
Tannic Acid-Stabilized Silver Nanoparticles Used in Biomedical Application as an Effective Antimelioidosis and Prolonged Efflux Pump Inhibitor against Melioidosis Causative Pathogen

 ,
,  and
and
Abstract
:1. Introduction
2. Results
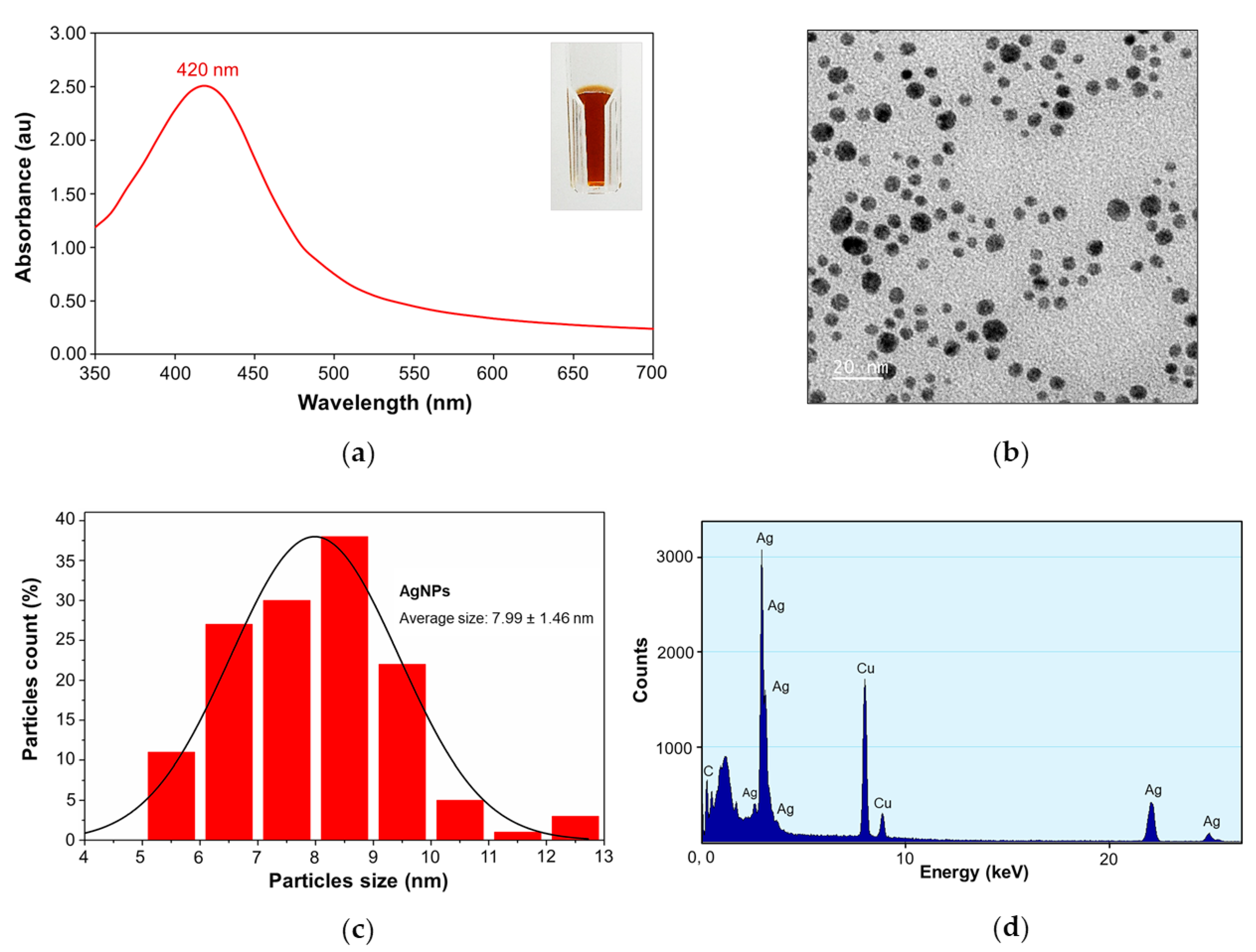
2.1. Physicochemical Characterization of Silver Nanoparticles
2.1.1. UV-visible Spectroscopy and Transmission Electron Microscopy
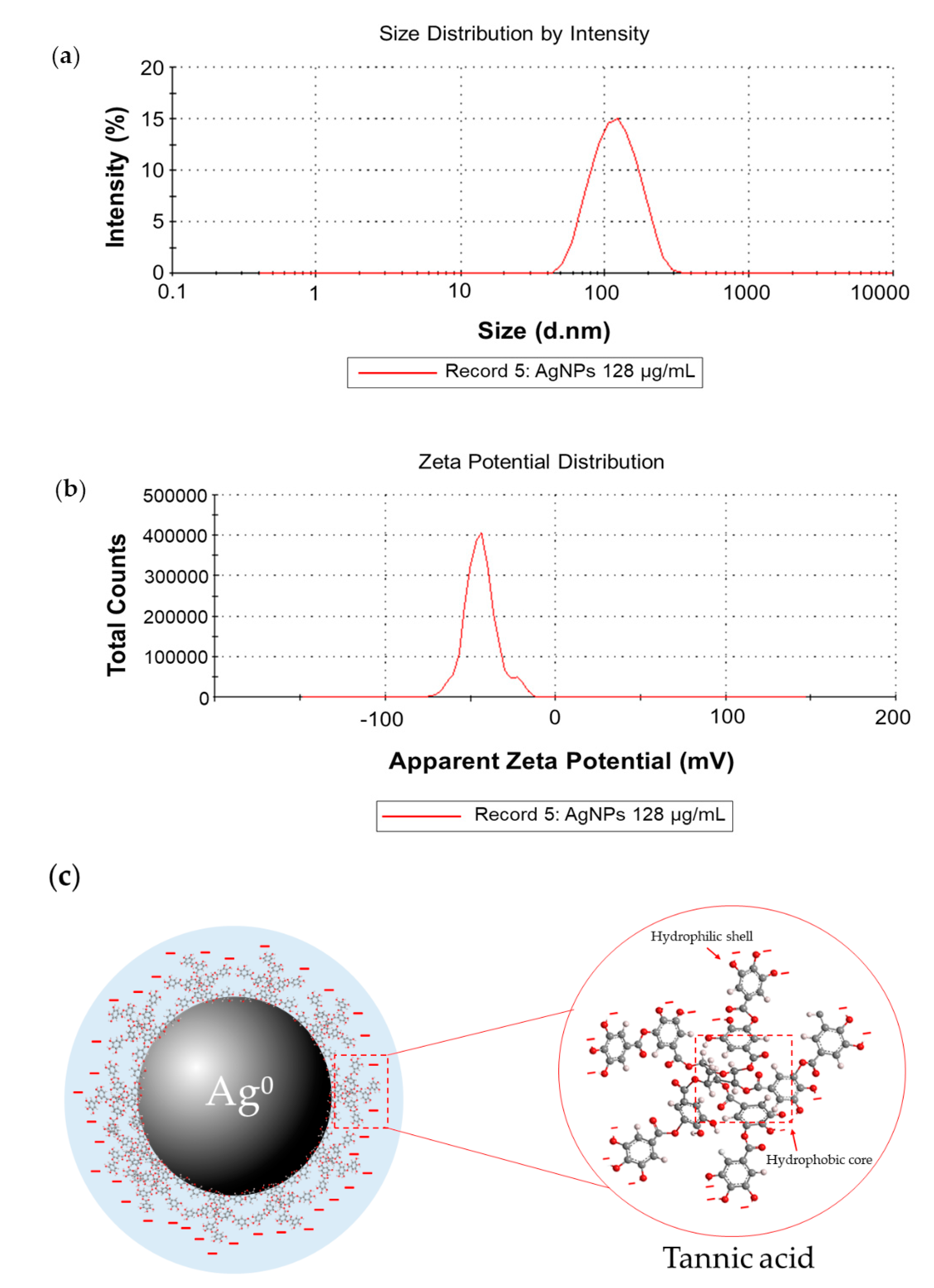
2.1.2. Dynamic Light Scattering and Zeta Potential Measurements
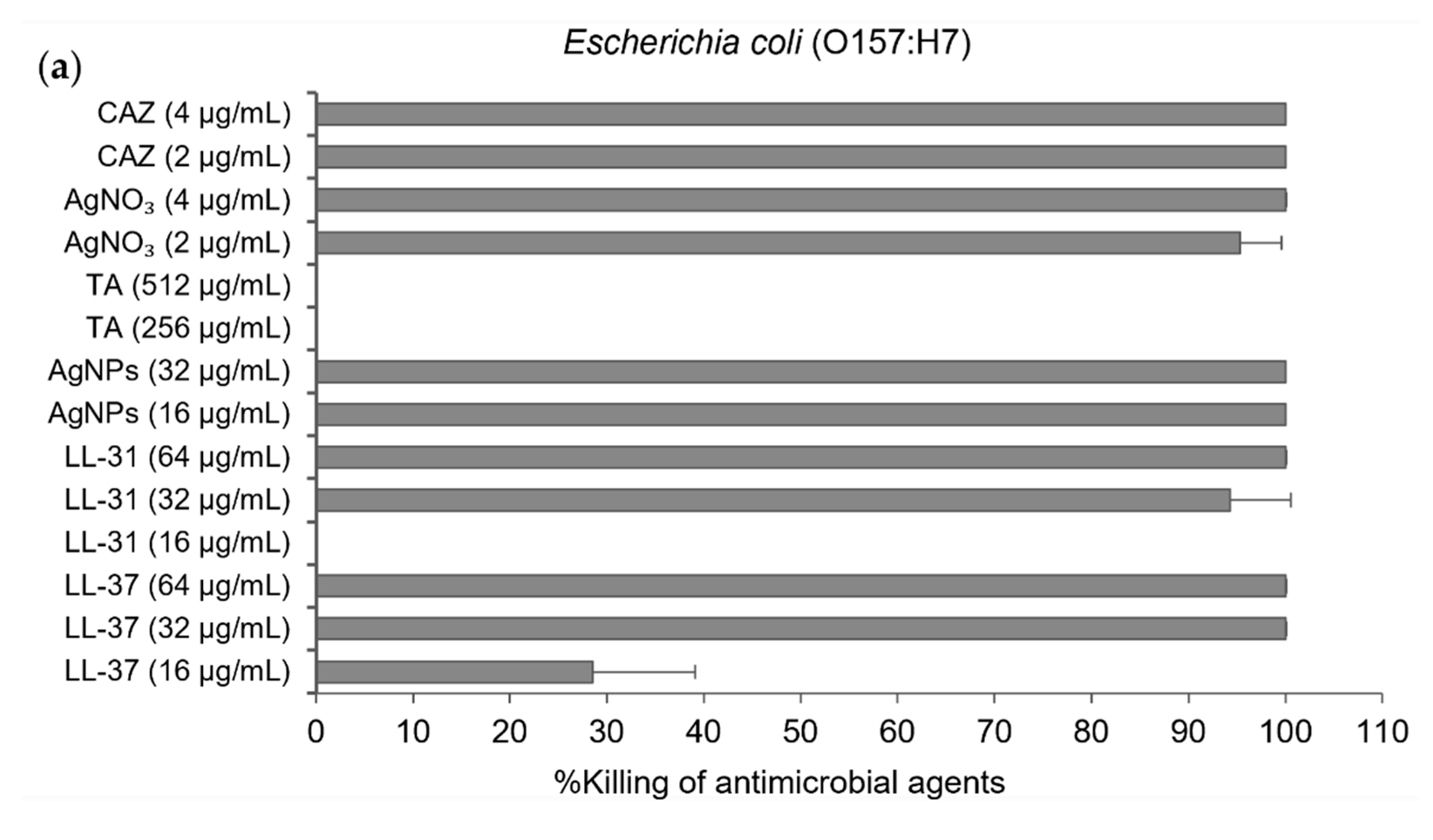
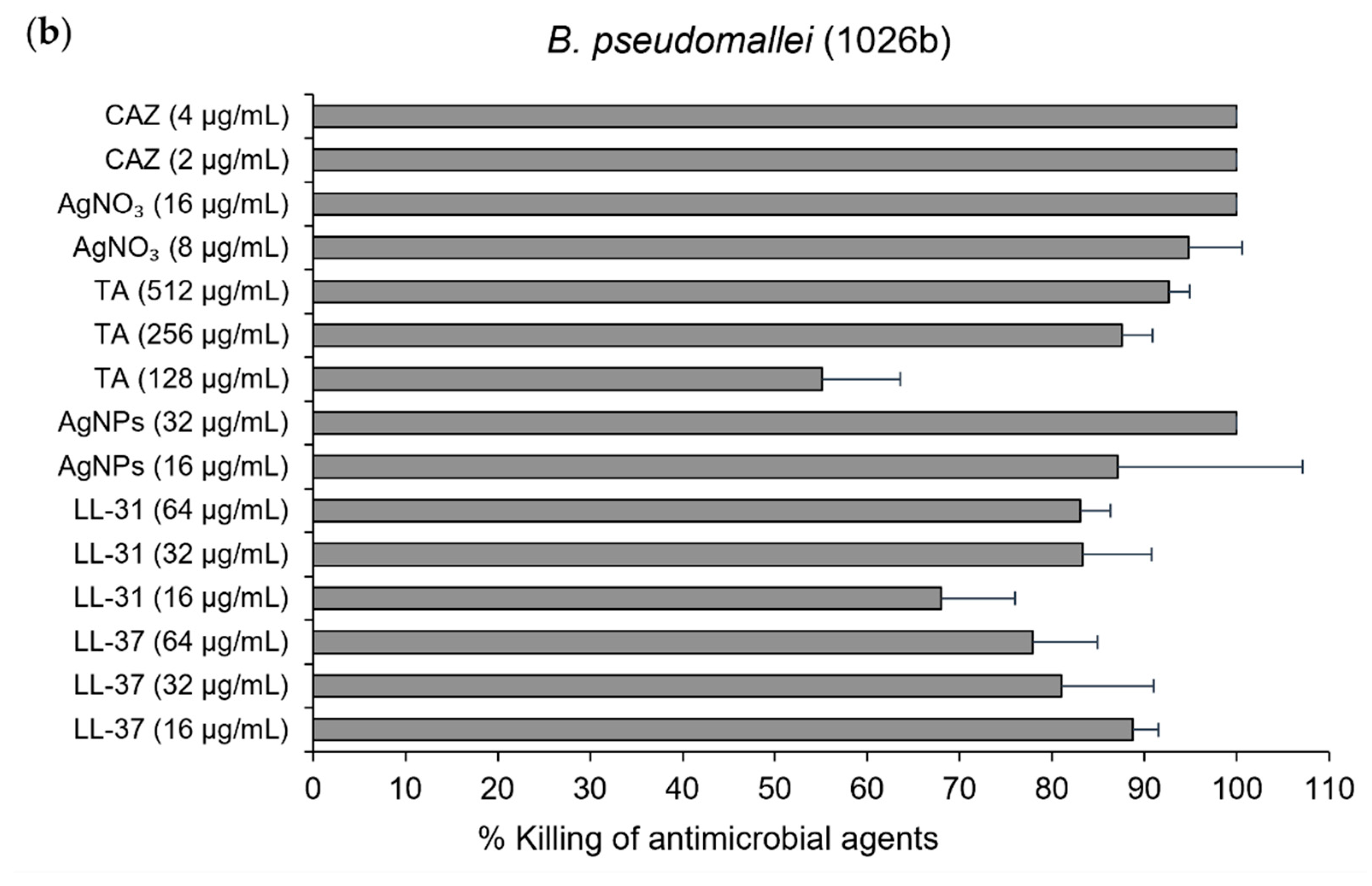
2.2. Efficient Antibacterial Activity of AgNPs against Both Susceptible and CAZ-Resistant Strains of B. pseudomallei
