
Highly Valuable Polyunsaturated Fatty Acids from Microalgae: Strategies to Improve Their Yields and Their Potential Exploitation in Aquaculture

 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Modulation of Growth Conditions to Enhance the Production of PUFAs
2.1. Temperature
2.2. Nutrients
2.3. Salinity
2.4. Irradiance
2.5. Autotrophic, Heterotrophic, and Mixotrophic Conditions
2.6. CO2
3. Genetic Engineering for PUFAs Production
3.1. Genetic Transformation and Gene Perturbation in Microalgae
3.2. Enhancement of the Fatty Acid Biosynthetic Pathway
3.3. Altering Elongation and Desaturation
3.4. Enhancement of the TAG Biosynthetic Pathway
3.5. Inhibition of Starch and Other Complex Polysaccharides’ Biosynthesis
3.6. Altering Pyruvate Metabolism
3.7. Reducing Lipid Catabolism
3.8. Overexpression of Transcription Factors
4. Potential of Microalgae for Sustainable Aquaculture
5. Extraction, Purification, and Stability of PUFAs
6. Summary, Conclusions, and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Ananthi, V.; Raja, R.; Carvalho, I.S.; Brindhadevi, K.; Pugazhendhi, A.; Arun, A. A realistic scenario on microalgae based biodiesel production: Third generation biofuel. Fuel 2021, 284, 118965. [Google Scholar] [CrossRef]
- D’Ippolito, G.; Sardo, A.; Paris, D.; Vella, F.M.; Adelfi, M.G.; Botte, P.; Gallo, C.; Fontana, A. Potential of lipid metabolism in marine diatoms for biofuel production. Biotechnol. Biofuels 2015, 8, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farahin, A.W.; Yusoff, F.M.; Basri, M.; Nagao, N.; Shariff, M. Use of microalgae: Tetraselmis tetrathele extract in formulation of nanoemulsions for cosmeceutical application. J. Appl. Phycol. 2019, 31, 1743–1752. [Google Scholar] [CrossRef]
- Morocho-Jacome, A.L.; Ruscinc, N.; Martinez, R.M.; de Carvalho, J.C.M.; de Almeida, T.S.; Rosado, C.; Costa, J.G.; Velasco, M.V.R.; Baby, A.R. (Bio)technological aspects of microalgae pigments for cosmetics. Appl. Microbiol. Biotechnol. 2020, 104, 9513–9522. [Google Scholar] [CrossRef] [PubMed]
- Lauritano, C.; Andersen, J.H.; Hansen, E.; Albrigtsen, M.; Escalera, L.; Esposito, F.; Helland, K.; Hanssen, K.Ø.; Romano, G.; Ianora, A. Bioactivity screening of microalgae for antioxidant, anti-inflammatory, anticancer, anti-diabetes, and antibacterial activities. Front. Mar. Sci. 2016, 3, 68. [Google Scholar] [CrossRef] [Green Version]
- Jimenez, R.; Markou, G.; Tayibi, S.; Barakat, A.; Chapsal, C.; Monlau, F. Production of microalgal slow-release fertilizer by valorizing liquid agricultural digestate: Growth experiments with tomatoes. Appl. Sci. 2020, 10, 3890. [Google Scholar] [CrossRef]
- Uysal, O.; Uysal, F.O.; Ekinci, K. Evaluation of microalgae as microbial fertilizer. Eur. J. Sustain. Dev. 2015, 4, 77–82. [Google Scholar] [CrossRef]
- Chandrashekharaiah, P.; Sanyal, D.; Dasgupta, S.; Banik, A. Cadmium biosorption and biomass production by two freshwater microalgae Scenedesmus acutus and Chlorella pyrenoidosa: An integrated approach. Chemosphere 2021, 269, 128755. [Google Scholar] [CrossRef]
- Hena, S.; Gutierrez, L.; Croue, J.P. Removal of pharmaceutical and personal care products (PPCPs) from wastewater using microalgae: A review. J. Hazard. Mater. 2021, 403, 124041. [Google Scholar] [CrossRef]
- Ferrazzano, G.F.; Papa, C.; Pollio, A.; Ingenito, A.; Sangianantoni, G.; Cantile, T. Cyanobacteria and microalgae as sources of functional foods to improve human general and oral health. Molecules 2020, 25, 5164. [Google Scholar] [CrossRef]
- Chauton, M.S.; Reitan, K.I.; Norsker, N.H.; Tveteras, R.; Kleivdal, H.T. A techno-economic analysis of industrial production of marine microalgae as a source of EPA and DHA-rich raw material for aquafeed: Research challenges and possibilities. Aquaculture 2015, 436, 95–103. [Google Scholar] [CrossRef]
- Chandra, R.; Iqbal, H.M.N.; Vishal, G.; Lee, H.S.; Nagra, S. Algal biorefinery: A sustainable approach to valorize algal-based biomass towards multiple product recovery. Bioresour. Technol. 2019, 278, 346–359. [Google Scholar] [CrossRef] [PubMed]
- ElMekawy, A.; Hegab, H.M.; Mohanakrishna, G.; Elbaz, A.F.; Bulut, M.; Pant, D. Technological advances in CO2 conversion electro-biorefinery: A step toward commercialization. Bioresour. Technol. 2016, 215, 357–370. [Google Scholar] [CrossRef]
- Menetrez, M.Y. An overview of algae biofuel production and potential environmental impact. Environ. Sci. Technol. 2012, 46, 7073–7085. [Google Scholar] [CrossRef] [PubMed]
- Dębowski, M.; Zieliński, M.; Kazimierowicz, J.; Kujawska, N.; Talbierz, S. Microalgae cultivation technologies as an opportunity for bioenergetic system development—Advantages and limitations. Sustainability 2020, 12, 9980. [Google Scholar] [CrossRef]
- Rosch, C.; Rossmann, M.; Weickert, S. Microalgae for integrated food and fuel production. GCB Bioenergy 2019, 11, 326–334. [Google Scholar] [CrossRef]
- Bhola, V.; Swalaha, F.; Kumar, R.R.; Singh, M.; Bux, F. Overview of the potential of microalgae for CO2 sequestration. Int. J. Environ. Sci. Technol. 2014, 11, 2103–2118. [Google Scholar] [CrossRef] [Green Version]
- Botte, P.; d’Ippolito, G.; Gallo, C.; Sardo, A.; Fontana, A. Combined exploitation of CO2 and nutrient replenishment for increasing biomass and lipid productivity of the marine diatoms Thalassiosira weissflogii and Cyclotella cryptica. J. Appl. Phycol. 2018, 30, 243–251. [Google Scholar] [CrossRef]
- Sepulveda, C.; Gomez, C.; Bahraoui, N.E.L.; Acien, G. Comparative evaluation of microalgae strains for CO2 capture purposes. J. CO2 Util. 2019, 30, 158–167. [Google Scholar] [CrossRef]
- Van den Hende, S.; Vervaeren, H.; Boon, N. Flue gas compounds and microalgae: (Bio-)chemical interactions leading to biotechnological opportunities. Biotechnol. Adv. 2012, 30, 1405–1424. [Google Scholar] [CrossRef] [PubMed]
- Chew, K.W.; Yap, J.Y.; Show, P.L.; Suan, N.H.; Juan, J.C.; Ling, T.C.; Lee, D.J.; Chang, J.S. Microalgae biorefinery: High value products perspectives. Bioresour. Technol. 2017, 229, 53–62. [Google Scholar] [CrossRef]
- Kapoor, B.; Kapoor, D.; Gautam, S.; Singh, R.; Bhardwaj, S. Dietary polyunsaturated fatty acids (PUFAs): Uses and potential health benefits. Curr. Nutr. Rep. 2021, 10, 232–242. [Google Scholar] [CrossRef]
- Mason, R.P.; Libby, P.; Bhatt, D.L. Emerging mechanisms of cardiovascular protection for the Omega-3 fatty acid eicosapentaenoic acid. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 1135–1147. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Prasad, P.; Sreedhar, R.V.; Akhilender Naidu, K.; Shang, X.; Keum, Y.-S. Omega–3 polyunsaturated fatty acids (PUFAs): Emerging plant and microbial sources, oxidative stability, bioavailability, and health benefits—A review. Antioxidants 2021, 10, 1627. [Google Scholar] [CrossRef]
- Ghasemi Fard, S.; Wang, F.L.; Sinclair, A.J.; Elliott, G.; Turchini, G.M. How does high DHA fish oil affect health? A systematic review of evidence. Crit. Rev. Food Sci. Nutr. 2019, 59, 1684–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khozin-Goldberg, I.; Iskandarov, U.; Cohen, Z. LC-PUFA from photosynthetic microalgae: Occurrence, biosynthesis, and prospects in biotechnology. Appl. Microbiol. Biotechnol. 2011, 91, 905–915. [Google Scholar] [CrossRef]
- Makrides, M.; Gibson, R.A. Are long-chain polyunsaturated fatty acids essential nutrients in infancy? In Proceedings of the 4th International Congress on Essential Fatty Acids and Eicosanoids, Edinburgh, UK, 20–24 July 1997; pp. 136–140. [Google Scholar]
- Simopoulos, A.P. An increase in the Omega-6/Omega-3 fatty acid ratio increases the risk for obesity. Nutrients 2016, 8, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yates, C.M.; Calder, P.C.; Rainger, G.E. Pharmacology and therapeutics of omega-3 polyunsaturated fatty acids in chronic inflammatory disease. Pharmacol. Ther. 2014, 141, 272–282. [Google Scholar] [CrossRef]
- Abedi, E.; Sahari, M.A. Long-chain polyunsaturated fatty acid sources and evaluation of their nutritional and functional properties. Food Sci. Nutr. 2014, 2, 443–463. [Google Scholar] [CrossRef]
- Klinger, D.; Naylor, R. Searching for solutions in aquaculture: Charting a sustainable course. Annu. Rev. Environ. Resour. 2012, 37, 247–276. [Google Scholar] [CrossRef] [Green Version]
- Qi, S.B.; Zhao, X.W.; Zhang, W.J.; Wang, C.H.; He, M.L.; Chang, Y.Q.; Ding, J. The effects of 3 different microalgae species on the growth, metamorphosis and MYP gene expression of two sea urchins, Strongylocentrotus intermedius and S. nudus. Aquaculture 2018, 492, 123–131. [Google Scholar] [CrossRef]
- Leal, E.; de Beyer, L.; O’Connor, W.; Dove, M.; Ralph, P.J.; Pernice, M. Production optimisation of Tisochrysis lutea as a live feed for juvenile Sydney rock oysters, Saccostrea glomerata, using large-scale photobioreactors. Aquaculture 2021, 533, 736077. [Google Scholar] [CrossRef]
- Gachelin, M.; Boutoute, M.; Carrier, G.; Talec, A.; Pruvost, E.; Guiheneuf, F.; Bernard, O.; Sciandra, A. Enhancing PUFA-rich polar lipids in Tisochrysis lutea using adaptive laboratory evolution (ALE) with oscillating thermal stress. Appl. Microbiol. Biotechnol. 2021, 105, 301–312. [Google Scholar] [CrossRef]
- Jethani, H.; Patel, P.; Mudliar, S.N.; Sarada, R.; Chauhan, V.S. Growth and biochemical response of an indigenous oleaginous microalga Scenedesmus obtusus cultivated in outdoor open ponds. Indian J. Exp. Biol. 2019, 57, 40–49. [Google Scholar]
- Almeyda, M.D.; Bilbao, P.G.S.; Popovich, C.A.; Constenla, D.; Leonardi, P.I. Enhancement of polyunsaturated fatty acid production under low-temperature stress in Cylindrotheca closterium. J. Appl. Phycol. 2020, 32, 989–1001. [Google Scholar] [CrossRef]
- Pasquet, V.; Ulmann, L.; Mimouni, V.; Guiheneuf, F.; Jacquette, B.; Morant-Manceau, A.; Tremblin, G. Fatty acids profile and temperature in the cultured marine diatom Odontella aurita. J. Appl. Phycol. 2014, 26, 2265–2271. [Google Scholar] [CrossRef]
- Chang, J.Y.; Le, K.; Song, X.Q.; Jiao, K.L.; Zeng, X.H.; Ling, X.P.; Shi, T.; Tang, X.; Sun, Y.; Lin, L. Scale-up cultivation enhanced arachidonic acid accumulation by red microalgae Porphyridium purpureum. Bioprocess Biosyst. Eng. 2017, 40, 1763–1773. [Google Scholar] [CrossRef]
- Qiao, H.J.; Cong, C.; Sun, C.X.; Li, B.S.; Wang, J.Y.; Zhang, L.M. Effect of culture conditions on growth, fatty acid composition and DHA/EPA ratio of Phaeodactylum tricornutum. Aquaculture 2016, 452, 311–317. [Google Scholar] [CrossRef]
- Krishnan, A.; Ananda, R.; Joseph, A. Culture medium and growth phase modulate the fatty acid composition of the diatom Nitzschia palea (Kutzing) W. Smith—Potential source for live feed and biodiesel. Fish. Technol. 2020, 57, 28–35. [Google Scholar]
- Fakhry, E.M.; El Maghraby, D.M. Lipid accumulation in response to nitrogen limitation and variation of temperature in Nannochloropsis salina. Bot. Stud. 2015, 56, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solovchenko, A.E.; Gorelova, O.A.; Baulina, O.I.; Selyakh, I.O.; Semenova, L.R.; Chivkunova, O.B.; Scherbakov, P.N.; Lobakova, E.S. Physiological plasticity of symbiotic Desmodesmus (Chlorophyceae) isolated from taxonomically distant white sea invertibrates. Russ. J. Plant Physiol. 2015, 62, 653–663. [Google Scholar] [CrossRef]
- Schreiber, C.; Behrendt, D.; Huber, G.; Pfaff, C.; Widzgowski, J.; Ackermann, B.; Muller, A.; Zachleder, V.; Moudrikova, S.; Mojzes, P.; et al. Growth of algal biomass in laboratory and in large-scale algal photobioreactors in the temperate climate of western Germany. Bioresour. Technol. 2017, 234, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Pal, D.; Khozin-Goldberg, I.; Cohen, Z.; Boussiba, S. The effect of light, salinity, and nitrogen availability on lipid production by Nannochloropsis sp. Appl. Microbiol. Biotechnol. 2011, 90, 1429–1441. [Google Scholar] [CrossRef]
- Su, C.-H.; Chien, L.-J.; Gomes, J.; Lin, Y.-S.; Yu, Y.; Liou, J.S.; Syu, R.-J. Factors affecting lipid accumulation by Nannochloropsis oculata in a two-stage cultivation process. J. Appl. Phycol. 2010, 23, 903–908. [Google Scholar] [CrossRef]
