
Chemical Profile of Launaea nudicaulis Ethanolic Extract and Its Antidiabetic Effect in Streptozotocin-Induced Rats

 , , ,
, , ,  ,
, 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Acute Toxicity
2.2. Effect on Blood Glucose Levels
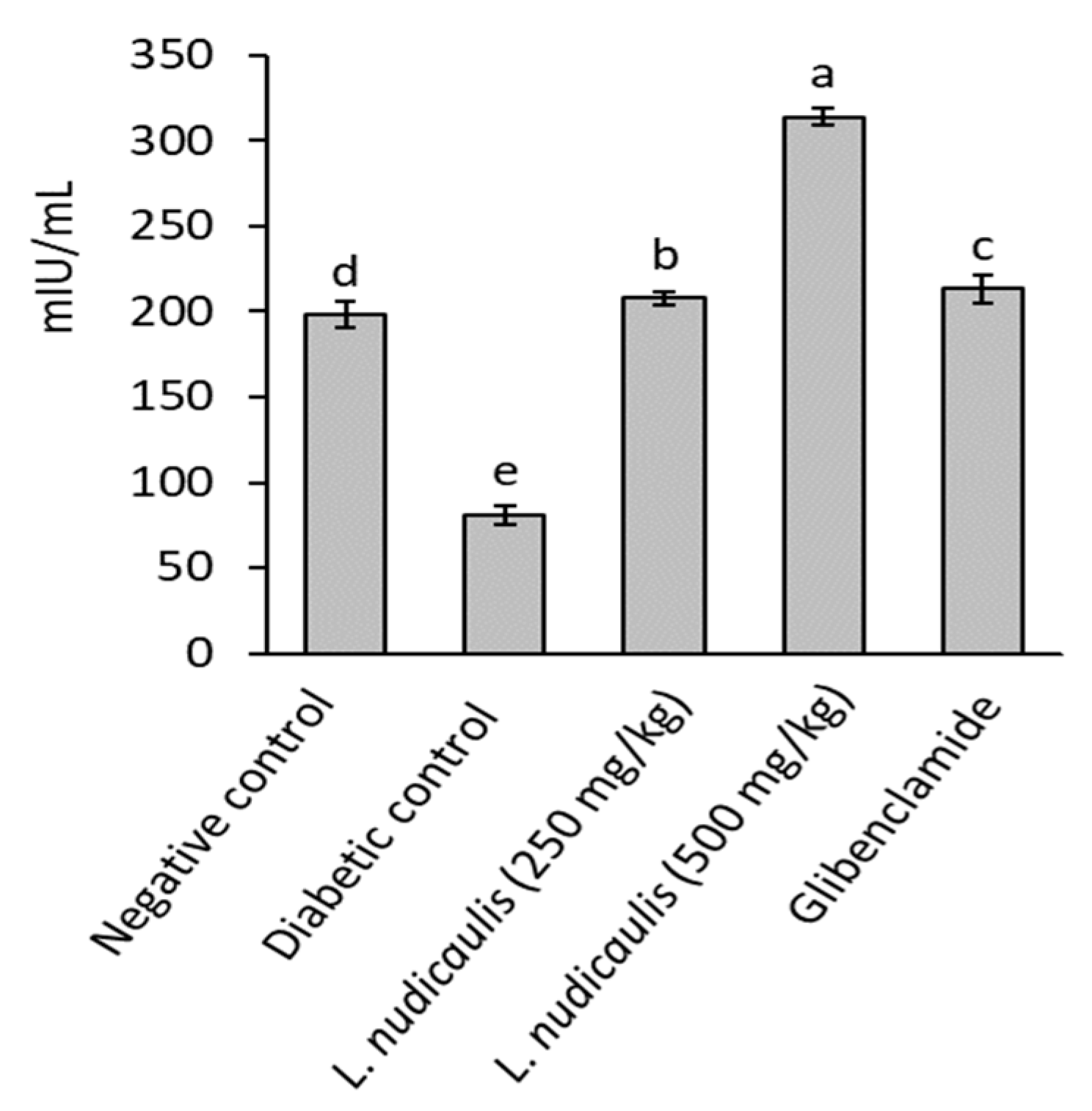
2.3. Effect on Serum Insulin Levels
2.4. Effect on Rats Weekly Body Weight
2.5. Effect on Lipid Profile
2.6. Effect on Liver and Kidney Functions
2.7. Effect On Relative Weight of Vital Organs
2.8. Effect on Serum Antioxidant and Oxidative Stress Biomarkers
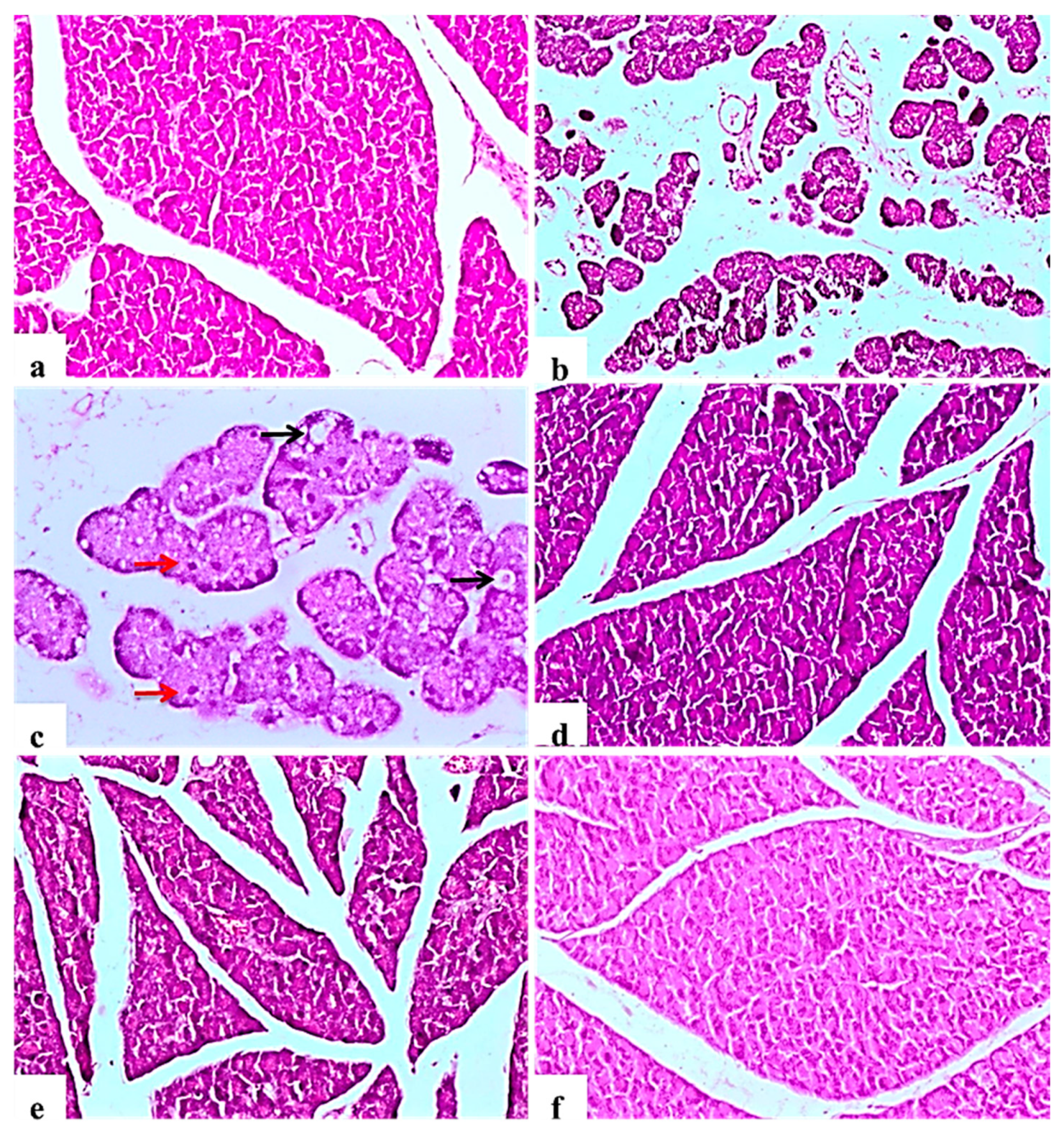
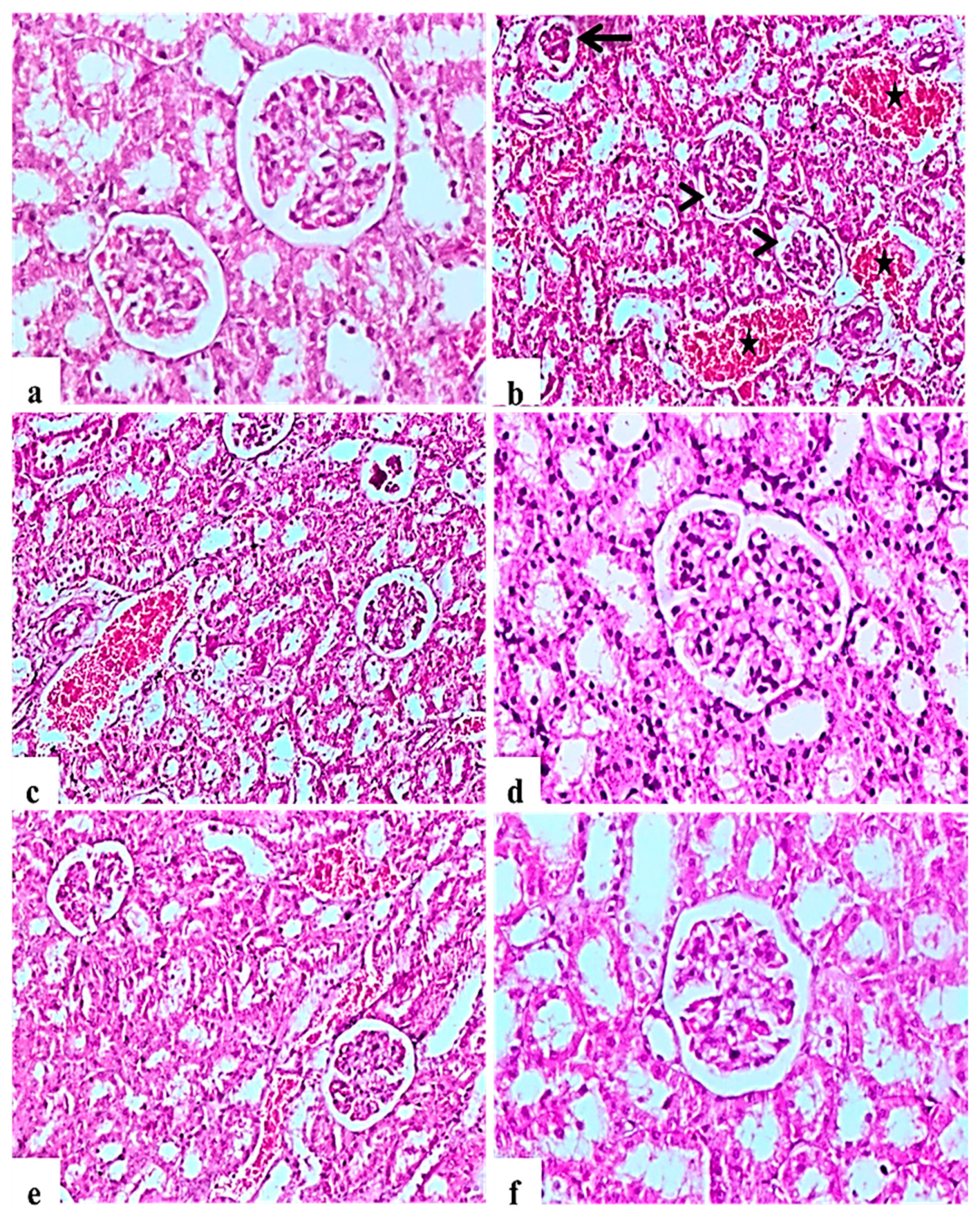
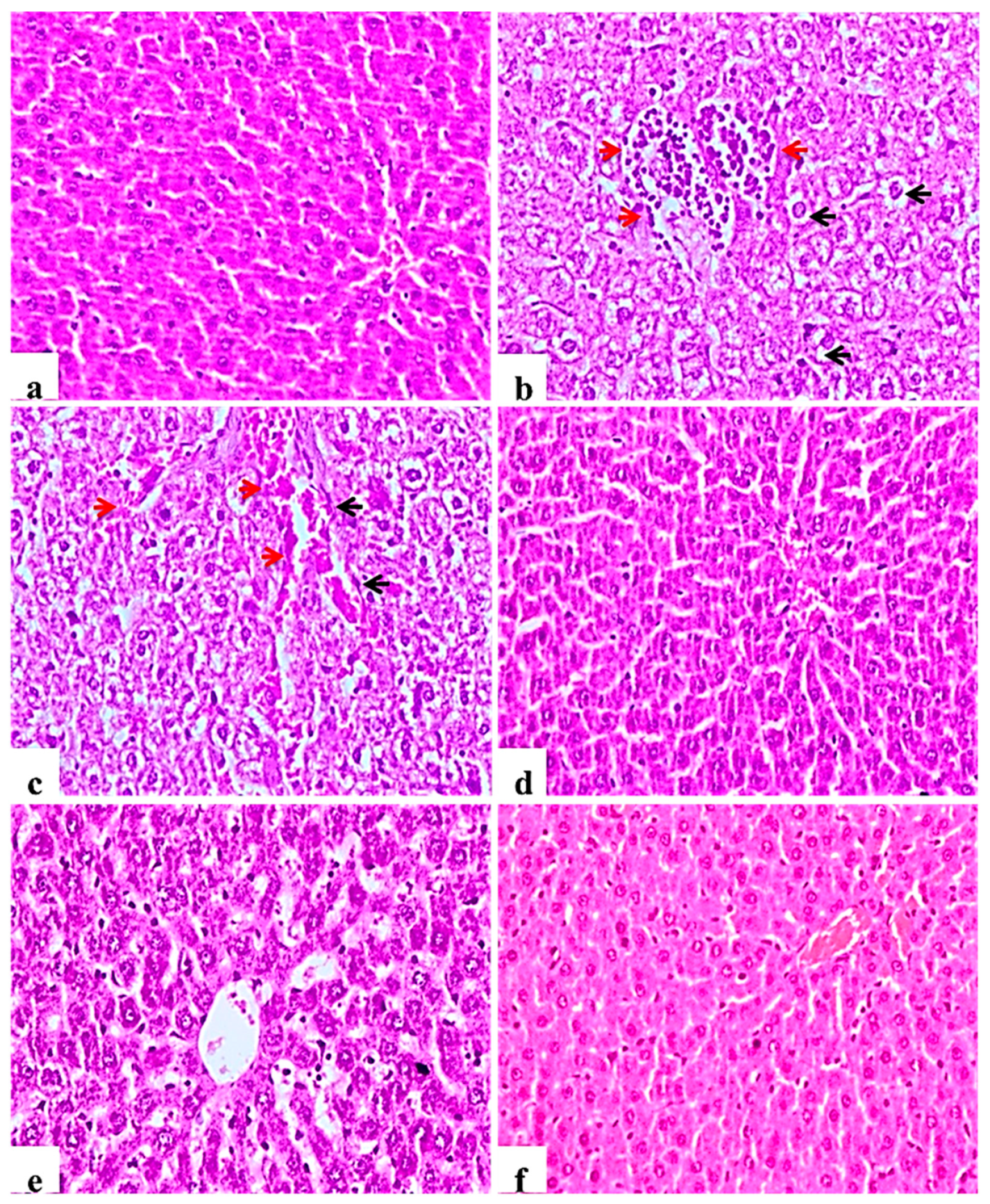
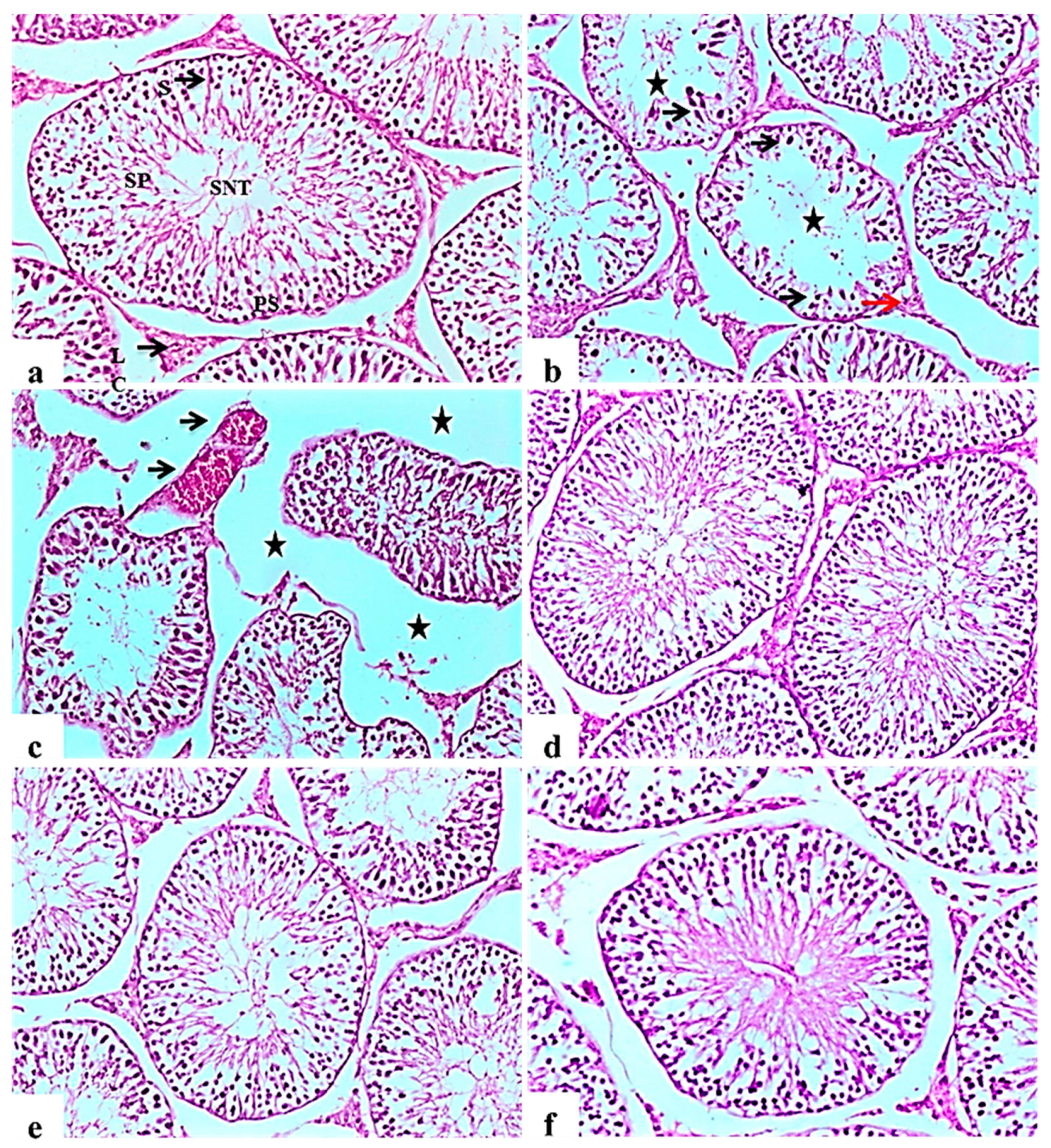
2.9. Histological Examination of Pancreas, Liver, Kidney, and Testis
2.9.1. Pancreas
2.9.2. Kidney
2.9.3. Liver
2.9.4. Testis
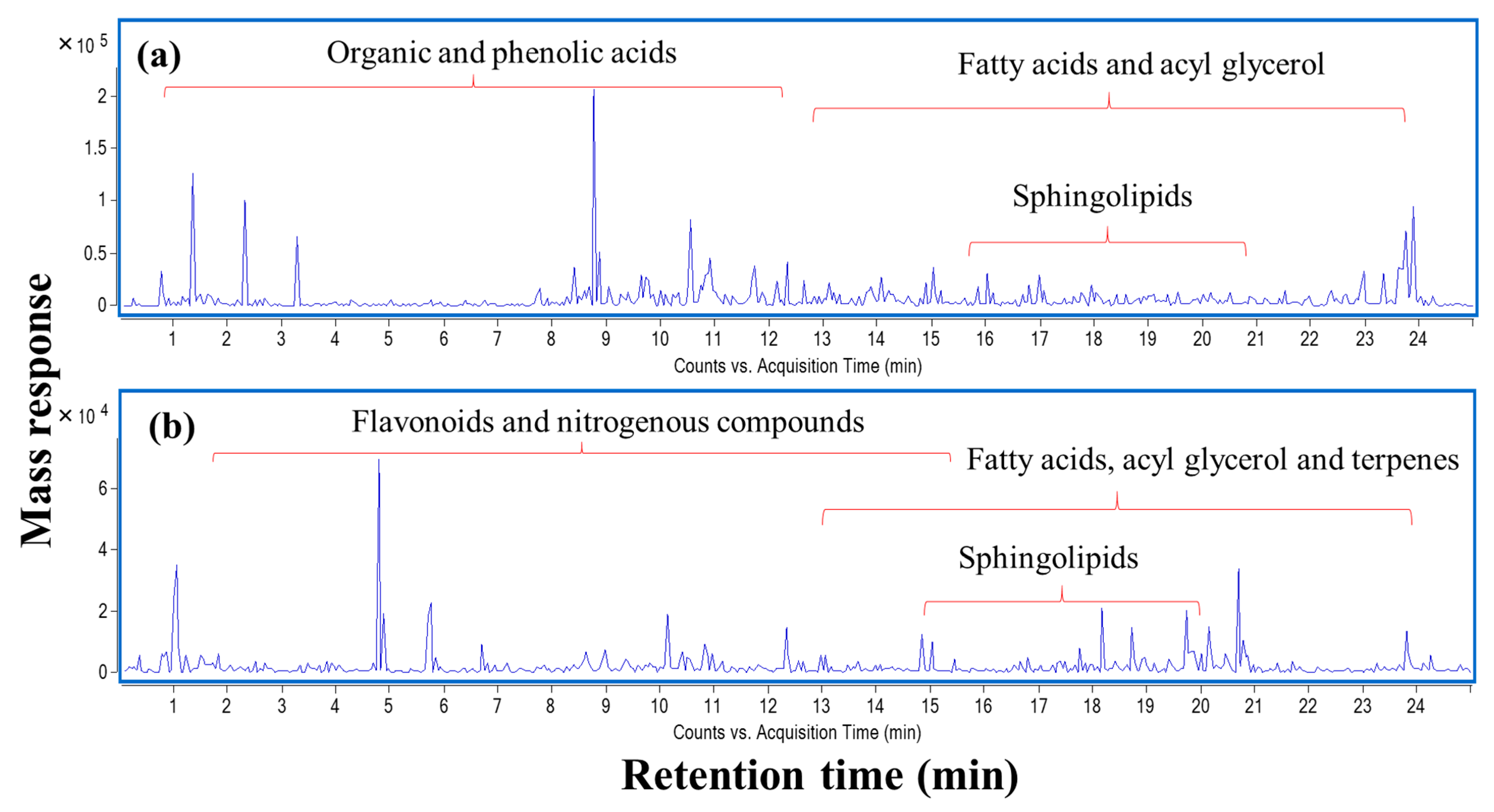
2.10. Comprehensive Metabolites Profiling of L. nudicaulis Extract
2.10.1. Flavonoids
2.10.2. Simple Phenolics
2.10.3. Fatty Acids and Sphingolipids
2.10.4. Amino Acids/Peptides
2.10.5. Organic Acids
2.10.6. Nitrogenous Compounds
3. Materials and Methods
3.1. Plant Materials Collection, Identification, and Extract Preparation
3.2. Chemicals
3.3. Experimental Animals
3.4. Acute Toxicity (LD50)
3.5. Hypoglycemic Experiment
3.5.1. Diabetes Induction
3.5.2. The L. nudicaulis Extract and Standard Drug Dosing Protocol
3.5.3. Experimental Design
3.5.4. Measurement of Blood Glucose Levels
3.5.5. Determination of Serum Insulin Level and Serum Lipid Profile
3.5.6. Determination of Liver and Kidney Functions
3.5.7. Determination of Antioxidant and Oxidative Stress Biomarkers of Serum
3.6. Histopathological Examination
3.7. High-Resolution Ultra-Performance Liquid Chromatography-Mass Spectrometry Analysis (UPLC-ESI–qTOF-MS)
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [Green Version]
- Thomas, R.; Halim, S.; Gurudas, S.; Sivaprasad, S.; Owens, D. IDF Diabetes Atlas: A review of studies utilising retinal photography on the global prevalence of diabetes related retinopathy between 2015 and 2018. Diabetes Res. Clin. Pract. 2019, 157, 107840. [Google Scholar] [CrossRef]
- Osawa, T.; Kato, Y. Protective role of antioxidative food factors in oxidative stress caused by hyperglycemia. Ann. N. Y. Acad. Sci. 2005, 1043, 440–451. [Google Scholar] [CrossRef]
- Yin, P.; Zhao, S.; Chen, S.; Liu, J.; Shi, L.; Wang, X.; Liu, Y.; Ma, C. Hypoglycemic and hypolipidemic effects of polyphenols from burs of Castanea mollissima Blume. Molecules 2011, 16, 9764–9774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Haleem, S.A.; Ibrahim, A.Y.; Ismail, R.F.; Shaffie, N.M.; Hendawy, S.; Omer, E. In-vivo hypoglycemic and hypolipidemic properties of Tagetes lucida alcoholic extract in streptozotocin-induced hyperglycemic Wistar albino rats. Ann. Agric. Sci. 2017, 62, 169–181. [Google Scholar] [CrossRef]
- Ricci, G.; Catizone, A.; Esposito, R.; Pisanti, F.; Vietri, M.T.; Galdieri, M. Diabetic rat testes: Morphological and functional alterations. Andrologia 2009, 41, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Shojaei Andebili, S.; Dehghani Firoozabadi, A.; Behnam Rassouli, M.; Mahdavi SHahri, N.; Haghparast, A. Morphological evaluation of testis tissue of rats in various time points after diabetes type 1 induction. Iran. J. Diabetes Obes. 2014, 5, 98–106. [Google Scholar]
- Farrag, A.R.H.; Abdallah, H.M.; Khattab, A.R.; Elshamy, A.I.; El Gendy, A.E.-N.G.; Mohamed, T.A.; Farag, M.A.; Efferth, T.; Hegazy, M.-E.F. Antiulcer activity of Cyperus alternifolius in relation to its UPLC-MS metabolite fingerprint: A mechanistic study. Phytomedicine 2019, 62, 152970. [Google Scholar] [CrossRef] [PubMed]
- Al-Mahrezi, J.A.; Al-Sabahi, J.N.; Akhtar, M.S.; Selim, D.; Weli, A.M.; JIJoPS, R. Essential oil composition and antimicrobial screening of Launaea nudicaulis grown in Oman. Int. J. Pharm. Sci. Res. 2011, 2, 3166–3169. [Google Scholar]
- Elshamy, A.I.; Abd-ElGawad, A.M.; El-Amier, Y.A.; El Gendy, A.E.N.G.; Al-Rowaily, S.L. Interspecific variation, antioxidant and allelopathic activity of the essential oil from three Launaea species growing naturally in heterogeneous habitats in Egypt. Flavour Fragr. J. 2019, 34, 316–328. [Google Scholar] [CrossRef]
- Cole, M. Key antifungal, antibacterial and anti-insect assays—A critical review. Biochem. Syst. Ecol. 1994, 22, 837–856. [Google Scholar] [CrossRef]
- El-Sharkawy, E.R.; Ed-Dra, A.; Abdallah, E.M. Phytochemical, antimicrobial and antioxidant properties of Launaea nudicaulis and Farsetia hamiltonii. J. Biol. Control 2017, 31, 102–109. [Google Scholar] [CrossRef] [Green Version]
- Saleem, M.; Parveen, S.; Riaz, N.; Tahir, M.N.; Ashraf, M.; Afzal, I.; Ali, M.S.; Malik, A.; Jabbar, A. New bioactive natural products from Launaea nudicaulis. Phytochem. Lett. 2012, 5, 793–799. [Google Scholar] [CrossRef]
- Chauhan, A.; Sharma, P.; Srivastava, P.; Kumar, N.; Dudhe, R. Plants having potential antidiabetic activity: A review. Der Pharm. Lett. 2010, 2, 369–387. [Google Scholar]
- Cheriti, A.; Belboukhari, M.; Belboukhari, N.; Djeradi, H. Phytochemical and biological studies on Launaea Cass. genus (Asteraceae) from Algerian Sahara. Phytochemistry 2012, 11, 67–80. [Google Scholar]
- Sepehri-Moghadam, H.; Rahbarian, R.; Sadoughi, S.D. The effect of aqueous extract of Launaea acanthodes (Boiss.) O. Kuntze on the serum level of insulin and blood glucose and histomorphological changes of pancreas in diabetic rats. Feyz J. Kashan Univ. Med. Sci. 2015, 19, 30–37. [Google Scholar]
- Adinortey, M.B.; Sarfo, J.K.; Kwarteng, J.; Adinortey, C.A.; Ekloh, W.; Kuatsienu, L.E.; Kwadwo Nyarko, A. The ethnopharmacological and nutraceutical relevance of Launaea taraxacifolia (Willd.) Amin ex C. Jeffrey. Evid.-Based Complementary Altern. Med. 2018, 2018, 7259146. [Google Scholar] [CrossRef]