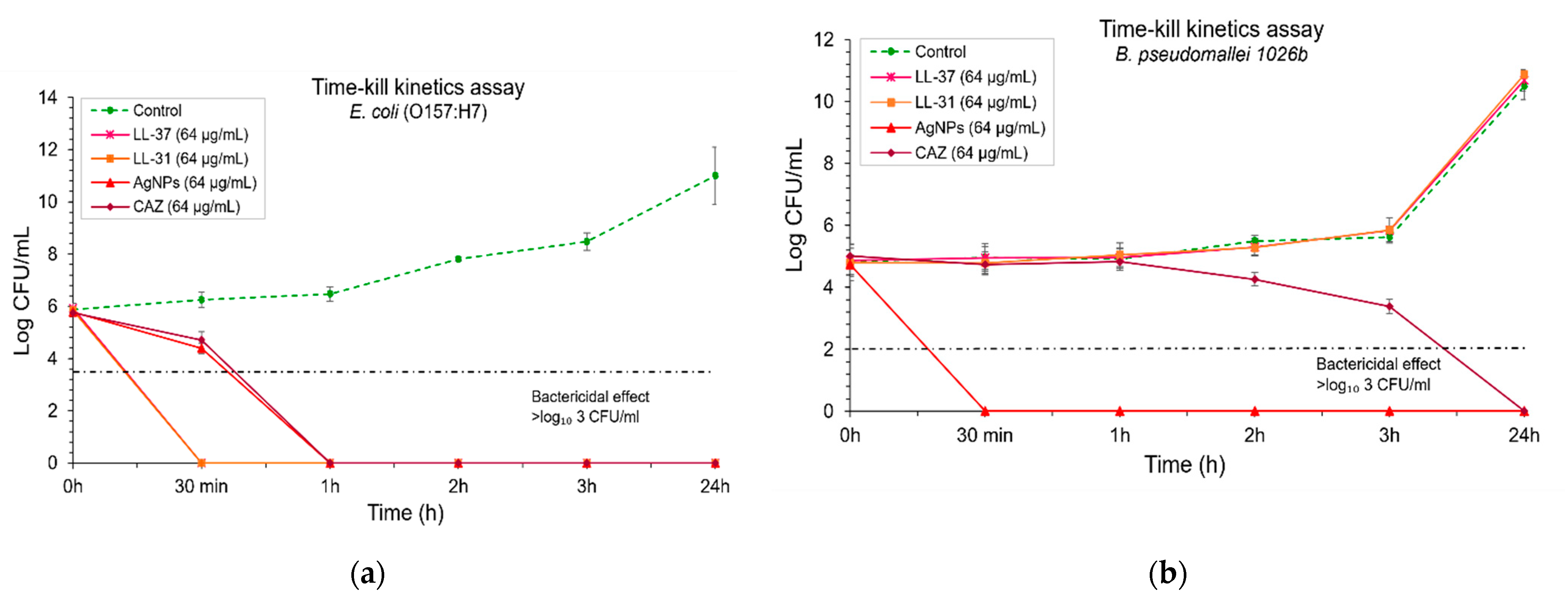
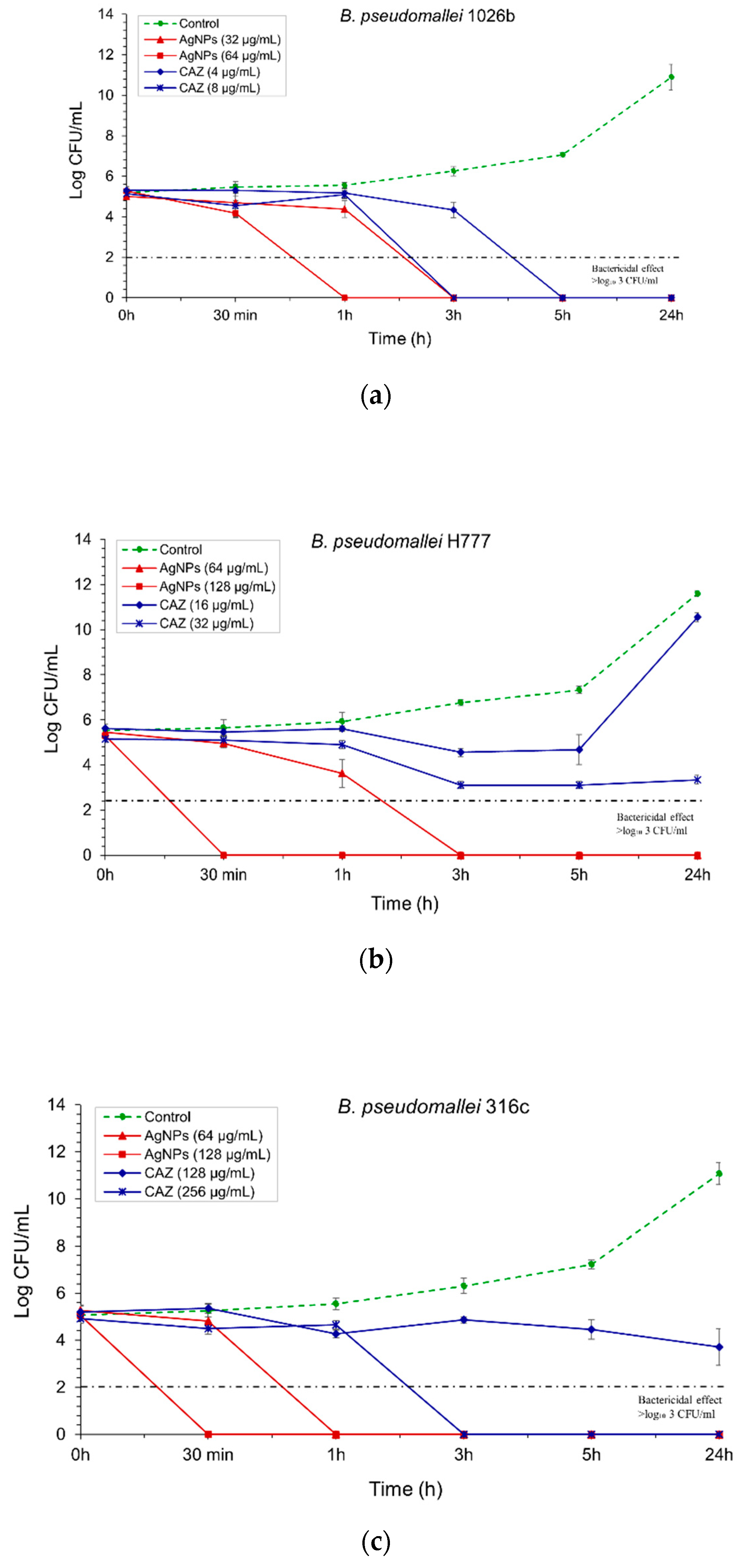
2.3. AgNPs Exhibited an Efficient, Fast Action of Bactericidal Effect Against B. pseudomallei
2.4. AgNPs Can Inhibit Biofilm Formation of B. pseudomallei
2.5. No Resistance Induction Developed by B. pseudomallei towards AgNPs after 30th Bacterial Generation Passage
2.6. AgNPs Still Showed Prolonged Efflux Pump Inhibiting Effect against B. pseudomallei via Phenotypic EtBr-agar Cartwheel Assay after 30th Bacterial Generation Passage
2.7. AgNPs Acted as an Effective Efflux Pump Inhibitor by Exhibiting a Maximum of 16-Fold Reduction in Efflux Pump Substrate MIC (EtBr)