- Guedes, A.C.; Meireles, L.A.; Amaro, H.M.; Malcata, F.X. Changes in lipid class and fatty acid composition of cultures of Pavlova lutheri, in response to light intensity. J. Am. Oil Chem. Soc. 2010, 87, 791–801. [Google Scholar] [CrossRef]
- Wang, X.X.; Fosse, H.K.; Li, K.S.; Chauton, M.S.; Vadstein, O.; Reitan, K.I. Influence of nitrogen limitation on lipid accumulation and EPA and DHA content in four marine microalgae for possible use in aquafeed. Front. Mar. Sci. 2019, 6, 95. [Google Scholar] [CrossRef]
- Breuer, G.; Lamers, P.P.; Martens, D.E.; Draaisma, R.B.; Wijffels, R.H. The impact of nitrogen starvation on the dynamics of triacylglycerol accumulation in nine microalgae strains. Bioresour. Technol. 2012, 124, 217–226. [Google Scholar] [CrossRef]
- Li, T.; Xu, J.; Wu, H.B.; Jiang, P.L.; Chen, Z.S.; Xiang, W.Z. Growth and biochemical composition of Porphyridium purpureum SCS-02 under different nitrogen concentrations. Mar. Drugs 2019, 17, 124. [Google Scholar] [CrossRef] [Green Version]
- Su, G.M.; Jiao, K.L.; Li, Z.; Guo, X.Y.; Chang, J.Y.; Ndikubwimana, T.; Sun, Y.; Zeng, X.H.; Lu, Y.H.; Lin, L. Phosphate limitation promotes unsaturated fatty acids and arachidonic acid biosynthesis by microalgae Porphyridium purpureum. Bioprocess Biosyst. Eng. 2016, 39, 1129–1136. [Google Scholar] [CrossRef]
- Khozin-Goldberg, I.; Cohen, Z. The effect of phosphate starvation on the lipid and fatty acid composition of the fresh water eustigmatophyte Monodus subterraneus. Phytochemistry 2006, 67, 696–701. [Google Scholar] [CrossRef]
- Sabu, S.; Singh, I.S.B.; Joseph, V. Improved lipid production in oleaginous brackish diatom Navicula phyllepta MACC8 using two-stage cultivation approach. 3 Biotech 2019, 9, 437. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Wu, M.; Fu, Q.Q.; Li, X.S.; Xu, J.T. A two-stage model with nitrogen and silicon limitation enhances lipid productivity and biodiesel features of the marine bloom-forming diatom Skeletonema costatum. Bioresour. Technol. 2019, 289, 121717. [Google Scholar] [CrossRef]
- Boelen, P.; van Mastrigt, A.; van de Bovenkamp, H.H.; Heeres, H.J.; Buma, A.G.J. Growth phase significantly decreases the DHA-to-EPA ratio in marine microalgae. Aquac. Int. 2017, 25, 577–587. [Google Scholar] [CrossRef] [Green Version]
- Adarme-Vega, T.C.; Thomas-Hall, S.R.; Lim, D.K.Y.; Schenk, P.M. Effects of long chain fatty acid synthesis and associated gene expression in microalga Tetraselmis sp. Mar. Drugs 2014, 12, 3381–3398. [Google Scholar] [CrossRef] [Green Version]
- Dammak, M.; Haase, S.M.; Miladi, R.; Ben Amor, F.; Barkallah, M.; Gosset, D.; Pichon, C.; Huchzermeyer, B.; Fendri, I.; Denis, M.; et al. Enhanced lipid and biomass production by a newly isolated and identified marine microalga. Lipids Health Dis. 2016, 15, 209. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Roldan, A.J.; Perales-Vela, H.V.; Canizares-Villanueva, R.O.; Torzillo, G. Physiological response of Nannochloropsis sp. to saline stress in laboratory batch cultures. J. Appl. Phycol. 2014, 26, 115–121. [Google Scholar] [CrossRef]
- Balzano, S.; Villanueva, L.; de Bar, M.; Sinninghe Damsté, J.S.; Schouten, S. Impact of culturing conditions on the abundance and composition of long chain alkyl diols in species of the genus Nannochloropsis. Org. Geochem. 2017, 108, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Rearte, T.A.; Figueroa, F.L.; Gomez-Serrano, C.; Velez, C.G.; Marsili, S.; Iorio, A.D.; Gonzalez-Lopez, C.V.; Ceron-Garcia, M.C.; Abdala-Diaz, R.T.; Acien-Fernandez, F.G. Optimization of the production of lipids and carotenoids in the microalga Golenkinia aff. brevispicula. Algal Res. Biomass Biofuels Bioprod. 2020, 51, 102004. [Google Scholar] [CrossRef]
- Cepak, V.; Pribyl, P.; Kohoutkova, J.; Kastanek, P. Optimization of cultivation conditions for fatty acid composition and EPA production in the eustigmatophycean microalga Trachydiscus minutus. J. Appl. Phycol. 2014, 26, 181–190. [Google Scholar] [CrossRef]
- Atikij, T.; Syaputri, Y.; Iwahashi, H.; Praneenararat, T.; Sirisattha, S.; Kageyama, H.; Waditee-Sirisattha, R. Enhanced lipid production and molecular dynamics under salinity stress in green microalga Chlamydomonas reinhardtii (137C). Mar. Drugs 2019, 17, 484. [Google Scholar] [CrossRef] [Green Version]
- Conceicao, D.; Lopes, R.G.; Derner, R.B.; Cella, H.; do Carmo, A.P.B.; D’Oca, M.G.M.; Petersen, R.; Passos, M.F.; Vargas, J.V.C.; Galli-Terasawa, L.V.; et al. The effect of light intensity on the production and accumulation of pigments and fatty acids in Phaeodactylum tricornutum. J. Appl. Phycol. 2020, 32, 1017–1025. [Google Scholar] [CrossRef]
- Russo, G.L.; Langellotti, A.L.; Oliviero, M.; Sacchi, R.; Masi, P. Sustainable production of food grade omega-3 oil using aquatic protists: Reliability and future horizons. New Biotechnol. 2021, 62, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Perez-Garcia, O.; Bashan, Y. Microalgal heterotrophic and mixotrophic culturing for bio-refining: From metabolic routes to techno-economics. In Algal Biorefineries: Products and Refinery Design; Prokop, A., Bajpai, R.K., Zappi, M.E., Eds.; Springer: Cham, Switzerland, 2015; pp. 61–131. [Google Scholar]
- Zhu, L.Y.; Zhang, X.C.; Ji, L.; Song, X.J.; Kuang, C.H. Changes of lipid content and fatty acid composition of Schizochytrium limacinum in response to different temperatures and salinities. Process Biochem. 2007, 42, 210–214. [Google Scholar] [CrossRef]
- Barone, R.; De Napoli, L.; Mayol, L.; Paolucci, M.; Volpe, M.G.; D’Elia, L.; Pollio, A.; Guida, M.; Gambino, E.; Carraturo, F.; et al. Autotrophic and heterotrophic growth conditions modify biomolecole production in the microalga Galdieria sulphuraria (Cyanidiophyceae, Rhodophyta). Mar. Drugs 2020, 18, 169. [Google Scholar] [CrossRef] [Green Version]
- Cupo, A.; Landi, S.; Morra, S.; Nuzzo, G.; Gallo, C.; Manzo, E.; Fontana, A.; D’Ippolito, G. Autotrophic vs. heterotrophic cultivation of the marine diatom Cyclotella cryptica for EPA production. Mar. Drugs 2021, 19, 355. [Google Scholar] [CrossRef]
- Selvakumar, P.; Umadevi, K. Enhanced lipid and fatty acid content under photoheterotrophic condition in the mass cultures of Tetraselmis gracilis and Platymonas convolutae. Algal Res. Biomass Biofuels Bioprod. 2014, 6, 180–185. [Google Scholar] [CrossRef]
- Oliver, L.; Dietrich, T.; Maranon, I.; Villaran, M.C.; Barrio, R.J. Producing Omega-3 polyunsaturated fatty acids: A review of sustainable sources and future trends for the EPA and DHA market. Resources 2020, 9, 148. [Google Scholar] [CrossRef]
- Rawat, I.; Ranjith Kumar, R.; Mutanda, T.; Bux, F. Dual role of microalgae: Phycoremediation of domestic wastewater and biomass production for sustainable biofuels production. Appl. Energy 2011, 88, 3411–3424. [Google Scholar] [CrossRef]
- Koutra, E.; Mastropetros, S.G.; Ali, S.S.; Tsigkou, K.; Kornaros, M. Assessing the potential of Chlorella vulgaris for valorization of liquid digestates from agro-industrial and municipal organic wastes in a biorefinery approach. J. Clean. Prod. 2021, 280, 124352. [Google Scholar] [CrossRef]
- Ljubic, A.; Safafar, H.; Holdt, S.L.; Jacobsen, C. Biomass composition of Arthrospira platensis during cultivation on industrial process water and harvesting. J. Appl. Phycol. 2018, 30, 943–954. [Google Scholar] [CrossRef] [Green Version]
- Kujawska, N.; Talbierz, S.; Dębowski, M.; Kazimierowicz, J.; Zieliński, M. Cultivation method effect on Schizochytrium sp. biomass growth and docosahexaenoic acid (DHA) production with the use of waste glycerol as a source of organic carbon. Energies 2021, 14, 2952. [Google Scholar] [CrossRef]
- Kujawska, N.; Talbierz, S.; Dębowski, M.; Kazimierowicz, J.; Zieliński, M. Optimizing docosahexaenoic acid (DHA) production by Schizochytrium sp. grown on waste glycerol. Energies 2021, 14, 1685. [Google Scholar] [CrossRef]
- Vargas, M.A.; Rodriguez, H.; Moreno, J.; Olivares, H.; Del Campo, J.A.; Rivas, J.; Guerrero, M.G. Biochemical composition and fatty acid content of filamentous nitrogen-fixing cyanobacteria. J. Phycol. 1998, 34, 812–817. [Google Scholar] [CrossRef]
- Tang, D.H.; Han, W.; Li, P.L.; Miao, X.L.; Zhong, J.J. CO2 biofixation and fatty acid composition of Scenedesmus obliquus and Chlorella pyrenoidosa in response to different CO2 levels. Bioresour. Technol. 2011, 102, 3071–3076. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.L.; Meng, F.P.; Wang, Y.J.; Yi, X.Y.; Cui, H.W. Effect of pH, temperature, and CO2 concentration on growth and lipid accumulation of Nannochloropsis sp. MASCC 11. J. Ocean. Univ. China 2020, 19, 1183–1192. [Google Scholar] [CrossRef]
- Kareya, M.S.; Mariam, I.; Shaikh, K.M.; Nesamma, A.A.; Jutur, P.P. Photosynthetic carbon partitioning and metabolic regulation in response to very-low and high CO2 in Microchloropsis gaditana NIES 2587. Front. Plant Sci. 2020, 11, 981. [Google Scholar] [CrossRef]
- Liang, M.H.; Jiang, J.G. Advancing oleaginous microorganisms to produce lipid via metabolic engineering technology. Prog. Lipid Res. 2013, 52, 395–408. [Google Scholar] [CrossRef]
- Goncalves, E.C.; Wilkie, A.C.; Kirst, M.; Rathinasabapathi, B. Metabolic regulation of triacylglycerol accumulation in the green algae: Identification of potential targets for engineering to improve oil yield. Plant Biotechnol. J. 2016, 14, 1649–1660. [Google Scholar] [CrossRef]
- Kindle, K.L.; Schnell, R.A.; Fernandez, E.; Lefebvre, P.A. Stable nuclear transformation of Chlamydomonas using the Chlamydomonas gene for nitrate reductase. J. Cell Biol. 1989, 109, 2589–2601. [Google Scholar] [CrossRef]
- Kindle, K.L. High-frequency nuclear transformation of Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 1990, 87, 1228–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karas, B.J.; Diner, R.E.; Lefebvre, S.C.; McQuaid, J.; Phillips, A.P.R.; Noddings, C.M.; Brunson, J.K.; Valas, R.E.; Deerinck, T.J.; Jablanovic, J.; et al. Designer diatom episomes delivered by bacterial conjugation. Nat. Commun. 2015, 6, 7925. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.V.; Misquitta, R.W.; Reddy, V.S.; Rao, B.J.; Rajam, M.V. Genetic transformation of the green alga Chlamydomonas reinhardtii by Agrobacterium tumefaciens. Plant Sci. 2004, 166, 731–738. [Google Scholar] [CrossRef]
- Shimogawara, K.; Fujiwara, S.; Grossman, A.; Usuda, H. High-efficiency transformation of Chlamydomonas reinhardtii by electroporation. Genetics 1998, 148, 1821–1828. [Google Scholar] [CrossRef] [PubMed]
- Arora, N.; Yen, H.W.; Philippidis, G.P. Harnessing the power of mutagenesis and adaptive laboratory evolution for high lipid production by oleaginous microalgae and yeasts. Sustainability 2020, 12, 5125. [Google Scholar] [CrossRef]
- Fajardo, C.; De Donato, M.; Carrasco, R.; Martinez-Rodriguez, G.; Mancera, J.M.; Fernandez-Acero, F.J. Advances and challenges in genetic engineering of microalgae. Rev. Aquac. 2020, 12, 365–381. [Google Scholar] [CrossRef]
- Liang, M.H.; Wang, L.; Wang, Q.M.; Zhu, J.H.; Jiang, J.G. High-value bioproducts from microalgae: Strategies and progress. Crit. Rev. Food Sci. Nutr. 2019, 59, 2423–2441. [Google Scholar] [CrossRef]
- Fang, Y.X.; Stroukov, W.; Cathomen, T.; Mussolino, C. Chimerization enables gene synthesis and lentiviral delivery of customizable TALE-based effectors. Int. J. Mol. Sci. 2020, 21, 795. [Google Scholar] [CrossRef] [Green Version]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. CRISPR provides acquired resistance against viruses in prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef]
- Balamurugan, S.; Wang, X.; Wang, H.L.; An, C.J.; Li, H.; Li, D.W.; Yang, W.D.; Liu, J.S.; Li, H.Y. Occurrence of plastidial triacylglycerol synthesis and the potential regulatory role of AGPAT in the model diatom Phaeodactylum tricornutum. Biotechnol. Biofuels 2017, 10, 97. [Google Scholar] [CrossRef]
- Niu, Y.F.; Wang, X.; Hu, D.X.; Balamurugan, S.; Li, D.W.; Yang, W.D.; Liu, J.S.; Li, H.Y. Molecular characterization of a glycerol-3-phosphate acyltransferase reveals key features essential for triacylglycerol production in Phaeodactylum tricornutum. Biotechnol. Biofuels 2016, 9, 60. [Google Scholar] [CrossRef] [Green Version]