- Abdallah, H.M.I.; Elshamy, A.I.; El Gendy, A.E.-N.G.; Abd El-Gawad, A.M.; Omer, E.A.; De Leo, M.; Pistelli, L. Anti-inflammatory, antipyretic, and antinociceptive effects of a Cressa cretica aqueous extract. Planta Med. 2017, 83, 1313–1320. [Google Scholar] [CrossRef] [PubMed]
- El-Newary, S.A.; Shaffie, N.M.; Omer, E. The protection of Thymus vulgaris leaves alcoholic extract against hepatotoxicity of alcohol in rats. Asian Pac. J. Trop. Med. 2017, 10, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Mohsen-Nia, M.; Amiri, H. Measurement and modelling of static dielectric constants of aqueous solutions of methanol, ethanol and acetic acid at T = 293.15 K and 91.3 kPa. J. Chem. Thermodyn. 2013, 57, 67–70. [Google Scholar] [CrossRef]
- Miguel, M.G.; Nunes, S.; Dandlen, S.A.; Cavaco, A.M.; Antunes, M.D. Phenols and antioxidant activity of hydro-alcoholic extracts of propolis from Algarve, South of Portugal. Food Chem. Toxicol. 2010, 48, 3418–3423. [Google Scholar] [CrossRef] [PubMed]
- Sahu, N.; Singh, A.; Khatoon, S.; Kumar, B.; Arya, K. Macro-microscopic and LCMS markers for identification and authentication of herbal formulations of Taraxacum officinale and Launaea procumbens. Int. J. Res. Pharm. Sci. 2017, 8, 301–310. [Google Scholar]
- Dhananjeyan, M.R.; Bykowski, C.; Trendel, J.A.; Sarver, J.G.; Ando, H.; Erhardt, P.W. Simultaneous determination of procaine and para-aminobenzoic acid by LC–MS/MS method. J. Chromatogr. B 2007, 847, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Fernández, R.; López-Martínez, J.C.; Romero-González, R.; Martínez-Vidal, J.L.; Flores, M.I.A.; Frenich, A.G. Simple LC–MS determination of citric and malic acids in fruits and vegetables. Chromatographia 2010, 72, 55–62. [Google Scholar] [CrossRef]
- Rhouati, S.; Ahmed, A.A.; Ouahrani, M.R. Isolation of coumarins and coumarin glucoside from Launaea resedifolia. Asian J. Chem. 2006, 18, 2348–2352. [Google Scholar]
- Pramai, P.; Thongsook, T.; Thanasukarn, P.; Jannoey, P.; Nuengchamnong, N.; Chen, F.; Maulidiani, M.; Abas, F.; Jiamyungyuen, S. Chemical profiles of three varieties of germinated rice based on LC-MS and their antioxidant activity. Food Appl. Biosci. J. 2019, 7, 11–32. [Google Scholar]
- Jun, M.; Shao, Y.; Ho, C.-T.; Koetter, U.; Lech, S. Structural identification of nonvolatile dimerization products of glucosamine by gas chromatography− mass spectrometry, liquid chromatography− mass spectrometry, and nuclear magnetic resonance analysis. J. Agric. Food Chem. 2003, 51, 6340–6346. [Google Scholar] [CrossRef]
- Xiao, B.; Dong, J.; Yan, M.; Liu, Y.; Li, Y.; Yang, G.; Sun, Z. Development of a liquid chromatography–tandem mass spectrometry method for simultaneous determination of five isoflavonoids and seven neurochemicals in rat brain dialysate and its application to a pharmacological study. J. Sep. Sci. 2018, 41, 2321–2330. [Google Scholar] [CrossRef]
- Odukoya, J.; Oshodi, A. Evaluation of the nutritional qualities of the leaves of Parquetina nigrescens, Launaea taraxacifolia and Solanum nigrum. Eur. J. Pure Appl. Chem. 2018, 5, 18–31. [Google Scholar]
- Gao, J.; Leung, K.S.; Wang, Y.; Lai, C.; Li, S.; Hu, L.; Lu, G.; Jiang, Z.; Yu, Z. Qualitative and quantitative analyses of nucleosides and nucleobases in Ganoderma spp. by HPLC–DAD-MS. J. Pharm. Biomed. Anal. 2007, 44, 807–811. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Kumar, S.; Pandey, R.; Hasanain, M.; Sarkar, J.; Kumar, B. Bioguided chemical characterization of the antiproliferative fraction of edible pseudo bulbs of Malaxis acuminata D. Don by HPLC-ESI-QTOF-MS. Med. Chem. Res. 2017, 26, 3307–3314. [Google Scholar] [CrossRef]
- Yamaoka, N.; Kaneko, K.; Kudo, Y.; Aoki, M.; Yasuda, M.; Mawatari, K.; Nakagomi, K.; Yamada, Y.; Yamamoto, T. Analysis of purine in purine-rich cauliflower. Nucleosides Nucleotides Nucleic Acids 2010, 29, 518–521. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.-Z.; Harnly, J.M. Identification of the phenolic components of chrysanthemum flower (Chrysanthemum morifolium Ramat). Food Chem. 2010, 120, 319–326. [Google Scholar] [CrossRef]
- t’Kindt, R.; De Veylder, L.; Storme, M.; Deforce, D.; Van Bocxlaer, J. LC–MS metabolic profiling of Arabidopsis thaliana plant leaves and cell cultures: Optimization of pre-LC–MS procedure parameters. J. Chromatogr. B 2008, 871, 37–43. [Google Scholar] [CrossRef]
- Ostrowski, W.; Wojakowska, A.; Grajzer, M.; Stobiecki, M. Mass spectrometric behavior of phenolic acids standards and their analysis in the plant samples with LC/ESI/MS system. J. Chromatogr. B 2014, 967, 21–27. [Google Scholar] [CrossRef]
- Toffali, K.; Zamboni, A.; Anesi, A.; Stocchero, M.; Pezzotti, M.; Levi, M.; Guzzo, F. Novel aspects of grape berry ripening and post-harvest withering revealed by untargeted LC-ESI-MS metabolomics analysis. Metabolomics 2011, 7, 424–436. [Google Scholar] [CrossRef]
- Khan, R.A.; Khan, M.R.; Sahreen, S.; Ahmed, M. Assessment of flavonoids contents and in vitro antioxidant activity of Launaea procumbens. Chem. Cent. J. 2012, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Cantos, E.; Espin, J.C.; Tomás-Barberán, F.A. Varietal differences among the polyphenol profiles of seven table grape cultivars studied by LC−DAD−MS−MS. J. Agric. Food Chem. 2002, 50, 5691–5696. [Google Scholar] [CrossRef]
- Ferrare, K.; Bidel, L.P.; Awwad, A.; Poucheret, P.; Cazals, G.; Lazennec, F.; Azay-Milhau, J.; Tournier, M.; Lajoix, A.-D.; Tousch, D. Increase in insulin sensitivity by the association of chicoric acid and chlorogenic acid contained in a natural chicoric acid extract (NCRAE) of chicory (Cichorium intybus L.) for an antidiabetic effect. J. Ethnopharmacol. 2018, 215, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Asif, M.; Saadullah, M.; Yaseen, H.S.; Saleem, M.; Yousaf, H.M.; Khan, I.U.; Yaseen, M.; Shams, M.U. Evaluation of in vivo anti-inflammatory and anti-angiogenic attributes of methanolic extract of Launaea spinosa. Inflammopharmacology 2020, 28, 993–1008. [Google Scholar] [CrossRef]