3. Discussion
4. Materials and Methods
4.1. Bacterial Strain and Culture Media
4.2. Antibacterial Agents and Preparations
4.3. Characterization of AgNPs
4.4. Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC) Determination by Broth Microdilution and Resazurin Colorimetric Assay
4.5. Serial Colony Plate Counting Assay
4.6. Time-Kill Kinetics Assay
4.7. Biofilm Inhibition by Crystal Violet Assay
4.8. Resistance Induction Study
4.9. Phenotypic Efflux Pump Activity and Efflux Pump Inhibition Detection by EtBr-Cartwheel Assay
4.10. Efflux Pump Inhibition Evaluation and Fold Reduction in MIC by Microdilution Assay
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Hesstvedt, L.; Reikvam, D.H.; Dunlop, O. Neurological melioidosis in Norway presenting with a cerebral abscess. IDCases 2015, 2, 16–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wongkaewkhiaw, S.; Taweechaisupapong, S.; Anutrakunchai, C.; Nazmi, K.; Bolscher, J.G.; Wongratanacheewin, S.; Kanthawong, S. D-LL-31 in combination with ceftazidime synergistically enhances bactericidal activity and biofilm destruction inBurkholderia pseudomallei. Biofouling 2019, 35, 573–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanthawong, S.; Bolscher, J.G.; Veerman, E.C.; Van Marle, J.; De Soet, H.J.; Nazmi, K.; Wongratanacheewin, S.; Taweechaisupapong, S. Antimicrobial