- Niu, Y.F.; Zhang, M.H.; Li, D.W.; Yang, W.D.; Liu, J.S.; Bai, W.B.; Li, H.Y. Improvement of neutral lipid and polyunsaturated fatty acid biosynthesis by overexpressing a type 2 diacylglycerol acyltransferase in marine diatom Phaeodactylum tricornutum. Mar. Drugs 2013, 11, 4558–4569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinamarca, J.; Levitan, O.; Kumaraswamy, G.K.; Lun, D.S.; Falkowski, P.G. Overespresssion of a diacylglycerol acyltransferase gene in Phaeodactylum tricornutum directs carbon towards lipid biosynthesis. J. Phycol. 2017, 53, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Li, D.W.; Cen, S.Y.; Liu, Y.H.; Balamurugan, S.; Zheng, X.Y.; Alimujiang, A.; Yang, W.D.; Liu, J.S.; Li, H.Y. A type 2 diacylglycerol acyltransferase accelerates the triacylglycerol biosynthesis in heterokont oleaginous microalga Nannochloropsis oceanica. J. Biotechnol. 2016, 229, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.H.; Shi, Y.; Ma, X.N.; Pan, Y.F.; Hu, H.H.; Li, Y.T.; Luo, M.; Gerken, H.; Liu, J. A type-I diacylglycerol acyltransferase modulates triacylglycerol biosynthesis and fatty acid composition in the oleaginous microalga, Nannochloropsis oceanica. Biotechnol. Biofuels 2017, 10, 174. [Google Scholar] [CrossRef]
- La Russa, M.; Bogen, C.; Uhmeyer, A.; Doebbe, A.; Filippone, E.; Kruse, O.; Mussgnug, J.H. Functional analysis of three type-2 DGAT homologue genes for triacylglycerol production in the green microalga Chlamydomonas reinhardtii. J. Biotechnol. 2012, 162, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Iwai, M.; Ikeda, K.; Shimojima, M.; Ohta, H. Enhancement of extraplastidic oil synthesis in Chlamydomonas reinhardtii using a type-2 diacylglycerol acyltransferase with a phosphorus starvation-inducible promoter. Plant Biotechnol. J. 2014, 12, 808–819. [Google Scholar] [CrossRef] [Green Version]
- Zou, L.G.; Chen, J.W.; Zheng, D.L.; Balamurugan, S.; Li, D.W.; Yang, W.D.; Liu, J.S.; Li, H.Y. High-efficiency promoter-driven coordinated regulation of multiple metabolic nodes elevates lipid accumulation in the model microalga Phaeodactylum tricornutum. Microb. Cell Factories 2018, 17, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muto, M.; Tanaka, M.; Liang, Y.; Yoshino, T.; Matsumoto, M.; Tanaka, T. Enhancement of glycerol metabolism in the oleaginous marine diatom Fistulifera solaris JPCC DA0580 to improve triacylglycerol productivity. Biotechnol. Biofuels 2015, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Lu, Y.; Peng, K.T.; Huang, T.; Niu, Y.F.; Xie, W.H.; Yang, W.D.; Liu, J.S.; Li, H.Y. Glycerol and neutral lipid production in the oleaginous marine diatom Phaeodactylum tricornutum promoted by overexpression of glycerol-3-phosphate dehydrogenase. Biotechnol. Biofuels 2014, 7, 110. [Google Scholar] [CrossRef] [Green Version]
- Yamaoka, Y.; Achard, D.; Jang, S.; Legéret, B.; Kamisuki, S.; Ko, D.; Schulz-Raffelt, M.; Kim, Y.; Song, W.Y.; Nishida, I.; et al. Identification of a Chlamydomonas plastidial 2-lysophosphatidic acid acyltransferase and its use to engineer microalgae with increased oil content. Plant Biotechnol. J. 2016, 14, 2158–2167. [Google Scholar] [CrossRef]
- Gong, Y.M.; Guo, X.J.; Wan, X.; Liang, Z.; Jiang, M. Characterization of a novel thioesterase (PtTE) from Phaeodactylum tricornutum. J. Basic Microbiol. 2011, 51, 666–672. [Google Scholar] [CrossRef] [PubMed]
- Lei, A.P.; Chen, H.; Shen, G.M.; Hu, Z.L.; Chen, L.; Wang, J.X. Expression of fatty acid synthesis genes and fatty acid accumulation in Haematococcus pluvialis under different stressors. Biotechnol. Biofuels 2012, 5, 18. [Google Scholar] [CrossRef] [Green Version]
- Xue, J.; Niu, Y.F.; Huang, T.; Yang, W.D.; Liu, J.S.; Li, H.Y. Genetic improvement of the microalga Phaeodactylum tricornutum for boosting neutral lipid accumulation. Metab. Eng. 2015, 27, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.W.; Liu, W.J.; Hu, D.X.; Wang, X.; Balamurugan, S.; Alimujiang, A.; Yang, W.D.; Liu, J.S.; Li, H.Y. Identification of a malonyl CoA-acyl carrier protein transacylase and its regulatory role in fatty acid biosynthesis in oleaginous microalga Nannochloropsis oceanica. Biotechnol. Appl. Biochem. 2017, 64, 620–626. [Google Scholar] [CrossRef]
- Li, Z.P.; Meng, T.; Ling, X.P.; Li, J.; Zheng, C.Q.; Shi, Y.Y.; Chen, Z.Y.; Li, Z.Q.; Li, Q.B.; Lu, Y.H.; et al. Overexpression of malonyl-CoA: ACP transacylase in Schizochytrium sp. to improve polyunsaturated fatty acid production. J. Agric. Food Chem. 2018, 66, 5382–5391. [Google Scholar] [CrossRef] [PubMed]
- Cook, O.; Hildebrand, M. Enhancing LC-PUFA production in Thalassiosira pseudonana by overexpressing the endogenous fatty acid elongase genes. J. Appl. Phycol. 2016, 28, 897–905. [Google Scholar] [CrossRef]
- Peng, K.T.; Zheng, C.N.; Xue, J.; Chen, X.Y.; Yang, W.D.; Liu, J.S.; Bai, W.B.; Li, H.Y. Delta 5 fatty acid desaturase upregulates the synthesis of polyunsaturated fatty acids in the marine diatom Phaeodactylum tricornutum. J. Agric. Food Chem. 2014, 62, 8773–8776. [Google Scholar] [CrossRef]
- Kaye, Y.; Grundman, O.; Leu, S.; Zarka, A.; Zorin, B.; Didi-Cohen, S.; Khozin-Goldberg, I.; Boussiba, S. Metabolic engineering toward enhanced LC-PUFA biosynthesis in Nannochloropsis oceanica: Overexpression of endogenous Delta 12 desaturase driven by stress-inducible promoter leads to enhanced deposition of polyunsaturated fatty acids in TAG. Algal Res. Biomass Biofuels Bioprod. 2015, 11, 387–398. [Google Scholar] [CrossRef]
- Kang, N.K.; Jeon, S.; Kwon, S.; Koh, H.G.; Shin, S.E.; Lee, B.; Choi, G.G.; Yang, J.W.; Jeong, B.R.; Chang, Y.K. Effects of overexpression of a bHLH transcription factor on biomass and lipid production in Nannochloropsis salina. Biotechnol. Biofuels 2015, 8, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibanez-Salazar, A.; Rosales-Mendoza, S.; Rocha-Uribe, A.; Ramirez-Alonso, J.I.; Lara-Hernandez, I.; Hernandez-Torres, A.; Paz-Maldonado, L.M.T.; Silva-Ramirez, A.S.; Banuelos-Hernandez, B.; Martinez-Salgado, J.L.; et al. Over-expression of Dof-type transcription factor increases lipid production in Chlamydomonas reinhardtii. J. Biotechnol. 2014, 184, 27–38. [Google Scholar] [CrossRef]
- Salas-Montantes, C.J.; Gonzalez-Ortega, O.; Ochoa-Alfaro, A.E.; Camarena-Rangel, R.; Paz-Maldonado, L.M.T.; Rosales-Mendoza, S.; Rocha-Uribe, A.; Soria-Guerra, R.E. Lipid accumulation during nitrogen and sulfur starvation in Chlamydomonas reinhardtii overexpressing a transcription factor. J. Appl. Phycol. 2018, 30, 1721–1733. [Google Scholar] [CrossRef]
- Bajhaiya, A.K.; Dean, A.P.; Zeef, L.A.H.; Webster, R.E.; Pittman, J.K. PSR1 is a global transcriptional regulator of phosphorus deficiency responses and carbon storage metabolism in Chlamydomonas reinhardtii. Plant Physiol. 2016, 170, 1216–1234. [Google Scholar] [CrossRef] [Green Version]
- Ngan, C.Y.; Wong, C.H.; Choi, C.; Yoshinaga, Y.; Louie, K.; Jia, J.; Chen, C.; Bowen, B.; Cheng, H.Y.; Leonelli, L.; et al. Lineage-specific chromatin signatures reveal a regulator of lipid metabolism in microalgae. Nat. Plants 2015, 1, 107. [Google Scholar] [CrossRef]
- Kwon, S.; Kang, N.K.; Koh, H.G.; Shin, S.E.; Lee, B.; Jeong, B.R.; Chang, Y.K. Enhancement of biomass and lipid productivity by overexpression of a bZIP transcription factor in Nannochloropsis salina. Biotechnol. Bioeng. 2018, 115, 331–340. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wei, W.; Li, N.J.; Yuan, W.S.Q.; Ding, Y.; Yang, W.D.; Liu, J.S.; Balamurugan, S.; Li, H.Y. Heterogeneous expression of human PNPLA3 triggers algal lipid accumulation and lipid droplet enlargement. Algal Res. Biomass Biofuels Bioprod. 2018, 31, 276–281. [Google Scholar] [CrossRef]
- Wang, X.; Hao, T.B.; Balamurugan, S.; Yang, W.D.; Liu, J.S.; Dong, H.P.; Li, H.Y. A lipid droplet-associated protein involved in lipid droplet biogenesis and triacylglycerol accumulation in the oleaginous microalga Phaeodactylum tricornutum. Algal Res. Biomass Biofuels Bioprod. 2017, 26, 215–224. [Google Scholar] [CrossRef]
- Dunahay, T.G.; Jarvis, E.E.; Dais, S.S.; Roessler, P.G. Manipulation of microalgal lipid production using genetic engineering. Appl. Biochem. Biotechnol. 1996, 57–58, 223–231. [Google Scholar] [CrossRef]
- Xue, J.; Balamurugan, S.; Li, D.W.; Liu, Y.H.; Zeng, H.; Wang, L.; Yang, W.D.; Liu, J.S.; Li, H.Y. Glucose-6-phosphate dehydrogenase as a target for highly efficient fatty acid biosynthesis in microalgae by enhancing NADPH supply. Metab. Eng. 2017, 41, 212–221. [Google Scholar] [CrossRef]
- Marechal, E.; Dolch, L. Increased Triacylglycerol Production in Microalgae. WO Patent 2017/129777 A1, 3 August 2017. [Google Scholar]
- Li, Y.T.; Han, D.X.; Hu, G.R.; Dauvillee, D.; Sommerfeld, M.; Ball, S.; Hua, Q. Chlamydomonas starchless mutant defective in ADP-glucose pyrophosphorylase hyper-accumulates triacylglycerol. Metab. Eng. 2010, 12, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Work, V.H.; Radakovits, R.; Jinkerson, R.E.; Meuser, J.E.; Elliott, L.G.; Vinyard, D.J.; Laurens, L.M.L.; Dismukes, G.C.; Posewitz, M.C. Increased lipid accumulation in the Chlamydomonas reinhardtii sta7-10 starchless isoamylase mutant and increased carbohydrate synthesis in complemented strains. Eukaryot. Cell 2010, 9, 1251–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daboussi, F.; Leduc, S.; Marechal, A.; Dubois, G.; Guyot, V.; Perez-Michaut, C.; Amato, A.; Falciatore, A.; Juillerat, A.; Beurdeley, M.; et al. Genome engineering empowers the diatom Phaeodactylum tricornutum for biotechnology. Nat. Commun. 2014, 5, 4831. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.H.; Shi, H.P.; Yang, G.P.; Lv, N.N.; Yang, M.; Pan, K.H. Silencing UDP-glucose pyrophosphorylase gene in Phaeodactylum tricornutum affects carbon allocation. New Biotechnol. 2016, 33, 237–244. [Google Scholar] [CrossRef]
- Hildebrand, M.; Manandhar-Shrestha, K.; Abbriano, R. Effects of chrysolaminarin synthase knockdown in the diatom Thalassiosira pseudonana: Implications of reduced carbohydrate storage relative to green algae. Algal Res. Biomass Biofuels Bioprod. 2017, 23, 66–77. [Google Scholar] [CrossRef]
- Breuer, G.; de Jaeger, L.; Artus, V.P.G.; Martens, D.E.; Springer, J.; Draaisma, R.B.; Eggink, G.; Wijffels, R.H.; Lamers, P.P. Superior triacylglycerol (TAG) accumulation in starchless mutants of Scenedesmus obliquus: (II) evaluation of TAG yield and productivity in controlled photobioreactors. Biotechnol. Biofuels 2014, 7, 70. [Google Scholar] [CrossRef] [Green Version]
- Kao, P.H.; Ng, I.S. CRISPRi mediated phosphoenolpyruvate carboxylase regulation to enhance the production of lipid in Chlamydomonas reinhardtii. Bioresour. Technol. 2017, 245, 1527–1537. [Google Scholar] [CrossRef]
- Deng, X.D.; Cai, J.J.; Li, Y.J.; Fei, X.W. Expression and knockdown of the PEPC1 gene affect carbon flux in the biosynthesis of triacylglycerols by the green alga Chlamydomonas reinhardtii. Biotechnol. Lett. 2014, 36, 2199–2208. [Google Scholar] [CrossRef]
- Wang, C.G.; Chen, X.; Li, H.; Wang, J.X.; Hu, Z.L. Artificial miRNA inhibition of phosphoenolpyruvate carboxylase increases fatty acid production in a green microalga Chlamydomonas reinhardtii. Biotechnol. Biofuels 2017, 10, 91. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Pan, Y.F.; Bowler, C.; Zhang, L.X.; Hu, H.H. Knockdown of phosphoenolpyruvate carboxykinase increases carbon flux to lipid synthesis in Phaeodactylum tricornutum. Algal Res. Biomass Biofuels Bioprod. 2016, 15, 50–58. [Google Scholar] [CrossRef]
- Ma, Y.H.; Wang, X.; Niu, Y.F.; Yang, Z.K.; Zhang, M.H.; Wang, Z.M.; Yang, W.D.; Liu, J.S.; Li, H.Y. Antisense knockdown of pyruvate dehydrogenase kinase promotes the neutral lipid accumulation in the diatom Phaeodactylum tricornutum. Microb. Cell Factories 2014, 13, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trentacoste, E.M.; Shrestha, R.P.; Smith, S.R.; Gle, C.; Hartmann, A.C.; Hildebrand, M.; Gerwick, W.H. Metabolic engineering of lipid catabolism increases microalgal lipid accumulation without compromising growth. Proc. Natl. Acad. Sci. USA 2013, 110, 19748–19753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.L.; Pan, Y.F.; Hu, H.H. Identification of the triacylglycerol lipase in the chloroplast envelope of the diatom Phaeodactylum tricornutum. Algal Res. Biomass Biofuels Bioprod. 2018, 33, 440–447. [Google Scholar] [CrossRef]