- Liu, X.; Yang, Q.; Lu, Y.; Li, Y.; Li, T.; Zhou, B.; Qiao, L. Effect of purslane (Portulaca oleracea L.) extract on anti-browning of fresh-cut potato slices during storage. Food Chem. 2019, 283, 445–453. [Google Scholar] [CrossRef]
- Kikowska, M.; Kruszka, D.; Derda, M.; Hadaś, E.; Thiem, B. Phytochemical screening and acanthamoebic activity of shoots from in vitro cultures and in vivo plants of Eryngium alpinum L.—The endangered and protected species. Molecules 2020, 25, 1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.-Q.; Leung, A.K.M.; Chan, C.L.; Su, T.; Li, W.-D.; Li, S.-M.; Fong, D.W.F.; Yu, Z.-L. UHPLC UHD Q-TOF MS/MS analysis of the impact of sulfur fumigation on the chemical profile of Codonopsis Radix (Dangshen). Analyst 2014, 139, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Knight, S.; Kuhnert, N. Discriminating between the six isomers of dicaffeoylquinic acid by LC-MS n. J. Agric. Food Chem. 2005, 53, 3821–3832. [Google Scholar] [CrossRef]
- Mishra, G.J.; Reddy, M.; Rana, J.S. Isolation of flavonoid constituent from Launaea procumbens Roxb. by preparative HPTLC method. IOSR J. Pharm 2012, 2, 5–11. [Google Scholar] [CrossRef]
- Moussaoui, F.; Zellagui, A.; Segueni, N.; Touil, A.; Rhouati, S. Flavonoid constituents from Algerian Launaea resedifolia (OK) and their antimicrobial activity. Rec. Nat. Prod. 2010, 4, 91–95. [Google Scholar]
- Bastos, K.X.; Dias, C.N.; Nascimento, Y.M.; Da Silva, M.S.; Langassner, S.M.Z.; Wessjohann, L.A.; Tavares, J.F. Identification of phenolic compounds from Hancornia speciosa (Apocynaceae) leaves by uhplc orbitrap-hrms. Molecules 2017, 22, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farag, M.A.; El-Ahmady, S.H.; Elian, F.S.; Wessjohann, L.A. Metabolomics driven analysis of artichoke leaf and its commercial products via UHPLC–q-TOF-MS and chemometrics. Phytochemistry 2013, 95, 177–187. [Google Scholar] [CrossRef]
- Fuchs, D.; Hamberg, M.; Sköld, C.M.; Wheelock, Å.M.; Wheelock, C.E. An LC-MS/MS workflow to characterize 16 regio-and stereoisomeric trihydroxyoctadecenoic acids. J. Lipid Res. 2018, 59, 2025–2033. [Google Scholar] [CrossRef] [Green Version]
- Dugo, P.; Mondello, L.; Dugo, L.; Stancanelli, R.; Dugo, G. LC-MS for the identification of oxygen heterocyclic compounds in citrus essential oils. J. Pharm. Biomed. Anal. 2000, 24, 147–154. [Google Scholar] [CrossRef]
- Farag, M.A.; Shakour, Z.T.A. Metabolomics driven analysis of 11 Portulaca leaf taxa as analysed via UPLC-ESI-MS/MS and chemometrics. Phytochemistry 2019, 161, 117–129. [Google Scholar] [CrossRef]
- Khaled, S.E.; Hashem, F.A.-M.; Shabana, M.H.; Hammam, A.-M.M.; Madboli, A.N.A.; Al-Mahdy, D.A.; Farag, M.A. A biochemometric approach for the assessment of Phyllanthus emblica female fertility effects as determined via UPLC-ESI-qTOF-MS and GC-MS. Food Funct. 2019, 10, 4620–4635. [Google Scholar] [CrossRef]
- Riaz, N.; Parveen, S.; Saleem, M.; Ali, M.S.; Malik, A.; Ashraf, M.; Afzal, I.; Jabbar, A. Lipoxygenase inhibitory sphingolipids from Launaea nudicaulis. J. Asian Nat. Prod. Res. 2012, 14, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Davis, R.A.; Sykes, M.L.; Avery, V.M.; Quinn, R.J. Iotrochamides A and B, antitrypanosomal compounds from the Australian marine sponge Iotrochota sp. Bioorganic Med. Chem. Lett. 2012, 22, 4873–4876. [Google Scholar] [CrossRef] [Green Version]
- Panche, A.; Diwan, A.; Chandra, S. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, R.; Clerc, A.-C.; Gori, I.; Russell, R.; Pellegrini, C.; Govender, L.; Wyss, J.-C.; Golshayan, D.; Canny, G.O. Lipoxin A 4 prevents the progression of de novo and established endometriosis in a mouse model by attenuating prostaglandin E 2 production and estrogen signaling. PLoS ONE 2014, 9, e89742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snoch, W.; Stępień, K.; Prajsnar, J.; Staroń, J.; Szaleniec, M.; Guzik, M. Influence of chemical modifications of polyhydroxyalkanoate-derived fatty acids on their antimicrobial properties. Catalysts 2019, 9, 510. [Google Scholar] [CrossRef] [Green Version]
- Mullen, T.; Obeid, L.M. Ceramide and apoptosis: Exploring the enigmatic connections between sphingolipid metabolism and programmed cell death. Anticancer Agents Med. Chem. 2012, 12, 340–363. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Guo, Y.-W.; Gu, Y. Secondary metabolites from the South China Sea invertebrates: Chemistry and biological activity. Curr. Med. Chem. 2006, 13, 2041–2090. [Google Scholar] [CrossRef]
- Belitz, H.-D.; Grosch, W.; Schieberle, P. Amino Acids, Peptides, Proteins. In Food Chemistry; Springer: Berlin/Heidelberg, Germany, 2004; pp. 8–91. [Google Scholar]
- Gbadmosi, I.; Adeyi, O.; Oyekanmi, O.; Somade, O. Launea taraxacifolia leaf partitions ameliorates alloxan-induced pathophysiological complications via antioxidant mechanisms in diabetic rats. Metab. Open 2020, 6, 100029. [Google Scholar] [CrossRef]
- Adeyi, A.O.; Nneji, L. Ameliorative potentials of medicinal plants on the pathophysiological complications of diabetes mellitus: A review. J. Med. Plants Res. 2015, 9, 262–288. [Google Scholar] [CrossRef]
- Al-Doaiss, A.A.; Al-Shehri, M.A. Efecto Protector de la Goma Arábiga/Insulina Contra los Cambios Histológicos en los Testículos de Ratas Diabéticas. Int. J. Morphol. 2020, 38, 340–347. [Google Scholar] [CrossRef] [Green Version]
