellular production of reactive oxygen species. Free Radic. Biol. Med. 2008, 45, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martín, C. Pathophysiology of Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef] [PubMed]
- Kaneto, H.; Matsuoka, T.A.; Nakatani, Y.; Kawamori, D.; Miyatsuka, T.; Matsuhisa, M.; Yamasaki, Y. Oxidative stress, ER stress, and the JNK pathway in type 2 diabetes. J. Mol. Med. 2005, 83, 429–439. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, J.; Sathish, S.; Mayilvanan, C.; Balasubramanian, K. Excess aldosterone-induced changes in insulin signaling molecules and glucose oxidation in gastrocnemius muscle of adult male rat. Mol. Cell Biochem. 2013, 372, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Devasagayam, T.; Tarachand, U. Decreased lipid peroxidation in the rat kidney during gestation. Biochem. Biophys. Res. Commun. 1987, 145, 134–138. [Google Scholar] [CrossRef]
- Pick, E.; Keisari, Y. Superoxide anion and hydrogen peroxide production by chemically elicited peritoneal macrophages induction by multiple nonphagocytic stimuli. Cell Immunol. 1981, 59, 301–318. [Google Scholar] [CrossRef]
- Puntarulo, S.; Cederbaum, A.I. Comparison of the ability of ferric complexes to catalyze microsomal chemiluminescence, lipid peroxidation, and hydroxyl radical generation. Arch. Biochem. Biophys. 1988, 264, 482–491. [Google Scholar] [CrossRef]
- Sinha, A.K. Colorimetric assay of catalase. Anal. Biochem. 1972, 47, 389–394. [Google Scholar] [CrossRef]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef]
- Moron, M.S.; Depierre, J.W.; Mannervik, B. Levels of glutathione, glutathione reductase and glutathione S-transferase activities in rat lung and liver. Biochim. Biophys. Acta 1979, 582, 67–78. [Google Scholar] [CrossRef]
- Rotruck, J.T.; Pope, A.L.; Ganther, H.E.; Swanson, A.B.; Hafeman, D.G.; Hoekstra, W.G. Selenium: Biochemical role as a component of glutathione peroxidase. Science 1973, 179, 588–590. [Google Scholar] [CrossRef] [PubMed]
- Valverde, A.M.; Navarro, P.; Teruel, T.; Conejo, R.; Benito, M.; Lorenzo, M. Insulin and insulin-like growth factor I up-regulate GLUT 4 gene expression in fetal brown adipocytes, in a phosphoinositide 3-kinase-dependent manner. Biochem. J. 1999, 337, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Kraft, L.A.; Johnson, A.D. Epididymal carbohydrate metabolism II. Substrates and pathway utilization of caput and cauda epididymal tissue from the rabbit, rat and mouse. Comp. Biochem. Physiol. 1972, 42, 451–461. [Google Scholar] [CrossRef]
- Maruthamuthu, M.K.; Ganesh, I.; Ravikumar, S.; Hong, S.H. Evaluation of zraP gene expression characteristics and construction of a lead (Pb) sensing and removal system in a recombinant Escherichia coli. Biotechnol. Lett. 2015, 37, 659–664. [Google Scholar] [CrossRef]
- Somasundaram, S.; Maruthamuthu, M.K.; Ganesh, I.; Eom, G.T.; Hong, S.H. Enchancement of Gamma-Aminobutyric Acid Production by Co-Localization of Neurospora crassa OR74A Glutamate Decarboxylase with Escherichia coli GABA Transporter Via Synthetic Scaffold Complex. J. Microbiol. Biotechnol. 2017, 27, 1664–1669. [Google Scholar] [CrossRef] [Green Version]
- Kannan, M.M.; Vanitha, J.; Jiang, S.; Ramachandran, S. Impact of colchicine treatment on sorghum bicolor BT×623. Mol. Plant Breed. 2013, 4, 128–135. [Google Scholar] [CrossRef]
- Gonzalez, C.; Alonso, A.; Fernandez, R.; Patterson, A.M. Regulation of insulin receptor substrate-1 in the liver, skeletal muscle and adipose tissue of rats throughout pregnancy. Gynecol. Endocrinol. 2003, 17, 187–197. [Google Scholar] [PubMed]
- Sharma, N.; Arias, E.B.; Cartee, G.D. Rapid reversal of insulin stimulated AS160 phosphorylation in rat skeletal muscle after insulin exposure. Physiol. Res. 2010, 59, 71–78. [Google Scholar] [CrossRef]
- Liu, Y.; Wan, Q.; Guan, Q.; Gao, L.; Zhao, J. High-fat diet feeding impairs both the expression and activity of AMPKa in rats’ skeletal muscle. Biochem. Biophys. Res. Commun. 2006, 339, 701–707. [Google Scholar] [CrossRef]