- Barka, F.; Angstenberger, M.; Ahrendt, T.; Lorenzen, W.; Bode, H.B.; Buchel, C. Identification of a triacylglycerol lipase in the diatom Phaeodactylum tricornutum. Biochim. Biophys. Acta 2016, 1861, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Li, X.B.; Benning, C.; Kuo, M.H. Rapid triacylglycerol turnover in Chlamydomonas reinhardtii requires a lipase with broad substrate specificity. Eukaryot. Cell 2012, 11, 1451–1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, F.T.; Liang, Y.X.; Legeret, B.; Beyly-Adriano, A.; Blangy, S.; Haslam, R.P.; Napier, J.A.; Beisson, F.; Peltier, G.; Li-Beisson, Y. Chlamydomonas carries out fatty acid beta-oxidation in ancestral peroxisomes using a bona fide acyl-CoA oxidase. Plant J. 2017, 90, 358–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moellering, E.R.; Benning, C. RNA interference silencing of a major lipid droplet protein affects lipid droplet size in Chlamydomonas reinhardtii. Eukaryot. Cell 2010, 9, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Yoon, K.; Han, D.X.; Li, Y.T.; Sommerfeld, M.; Hu, Q. Phospholipid: Diacylglycerol acyltransferase is a multifunctional enzyme involved in membrane lipid turnover and degradation while synthesizing triacylglycerol in the unicellular green microalga Chlamydomonas reinhardtii. Plant Cell 2012, 24, 3708–3724. [Google Scholar] [CrossRef] [Green Version]
- De Jaeger, L.; Springer, J.; Wolbert, E.J.H.; Martens, D.E.; Eggink, G.; Wijffels, R.H. Gene silencing of stearoyl-ACP desaturase enhances the stearic acid content in Chlamydomonas reinhardtii. Bioresour. Technol. 2017, 245, 1616–1626. [Google Scholar] [CrossRef] [PubMed]
- Lau, C.C.; Loh, S.H.; Aziz, A.; Cha, T.S. Effects of disrupted omega-3 desaturase gene construct on fatty acid composition and expression of four fatty acid biosynthetic genes in transgenic Chlorella vulgaris. Algal Res. Biomass Biofuels Bioprod. 2017, 26, 143–152. [Google Scholar] [CrossRef]
- Hao, X.H.; Luo, L.; Jouhet, J.; Rebeille, F.; Marechal, E.; Hu, H.H.; Pan, Y.F.; Tan, X.M.; Chen, Z.; You, L.J.; et al. Enhanced triacylglycerol production in the diatom Phaeodactylum tricornutum by inactivation of a Hotdog-fold thioesterase gene using TALEN-based targeted mutagenesis. Biotechnol. Biofuels 2018, 11, 312. [Google Scholar] [CrossRef]
- Shtaida, N.; Khozin-Goldberg, I.; Solovchenko, A.; Chekanov, K.; Didi-Cohen, S.; Leu, S.; Cohen, Z.; Boussiba, S. Downregulation of a putative plastid PDC E1 alpha subunit impairs photosynthetic activity and triacylglycerol accumulation in nitrogen-starved photoautotrophic Chlamydomonas reinhardtii. J. Exp. Bot. 2014, 65, 6563–6576. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Han, D.X.; Yoon, K.; Hu, Q.; Li, Y.T. Characterization of type 2 diacylglycerol acyltransferases in Chlamydomonas reinhardtii reveals their distinct substrate specificities and functions in triacylglycerol biosynthesis. Plant J. 2016, 86, 3–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, J.K.; Smith, S.R.; McCrow, J.P.; Tan, M.; Zheng, H.; Beeri, K.; Roth, R.; Lichtle, C.; Goodenough, U.; Bowler, C.P.; et al. Nitrate reductase knockout uncouples nitrate transport from nitrate assimilation and drives repartitioning of carbon flux in a model pennate diatom. Plant Cell 2017, 29, 2047–2070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levitan, O.; Dinamarca, J.; Zelzion, E.; Gorbunov, M.Y.; Falkowski, P.G. An RNA interference knock-down of nitrate reductase enhances lipid biosynthesis in the diatom Phaeodactylum tricornutum. Plant J. 2015, 84, 963–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ajjawi, I.; Verruto, J.; Aqui, M.; Soriaga, L.B.; Coppersmith, J.; Kwok, K.; Peach, L.; Orchard, E.; Kalb, R.; Xu, W.D.; et al. Lipid production in Nannochloropsis gaditana is doubled by decreasing expression of a single transcriptional regulator. Nat. Biotechnol. 2017, 35, 647–652. [Google Scholar] [CrossRef]
- Meireles, L.A.; Guedes, A.C.; Malcata, F.X. Increase of the yields of eicosapentaenoic and docosahexaenoic acids by the microalga Pavlova lutheri following random mutagenesis. Biotechnol. Bioeng. 2003, 81, 50–55. [Google Scholar] [CrossRef]
- Osorio, H.; Jara, C.; Fuenzalida, K.; Rey-Jurado, E.; Vasquez, M. High-efficiency nuclear transformation of the microalgae Nannochloropsis oceanica using Tn5 Transposome for the generation of altered lipid accumulation phenotypes. Biotechnol. Biofuels 2019, 12, 134. [Google Scholar] [CrossRef] [PubMed]
- Iskandarov, U.; Sitnik, S.; Shtaida, N.; Didi-Cohen, S.; Leu, S.; Khozin-Goldberg, I.; Cohen, Z.; Boussiba, S. Cloning and characterization of a GPAT-like gene from the microalga Lobosphaera incisa (Trebouxiophyceae): Overexpression in Chlamydomonas reinhardtii enhances TAG production. J. Appl. Phycol. 2016, 28, 907–919. [Google Scholar] [CrossRef]
- Hamilton, M.L.; Haslam, R.P.; Napier, J.A.; Sayanova, O. Metabolic engineering of Phaeodactylum tricornutum for the enhanced accumulation of omega-3 long chain polyunsaturated fatty acids. Metab. Eng. 2014, 22, 3–9. [Google Scholar] [CrossRef]
- Kobayashi, T.; Sakaguchi, K.; Matsuda, T.; Abe, E.; Hama, Y.; Hayashi, M.; Honda, D.; Okita, Y.; Sugimoto, S.; Okino, N.; et al. Increase of eicosapentaenoic acid in Thraustochytrids through thraustochytrid ubiquitin promoter-driven expression of a fatty acid delta 5 desaturase gene. Appl. Environ. Microbiol. 2011, 77, 3870–3876. [Google Scholar] [CrossRef] [Green Version]
- Talebi, A.F.; Tohidfar, M.; Derazmahalleh, S.M.M.; Sulaiman, A.; Baharuddin, A.S.; Tabatabaei, M. Biochemical modulation of lipid pathway in microalgae Dunaliella sp. for biodiesel production. BioMed Res. Int. 2015, 2015, 597198. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.W.M.; Lee, Y.K. Expression of the heterologous Dunaliella tertiolecta fatty acyl-ACP thioesterase leads to increased lipid production in Chlamydomonas reinhardtii. J. Biotechnol. 2017, 247, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Kao, A.L.; Tsai, Z.C.; Chow, T.J.; Chang, H.Y.; Zhao, X.Q.; Chen, P.T.; Su, H.Y.; Chang, J.S. Expression of type 2 diacylglycerol acyltransferse gene DGTT1 from Chlamydomonas reinhardtii enhances lipid production in Scenedesmus obliquus. Biotechnol. J. 2016, 11, 336–344. [Google Scholar] [CrossRef]
- Ahmad, I.; Sharma, A.K.; Daniell, H.; Kumar, S. Altered lipid composition and enhanced lipid production in green microalga by introduction of brassica diacylglycerol acyltransferase 2. Plant Biotechnol. J. 2015, 13, 540–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zulu, N.N.; Popko, J.; Zienkiewicz, K.; Tarazona, P.; Herrfurth, C.; Feussner, I. Heterologous co-expression of a yeast diacylglycerol acyltransferase (ScDGA1) and a plant oleosin (AtOLEO3) as an efficient tool for enhancing triacylglycerol accumulation in the marine diatom Phaeodactylum tricornutum. Biotechnol. Biofuels 2017, 10, 187. [Google Scholar] [CrossRef]
- Hsieh, H.J.; Su, C.H.; Chien, L.J. Accumulation of lipid production in Chlorella minutissima by triacylglycerol biosynthesis-related genes cloned from Saccharomyces cerevisiae and Yarrowia lipolytica. J. Microbiol. 2012, 50, 526–534. [Google Scholar] [CrossRef]
- Ren, L.J.; Zhuang, X.Y.; Chen, S.L.; Ji, X.J.; Huang, H. Introduction of omega-3 desaturase obviously changed the fatty acid profile and sterol content of Schizochytrium sp. J. Agric. Food Chem. 2015, 63, 9770–9776. [Google Scholar] [CrossRef] [PubMed]
- Sumiya, N.; Kawase, Y.; Hayakawa, J.; Matsuda, M.; Nakamura, M.; Era, A.; Tanaka, K.; Kondo, A.; Hasunuma, T.; Imamura, S.; et al. Expression of cyanobacterial Acyl-ACP reductase elevates the triacylglycerol level in the red alga Cyanidioschyzon merolae. Plant Cell Physiol. 2015, 56, 1962–1980. [Google Scholar] [CrossRef] [Green Version]
- Radakovits, R.; Eduafo, P.M.; Posewitz, M.C. Genetic engineering of fatty acid chain length in Phaeodactylum tricornutum. Metab. Eng. 2011, 13, 89–95. [Google Scholar] [CrossRef]
- Lin, H.X.; Lee, Y.K. Genetic engineering of medium-chain-length fatty acid synthesis in Dunaliella tertiolecta for improved biodiesel production. J. Appl. Phycol. 2017, 29, 2811–2819. [Google Scholar] [CrossRef] [Green Version]
- Inaba, Y.; Nakahigashi, K.; Ito, T.; Tomita, M. Alteration of fatty acid chain length of Chlamydomonas reinhardtii by simultaneous expression of medium-chain-specific thioesterase and acyl carrier protein. Phycol. Res. 2017, 65, 94–99. [Google Scholar] [CrossRef]
- Gomma, A.E.; Lee, S.K.; Sun, S.M.; Yang, S.H.; Chung, G. Improvement in oil production by increasing malonyl-CoA and glycerol-3-phosphate pools in Scenedesmus quadricauda. Indian J. Microbiol. 2015, 55, 447–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.F.; Cheng, R.B.; Lin, X.Z.; You, S.; Li, K.; Rong, H.; Ma, Y. Overexpression of acetyl-CoA synthetase increased the biomass and fatty acid proportion in microalga Schizochytrium. Appl. Microbiol. Biotechnol. 2013, 97, 1933–1939. [Google Scholar] [CrossRef]
- Pudney, A.; Gandini, C.; Economou, C.K.; Smith, R.; Goddard, P.; Napier, J.A.; Spicer, A.; Sayanova, O. Multifunctionalizing the marine diatom Phaeodactylum tricornutum for sustainable co-production of omega-3 long chain polyunsaturated fatty acids and recombinant phytase. Sci. Rep. 2019, 9, 11444. [Google Scholar] [CrossRef]
- Zhang, J.H.; Hao, Q.; Bai, L.L.; Xu, J.; Yin, W.B.; Song, L.Y.; Xu, L.; Guo, X.J.; Fan, C.M.; Chen, Y.H.; et al. Overexpression of the soybean transcription factor GmDof4 significantly enhances the lipid content of Chlorella ellipsoidea. Biotechnol. Biofuels 2014, 7, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, N.K.; Kim, E.K.; Kim, Y.U.; Lee, B.; Jeong, W.J.; Jeong, B.R.; Chang, Y.K. Increased lipid production by heterologous expression of AtWRI1 transcription factor in Nannochloropsis salina. Biotechnol. Biofuels 2017, 10, 231. [Google Scholar] [CrossRef]
- Hou, L.; Jia, X.; Shi, D.; Wang, X. Expression of PEPC gene, lipid content and photosynthesis in Anabaena 7120. In Photosynthesis Research for Food, Fuel and the Future; Advanced topics in science and technology in China; Springer: Berlin/Heidelberg, Germany, 2008; pp. 335–338. [Google Scholar]
- Xie, W.H.; Pang, F.; Niu, Y.F.; Zhang, M.H.; Yang, W.D.; Liu, J.S.; Yan, D.G.; Li, H.Y. Functional characterization of an ACCase subunit from the diatom Phaeodactylum tricornutum expressed in Escherichia coli. Biotechnol. Appl. Biochem. 2013, 60, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Qi, B.X.; Fraser, T.; Mugford, S.; Dobson, G.; Sayanova, O.; Butler, J.; Napier, J.A.; Stobart, A.K.; Lazarus, C.M. Production of very long chain polyunsaturated omega-3 and omega-6 fatty acids in plants. Nat. Biotechnol. 2004, 22, 739–745. [Google Scholar] [CrossRef]
- Xia, F.; Li, X.Y.; Li, X.Z.; Zheng, D.S.; Sun, Q.X.; Liu, J.; Li, Y.X.; Hua, J.P.; Qi, B.X. Elevation of the yields of very long chain polyunsaturated fatty acids via minimal codon optimization of two key biosynthetic enzymes. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [Green Version]
- Petrie, J.R.; Shrestha, P.; Mansour, M.P.; Nichols, P.D.; Liu, Q.; Singh, S.P. Metabolic engineering of omega-3 long-chain polyunsaturated fatty acids in plants using an acyl-CoA Delta 6-desaturase with omega 3-preference from the marine microalga Micromonas pusilla. Metab. Eng. 2010, 12, 233–240. [Google Scholar] [CrossRef]
- Abbadi, A.; Domergue, F.; Bauer, J.; Napier, J.A.; Welti, R.; Zahringer, U.; Cirpus, P.; Heinz, E. Biosynthesis of very-long-chain polyunsaturated fatty acids in transgenic oilseeds: Constraints on their accumulation. Plant Cell 2004, 16, 2734–2748. [Google Scholar] [CrossRef] [Green Version]