- Navarro-Casado, L.; Juncos-Tobarra, M.; Chafer-Rudilla, M.; De Onzono, L.Í.; Blazquez-Cabrera, J.; Miralles-Garcia, J. Effect of experimental diabetes and STZ on male fertility capacity. Study in rats. J. Androl. 2010, 31, 584–592. [Google Scholar] [CrossRef] [PubMed]
- Kianifard, D.; Sadrkhanlou, R.A.; Hasanzadeh, S. The ultrastructural changes of the sertoli and leydig cells following streptozotocin induced diabetes. Iran. J. Basic Med. Sci. 2012, 15, 623–635. [Google Scholar] [PubMed]
- Du, Z.; Xu, S.; Hu, S.; Yang, H.; Zhou, Z.; Sidhu, K.; Miao, Y.; Liu, Z.; Shen, W.; Reiter, R.J. Melatonin attenuates detrimental effects of diabetes on the niche of mouse spermatogonial stem cells by maintaining Leydig cells. Cell Death Dis. 2018, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Adejuwon, S.; Femi-Akinlosotu, O.; Omirinde, J. Cisplatin-induced testicular dysfunction and its amelioration by Launaea taraxacifolia leaf extract. Andrologia 2015, 47, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.A. Protective effects of Launaea procumbens on rat testis damage by CCl4. Lipids Health Dis. 2012, 11, 103. [Google Scholar] [CrossRef] [Green Version]
- Adinortey, M.B.; Agbeko, R.; Boison, D.; Ekloh, W.; Kuatsienu, L.E.; Biney, E.E.; Affum, O.O.; Kwarteng, J.; Nyarko, A.K. Phytomedicines used for diabetes mellitus in Ghana: A systematic search and review of preclinical and clinical evidence. Evid.-Based Complementary Altern. Med. 2019, 2019, 6021209. [Google Scholar] [CrossRef] [Green Version]
- AL-Ishaq, R.K.; Abotaleb, M.; Kubatka, P.; Kajo, K.; Büsselberg, D. Flavonoids and their anti-diabetic effects: Cellular mechanisms and effects to improve blood sugar levels. Biomolecules 2019, 9, 430. [Google Scholar] [CrossRef] [Green Version]
- Tangvarasittichai, S. Oxidative stress, insulin resistance, dyslipidemia and type 2 diabetes mellitus. World J. Diabetes 2015, 6, 456. [Google Scholar] [CrossRef]
- Halban, P.A.; Polonsky, K.S.; Bowden, D.W.; Hawkins, M.A.; Ling, C.; Mather, K.J.; Powers, A.C.; Rhodes, C.J.; Sussel, L.; Weir, G.C. β-cell failure in type 2 diabetes: Postulated mechanisms and prospects for prevention and treatment. J. Clin. Endocrinol. Metab. 2014, 99, 1983–1992. [Google Scholar] [CrossRef] [PubMed]
- Erejuwa, O.O.; Sulaiman, S.A.; Wahab, M.S.A.; Sirajudeen, K.N.S.; Salleh, M.S.M.; Gurtu, S. Antioxidant protective effect of glibenclamide and metformin in combination with honey in pancreas of streptozotocin-induced diabetic rats. Int. J. Mol. Sci. 2010, 11, 2056–2066. [Google Scholar] [CrossRef] [Green Version]
- Khan, R.A.; Khan, M.R.; Sahreen, S.; Ahmed, M.; Shah, N.A. Carbon tetrachloride-induced lipid peroxidation and hyperglycemia in rat: A novel study. Toxicol. Ind. Health 2015, 31, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Alqasoumi, S.I.; Abdel-Kader, M.S. Terpenoids from Juniperus procera with hepatoprotective activity. Pak. J. Pharm. Sci. 2012, 25, 315–322. [Google Scholar] [PubMed]
- El-Newary, S.A.; Ismail, R.F.; Shaffie, N.M.; Hendawy, S.; Omer, E. Hepatoprotective, therapeutic and in vivo anti-oxidant activities of Tagetes lucida leaves alcoholic extract against paracetamol-induced hepatotoxicity rats. Int. J. PharmTech Res. 2016, 9, 327–341. [Google Scholar]
- Abd-ElGawad, A.M.; Elshamy, A.; El-Amier, Y.A.; El Gendy, A.; Al-Barati, S.; Dar, B.; Al-Rowaily, S.; Assaeed, A. Chemical composition variations, allelopathic, and antioxidant activities of Symphyotrichum squamatum (Spreng.) Nesom essential oils growing in heterogeneous habitats. Arab. J. Chem. 2020, 13, 237–4245. [Google Scholar] [CrossRef]
- Sun, C.; Zhao, C.; Guven, E.C.; Paoli, P.; Simal-Gandara, J.; Ramkumar, K.M.; Wang, S.; Buleu, F.; Pah, A.; Turi, V. Dietary polyphenols as antidiabetic agents: Advances and opportunities. Food Front. 2020, 1, 18–44. [Google Scholar] [CrossRef] [Green Version]
- Vinayagam, R.; Jayachandran, M.; Xu, B. Antidiabetic effects of simple phenolic acids: A comprehensive review. Phytother. Res. 2016, 30, 184–199. [Google Scholar] [CrossRef]
- Zhang, Z.; Ding, Y.; Dai, X.; Wang, J.; Li, Y. Epigallocatechin-3-gallate protects pro-inflammatory cytokine induced injuries in insulin-producing cells through the mitochondrial pathway. Eur. J. Pharmacol. 2011, 670, 311–316. [Google Scholar] [CrossRef]
- Zanatta, L.; Rosso, A.; Folador, P.; Figueiredo, M.S.; Pizzolatti, M.G.; Leite, L.D.; Silva, F.R. Insulinomimetic effect of kaempferol 3-neohesperidoside on the rat soleus muscle. J. Nat. Prod. 2008, 71, 532–535. [Google Scholar] [CrossRef]
- Nguyen-Ngo, C.; Salomon, C.; Quak, S.; Lai, A.; Willcox, J.C.; Lappas, M. Nobiletin exerts anti-diabetic and anti-inflammatory effects in an in vitro human model and in vivo murine model of gestational diabetes. Clin. Sci. 2020, 134, 571–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundaram, R.; Shanthi, P.; Sachdanandam, P. Effect of tangeretin, a polymethoxylated flavone on glucose metabolism in streptozotocin-induced diabetic rats. Phytomedicine 2014, 21, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Galadari, S.; Rahman, A.; Pallichankandy, S.; Galadari, A.; Thayyullathil, F. Role of ceramide in diabetes mellitus: Evidence and mechanisms. Lipids Health Dis. 2013, 12, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulos, L. Flora of Egypt: Verbenaceae—Compositae; Al Hadara Publishing: Cairo, Egypt, 2002; Volume 3. [Google Scholar]
- Tackholm, V. Students Flora of Egypt, 2nd ed.; Cairo University: Cairo, Egypt, 1974. [Google Scholar]
- AOAC. Official Methods of Analysis; Association of Official Analytical Chemists: Washington, DC, USA, 2000. [Google Scholar]
- OECD. OECD Guidelines for the Testing of Chemicals No. 423: Acute Oral Toxicity—Acute Toxic Class Method; OECD: Paris, France, 1996. [Google Scholar]