- Peinnequin, A.; Mouret, C.; Birot, O.; Alonso, A.; Mathieu, J.; Clarençon, D.; Agay, D.; Chancerelle, Y.; Multon, E. Rat pro-inflammatory cytokine and cytokine related mRNA quantification by real-time polymerase chain reaction using SYBR green. BMC Immunol. 2004, 5, 3. [Google Scholar] [CrossRef] [Green Version]
- Maruthamuthu, M.K.; Hong, J.; Arulsamy, K.; Somasundaram, S.; Hong, S.; Choe, W.S.; Yoo, I.K. Development of bisphenol A-removing recombinant Escherichia coli by monomeric and dimeric surface display of bisphenol A-binding peptide. Bioprocess Biosyst. Eng. 2018, 41, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Davidson, J.L.; Wang, J.; Maruthamuthu, M.K.; Dextre, A.; Pascual-Garrigos, A.; Mohan, S.; Putikam, S.V.S.; Osman, F.O.I.; McChesney, D.; Seville, J.; et al. A paper-based colorimetric molecular test for SARS-CoV-2 in saliva. Biosens. Bioelectron. X 2021, 9, 100076. [Google Scholar] [CrossRef] [PubMed]
- Maruthamuthu, M.K.; Nadarajan, S.P.; Ganesh, I.; Ravikumar, S.; Yun, H.; Yoo, I.K.; Hong, S.H. Construction of a high efficiency copper adsorption bacterial system via peptide display and its application on copper dye polluted wastewater. Bioprocess Biosyst. Eng. 2015, 38, 2077–2084. [Google Scholar] [CrossRef] [PubMed]
- Maruthamuthu, M.K.; Selvamani, V.; Nadarajan, S.P.; Yun, H.; Oh, Y.K.; Eom, G.T.; Hong, S.H. Manganese and cobalt recovery by surface display of metal binding peptide on various loops of OmpC in Escherichia coli. J. Ind. Microbiol. Biotechnol. 2018, 45, 31–41. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Binding Affinity (Kcal/mol) | H-Bond Details | Distance |
|---|---|---|---|
| GLUT 4 | −7.8 | GLN-113 | 2.3 |
| TYR-168 | 2.5 | ||
| TRP-173 | 2.2 | ||
| ARG-188 | 2.3 | ||
| ALA-190 | 2.3 | ||
| ARG-336 | 2.6 | ||
| GLU-359 | 2.1 | ||
| GLY-404 | 2.2 | ||
| TYR-405 | 2.6 | ||
| ARG-433 | 2.0 | ||
| IRS-1 | −8.2 | GLN-977 | 2.5 |
| SER-979 | 1.8 | ||
| LYS-1003 | 2.0 | ||
| ASP-1105 | 2.2 | ||
| ASN-1110 | 2.7 |
| Name of the Gene | Primer Sequence | Reference |
|---|---|---|
| Rat IR | Sense primer: 5′-GCC ATC CCG AAA GCG AAG ATC-3′ | [49] |
| Anti-sense primer: 5′-TCT GGG TCC TGA TTG CAT-3′ | ||
| Rat Akt | Sense primer: 5′-GGA AGC CTT CAG TTT GGA TCC CAA-3′ | [50] |
| Anti-sense primer: 5′-AGT GGA AAT CCA GTT CCG AGC TTG-3′ | ||
| Rat GLUT 4 | Sense primer: 5′-GGG CTG TGA GTG AGT GCT TTC-3′ | [51] |
| Anti-sense primer: 5′-CAG CGA GGC AAG GCT AGA-3′ | ||
| Rat β-actin | Sense primer: 5′-AAG TCC CTC ACC CTC CCA AAA G-3′ | [52] |
| Anti-sense primer: 5′-AAG CAA TGC TGT CAC CTT CCC-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deenadayalan, A.; Subramanian, V.; Paramasivan, V.; Veeraraghavan, V.P.; Rengasamy, G.; Coiambatore Sadagopan, J.; Rajagopal, P.; Jayaraman, S. Stevioside Attenuates Insulin Resistance in Skeletal Muscle by Facilitating IR/IRS-1/Akt/GLUT 4 Signaling Pathways: An In Vivo and In Silico Approach. Molecules 2021, 26, 7689. https://doi.org/10.3390/molecules26247689
Deenadayalan A, Subramanian V, Paramasivan V, Veeraraghavan VP, Rengasamy G, Coiambatore Sadagopan J, Rajagopal P, Jayaraman S. Stevioside Attenuates Insulin Resistance in Skeletal Muscle by Facilitating IR/IRS-1/Akt/GLUT 4 Signaling Pathways: An In Vivo and In Silico Approach. Molecules. 2021; 26(24):7689. https://doi.org/10.3390/molecules26247689
Chicago/Turabian StyleDeenadayalan, Abilasha, Vijayalakshmi Subramanian, Vijayalakshmi Paramasivan, Vishnu Priya Veeraraghavan, Gayathri Rengasamy, Janaki Coiambatore Sadagopan, Ponnulakshmi Rajagopal, and Selvaraj Jayaraman. 2021. "Stevioside Attenuates Insulin Resistance in Skeletal Muscle by Facilitating IR/IRS-1/Akt/GLUT 4 Signaling Pathways: An In Vivo and In Silico Approach" Molecules 26, no. 24: 7689. https://doi.org/10.3390/molecules26247689
APA StyleDeenadayalan, A., Subramanian, V., Paramasivan, V., Veeraraghavan, V. P., Rengasamy, G., Coiambatore Sadagopan, J., Rajagopal, P., & Jayaraman, S. (2021). Stevioside Attenuates Insulin Resistance in Skeletal Muscle by Facilitating IR/IRS-1/Akt/GLUT 4 Signaling Pathways: An In Vivo and In Silico Approach. Molecules, 26(24), 7689. https://doi.org/10.3390/molecules26247689