- Walsh, T.A.; Bevan, S.A.; Gachotte, D.J.; Larsen, C.M.; Moskal, W.A.; Merlo, P.A.O.; Sidorenko, L.V.; Hampton, R.E.; Stoltz, V.; Pareddy, D.; et al. Canola engineered with a microalgal polyketide synthase-like system produces oil enriched in docosahexaenoic acid. Nat. Biotechnol. 2016, 34, 881–887. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.L.; Guo, B.; Wan, X.; Gong, Y.M.; Zhang, Y.B.; Hu, C.J. Isolation and characterization of the diatom Phaeodactylum delta 5-elongase gene for transgenic LC-PUFA production in Pichia pastoris. Mar. Drugs 2014, 12, 1317–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.D.; Chi, X.Y.; Yang, Q.L.; Li, Z.X.; Liu, S.F.; Gan, Q.H.; Qin, S. Molecular cloning and stress-dependent expression of a gene encoding Delta(12)-fatty acid desaturase in the Antarctic microalga Chlorella vulgaris NJ-7. Extremophiles 2009, 13, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.H.; Ho, M.Y.; Kanehara, K.; Nakamura, Y. Functional study of diacylglycerol acyltransferase type 2 family in Chlamydomonas reinhardtii. FEBS Lett. 2013, 587, 2364–2370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domergue, F.; Lerchl, J.; Zahringer, U.; Heinz, E. Cloning and functional characterization of Phaeodactylum tricornutum front-end desaturases involved in eicosapentaenoic acid biosynthesis. Eur. J. Biochem. 2002, 269, 4105–4113. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.S.; Chen, H.Q.; Gu, Z.N.; Zhang, H.; Chen, W.; Chen, Y.Q. Application of a delta-6 desaturase with alpha-linolenic acid preference on eicosapentaenoic acid production in Mortierella alpina. Microb. Cell Factories 2016, 15, 117. [Google Scholar] [CrossRef] [Green Version]
- Thiyagarajan, S.; Arumugam, M.; Senthil, N.; Vellaikumar, S.; Kathiresan, S. Functional characterization and substrate specificity analysis of Delta 6-desaturase from marine microalga Isochrysis sp. Biotechnol. Lett. 2018, 40, 577–584. [Google Scholar] [CrossRef]
- Thiyagarajan, S.; Arumugam, M.; Kathiresan, S. Identification and functional characterization of two novel fatty acid genes from marine microalgae for eicosapentaenoic acid production. Appl. Biochem. Biotechnol. 2020, 190, 1371–1384. [Google Scholar] [CrossRef]
- Li-Beisson, Y.; Thelen, J.J.; Fedosejevs, E.; Harwood, J.L. The lipid biochemistry of eukaryotic algae. Prog. Lipid Res. 2019, 74, 31–68. [Google Scholar] [CrossRef]
- Davis, D.; Rudenko, G.; Somanchi, A.; Casolari, J.; Franklin, S.; Ewing, A. Novel Ketoacyl ACP Synthase Genes and Uses Thereof. U.S. Patent 2019/0345463 A1, 14 January 2016. [Google Scholar]
- Ozaki, T. Lipid Production Method Using Acyl-ACP Thioesterase. JP Patent 2016007154 A, 18 January 2016. [Google Scholar]
- Moreno-Sanchez, R.; Saavedra, E.; Rodriguez-Enriquez, S.; Olin-Sandoval, V. Metabolic control analysis: A tool for designing strategies to manipulate metabolic pathways. J. Biomed. Biotechnol. 2008, 2008, 597913. [Google Scholar] [CrossRef] [Green Version]
- Fell, D.A. Understanding the Control of Metabolism; Portland Press: London, UK, 2007; 306p. [Google Scholar]
- Sayanova, O.; Mimouni, V.; Ulmann, L.; Morant-Manceau, A.; Pasquet, V.; Schoefs, B.; Napier, J.A. Modulation of lipid biosynthesis by stress in diatoms. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krogdahl, A.; Penn, M.; Thorsen, J.; Refstie, S.; Bakke, A.M. Important antinutrients in plant feedstuffs for aquaculture: An update on recent findings regarding responses in salmonids. Aquac. Res. 2010, 41, 333–344. [Google Scholar] [CrossRef]
- Fan, J.L.; Andre, C.; Xu, C.C. A chloroplast pathway for the de novo biosynthesis of triacylglycerol in Chlamydomonas reinhardtii. FEBS Lett. 2011, 585, 1985–1991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.U.; Huang, A.H.C. Plastid lysophosphatidyl acyltransferase is essential for embryo development in Arabidopsis. Plant Physiol. 2004, 134, 1206–1216. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.U.; Lee, K.R.; Jung, S.J.; Shin, H.A.; Go, Y.S.; Suh, M.C.; Kim, J.B. Senescence-inducible LEC2 enhances triacylglycerol accumulation in leaves without negatively affecting plant growth. Plant Biotechnol. J. 2015, 13, 1346–1359. [Google Scholar] [CrossRef] [Green Version]
- Jiang, P.; Yang, G.; Jin, Z.; Huaxin, C. Method for Increasing Lipid Content of Diatom Through Metabolism Gene Engineering. CN Patent 105219649 A, 6 January 2016. [Google Scholar]
- Yi, X.; Xy, J. Gene with Triglyceride (TAG) Synthesis Function and Construction Method and Application Thereof. CN Patent 110305883 A, 8 October 2019. [Google Scholar]
- Chen, W.; Zhou, P.P.; Zhu, Y.M.; Xie, C.; Ma, L.; Wang, X.P.; Bao, Z.D.; Yu, L.J. Improvement in the docosahexaenoic acid production of Schizochytrium sp. S056 by replacement of sea salt. Bioprocess Biosyst. Eng. 2016, 39, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Fan, C.; Chen, Y. Chlorella ellipsoidea NF-YC Gene and Application Thereof. CN Patent 108101973 A, 1 July 2018. [Google Scholar]
- Li, Y.T.; Han, D.X.; Hu, G.R.; Sommerfeld, M.; Hu, Q.A. Inhibition of starch synthesis results in overproduction of lipids in Chlamydomonas reinhardtii. Biotechnol. Bioeng. 2010, 107, 258–268. [Google Scholar] [CrossRef]
- Deng, W.; Cai, J.X.; Zhang, J.Y.; Chen, Y.Y.; Chen, Y.R.; Di, Y.J.; Yuan, S.Z. Molecular basis of resistance to ACCase-inhibiting herbicide cyhalofop-butyl in Chinese sprangletop (Leptochloa chinensis (L.) Nees) from China. Pestic. Biochem. Physiol. 2019, 158, 143–148. [Google Scholar] [CrossRef]
- Cygler, M.; Schrag, J.D. Structure as basis for understanding interfacial properties of lipases. Methods Enzymol. 1997, 284, 3–27. [Google Scholar]
- Sayanova, O.; Napier, J.A. Metabolic engineering of microalgae for sustainable production of omega-3 long chain polyunsaturated fatty acids. Curr. Biotechnol. 2016, 5, 198–212. [Google Scholar] [CrossRef]
- Yang, Y.X.; Ahammed, G.J.; Wan, C.P.; Liu, H.J.; Chen, R.R.; Zhou, Y. Comprehensive analysis of TIFY transcription factors and their expression profiles under jasmonic acid and abiotic stresses in watermelon. Int. J. Genom. 2019, 2019, 6813086. [Google Scholar] [CrossRef]
- Liu, A.F.; Yu, X.Y.; Liu, S.R. Pluripotency transcription factors and cancer stem cells: Small genes make a big difference. Chin. J. Cancer 2013, 32, 483–487. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.C.; Liang, M.H.; Jiang, J.G. Transgenic microalgae as bioreactors. Crit. Rev. Food Sci. Nutr. 2020, 60, 3195–3213. [Google Scholar] [CrossRef]
- Bajhaiya, A.K.; Moreira, J.Z.; Pittman, J.K. Transcriptional engineering of microalgae: Prospects for high-value chemicals. Trends Biotechnol. 2017, 35, 95–99. [Google Scholar] [CrossRef]
- Hu, Z.; Fan, C.; Sun, P.; Chen, Y. Chlorella ellipsoidea NF-YA Gene and Application Thereof. CN Patent 107936098 A, 20 April 2018. [Google Scholar]
- Hu, Z.; Fan, C.; Chen, Y. Chlorella ellipsoidea NF-YB Gene and Application Thereof. CN Patent 108003226 A, 8 May 2018. [Google Scholar]
- Yang, J.W.; Kang, N.K.; Jeong, B.R.; Park, M.S.; Lee, B.S.; Choi, G.G.; Jeon, S.J.; Kwon, S.H. Recombinant Vector for Increasing Biomass and Lipid Productivity of Microalgae and Use Thereof. WO Patent 2015/105233 A1, 16 July 2015. [Google Scholar]
- Li, J.W.; Zhang, X.Y.; Wu, H.; Bai, Y.P. Transcription factor engineering for high-throughput strain evolution and organic acid bioproduction: A review. Front. Bioeng. Biotechnol. 2020, 8, 98. [Google Scholar] [CrossRef] [PubMed]
- Tibbetts, S.M.; Scaife, M.A.; Armenta, R.E. Apparent digestibility of proximate nutrients, energy and fatty acids in nutritionally-balanced diets with partial or complete replacement of dietary fish oil with microbial oil from a novel Schizochytrium sp. (T18) by juvenile Atlantic salmon (Salmo salar L.). Aquaculture 2020, 520, 735003. [Google Scholar] [CrossRef]
- Cornelio, F.H.G.; da Cunha, D.A.; Silveira, J.; Alexandre, D.; Silva, C.P.; Fracalossi, D.M. Dietary protein requirement of juvenile cachara catfish, Pseudoplatystoma reticulatum. J. World Aquac. Soc. 2014, 45, 45–54. [Google Scholar] [CrossRef]
- Sarker, P.K.; Kapuscinski, A.R.; McKuin, B.; Fitzgerald, D.S.; Nash, H.M.; Greenwood, C. Microalgae-blend tilapia feed eliminates fishmeal and fish oil, improves growth, and is cost viable. Sci. Rep. 2020, 10, 19328. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-Othon, C.A.; Perez-Velazquez, M.; Gatlin, D.M.; Gonzalez-Felix, M.L. Replacement of of fish oil by soybean oil and microalgal meals in diets for Totoaba macdonaldi (Gilbert, 1890) juveniles. Aquaculture 2020, 529, 735705. [Google Scholar] [CrossRef]
- Olsen, R.L.; Hasan, M.R. A limited supply of fishmeal: Impact on future increases in global aquaculture production. Trends Food Sci. Technol. 2012, 27, 120–128. [Google Scholar] [CrossRef]
- Beal, C.M.; Gerber, L.N.; Thongrod, S.; Phromkunthong, W.; Kiron, V.; Granados, J.; Archibald, I.; Greene, C.H.; Huntley, M.E. Marine microalgae commercial production improves sustainability of global fisheries and aquaculture. Sci. Rep. 2018, 8, 15064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sørensen, M.; Berge, G.M.; Reitan, K.I.; Ruyter, B. Microalga Phaeodactylum tricornutum in feed for Atlantic salmon (Salmo salar)—Effect on nutrient digestibility, growth and utilization of feed. Aquaculture 2016, 460, 116–123. [Google Scholar] [CrossRef]
- Gonzalez-Fernandez, C.; Munoz, R. (Eds.) Microalgae-Based Biofuels and Bioproducts: From Feedstock Cultivation to End-Products; Elsevier: Amsterdam, The Netherlands, 2017; 540p. [Google Scholar]
- De la Fuente-Vazquez, J.; Diaz-Diaz-Chiron, M.T.; Perez-Marcos, C.; Caneque-Martinez, V.; Sanchez-Gonzalez, C.I.; Alvarez-Acero, I.; Fernandez-Bermejo, C.; Rivas-Canedo, A.; Lauzurica-Gomez, S. Linseed, microalgae or fish oil dietary supplementation affects performance and quality characteristics of light lambs. Span. J. Agric. Res. 2014, 12, 436–447. [Google Scholar] [CrossRef]
- Barron, J.M.; Twibell, R.G.; Hill, H.A.; Hanson, K.C.; Gannam, A.L. Development of diets for the intensive culture of Pacific lamprey. Aquac. Res. 2016, 47, 3899–3906. [Google Scholar] [CrossRef]
- Miller, M.R.; Nichols, P.D.; Carter, C.G. Replacement of fish oil with thraustochytrid Schizochytrium sp. L. oil in Atlantic salmon parr (Salmo salar L.) diets. Comp. Biochem. Physiol. A—Mol. Integr. Physiol. 2007, 148, 382–392. [Google Scholar] [CrossRef] [PubMed]
- Sarker, P.K.; Kapuscinski, A.R.; Lanois, A.J.; Livesey, E.D.; Bernhard, K.P.; Coley, M.L. Towards sustainable aquafeeds: Complete substitution of fish oil with marine microalga Schizochytrium sp. improves growth and fatty acid deposition in juvenile Nile tilapia (Oreochromis niloticus). PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [Green Version]
- Cortegano, C.A.A.; de Alcantara, A.M.; da Silva, A.F.; Epifanio, C.M.F.; Bentes, S.P.C.; dos Santos, V.J.; Visentainer, J.V.; Goncalves, L.U. Finishing plant diet supplemented with microalgae meal increases the docosahexaenoic acid content in Colossoma macropomum flesh. Aquac. Res. 2019, 50, 1291–1299. [Google Scholar] [CrossRef]
- Belanger, A.; Sarker, P.K.; Bureau, D.P.; Chouinard, Y.; Vandenberg, G.W. Apparent digestibility of macronutrients and fatty acids from microalgae (Schizochytrium sp.) fed to rainbow trout (Oncorhynchus mykiss): A potential candidate for fish oil substitution. Animals 2021, 11, 456. [Google Scholar] [CrossRef]
- Seong, T.; Matsutani, H.; Haga, Y.; Kitagima, R.; Satoh, S. First step of non-fish meal, non-fish oil diet development for red seabream, (Pagrus major), with plant protein sources and microalgae Schizochytrium sp. Aquac. Res. 2019, 50, 2460–2468. [Google Scholar] [CrossRef]
- Yan, L.; Kim, I.H. Effects of dietary omega-3 fatty acid-enriched microalgae supplementation on growth performance, blood profiles, meat quality, and fatty acid composition of meat in broilers. J. Appl. Anim. Res. 2013, 41, 392–397. [Google Scholar] [CrossRef] [Green Version]
- Li, M.H.H.; Robinson, E.H.; Tucker, C.S.; Manning, B.B.; Khoo, L. Effects of dried algae Schizochytrium sp., a rich source of docosahexaenoic acid, on growth, fatty acid composition, and sensory quality of channel catfish Ictalurus punctatus. Aquaculture 2009, 292, 232–236. [Google Scholar] [CrossRef]