- Ravi, K.; Rajasekaran, S.; Subramanian, S. Antihyperglycemic effect of Eugenia jambolana seeds kernel on streptozotocin-induced diabetes in rats. Food Chem. Toxicol. 2005, 43, 1433–1439. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, M. Fundamentals of experimental pharmacology. Indian J. Pharmacol. 2007, 39, 216. [Google Scholar]
- Garg, R.; Kumar, R.; Nathiya, D.; Goshain, O.; Trivedi, V.; Sharma, A.K.; Murti, K. Comparative acute toxicity studies of selected indigenous herbal plants in Swiss albino mice. IOSR J. Pharm. Biol. Sci. 2016, 11, 20–27. [Google Scholar]
- Ekeanyanwu, R.; Udeme, A.; Onuigbo, A.; Etienajirhevwe, O. Anti-diabetic effect of ethanol leaf extract of Cissampelos owariensis (lungwort) on alloxan induced diabetic rats. Afr. J. Biotechnol. 2012, 11, 6758–6762. [Google Scholar] [CrossRef]
- Van Pelt, L. Ketamine and xylazine for surgical anesthesia in rats. J. Am. Vet. Med. Assoc. 1977, 171, 842–844. [Google Scholar]
- Allain, C.C.; Poon, L.S.; Chan, C.S.; Richmond, W.; Fu, P.C. Enzymatic determination of total serum cholesterol. Clin. Chem. 1974, 20, 470–475. [Google Scholar] [CrossRef]
- Naito, H.K. High-density lipoprotein (HDL) cholesterol. Methods Clin. Chem. 1984, 437, 1207–1213. [Google Scholar]
- Fossati, P.; Prencipe, L. Serum triglycerides determined colorimetrically with an enzyme that produces hydrogen peroxide. Clin. Chem. 1982, 28, 2077–2080. [Google Scholar] [CrossRef]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi-Hayakawa, H.; Onodera, N.; Matsubara, S.; Yasuda, E.; Chonan, O.; Takahashi, R.; Ishikawa, F. Effects of soy milk and bifidobacterium fermented soy milk on lipid metabolism in aged ovariectomized rats. Biosci. Biotechnol. Biochem. 1998, 62, 1688–1692. [Google Scholar] [CrossRef] [Green Version]
- Henry, R. Clinical Chemistry. Principles and Techniques; Harper & Row, Publishers: New York, NY, USA, 1964. [Google Scholar]
- Doumas, B.T.; Watson, W.A.; Biggs, H.G. Albumin standards and the measurement of serum albumin with bromcresol green. Clin. Chim. Acta 1971, 31, 87–96. [Google Scholar] [CrossRef]
- Belfield, A.; Goldberg, D.M. Application of a continuous spectrophotometric assay for 5’nucleotidase activity in normal subjects and patients with liver and bone disease. Clin. Chem. 1969, 15, 931–939. [Google Scholar] [CrossRef] [PubMed]
- Rettman, S.; Frankel, L.S. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol. 1957, 28, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Reinhold, J.G. Standard Methods in Clinical Chemistry; Academic Press: New York, NY, USA, 1953. [Google Scholar]
- Tabacco, A.; Meiattini, F.; Moda, E.; Tarli, P. Simplified enzymic/colorimetric serum urea nitrogen determination. Clin. Chem. 1979, 25, 336–337. [Google Scholar] [CrossRef] [PubMed]
- Gochman, N.; Schmitz, J.M. Automated determination of uric acid, with use of a uricase—Peroxidase system. Clin. Chem. 1971, 17, 1154–1159. [Google Scholar] [CrossRef]
- Faulkner, W.; King, J. Colorimetric detemination of creatinine. In Fundamentals of Clinical Chemistry, 2nd ed.; Tietz, N.W., Ed.; Saunders: Philadelphia, PA, USA, 1976; Volume 994. [Google Scholar]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Griffith, O.W. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine. Anal. Biochem. 1980, 106, 207–212. [Google Scholar] [CrossRef]
- Goldberg, D.M.; Spooner, R.J. Glutathione reductase. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Bergmeyer, J., GraBI, M., Eds.; Verlag Chemie: Basel, Switzerland, 1983; Volume 3, pp. 258–265. [Google Scholar]
- Paglia, D.E.; Valentine, W.N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases the first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Beers, R.F.; Sizer, I.W. A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase. J. Biol. Chem. 1952, 195, 133–140. [Google Scholar] [CrossRef]
- Suvarna, K.S.; Layton, C.; Bancroft, J.D. Bancroft’s Theory and Practice of Histological Techniques, 8th ed.; Elsevier: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Sakna, S.T.; Mocan, A.; Sultani, H.N.; El-Fiky, N.M.; Wessjohann, L.A.; Farag, M.A. Metabolites profiling of Ziziphus leaf taxa via UHPLC/PDA/ESI-MS in relation to their biological activities. Food Chem. 2019, 293, 233–246. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Blood Glucose Level (mg/dL) | |||||
|---|---|---|---|---|---|---|
| Start Time | 1st Week | 2nd Week | 3rd Week | 4th Week | 5th Week | |
| Negative control | 124.0 ± 6.12 o | 126.6 ± 7.77 o | 127.6 ± 8.32 o | 123.8 ± 7.92 o | 125.8 ± 7.46 o | 126.2 ± 5.63 o |
| 0.2% * | 2.9% | −0.2% | 1.5% | 1.8% | ||
| Diabetic control | 411.9 ± 17.53 d | 434.4 ± 15.19 c | 467.2 ± 15.56 b | 485.9 ± 17.17 a | 486.3 ± 20.07 a | 486.00 ± 25.22 a |
| 5.5% | 13.42% | 18.0% | 18.0% | 18.0% | ||
| L. nudicaulis (250 mg/kg/day) | 408.5 ± 8.86 d | 302.5 ± 17.23h h | 254.1 ± 11.48 k | 225.9 ± 11.47 l | 217.8 ± 10.28 l | 188.4 ± 7.71 m |
| −26.0% | −37.8% | −44.7% | −46.70% | −53.9% | ||
| L. nudicaulis (500 mg/kg/day) | 410.0 ± 9.13 d | 364.4 ± 11.97 f | 263.6 ± 14.67 j | 194.6 ± 9.41 m | 161.1 ± 4.85 n | 130.9 ± 5.61 o |
| −11.1% | −35.7% | −52.5% | −60.7% | −68.1% | ||
| Glibenclamide (5 mg/kg/day) | 408.7 ± 5.71 d | 380.7 ± 17.38 e | 354.7 ± 10.72 g | 289.9 ± 14.16 i | 244.7 ± 9.76 k | 198.4 ± 7.87 m |
| −6.9% | −13.2% | −29.1% | −40.1% | −51.5% | ||
| Time | Start Time | 1-Week | 2-Weeek | 3-Week | 4-Week | 5-Week |
|---|---|---|---|---|---|---|
| Negative control | 167.0 ± 4.97 g,h | 180.4 ± 7.12 f | 190.0 ± 5.30 d | 197.8 ± 3.71 c | 212.4 ± 5.15 b | 227.0 ± 3.92 a |
| +13.4 * | +9.6 | +7.8 | +14.6 | +14.6 | ||
| Diabetic Control | 166.1 ± 4.36 g,h | 153.8 ± 3.69 j,k | 138.8 ± 5.80 m,n | 133.4 ± 3.61 n | 123.4 ± 1.80 o | 107.5 ± 1.67 p |
| −12.3 | −15.0 | −5.4 | −10.0 | −15.5 | ||