- Kumar, V.; Habte-Tsion, H.M.; Allen, K.M.; Bowman, B.A.; Thompson, K.R.; El-Haroun, E.; Filer, K.; Tidwell, J.H. Replacement of fish oil with Schizochytrium meal and its impacts on the growth and lipid metabolism of Pacific white shrimp (Litopenaeus vannamei). Aquac. Nutr. 2018, 24, 1769–1781. [Google Scholar] [CrossRef]
- Urrutia, O.; Mendizabal, J.A.; Insausti, K.; Soret, B.; Purroy, A.; Arana, A. Effects of addition of linseed and marine algae to the diet on adipose tissue development, fatty acid profile, lipogenic gene expression, and meat quality in lambs. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [Green Version]
- Mordenti, A.L.; Sardi, L.; Bonaldo, A.; Pizzamiglio, V.; Brogna, N.; Cipollini, I.; Tassinari, M.; Zaghini, G. Influence of marine algae (Schizochytrium spp.) dietary supplementation on doe performance and progeny meat quality. Livest. Sci. 2010, 128, 179–184. [Google Scholar] [CrossRef]
- Kousoulaki, K.; Morkore, T.; Nengas, I.; Berge, R.K.; Sweetman, J. Microalgae and organic minerals enhance lipid retention efficiency and fillet quality in Atlantic salmon (Salmo salar L.). Aquaculture 2016, 451, 47–57. [Google Scholar] [CrossRef]
- Garcia-Ortega, A.; Kissinger, K.R.; Trushenski, J.T. Evaluation of fish meal and fish oil replacement by soybean protein and algal meal from Schizochytrium limacinum in diets for giant grouper Epinephelus lanceolatus. Aquaculture 2016, 452, 1–8. [Google Scholar] [CrossRef]
- Bouras, S.; Katsoulas, N.; Antoniadis, D.; Karapanagiotidis, I.T. Use of biofuel industry wastes as alternative nutrient sources for DHA-Yielding Schizochytrium limacinum production. Appl. Sci. 2020, 10, 4398. [Google Scholar] [CrossRef]
- Jaseera, K.V.; Ebeneezar, S.; Sayooj, P.; Nair, A.V.; Kaladharan, P. Dietary supplementation of microalgae, Aurantiochytrium sp. and co-feeding with Artemia enhances the growth, stress tolerance and survival in Penaeus monodon (Fabricius, 1798) post larvae. Aquaculture 2021, 533, 736176. [Google Scholar] [CrossRef]
- Tibaldi, E.; Zittelli, G.C.; Parisi, G.; Bruno, M.; Giorgi, G.; Tulli, F.; Venturini, S.; Tredici, M.R.; Poli, B.M. Growth performance and quality traits of European sea bass (D. labrax) fed diets including increasing levels of freeze-dried Isochrysis sp. (T-ISO) biomass as a source of protein and n-3 long chain PUFA in partial substitution of fish derivatives. Aquaculture 2015, 440, 60–68. [Google Scholar] [CrossRef]
- He, Y.J.; Lin, G.; Rao, X.Z.; Chen, L.J.; Jian, H.; Wang, M.Z.; Guo, Z.; Chen, B.L. Microalga Isochrysis galbana in feed for Trachinotus ovatus: Effect on growth performance and fatty acid composition of fish fillet and liver. Aquac. Int. 2018, 26, 1261–1280. [Google Scholar] [CrossRef]
- Atalah, E.; Cruz, C.M.H.; Izquierdo, M.S.; Rosenlund, G.; Caballero, M.J.; Valencia, A.; Robaina, L. Two microalgae Crypthecodinium cohnii and Phaeodactylum tricornutum as alternative source of essential fatty acids in starter feeds for seabream (Sparus aurata). Aquaculture 2007, 270, 178–185. [Google Scholar] [CrossRef]
- Sheikhzadeh, N.; Tayefi-Nasrabadi, H.; Oushani, A.K.; Enferadi, M.H.N. Effects of Haematococcus pluvialis supplementation on antioxidant system and metabolism in rainbow trout (Oncorhynchus mykiss). Fish Physiol. Biochem. 2012, 38, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Austic, R.E.; Mustafa, A.; Jung, B.; Gatrell, S.; Lei, X.G. Potential and limitation of a new defatted diatom microalgal biomass in replacing soybean meal and corn in diets for broiler chickens. J. Agric. Food Chem. 2013, 61, 7341–7348. [Google Scholar] [CrossRef]
- Velasquez, S.F.; Chan, M.A.; Abisado, R.G.; Traifalgar, R.F.M.; Tayamen, M.M.; Maliwat, G.C.F.; Ragaza, J.A. Dietary Spirulina (Arthrospira platensis) replacement enhances performance of juvenile Nile tilapia (Oreochromis niloticus). J. Appl. Phycol. 2016, 28, 1023–1030. [Google Scholar] [CrossRef]
- Bonos, E.; Kasapidou, E.; Kargopoulos, A.; Karampampas, A.; Christaki, E.; Florou-Paneri, P.; Nikolakakis, I. Spirulina as a functional ingredient in broiler chicken diets. S. Afr. J. Anim. Sci. 2016, 46, 94–102. [Google Scholar] [CrossRef] [Green Version]
- Raji, A.A.; Alaba, P.A.; Yusuf, H.; Abu Bakar, N.H.; Taufek, N.M.; Muin, H.; Alias, Z.; Milow, P.; Razak, S.A. Fishmeal replacement with Spirulina Platensis and Chlorella vulgaris in African catfish (Clarias gariepinus) diet: Effect on antioxidant enzyme activities and haematological parameters. Res. Vet. Sci. 2018, 119, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Luo, Z.; Chen, F.; Wei, C.C.; Wu, K.; Zhu, X.M.; Liu, X. Effect of fish meal replacement by Chlorella meal with dietary cellulase addition on growth performance, digestive enzymatic activities, histology and myogenic genes’ expression for crucian carp Carassius auratus. Aquac. Res. 2017, 48, 3244–3256. [Google Scholar] [CrossRef]
- Kupchinsky, Z.A.; Coyle, S.D.; Bright, L.A.; Tidwell, J.H. Evaluation of heterotrophic algae meal as a diet ingredient for channel catfish, Ictalurus punctatus. J. World Aquac. Soc. 2015, 46, 445–452. [Google Scholar] [CrossRef]
- Castro, C.; Coutinho, F.; Iglesias, P.; Oliva-Teles, A.; Couto, A. Chlorella sp. and Nannochloropsis sp. inclusion in plant-based diets modulate the intestine and liver antioxidant mechanisms of European sea bass juveniles. Front. Vet. Sci. 2020, 7, 607575. [Google Scholar] [CrossRef]
- Teuling, E.; Wierenga, P.A.; Agboola, J.O.; Gruppen, H.; Schrama, J.W. Cell wall disruption increases bioavailability of Nannochloropsis gaditana nutrients for juvenile Nile tilapia (Oreochromis niloticus). Aquaculture 2019, 499, 269–282. [Google Scholar] [CrossRef]
- Sales, R.; Galafat, A.; Vizcaino, A.J.; Saez, M.I.; Martinez, T.F.; Ceron-Garcia, M.C.; Navarro-Lopez, E.; Tsuzuki, M.Y.; Acien-Fernandez, F.G.; Molina-Grima, E.; et al. Effects of dietary use of two lipid extracts from the microalga Nannochloropsis gaditana (Lubian, 1982) alone and in combination on growth and muscle composition in juvenile gilthead seabream, Sparus aurata. Algal Res. Biomass Biofuels Bioprod. 2021, 53, 102162. [Google Scholar] [CrossRef]
- Oswald, A.T.O.; Ishikawa, M.; Koshio, S.; Yokoyama, S.; Moss, A.S.; Serge, D. Nutritional evaluation of Nannochloropsis powder and lipid as alternative to fish oil for kuruma shrimp, Marsupenaeus japonicus. Aquaculture 2019, 504, 427–436. [Google Scholar] [CrossRef]
- Valente, L.M.P.; Custodio, M.; Batista, S.; Fernandes, H.; Kiron, V. Defatted microalgae (Nannochloropsis sp.) from biorefinery as a potential feed protein source to replace fishmeal in European sea bass diets. Fish Physiol. Biochem. 2019, 45, 1067–1081. [Google Scholar] [CrossRef]
- Gbadamosi, O.K.; Lupatsch, I. Effects of dietary Nannochloropsis salina on the nutritional performance and fatty acid profile of Nile tilapia, Oreochromis niloticus. Algal Res. Biomass Biofuels Bioprod. 2018, 33, 48–54. [Google Scholar] [CrossRef]
- Haas, S.; Bauer, J.L.; Adakli, A.; Meyer, S.; Lippemeier, S.; Schwarz, K.; Schulz, C. Marine microalgae Pavlova viridis and Nannochloropsis sp. as n-3 PUFA source in diets for juvenile European sea bass (Dicentrarchus labrax L.). J. Appl. Phycol. 2016, 28, 1011–1021. [Google Scholar] [CrossRef]
- Ponis, E.; Parisi, G.; Le Coz, J.R.; Robert, R.; Zittelli, G.C.; Tredici, R. Effect of the culture system and culture technique on biochemical characteristics of Pavlova lutheri and its nutritional value for Crassostrea gigas larvae. Aquac. Nutr. 2006, 12, 322–329. [Google Scholar] [CrossRef] [Green Version]
- Sharawy, Z.Z.; Ashour, M.; Abbas, E.; Ashry, O.; Helal, M.; Nazmi, H.; Kelany, M.; Kamel, A.; Hassaan, M.; Rossi, W.; et al. Effects of dietary marine microalgae, Tetraselmis suecica, on production, gene expression, protein markers and bacterial count of Pacific white shrimp Litopenaeus vannamei. Aquac. Res. 2020, 51, 2216–2228. [Google Scholar] [CrossRef]
- Rahman, N.A.; Khatoon, H.; Yusuf, N.; Banerjee, S.; Haris, N.A.; Lananan, F.; Tomoyo, K. Tetraselmis chuii biomass as a potential feed additive to improve survival and oxidative stress status of Pacific white-leg shrimp Litopenaeus vannamei postlarvae. Int. Aquat. Res. 2017, 9, 235–247. [Google Scholar] [CrossRef] [Green Version]
- Pereira, H.; Sardinha, M.; Santos, T.; Gouveia, L.; Barreira, L.; Dias, J.; Varela, J. Incorporation of defatted microalgal biomass (Tetraselmis sp. CTP4) at the expense of soybean meal as a feed ingredient for juvenile gilthead seabream (Sparus aurata). Algal Res. Biomass Biofuels Bioprod. 2020, 47, 101869. [Google Scholar] [CrossRef]
- Messina, M.; Bulfon, C.; Beraldo, P.; Tibaldi, E.; Cardinaletti, G. Intestinal morpho-physiology and innate immune status of European sea bass (Dicentrarchus labrax) in response to diets including a blend of two marine microalgae, Tisochrysis lutea and Tetraselmis suecica. Aquaculture 2019, 500, 660–669. [Google Scholar] [CrossRef]
- Kiron, V.; Sørensen, M.; Huntley, M.; Vasanth, G.K.; Gong, Y.Y.; Dahle, D.; Palihawadana, A.M. Defatted biomass of the microalga, Desmodesmus sp., can replace fishmeal in the feeds for Atlantic salmon. Front. Mar. Sci. 2016, 3, 67. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, M.; Montero, D.; Torrecillas, S.; Castro, P.; Zamorano, M.J.; Izquierdo, M. Hepatic biochemical, morphological and molecular effects of feeding microalgae and poultry oils to gilthead sea bream (Sparus aurata). Aquaculture 2021, 532, 736073. [Google Scholar] [CrossRef]
- Sarker, P.K.; Kapuscinski, A.R.; Vandenberg, G.W.; Proulx, E.; Sitek, A.J. Towards sustainable and ocean-friendly aquafeeds: Evaluating a fish-free feed for rainbow trout (Oncorhynchus mykiss) using three marine microalgae species. Elem. Sci. Anthr. 2020, 8, 404. [Google Scholar] [CrossRef] [Green Version]
- Perez-Velazquez, M.; Gatlin, D.M.; Gonzalez-Felix, M.L.; Garcia-Ortega, A.; de Cruz, C.R.; Juarez-Gomez, M.L.; Chen, K.Q. Effect of fishmeal and fish oil replacement by algal meals on biological performance and fatty acid profile of hybrid striped bass (Morone crhysops female × M. saxatilis male). Aquaculture 2019, 507, 83–90. [Google Scholar] [CrossRef]
- Bruce, J.R.; Knight, M.; Parke, M.W. The rearing of oyster larvae on an algal diet. J. Mar. Biol. Assoc. UK 2009, 24, 337–374. [Google Scholar] [CrossRef] [Green Version]
- Ryckebosch, E.; Bruneel, C.; Termote-Verhalle, R.; Goiris, K.; Muylaert, K.; Foubert, I. Nutritional evaluation of microalgae oils rich in omega-3 long chain polyunsaturated fatty acids as an alternative for fish oil. Food Chem. 2014, 160, 393–400. [Google Scholar] [CrossRef] [Green Version]
- Jusadi, D.; Ekasari, J.; Suprayudi, M.A.; Setiawati, M.; Fauzi, I.A. Potential of underutilized marine organisms for aquaculture feeds. Front. Mar. Sci. 2021, 8, 609471. [Google Scholar] [CrossRef]
- Lindsay, G.J.H.; Harris, J.E. Carboxymethylcellulase activity in the digestive tracts of fish. J. Fish Biol. 1980, 16, 219–233. [Google Scholar] [CrossRef]
- Archibeque, S.L.; Ettinger, A.; Willson, B.D. Nannochloropsis oculata as a source of animal feed. Acta Agron. Hung. 2009, 57, 245–248. [Google Scholar] [CrossRef]
- Li, X.P.; Liu, J.P.; Chen, G.Y.; Zhang, J.G.; Wang, C.B.; Liu, B. Extraction and purification of eicosapentaenoic acid and docosahexaenoic acid from microalgae: A critical review. Algal Res. Biomass Biofuels Bioprod. 2019, 43, 101619. [Google Scholar] [CrossRef]
- Lee, S.Y.; Cho, J.M.; Chang, Y.K.; Oh, Y.K. Cell disruption and lipid extraction for microalgal biorefineries: A review. Bioresour. Technol. 2017, 244, 1317–1328. [Google Scholar] [CrossRef]
- Adam, F.; Abert-Vian, M.; Peltier, G.; Chemat, F. “Solvent-free” ultrasound-assisted extraction of lipids from fresh microalgae cells: A green, clean and scalable process. Bioresour. Technol. 2012, 114, 457–465. [Google Scholar] [CrossRef]
- Lee, J.Y.; Yoo, C.; Jun, S.Y.; Ahn, C.Y.; Oh, H.M. Comparison of several methods for effective lipid extraction from microalgae. Bioresour. Technol. 2010, 101, S75–S77. [Google Scholar] [CrossRef]
- Goettel, M.; Eing, C.; Gusbeth, C.; Straessner, R.; Frey, W. Pulsed electric field assisted extraction of intracellular valuables from microalgae. Algal Res. Biomass Biofuels Bioprod. 2013, 2, 401–408. [Google Scholar] [CrossRef]