| L. nudicaulis 250 mg/kg | 150.0 ± 13.30 k,l | 134.1 ± 1.51 n | 147.0 ± 4.97 l | 161.2 ± 9.66 h,i | 167.0 ± 7.42 g | 196.2 ± 11.22 c |
| −15.9 | +12.9 | +14.2 | +5.8 | +29.2 | ||
| L. nudicaulis 500 mg/kg | 161.0 ± 11.81 g,h,i | 150.1 ± 8.28 k,l | 156.9 ± 6.20 i,j | 165.7 ± 6.37 g,h | 183.9 ± 7.45 e,f | 210.2 ± 9.12 b |
| −10.9 | +6.8 | +8.9 | +18.2 | +26.4 | ||
| gilbenclamide 5 mg/kg/day | 162.0 ± 7.47 g,h,i | 140.0 ± 6.73 m | 153.0 ± 10.21 j,k | 167.0 ± 3.08 g,h | 181.3 ± 6.39 f | 189.8 ± 3.40 d,e |
| −22.0 | +13.0 | +14.0 | +14.3 | +8.5 |
| Groups | TC (mg/dL) | HDL-C (mg/dL) | TG (mg/dL) | VLDL-C (mg/dL) | LDL-C (mg/dL) | Risk Ratio % |
|---|---|---|---|---|---|---|
| Negative control | 82.30 ± 7.33 c | 49.10 ± 2.91 a | 142.87 ± 2.85 c | 28.57 ± 0.57 c | 5.09 ± 0.94 d | 0.11 ± 0.01 d |
| Diabetic control | 194.86 ± 10.65 a | 34.45 ± 1.40 c | 232.90 ± 8.43 a | 46.58 ± 1.69 a | 113.83 ± 6.75 a | 3.33 ± 0.25 a |
| L. nudicaulis 250 mg/kg/day | 88.29 ± 6.88 b | 44.96 ± 1.61 b | 148.26 ± 7.72 b | 29.65 ± 1.54 b | 13.68 ± 0.94 c | 0.31 ± 0.03 c |
| L. nudicaulis 500 mg/kg/day | 92.28 ± 5.25 b | 44.20 ± 2.46 b | 136.08 ± 7.35 d | 27.22 ± 1.47 d | 20.88 ± 1.43 b | 0.47 ± 0.04 b |
| Glibenclamide 5 mg/kg/day | 94.54 ± 5.00 b | 46.05 ± 3.12 b | 150.18 ± 4.81 b | 30.05 ± 0.96 b | 18.60 ± 1.90 b | 0.41 ± 0.03 b |
| Groups | Total Protein g/dL | Albumin g/dL | Globulin g/dL | AST U/L | ALT U/L | ALP U/L |
|---|---|---|---|---|---|---|
| Negative control | 7.39 ± 0.44 a | 4.08 ± 0.34 c | 3.31 ± 0.64 a | 57.35 ± 4.47 c | 31.08 ± 3.77 c | 64.84 ± 36.80 c |
| Diabetic control | 4.81 ± 0.26 b | 2.91 ± 0.13 d | 1.89 ± 0.32 c | 119.56 ± 8.87 a | 84.41 ± 6.16 a | 95.84 ± 3.40 a |
| L. nudicaulis 250 mg/kg/day | 7.15 ± 0.65 a | 5.21 ± 0.23 a | 1.94 ± 0.66 c | 86.23 ± 3.89 b | 42.60 ± 3.40 b | 75.20 ± 0.79 b |
| L. nudicaulis 500 mg/kg/day | 7.22 ± 0.54 a | 4.66 ± 0.40 b | 2.57 ± 0.83 b | 82.19 ± 6.36 b | 36.21 ± 3.11 b | 71.49 ± 0.44 b |
| Glibenclamide 5 mg/kg/day | 7.19 ± 0.55 a | 4.81 ± 0.28 b | 2.39 ± 0.34 b,c | 91.99 ± 5.20 b | 44.48 ± 2.45 b | 71.35 ± 0.76 b |
| Groups | Creatinine mg/dL | Uric Acid mg/dL | Urea mg/dL |
|---|---|---|---|
| Negative control | 1.21 ± 0.02 d | 4.11 ± 0.08 d | 4.32 ± 0.05 e |
| Diabetic control | 2.42 ± 0.08 a | 11.91 ± 0.30 a | 8.29 ± 0.09 a |
| L. nudicaulis 250 mg/kg/day | 1.71 ± 0.08 b | 7.10 ± 0.20 b | 5.42 ± 0.12 c |
| L. nudicaulis 500 mg/kg/day | 1.64 ± 0.05 b | 5.10 ± 0.14 c | 4.91 ± 0.10 d |
| Glibenclamide 5 mg/kg/day | 1.41 ± 0.04 c | 6.89 ± 0.14 b | 6.55 ± 0.17 b |
| Groups | Liver | Kidney | Spleen | Brain | Pancreas | Heart | Lung | Testes |
|---|---|---|---|---|---|---|---|---|
| Negative control | 4.14 ± 0.26 c,d | 0.84 ± 0.05 c | 0.67 ± 0.07 b | 0.80 ± 0.10 b,c | 0.37 ± 0.07 b | 0.37 ± 0.04 b | 0.74 ± 0.10 b,c | 1.31 ± 0.15 c |
| Diabetic control | 6.23 ± 0.16 a | 1.13 ± 0.04 a | 0.75 ± 0.05 a | 1.09 ± 0.08 a | 0.26 ± 0.08 c | 0.45 ± 0.09 a | 1.00 ± 0.22 a | 1.95 ± 0.28 a |
| L. nudicaulis 250 mg/kg/day | 4.84 ± 0.14 b | 0.97 ± 0.19 b | 0.57 ± 0.13 c | 0.8 ± 0.29 b,c | 0.47 ± 0.04 a | 0.38 ± 0.06 b | 0.85 ± 0.23 b | 1.60 ± 0.04 b |
| L. nudicaulis 500 mg/kg/day | 4.29 ± 0.37 c | 0.79 ± 0.05 c | 0.52 ± 0.11 c | 0.76 ± 0.02 c | 0.36 ± 0.06 b | 0.36 ± 0.04 b | 0.70 ± 0.03 c | 1.29 ± 0.03 c |
| Glibenclamide 5 mg/kg/day | 4.06 ± 0.26 d | 1.01 ± 0.10 b | 0.57 ± 0.09 c | 0.93 ± 0.13 b | 0.37 ± 0.04 b | 0.45 ± 0.08 a | 0.78 ± 0.04 b,c | 1.47 ± 0.24 b |
| Groups | MDA nmol/mL | CAT (U/L) | GSH (mg/dL) | GR (µmol/mg protein/min) | GST (µmol/mg protein/min) | GPx (µmol/mg protein/min) |
|---|---|---|---|---|---|---|
| Negative control | 3.54 ± 0.31 e | 118.82 ± 7.77 a | 4.70 ± 0.24 c | 5.80 ± 0.29 c | 3.91 ± 0.20 d | 2.40 ± 0.12 b |
| Diabetic control | 10.414 ± 0.23 a | 86.96 ± 9.26 d | 1.76 ± 0.20 d | 1.86 ± 0.21 d | 1.32 ± 0.15 e | 0.80 ± 0.09 c |
| L. nudicaulis 250 mg/kg/day | 6.43 ± 0.33 b | 108.89 ± 4.49 b,c | 4.682 ± 0.09 c | 5.92 ± 0.12 c | 4.11 ± 0.08 c | 2.26 ± 0.06 b |
| L. nudicaulis 500 mg/kg/day | 4.46 ± 0.31 d | 113.46 ± 5.47 a,b | 5.22 ± 0.14 a | 6.89 ± 0.18 a | 4.34 ± 0.20 b | 2.78 ± 0.07 a |
| Glibenclamide 5 mg/kg/day | 5.02 ± 0.36 c | 103.20 ± 6.04 c | 4.94 ± 0.29 b | 6.37 ± 0.37 b | 5.40 ± 0.31 a | 2.67 ± 0.14 a |
| No. | RT | Formula | Name | Class | Precursor | Fragmentation | Error | Ref. |
|---|---|---|---|---|---|---|---|---|
| 1 | 1.088 | C5H9O5− | Pentose | Carbohydrate | 149.0453 | 101, 89, 75, 73 | 1.85 | |
| 2 | 1.179 | C7H11O6− | Quinic acid | Organic acid | 191.0574 | 127, 105, 85, 75 | −7.12 | [22] |
| 3 | 1.225 | C7H14NO2+ | Unknwon | Nitrogenous compound | 144.1025 | 98, 84, 70 | 2.47 | |
| 4 | 1.271 | C7H8NO2+ | Aminobenzoic acid | Nitrogenous compound | 138.0546 | 92, 78, 67 | 2.39 | [23] |
| 5 | 1.271 | C4H5O5− | Malic acid | Organic acid | 133.0156 | 115, 75, 73, 71 | −9.73 | [24] |
| 6 | 1.476 | C10H7O4− | Scopoletin | Coumarin | 191.0368 | - | 9.51 | [25] |
| 7 | 1.636 | C11H16NO8− | Deoxy-dehydro-N-acetylneuraminic acid | Organic acid | 290.0898 | 200, 128 | −5.61 | [26] |
| 8 | 1.865 | C12H19N2O7− | Unknown | Pyrazine | 303.1216 | 213, 123, 87 | −7.28 | [27] |
| 9 | 2.642 | C7H10NO+ | Hydroxybenzylamine | Nitrogenous compound | 124.0753 | 108, 94 | 5.8 | [28] |
| 10 | 2.733 | C6H14NO2+ | Leucine | Amino acid | 132.1029 | 86, 69 | −6.85 | [29] |
| 11 | 2.779 | C17H19O4+ | Unknown | Phenolic acid | 287.127 | 229, 193, 175 | −2.73 | |
| 12 | 3.098 | C9H12NO+ | Dimethylamino-benzaldehyde | Nitrogenous compound | 150.0904 | 135, 108 | 7.83 | |
| 13 | 3.19 | C6H10NO3− | N-Acetylglycine ethyl ester | Amino acid | 144.067 | 102, 87, 71 | 0.99 | |
| 14 | 3.556 | C5H6N5O+ | Guanine | Purine | 152.0576 | 135, 110, 94 | −1.25 | [30] |
| 15 | 3.647 | C19H23O6+ | Trihydroxy-dimethoxy-dimethylflavan | Flavonoid | 347.1489 | 332, 329, 284, 227, 209 | −0.04 | |