- Yoo, G.; Park, W.K.; Kim, C.W.; Choi, Y.E.; Yang, J.W. Direct lipid extraction from wet Chlamydomonas reinhardtii biomass using osmotic shock. Bioresour. Technol. 2012, 123, 717–722. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Kong, X.Y.; Wang, Z.M.; Sun, Y.M.; Zhu, S.N.; Li, L.H.; Lv, P.M. Optimization of enzymatic hydrolysis for effective lipid extraction from microalgae Scenedesmus sp. Renew. Energy 2018, 125, 1049–1057. [Google Scholar] [CrossRef]
- Zuorro, A.; Maffei, G.; Lavecchia, R. Optimization of enzyme-assisted lipid extraction from Nannochloropsis microalgae. J. Taiwan Inst. Chem. Eng. 2016, 67, 106–114. [Google Scholar] [CrossRef]
- Ryckebosch, E.; Muylaert, K.; Foubert, I. Optimization of an analytical procedure for extraction of lipids from microalgae. J. Am. Oil Chem. Soc. 2012, 89, 189–198. [Google Scholar] [CrossRef]
- Sharma, A.K.; Chintala, V.; Ghodke, P.; Prasher, P.; Patel, A. Extraction and purification of PUFA from microbial biomass. In Nutraceutical Fatty Acids from Oleaginous Microalgae; John Wiley and Sons: Hoboken, NJ, USA, 2020; pp. 249–279. [Google Scholar]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Matyash, V.; Liebisch, G.; Kurzchalia, T.V.; Shevchenko, A.; Schwudke, D. Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. J. Lipid Res. 2008, 49, 1137–1146. [Google Scholar] [CrossRef] [Green Version]
- Patel, A.; Matsakas, L.; Sartaj, K.; Chandra, R. Extraction of lipids from algae using supercritical carbon dioxide. In Green Sustainable Process for Chemical and Environmental Engineering and Science; Elsevier: Amsterdam, The Netherlands, 2020; pp. 17–39. [Google Scholar]
- Daintree, L.S.; Kordikowski, A.; York, P. Separation processes for organic molecules using SCF Technologies. Adv. Drug Deliv. Rev. 2008, 60, 351–372. [Google Scholar] [CrossRef] [PubMed]
- Sahena, F.; Zaidul, I.S.M.; Jinap, S.; Karim, A.A.; Abbas, K.A.; Norulaini, N.A.N.; Omar, A.K.M. Application of supercritical CO2 in lipid extraction—A review. J. Food Eng. 2009, 95, 240–253. [Google Scholar] [CrossRef]
- Mendes, A.; da Silva, T.L.; Reis, A. DHA concentration and purification from the marine heterotrophic microalga Crypthecodinium cohnii CCMP 316 by winterization and urea complexation. Food Technol. Biotechnol. 2007, 45, 38–44. [Google Scholar]
- Ryckebosch, E.; Bruneel, C.; Termote-Verhalle, R.; Lemahieu, C.; Muylaert, K.; Van Durme, J.; Goiris, K.; Foubert, I. Stability of Omega-3 LC-PUFA-rich photoautotrophic microalgal oils compared to commercially available Omega-3 LC-PUFA oils. J. Agric. Food Chem. 2013, 61, 10145–10155. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, T.Z.; Zhang, W.; Chen, X.L.; Wang, J.F. Biodiesel production from algae oil high in free fatty acids by two-step catalytic conversion. Bioresour. Technol. 2012, 111, 208–214. [Google Scholar] [CrossRef]
- Gatrell, S.K.; Kim, J.; Derksen, T.J.; O’Neil, E.V.; Lei, X.G. Creating omega-3 fatty-acid-enriched chicken using defatted green microalgal biomass. J. Agric. Food Chem. 2015, 63, 9315–9322. [Google Scholar] [CrossRef]
- Camacho, F.; Macedo, A.; Malcata, F. Potential industrial applications and commercialization of microalgae in the functional food and feed industries: A short review. Mar. Drugs 2019, 17, 312. [Google Scholar] [CrossRef] [Green Version]



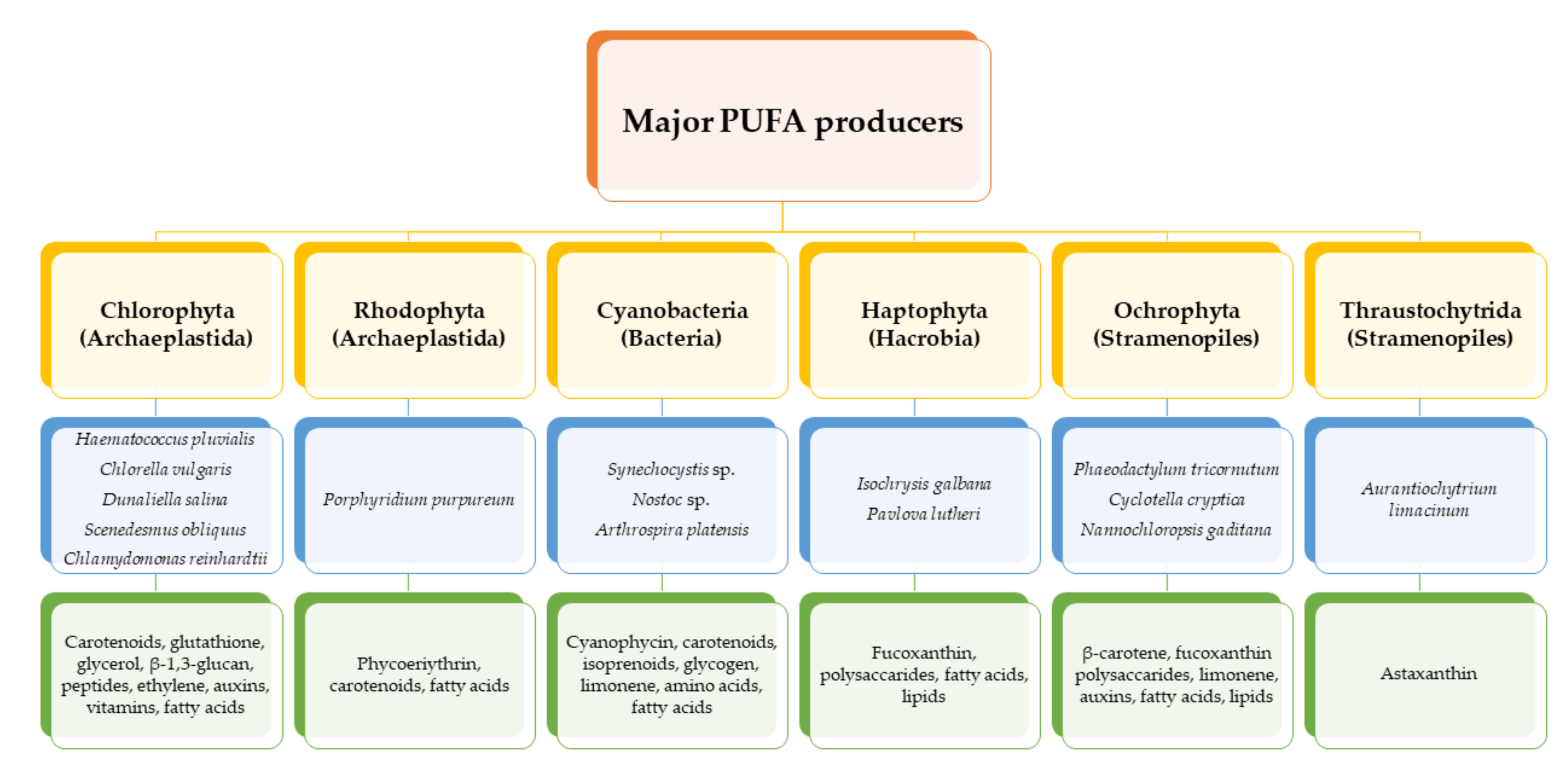{kind=link}
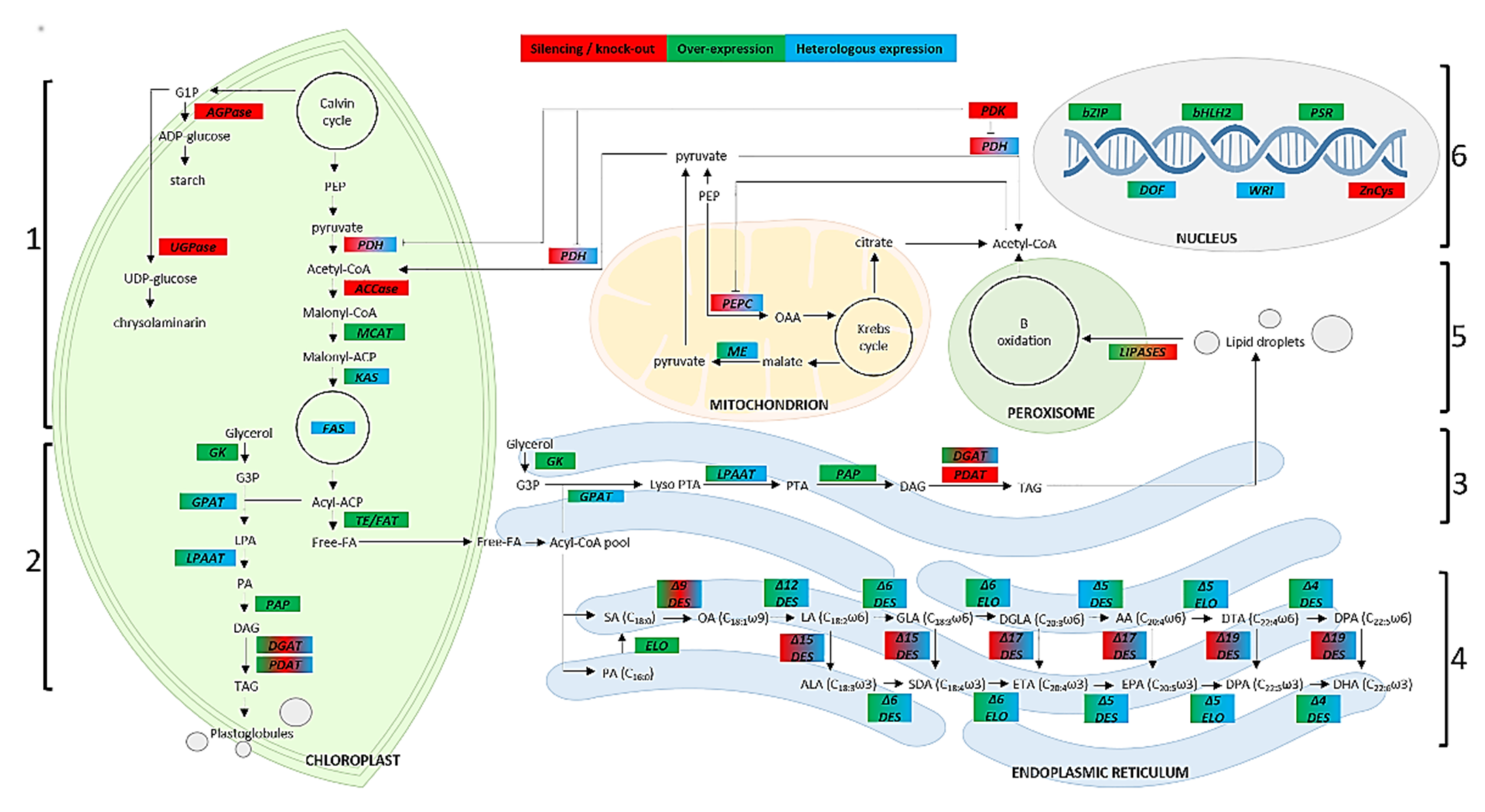{kind=link}
| Fatty Acid | Acronym | IUPAC Name | Structure |
|---|---|---|---|
| Palmitic acid (C16:0) | PA | Hexadecanoic acid |  |
| Stearic acid (C18:0) | SA | Octadecanoic acid |  |
| Oleic acid (C18:1ω9) | OA | (9Z)-Octadec-9-enoic acid |  |
| Linoleic acid (C18:2ω6) | LA | (9Z,12Z)-Octadeca-9,12-dienoic acid |  |
| α-linolenic acid (C18:3ω3) | ALA | (9Z,12Z,15Z)-Octadeca-9,12,15-trienoic acid |  |
| γ-linolenic acid (C18:3ω6) | GLA | (6Z,9Z,12Z)-Octadeca-6,9,12-trienoic acid |  |
| Eicosatetraenoic acid (C20:4ω3) | ETA | (8Z,11Z,14Z,17Z)-Icosa-8,11,14,17- tetraenoic acid |  |
| Dihomo-γ-linoleic acid (C20:3ω6) | DHGLA | (8Z,11Z,14Z)-Icosa-8,11,14-trienoic acid |  |
| Arachidonic Acid (C20:4ω6) | AA | (5Z,8Z,11Z,14Z)-Icosa-5,8,11,14-tetraenoic acid |  |
| Eicosapentaenoic acid (C20:5ω3) | EPA | (5Z,8Z,11Z,14Z,17Z)-Icosa-5,8,11,14,17-pentaenoic acid |  |
| Docosatetraenoic acid (C22:4ω6) | DTA | (7Z,10Z,13Z,16Z)-Docosa-7,10,13,16-tetraenoic acid |  |
| Docosahexaenoic acid (C22:6ω3) | DHA | (4Z,7Z,10Z,13Z,16Z,19Z)-Docosa-4,7,10,13,16,19-hexaenoic acid |  |
| Docosapentaenoic acid (C22:5ω3) | DPA | (7Z,10Z,13Z,16Z,19Z)-Docosa-7,10,13,16,19-pentaenoic acid |  |
| Overexpression | |||||
| Gene | Pathway | Microalgal Species | Results | Reference | |
| AGPAT1 | TAG biosynthesis | P. tricornutum | +50% EPA, +50% DHA content, +80% TAG content | [91] | |
| GPAT | TAG biosynthesis | P. tricornutum | +41% PUFA content | [92] | |
| DGAT2 | TAG biosynthesis | P. tricornutum | +76% EPA content, +35% neutral lipid content | [93] | |
| DGAT2 | TAG biosynthesis | P. tricornutum | +100% total lipid, +80% TAG content, +20% EPA content | [94] | |
| DGAT2 | TAG biosynthesis | N. oceanica | +69% neutral lipid content | [95] | |
| DGAT1A | TAG biosynthesis | N. oceanica | +39% TAG content | [96] | |
| DGTT1-3 | TAG biosynthesis | C. reinhardtii | no changes in lipid content | [97] | |
| DGTT4 | TAG biosynthesis | C. reinhardtii | +2800%TAG content | [98] | |
| GPAT, DGAT2 | TAG biosynthesis | P. tricornutum | +170% total lipid content | [99] | |
| GK | TAG biosynthesis | F. solaris | +12% total lipid content | [100] | |
| GPDH | TAG biosynthesis | P. tricornutum | +60% TAG content | [101] | |
| LPAAT1 | TAG biosynthesis | C. reinhardtii | +20% TAG content | [102] | |
| thioesterase | FA biosynthesis | P. tricornutum | +72% total lipid, +10% EPA content | [103] | |
| TE | FA biosynthesis | P. tricornutum | +16% EPA content | [103] | |
| ACP, KAS, FAT | FA biosynthesis | H. pluvialis | +100% EPA, +340%DHA content, +32% total FA content | [104] | |
| ME | FA biosynthesis | P. tricornutum | +150% total lipid content, −10% PUFA content | [105] | |
| MCAT | FA biosynthesis | N. oceanica | +31% neutral lipid, +8% EPA content | [106] | |
| MCAT | FA biosynthesis | Schizochytrium sp. | +172.5% EPA, +81.5% DHA, +69.2% DPA content | [107] | |
| FA elongase | FA biosynthesis | T. pseudonana | +40% EPA, +350% DHA content | [108] | |
| Δ5 desaturase | FA biosynthesis | P. tricornutum | +65% TAG content, +58% EPA content | [109] | |
| Δ12 desaturase | FA biosynthesis | N. oceanica | +75% AA content | [110] | |
| bHLH2 | transcription factor | N. salina | +33% total lipid content | [111] | |
| DOF | transcription factor | C. reinhardtii | +100% total lipid content | [112] | |
| DOF | transcription factor | C. reinhardtii | +170% total lipid content | [113] | |
| PSR1 | transcription factor | C. reinhardtii | no quantified reduction of neutral lipid content | [114] | |
| PSR1 | transcription factor | C. reinhardtii | +10% TAG content | [115] | |
| bZIP | transcription factor | N. salina | +50% total lipid content | [116] | |
| PNPLA3 | lipid turnover | P. tricornutum | +70% neutral lipid, +26% PUFA content | [117] | |
| LDP1 | lipid droplet metabolism | P. tricornutum | +30% total lipid, +40% neutral lipid content | [118] | |
| ACCase | pyruvate metabolism | C. cryptica | no changes in lipid content | [119] | |
| G6PD | carbohydrates metabolism | P. tricornutum | +170% total lipid content | [120] | |
| NOA | nitric oxide metabolism | P. tricornutum | +80% neutral lipid, +400% TAG content | [121] | |
| Silencing | |||||
|---|---|---|---|---|---|
| Gene | Pathway | Method | Microalgal Species | Results | Reference |
| AGPase | Carbohydrates’ metabolism | random mutagenesis | C. reinhardtii | +250% total lipids, +900% TAG content | [122] |
| isoamylase | Carbohydrates’ metabolism | random mutagenesis | C. reinhardtii | +450% total lipid content | [123] |
| UGPase | Carbohydrates’ metabolism | TALEN | P. tricornutum | +4400% TAG content | [124] |
| UGPase | Carbohydrates’ metabolism | RNAi | P. tricornutum | +4% total lipid content | [125] |
| CS | Carbohydrates’ metabolism | RNAi | T. pseudonana | +200% TAG content | [126] |
| SLM1 | Carbohydrates’ metabolism | random mutagenesis | S. obliquus | +51% TAG content | [127] |
| PEPC1 | pyruvate metabolism | CRISPRi | C. reinhardtii | +74% total lipid content | [128] |
| PEPC1 | pyruvate metabolism | RNAi | C. reinhardtii | +20% TAG content | [129] |
| PEPC1, PEPC2 | pyruvate metabolism | RNAi | C. reinhardtii | +48% FA content | [130] |
| PEPCK | pyruvate metabolism | RNAi | P. tricornutum | +40% total lipid content | [131] |
| CIS | pyruvate metabolism | RNAi | C. reinhardtii | +170% TAG content | [129] |
| PDK | pyruvate metabolism | RNAi | P. tricornutum | +82% neutral lipid, no changes in FA content | [132] |
| lipase | lipid turnover | RNAi | T. pseudonana | +300% EPA, +220% DHA content | [133] |
| omTGL | lipid turnover | RNAi | P. tricornutum | +70% EPA content | [134] |
| TGL1 | lipid turnover | RNAi | P. tricornutum | +200% TAG, +10% EPA content | [135] |
| LIP1 | lipid turnover | RNAi | C. reinhardtii | +150% TAG content | [136] |
| ACX2 | Β oxidation | insertional mutagenesis | C. reinhardtii | +400% neutral lipid, +70% TAG content | [137] |
| MLDP | lipid droplet metabolism | RNAi | C. reinhardtii | no changes in TAG content | [138] |
| LDP1 | lipid droplet metabolism | RNAi | P. tricornutum | −20% total lipid content | [130] |
| PDAT | TAG biosynthesis | RNAi | C. reinhardtii | general reduction of all TAG classes content | [139] |
| SAD | FA biosynthesis | RNAi | C. reinhardtii | +40% stearic acid content | [140] |
| ω-3-DES | FA biosynthesis | homologous recombination | C. vulgaris | no changes in PUFA and FA content | [141] |
| TES1 | FA biosynthesis | TALEN | P. tricornutum | +70% TAG content | [142] |
| PDH | FA biosynthesis | RNAi | C. reinhardtii | −50% FA content | [143] |
| DGTT | FA biosynthesis | RNAi | C. reinhardtii | −35% TAG content | [144] |
| NR | N assimilation | TALEN | P. tricornutum | +20% TAG content | [145] |
| NR | N assimilation | RNAi | P. tricornutum | +43% total lipid content | [146] |
| ZnCys | transcription factor | RNAi | N. gaditana | +35% total lipid content | [147] |
| - | - | random mutagenesis | P. lutheri | +33% EPA, +33% DHA content | [148] |
| - | - | insertional mutagenesis | N. oceanica | +180% PUFA, +40% EPA content | [149] |
| Heterologous expression | |||||
| Gene | Pathway | Source species | Receiver species | Results | Reference |
| Genes from microalgae in other microalgae | |||||
| GPAT | TAG biosynthesis | L. incisa | C. reinhardtii | +50% FA content | [150] |
| ELO5 | FA biosynthesis | O. tauri | P. tricornutum | +700% DHA content | [151] |
| ELO5, DES6 | FA biosynthesis | O. tauri | P. tricornutum | +800% DHA content | [151] |
| Δ5DES | FA biosynthesis | T. aureum | A. limacinum | +360% EPA, +1220% AA content | [152] |
| ME | FA biosynthesis | P. tricornutum | C. pyrenoidosa | +220% neutral lipid content | [105] |
| (Bn)AccD, (Cr)ME | pyruvate metabolism | B. napus, C. reinhardtii | D. salina | +12% total lipid content | [153] |
| ACCase | pyruvate metabolism | C. cryptica | N. saprophila | no changes in lipid content | [119] |
| thioesterase | FA biosynthesis | D. tertiolecta | C. reinhardtii | +50% FA content | [154] |
| DGAT2 | TAG biosynthesis | C. reinhardtii | S. obliquus | +85% total lipid content | [155] |
| Genes from other organisms in microalgae | |||||
| DGAT2 | TAG biosynthesis | B. napus (plant) | P. tricornutum | +12% ALA content | [156] |
| DGA1 | TAG biosynthesis | S. cerevisiae (yeast) | P. tricornutum | +130% TAG content | [157] |
| OLEO3 | TAG biosynthesis | A. thaliana (plant) | P. tricornutum | +40% TAGcontent | [157] |
| (Sc)DGA1, (At)OLEO3 | TAG biosynthesis | S. cerevisiae (yeast), A. thaliana (plant) | P. tricornutum | +260% TAG content | [157] |
| (Sc)G3PDH-GPAT-LPAAT, (Yl)DGATs | TAG biosynthesis | S. cerevisiae, Y. lipolytica (yeasts) | C. minutissima | +120% total lipid content | [158] |