| 16 | 3.693 | C10H12N5O5− | Guanosine | Purine | 282.0862 | 150, 133 | 6.4 | [30] |
| 17 | 4.195 | C7H7O+ | Benzaldehyde | Aldehyde | 107.0492 | 79, 77 | 3.12 | |
| 18 | 5.064 | C10H13N2O5− | Thymidine | Pyrimidine | 241.0838 | 208, 180, 150, 131 | 1.01 | [30] |
| 19 | 5.338 | C16H19O3+ | Methylbatatasin III | Phenolic | 259.1326 | 234, 201, 160, 136 | −1.04 | [31] |
| 20 | 5.338 | C10H11N4O6− | Xanthosine | Purine | 283.0712 | 197, 151, 66 | 9.86 | [32] |
| 21 | 5.566 | C19H23O6+ | Trihydroxy-dimethoxy-dimethylflavan isomer | Flavonoid | 347.1521 | 332, 330, 318, 287, 251, 168 | 9.17 | |
| 22 | 6.343 | C8H12N+ | Phenylethylamine | Nitrogenous compound | 122.0965 | 105 | 1.45 | |
| 23 | 6.846 | C15H17O9− | O-caffeoylhexose | Phenolic acid | 341.0912 | 179, 161 | −3.31 | [33] |
| 24 | 6.891 | C12H21O4+ | Hydroxy-dihydrojasmonic acid | Organic acid | 229.1443 | 170, 70 | 3.77 | |
| 25 | 8.034 | C10H13N2O4− | Unknwon nitrogenous | Nitrogenous | 225.0891 | 210, 181, 165, 139 | −5.16 | |
| 26 | 8.125 | C10H12N5O− | Zeatin | Purine | 218.1048 | 204, 177, 146, 137 | 0.44 | [34] |
| 27 | 8.216 | C17H19O3+ | Hydroxy-methoxy-methylflavan | Flavonoid | 271.1311 | 256, 228, 193, 177, 147, 137, 121 | −6.53 | |
| 28 | 8.262 | C18H19O2+ | Phenylpropyl cinnamate | Ester | 267.1374 | 189, 147, 120, 86 | −2.08 | |
| 29 | 8.308 | C14H15O+ | Phenethylphenol | Phenolic | 199.1099 | 135, 109 | 2.88 | |
| 30 | 8.354 | C9H11O3+ | Methoxyphenylacetic acid | Phenolic | 167.0702 | 123, 109, 78 | −0.42 | [35] |
| 31 | 8.765 | C13H9O9− | Caffeoyl-O-dihydroxymaleic acid | Phenolic acid | 309.0283 | 179, 135, 133 | −9.93 | |
| 32 | 8.811 | C16H17O9− | O-Caffeoylquinic acid | Phenolic acid | 353.0911 | 191, 179 | 9.32 | [15] |
| 33 | 8.856 | C13H11O9− | Caffeoyl tartaric acid | Phenolic acid | 311.0438 | 179, 149, 135 | −9.73 | [36] |
| 34 | 8.902 | C7H5O5− | Gallic acid | Phenolic acid | 169.0161 | 151, 125, 107, 95 | −9.24 | [37] |
| 35 | 8.993 | C9H8NO+ | Unknwon | Aldehyde | 146.0608 | 118, 91, 77 | −3.73 | |
| 36 | 9.176 | C20H19O2+ | Unknown | - | 291.1363 | 205, 190, 141, 128 | −0.26 | |
| 37 | 9.359 | C13H11O8− | p-coumaroyltartaric acid | Phenolic acid | 295.0488 | 251, 163, 119, 87 | 9.69 | [38] |
| 38 | 9.404 | C10H10N+ | Unknown | Nitrogenous compound | 144.0811 | 127, 116, 103, 91 | −2.61 | |
| 39 | 9.633 | C14H13O9− | Feruloyl-O-tartaric acid | Phenolic acid | 325.0599 | 193, 134 | −9.13 | [39] |
| 40 | 9.953 | C12H17O4− | Hydroxyjasmonic acid | Organic acid | 225.1157 | 165, 137, 97,81 | −9.87 | |
| 41 | 9.999 | C9H5O4− | Caffeic acid quinone | Phenolic acid | 177.0209 | 162, 149, 135,121 | −8.28 | |
| 42 | 10.09 | C12H12NO5− | p-Coumaroyl-serine | Phenolic acid | 250.0745 | 207, 161, 132, 115 | −9.12 | |
| 43 | 10.227 | C8H7N4O− | Unknown | Nitrogenous compound | 175.0631 | 133, 105, 89, 77 | −3.26 | |
| 44 | 10.273 | C9H7O4− | Caffeic acid | Phenolic acid | 179.0366 | 135, 124, 107, 93 | 5.95 | [40] |
| 45 | 10.501 | C11H12NO3− | N-Acetyl-l-phenylalanine | Amino acid | 206.0835 | 164, 147, 103, 91 | −6 | [41] |
| 46 | 10.867 | C10H17N2O3S− | Prolyl-methionine | Amino acid | 245.0961 | 203, 186, 159, 142 | 0.27 | |
| 47 | 10.913 | C24H35N8OS4− | Unknown | - | 579.1791 | 254, 203, 116,72 | 0.75 | |
| 48 | 11.232 | C25H23O12− | Scopoletin-7-O-dihexoside | Coumarin | 515.1237 | 353, 191, 179, 173 | 8.15 | [42] |
| 49 | 11.552 | C9H15O4− | Nonanedioic acid | Organic acid | 187.0995 | 170, 125, 97, 87 | 7.29 | [43] |
| 50 | 11.849 | C10H11O4+ | Methyl caffeate | Phenolic acid | 195.0657 | 177, 163, 150, 145 | −2.98 | [44] |
| 51 | 12.009 | C20H31O6+ | Unknown | - | 367.2151 | 349, 325, 204, 112 | 9.76 | |
| 52 | 12.329 | C15H11O6+ | Kaempferol | Flavonoid | 287.0567 | 153, 135, 93 | 5.87 | [45] |
| 53 | 12.512 | C11 H17O3+ | Hydroxybutyl-methoxy-phenol | Phenolic | 197.1188 | 180, 166, 153, 141 | −9.75 | |
| 54 | 12.74 | C11H11O4− | Unknown | Phenolic acid | 207.068 | 179,161,135 | 8.29 | |
| 55 | 12.786 | C13H23O2+ | Tridecadienoic acid | Fatty acid | 211.1694 | 151, 135, 83, 67 | −0.79 | |
| 56 | 13.06 | C15H11O5+ | Apigenin | Flavonoid | 271.0627 | 153, 131, 95 | 9.59 | [46] |
| 57 | 13.106 | C18H35O5+ | Trihydroxy-octadecenoic acid | Fatty acid | 331.2476 | 288, 244, 175, 69 | −0.9 | [47] |
| 58 | 13.151 | C18H33O5− | Trihydroxyoctadecenoic acid isomer | Fatty acid | 329.2364 | 311, 201,171 | −9.19 | [47] |
| 59 | 13.288 | C13H21O2+ | Tridecatrienoic acid | Fatty acid | 209.1542 | 163, 93, 79,71 | −3.85 | |
| 60 | 13.334 | C15H23O2+ | Unknown terpene | Terpene | 235.1712 | 219, 161, 121, 93 | −5.11 | |
| 61 | 13.38 | C15H23O+ | Unknown terpene | Terpene | 219.175 | 204, 175, 121, 79 | −1.84 | |
| 62 | 13.562 | C20H33O5+ | Trihydroxyicosa-tetraenoic acid (lipoxin) | Fatty acid | 353.2317 | 308, 277, 222, 199 | −1.55 | |
| 63 | 13.7 | C18H29O4+ | Hydroxy-oxo-octadecatrienoic acid | Fatty acid | 309.207 | 292, 273, 219, 165 | 1.31 | [48] |
| 64 | 13.837 | C18H27O3+ | Unknown | Terpene | 291.1942 | 204, 177, 133, 119 | 5.55 | |
| 65 | 13.882 | C16H29O2+ | Hexadecadienoic acid | Fatty acid | 253.2169 | 237, 209, 193, 174 | −1.34 | [49] |
| 66 | 14.157 | C20H31O5+ | Dihydroxy-oxoicosa-tetraenoic acid | Fatty acid | 351.2159 | 334, 149, 106, 81 | −1.99 | |
| 67 | 14.385 | C45H83O16P2− | Phosphatidylinositol phosphate (18:0/18:2) | Acyl glycerol | 941.5213 | 880, 471, 394, 259 | 5.43 | |
| 68 | 14.431 | C15H13O3+ | Unknown | Terpene | 241.0856 | 227, 211, 180, 157 | −1.33 | |
| 69 | 14.705 | C18H27O2+ | Unknown | Terpene | 275.2024 | 171, 147, 119,79 | −6.62 | |
| 70 | 14.751 | C20H29O4+ | Unknown | - | 333.2056 | 307, 188, 135, 87 | −1.3 | |
| 71 | 14.842 | C21H23O8+ | Hexamethoxyflavone | Flavonoid | 403.1394 | 388, 373, 212, 183 | 1.62 | [50] |