| Δ3DES | FA biosynthesis | S. dicilina (yeast) | Schizochytrium sp. | +3% DHA content | [159] |
| ACP reductase | FA biosynthesis | Synechocystis sp. (cyanobacteria) | C. merolae | +133% TAG content | [160] |
| (Cc)C14-TE, (Uc)C12-TE | FA biosynthesis | C. camphora, U. californica (plants) | P. tricornutum | +80% TAG content | [161] |
| (Cc)C14-TE, (Uc)C12-TE, (Ch)KAS | FA biosynthesis | C. camphora, U. californica, C. hookeriana (plants) | D. tertiolecta | +4% FA content | [162] |
| C14-TE, C10-TE, ACP | FA biosynthesis | C. lanceolata (plant) | C. reinhardtii | general increase in different FAs classes content | [163] |
| (Bn)AccD, (Cr)ME | pyruvate metabolism | B. napus (plant), C. reinhardtii | D. salina | +12% total lipid content | [153] |
| ACC1 | pyruvate metabolism | S. cerevisiae (yeast) | S. quadricauda | +60% FA content | [164] |
| ACC1, GDP1, GUT1 | pyruvate metabolism | S. cerevisiae (yeast) | S. quadricauda | +50% total lipid content | [164] |
| ACS | pyruvate metabolism | E. coli (bacteria) | Schizochytrium sp. | no changes in lipid content | [165] |
| (An)PhyA, (Ot)Elo5 | phytate metabolism | A. niger (yeast), O. tauri | P. tricornutum | +10% DHA, −25% EPA content | [166] |
| (Ec)AppA, (Ot)Elo5 | phytate metabolism | E. coli (bacteria), O. tauri | P. tricornutum | +12% DHA, −18% EPA content | [166] |
| DOF4 | transcription factor | G. max (plant) | C. ellipsoidea | +53% total lipid content | [167] |
| WRI1 | transcription factor | A. thaliana (plant) | N. salina | +64% tota lipid content | [168] |
| Genes from microalgae in other organisms | |||||
| antisense PEPC | pyruvate metabolism | Anabaena sp. | E. coli (bacteria) | +47% lipid content | [169] |
| ACCase | pyruvate metabolism | P. tricornutum | E. coli (bacteria) | +100% neutral lipid content | [170] |
| Δ9-ELO | FA biosynthesis | I. galbana | A. thaliana (plant) | +18% PUFA content | [171] |
| Δ9-ELO (codon optimized) | FA biosynthesis | I. galbana | A. thaliana (plant) | +64% PUFA content | [172] |
| Δ6-DES | FA biosynthesis | M. pusilla | A. thaliana (plant) | +26% EPA content | [173] |
| (Ig)Δ9E, (Eg)Δ8D, (Ma)Δ5D | FA biosynthesis | I. galbana, E. gracilis, M. alpina | A. thaliana (plant) | +23% PUFA, +3% EPA, +7% AA content | [171] |
| (Pt)Δ5D, (Pt)Δ6D, (Pp)Δ6E | FA biosynthesis | P. tricornutum, Physcomitrella patens | N. tabacum (plant) | +30% PUFA content | [174] |
| (Pt)Δ5D, (Pt)Δ6D, (Pp)Δ6E | FA biosynthesis | P. tricornutum, P. patens | L. usitatissimum (plant) | +30% PUFA content | [174] |
| (Sc)PUFA-synthase, (No)PPTase | FA biosynthesis | Schizochytrium sp., Nostoc sp. | A. thaliana (plant) B. napus (plant) | +4% DHA, +1% EPA content | [175] |
| ELO5 | FA biosynthesis | P. tricornutum | P. pastoris (yeast) | no quantified increase in DPA and DTA content | [176] |
| (Pt)ELO5, (Is)DES4 | FA biosynthesis | P. tricornutum, I. sphaerica | P. pastoris (yeast) | +3% DPA, +2.35% DHA content | [176] |
| DES2 | FA biosynthesis | C. vulgaris | S. cerevisiae (yeast) | no quantified reduction of LA content | [177] |
| DGTT2 | FA biosynthesis | C. reinhardtii | S. cerevisiae (yeast) | +800% TAG content | [178] |
| (Pt)Δ5D, (Pt)Δ6D, (Pp)Δ6E | FA biosynthesis | P. tricornutum, P. patens | S. cerevisiae (yeast) | +0.23% EPA, +0.17% AA content | [179] |
| DES6 | FA biosynthesis | M. pusilla | M. alpina (yeast) | +2500% EPA content | [180] |
| Δ6ELO | FA biosynthesis | Isochrysis sp. | E. coli (bacteria) | +6% SDA, +3% GLA content | [181] |
| (Iso)Δ6ELO, (Pav)Δ5DES | FA biosynthesis | Isochrysis sp., Pavlova sp. | E. coli (bacteria) | no quantified increase of AA and EPA content | [182] |
| Microalgal Species | Application (as Feed or Food) | Innovation | Nutritional Value (Respect to Basal Diet) | Reference |
|---|---|---|---|---|
| generic microalgae | Feed for light lamb Ovis aries farming | mixed with extruded linseed could, in part, replace fish meat | +520% ALA | [217] |
| generic microalgae | Feed for Pacific lamprey Entosphenus tridentatus aquaculture | complete replacement of yeast + fish oil | +21% total LC-PUFA; +9% total ω3 PUFA; +11% EPA | [218] |
| Schizochytrium sp. | Feed for Atlantic salmon Salmo salar aquaculture | complete replacement of fish oil | +100% DHA | [219] |
| Schizochytrium sp. | Feed for Nile tilapia Oreochromis niloticus aquaculture | complete replacement of fish meat | +23% total PUFA; +30.2% DHA | [220] |
| Schizochytrium sp. | Feed for tambaqui Colossoma macropomum aquaculture | complete replacement of fish meat | +300% total ω3 PUFA; +200% PA; +126% EPA; +51,200% DHA; +512% ω3:ω6 | [221] |
| Schizochytrium sp. | Feed for Atlantic salmon Salmo salar aquaculture | the complete replacement of fish | +6% SFA; +3% DHA; +1% MUFA; +1% total ω6 PUFA | [209] |
| Schizochytrium sp. | Feed for Rainbow Trout Oncorhynchus mykiss aquaculture | complete replacement of fish meat | +18% total PUFA | [222] |
| Schizochytrium sp. | Feed for red seabream Pagrus major aquaculture | complete replacement of fish meat | +130% SFA; +180% PA; +100% DHA; +2070% DHA:EPA | [223] |
| Schizochytrium sp. | Feed for broiler chicken Gallus domesticus farming | 0.2% Schiochytrium inclusion | +2.5% total USFA | [224] |
| Schizochytrium sp. | Feed for channel catfish Ictalurus punctatus aquaculture | +2% dried Schizochytrium | +3.71% total ω3 LC-PUFA; +3.35% DHA | [225] |
| Schizochytrium sp. | Feed for shrimps Litopenaeus vannamei aquaculture | until 75% replacement of fish meat | +100% total ω6 PUFA; +200% DHA | [226] |
| Schizochytrium sp. | Feed for light lamb Ovis aries farming | 3.8% Schizochytrium and 5% linseed inclusion | +400% ALA; +6500% DHA | [227] |
| Schizochytrium sp. | Feed for rabbit Oryctolagus sp. farming | 4 gr Schizochytrium per kg feed | +50% EPA; +180% DHA | [228] |
| Schizochytrium sp. | Feed for Atlantic salmon Salmo salar aquaculture | 50% Schizochytrium inclusion | +2% PA; +22% total ω6 PUFA; +15% total PUFA; +340% DHA:EPA | [229] |
| Schizochytrium limacinum | Feed for grouper Epinephelus lanceolatus aquaculture | in combination with soybean meal, soy protein concentrate could replace 40% of fish meat | +100% DHA; +550% DHA:EPA | [230] |
| Schizochytrium limacinum | General aquaculture feed | 48% v/v effluent concentration from biofuel industry | +80% SA; +120% DHA | [231] |
| Aurantiochytrium sp. | Feed for black tiger shrimp Penaeus monodon aquaculture | 1–2% Aurantiochytrium inclusion | +200% total MUFA; +10% total SFA; +12% total PUFA; +20% total ω3 PUFA; +37% DHA | [232] |
| Isochrysis sp. | Feed for European seabass Dicentrarchus labrax aquaculture | 20% of protein and 36% of lipid could be replaced using the freeze-dried Isochrysis | +4% total ω3 PUFA; +13% total SFA | [233] |
| Isochrysis galbana | Feed for silverfish Trachinotus ovatus aquaculture | 24–26% fish oil replacement | +10% DHA; +10% total LC-ω3 PUFA | [234] |
| Phaeodactylum tricornutum | Feed for Atlantic salmon Salmo salar aquaculture | 6% dried Phaeodactylum inclusion | same nutritional content | [215] |
| Phaeodactylum tricornutum or Crypthecodinium cohnii | Feed for gilthead seabream Sparus aurata aquaculture | 2–5% Crypthecodinium or Phaeodactylum inclusion | +16% total SFA; +20% DHA | [235] |
| Haematococcus pluvialis | Feed for Rainbow Trout Oncorhynchus mykiss aquaculture | 10 gr Haematococcus per kg feed | - | [236] |
| Staurosira sp. | Feed for broiler chicken Gallus domesticus farming | 7.5% Staurosira inclusion | +25% total lipids | [237] |
| Arthrospira platensis | Feed for Nile tilapia Oreochromis niloticus aquaculture | 30% Arthrospira inclusion | - | [238] |
| Spirulina platensis | Feed for broiler chicken Gallus domesticus farming | 10 gr Spirulina per kg feed | same nutritional content | [239] |
| Spirulina platensis or Chlorella vulgaris | Feed for African catfish Clarias gariepinus aquaculture | ultill 75% Spirulina or Chlorella inclusion | +20% total ω6 PUFA; +50% DHA | [240] |
| Chlorella sp. | Feed for crucian carp Carassius auratus aquaculture | in combination with 2 gr cellulases per kg could completely replace fish meat | - | [241] |
| Chlorella spp. | Feed for Channel Catfish Ictalurus punctatus aquaculture | 15% Chlorella inclusion | +75% PA; +30% OA; +100% EPA; +70% DHA; +32% total ω3 PUFA | [242] |
| Chlorella sp. or Nannochloropsis sp. | Feed for European seabass Dicentrarchus labrax aquaculture | 15% Chlorella or Nannochloropsis inclusion | +2% total ω6 PUFA | [243] |
| Nannochloropsis gaditana | Feed for Nile tilapia Oreochromis niloticus aquaculture | 30% Nannochloropsis inclusion | - | [244] |
| Nannochloropsis gaditana | Feed for gilthead seabream Sparus aurata aquaculture | 2% Nannochloropsis inclusion | +80% EPA; +200% EPA:DHA | [245] |
| Nannochloropsis sp. | Feed for kuruma shrimp Marsupenaeus japonicus aquaculture | 4–7–10% Nannochloropsis biomass or lipid inclusion | +13% total ω3 PUFA; +44% total ω6 PUFA; +37% EPA | [246] |
| Nannochloropsis sp. | Feed for European seabass Dicentrarchus labrax aquaculture | 5–10–15% Nannochloropsis inclusion | same nutritional content | [247] |
| Nannochloropsis salina | Feed for Nile tilapia Oreochromis niloticus aquaculture | complete replacement of fish meat | +47% total ω6 PUFA; +130% EPA | [248] |
| Nannochloropsis sp. or Pavlova viridis | Feed for European seabass Dicentrarchus labrax aquaculture | complete replacement of fish meat | +50% total PUFA; +70% total ω6 PUFA | [249] |
| Pavlova lutheri | Feed for oyster Crassostrea gigas aquaculture | complete replacement of fish meat | - | [250] |
| Tetraselmis suecica | Feed for shrimps Litopenaeus vannamei aquaculture | complete replacement of fish meat | - | [251] |
| Tetraselmis chuii | Feed for shrimps Litopenaeus vannamei aquaculture | 50% Tetraselmis inclusion | +2% total lipids | [252] |
| Tetraselmis sp. | Feed for gilthead seabream Sparus aurata aquaculture | 10% Tetraselmis inclusion | same total lipids | [253] |
| Tetraselmis suecica or Tisochrysis lutea | Feed for European seabass Dicentrarchus labrax aquaculture | combination of both microalgae for a complete replacement of fish meat | - | [254] |
| Desmodesmus sp. | Feed for Atlantic salmon Salmo salar aquaculture | 20% Desmodesmus inclusion | - | [255] |
| Aurantiochytrium sp. and Schizochytrium sp. | Feed for gilthead seabream Sparus aurata aquaculture | blend of poultry and one of two algal oils | +200% total ω6 LC-PUFA; +3% PA; +500% DHA:EPA | [256] |
| Nannochloropsis sp, Isochrysis sp. and Schizochytrium sp. | Feed for Rainbow Trout Oncorhynchus mykiss aquaculture | blend of three microalgae for a complete replacement of fish meat | +80% total PUFA; +170% total ω6 PUFA | [257] |
| Nannochloropsis sp. and Schizochytrium sp | Feed for Nile tilapia Oreochromis niloticus aquaculture | Blend of both microalgae for a complete replacement of fish meat | +20% PA; +50% DHA; +20% total ω6 LC-PUFA | [211] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santin, A.; Russo, M.T.; Ferrante, M.I.; Balzano, S.; Orefice, I.; Sardo, A. Highly Valuable Polyunsaturated Fatty Acids from Microalgae: Strategies to Improve Their Yields and Their Potential Exploitation in Aquaculture. Molecules 2021, 26, 7697. https://doi.org/10.3390/molecules26247697
Santin A, Russo MT, Ferrante MI, Balzano S, Orefice I, Sardo A. Highly Valuable Polyunsaturated Fatty Acids from Microalgae: Strategies to Improve Their Yields and Their Potential Exploitation in Aquaculture. Molecules. 2021; 26(24):7697. https://doi.org/10.3390/molecules26247697
Chicago/Turabian StyleSantin, Anna, Monia Teresa Russo, Maria Immacolata Ferrante, Sergio Balzano, Ida Orefice, and Angela Sardo. 2021. "Highly Valuable Polyunsaturated Fatty Acids from Microalgae: Strategies to Improve Their Yields and Their Potential Exploitation in Aquaculture" Molecules 26, no. 24: 7697. https://doi.org/10.3390/molecules26247697
APA StyleSantin, A., Russo, M. T., Ferrante, M. I., Balzano, S., Orefice, I., & Sardo, A. (2021). Highly Valuable Polyunsaturated Fatty Acids from Microalgae: Strategies to Improve Their Yields and Their Potential Exploitation in Aquaculture. Molecules, 26(24), 7697. https://doi.org/10.3390/molecules26247697