| 72 | 14.933 | C17H27O5− | Glycerol-hydroxy-tetradecatrienoate | Acyl glycerol | 311.1895 | 267, 179, 135 | 9.96 | |
| 73 | 14.979 | C18H27O3+ | Oxo-octadecatetraenoic acid | Fatty acid | 291.1965 | 209, 157, 133, 121 | −0.64 | |
| 74 | 15.208 | C18H31O4− | Dihydroxy-octadecadienoic acid | Fatty acid | 311.2245 | 267, 179, 135, 87 | −6.15 | [47] |
| 75 | 15.436 | C20H21O7+ | Pentamethoxyflavone | Flavonoid | 373.1281 | 358, 343, 312, 266, 197 | −0.21 | [50] |
| 76 | 15.619 | C18H40NO3+ | Amino-octadecanetriol | Sphingolipid | 318.2982 | 303, 242, 150, 62 | −6.5 | |
| 77 | 15.847 | C13H23O3+ | Methyl dihydrojasmonate | Oxylipid | 227.1645 | 212, 183, 141, 125 | 1.44 | |
| 78 | 15.893 | C20H33O4+ | Unknown | Terpene | 337.2368 | 293, 232, 195, 168 | −1.58 | |
| 79 | 15.938 | C18H29O3+ | Oxophytodienoic acid | Oxylipid | 293.2117 | 238, 151, 145, 101 | −0.02 | |
| 80 | 15.984 | C19H27O4+ | Unknown | Terpene | 319.1893 | 303, 235, 165, 105 | −3.4 | |
| 81 | 16.304 | C20H31O4+ | Unknown | - | 335.2218 | 318, 273,158,83 | 0.34 | |
| 82 | 16.578 | C18H29O3− | Hydroxylinolenic acid | Fatty acid | 293.215 | 275, 249, 233, 183 | −9.51 | |
| 83 | 16.578 | C18H29O2+ | Unknown | Terpene | 277.2164 | 149, 121, 107, 93 | 0.85 | |
| 84 | 16.67 | C18H27O2+ | Unknown | Terpene | 275.2015 | 149, 135, 121, 93 | −2.45 | |
| 85 | 16.761 | C20H29O3+ | Unknown | Terpene | 317.2088 | 299, 187, 164, 110 | −7.31 | |
| 86 | 16.898 | C18H29O3+ | Oxooctadeca-trienoic acid | Fatty acid | 293.2126 | 275, 248, 125, 93 | −5.77 | |
| 87 | 17.081 | C13H25O3+ | Hydroxy-tridecenoic acid | Fatty acid | 229.181 | 155, 109, 67 | 0.45 | |
| 88 | 17.172 | C18H37O6+ | Tetrahydroxy-octadecanoic acid (saturated) | Fatty acid | 349.2602 | 306, 277, 190, 83 | 4.96 | [51] |
| 89 | 17.264 | C18H31O2+ | Octadeca-trienoic acid (Linolenic acid) | Fatty acid | 279.2332 | 95, 81, 67 | −7.17 | |
| 90 | 17.538 | C22H35O3+ | Unknown | Terpene | 347.2564 | 276, 263, 174, 163 | 2.03 | |
| 91 | 17.583 | C18H29O2+ | Unknown | Terpene | 277.2172 | 150, 121, 95, 81 | −1.14 | |
| 92 | 17.629 | C22H37O3+ | Unknown | Terpene | 349.2737 | 315, 275, 214, 165 | −0.06 | |
| 93 | 17.995 | C24H41O5+ | Unknown | - | 409.294 | 391, 353, 123, 86 | −2.07 | |
| 94 | 18.132 | C18H21O4+ | Unknown | Terpene | 301.1426 | 214, 182, 135, 93 | −2.77 | |
| 95 | 18.269 | C18H31O3− | Hydroxyolinoleic acid | Fatty acid | 295.2303 | 277, 233, 195, 183 | −8.22 | |
| 96 | 18.863 | C25H48O12P− | Phosphoinositol (16:0/0:0) | Acyl glycerol | 571.2935 | 434, 409, 380, 315 | 8.07 | |
| 97 | 19 | C24H39O4+ | Unknown | Terpene | 391.283 | 373, 187, 85 | −3.28 | |
| 98 | 19.183 | C19H33O2+ | Methyl-octadeca-trienoate | Fatty acid | 293.2487 | 248, 169, 95 | −3.5 | [52] |
| 99 | 19.228 | C22H35O2+ | Docosapentaenoic acid | Fatty acid | 331.266 | 275, 235, 146, 67 | 8.58 | |
| 100 | 19.594 | C28H47O4+ | Unknown | Sterol | 447.3459 | 429, 359, 206, 149 | −2.2 | |
| 101 | 20.096 | C24H37O3+ | Unknown | Oxylipid | 373.2719 | 356, 329,284,235 | −4.8 | |
| 102 | 20.234 | C43H79NO10P− | Phosphoserine (19:0/18:2) | Acyl glycerol | 800.5521 | 741, 227 | 9.23 | |
| 103 | 20.371 | C16H31O3− | Hydroxypalmitate | Fatty acid | 271.2302 | 227, 225, 195, 151 | −7.7 | |
| 104 | 20.417 | C43H76O12P− | Phosphoinositol (P-16:0/18:4) | Acyl glycerol | 815.5059 | 758, 475, 428, 281 | −2.56 | |
| 105 | 20.599 | C45H81NO10P− | Phosphoserine (19:1/20:2) | Acyl glycerol | 826.566 | 767, 477,279 | 6.82 | |
| 106 | 20.691 | C34H68NO5− | Ceramide (t18:0/16:0(2OH)) | Sphingolipid | 570.5157 | 314, 255 | 9.46 | [53] |
| 107 | 21.056 | C30H49O2+ | Unknown | Sterol | 441.369 | 423, 361, 259,219 | −8.39 | |
| 108 | 22.472 | C42H84NO5+ | Ceramide (t18:1/24:0(2OH)) | Sphingolipid | 682.6342 | 665, 427, 372, 162, 74 | −0.29 | [53] |
| 109 | 22.747 | C48H92NO10− | Iotroridoside | Glycosphingolipid | 842.6782 | 438, 179, 89 | 6.56 | [54] |
| 110 | 22.838 | C43H77O13P− | Phosphatidylinositol (16:0/18:2) | Acyl glycerol | 832.5031 | 520, 366, 273 | −9.16 | |
| 111 | 22.93 | C43H74O13P− | Phosphatidylinositol (14:0/20:4) | Acyl glycerol | 829.4852 | 787, 225, 89 | −2.47 | |
| 112 | 23.295 | C45H74O13P− | Phosphatidylinositol (16:1/20:5) | Acyl glycerol | 853.4843 | 610, 339, 108 | −3.45 | |
| 113 | 23.706 | C43H76O13P− | Phosphatidylinositol (20:1/14:1) | Acyl glycerol | 831.5004 | 772, 459,80 | −3.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Newary, S.A.; Afifi, S.M.; Aly, M.S.; Ahmed, R.F.; El Gendy, A.E.-N.G.; Abd-ElGawad, A.M.; Farag, M.A.; Elgamal, A.M.; Elshamy, A.I. Chemical Profile of Launaea nudicaulis Ethanolic Extract and Its Antidiabetic Effect in Streptozotocin-Induced Rats. Molecules 2021, 26, 1000. https://doi.org/10.3390/molecules26041000
El-Newary SA, Afifi SM, Aly MS, Ahmed RF, El Gendy AE-NG, Abd-ElGawad AM, Farag MA, Elgamal AM, Elshamy AI. Chemical Profile of Launaea nudicaulis Ethanolic Extract and Its Antidiabetic Effect in Streptozotocin-Induced Rats. Molecules. 2021; 26(4):1000. https://doi.org/10.3390/molecules26041000
Chicago/Turabian StyleEl-Newary, Samah A., Sherif M. Afifi, Mohamed S. Aly, Rania F. Ahmed, Abd El-Nasser G. El Gendy, Ahmed M. Abd-ElGawad, Mohamed A. Farag, Abdelbaset M. Elgamal, and Abdelsamed I. Elshamy. 2021. "Chemical Profile of Launaea nudicaulis Ethanolic Extract and Its Antidiabetic Effect in Streptozotocin-Induced Rats" Molecules 26, no. 4: 1000. https://doi.org/10.3390/molecules26041000
APA StyleEl-Newary, S. A., Afifi, S. M., Aly, M. S., Ahmed, R. F., El Gendy, A. E. -N. G., Abd-ElGawad, A. M., Farag, M. A., Elgamal, A. M., & Elshamy, A. I. (2021). Chemical Profile of Launaea nudicaulis Ethanolic Extract and Its Antidiabetic Effect in Streptozotocin-Induced Rats. Molecules, 26(4), 1000. https://doi.org/10.3390/molecules26041000