and antibiofilm activity of LL-37 and its truncated variants against Burkholderia pseudomallei. Int. J. Antimicrob. Agents 2012, 39, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Fjell, C.D.; Hiss, J.A.; Hancock, R.E.W.; Schneider, G. Designing antimicrobial peptides: Form follows function. Nat. Rev. Drug Discov. 2011, 11, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Inglis, T.J.J.; Rodrigues, F.; Rigby, P.; Norton, R.; Currie, B.J. Comparison of the Susceptibilities of Burkholderia pseudomallei to Meropenem and Ceftazidime by Conventional and Intracellular Methods. Antimicrob. Agents Chemother. 2004, 48, 2999–3005. [Google Scholar] [CrossRef] [Green Version]
- Sarovich, D.S.; Price, E.P.; Peacock, S.J.; Cook, J.M.; Schulze, V.; Wolken, S.R.; Keim, P.; Pearson, T.; Limmathurotsakul, D. Development of ceftazidime resistance in an acute Burkholderia pseudomallei infection. Infect. Drug Resist. 2012, 5, 129–132. [Google Scholar] [CrossRef] [Green Version]
- Ravindran, A.; Chandran, P.; Khan, S.S. Biofunctionalized silver nanoparticles: Advances and prospects. Colloids Surf. B Biointerfaces 2013, 105, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Soliman, H.; Elsayed, A.; Dyaa, A. Antimicrobial activity of silver nanoparticles biosynthesised by Rhodotorula sp. strain ATL72. Egypt. J. Basic Appl. Sci. 2018, 5, 228–233. [Google Scholar] [CrossRef] [Green Version]
- Graf, C.; Vossen, D.L.J.; Imhof, A.; Van Blaaderen, A. A General Method To Coat Colloidal Particles with Silica. Langmuir 2003, 19, 6693–6700. [Google Scholar] [CrossRef]
- Chen, X.; Schluesener, H.J. Nanosilver: A nanoproduct in medical application. Toxicol. Lett. 2008, 176, 1–12. [Google Scholar] [CrossRef]
- Pal, S.; Tak, Y.K.; Song, J.M. Does the Antibacterial Activity of Silver Nanoparticles Depend on the Shape of the Nanoparticle? A Study of the Gram-Negative Bacterium Escherichia coli. Appl. Environ. Microbiol. 2007, 73, 1712–1720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dong, P.; Ha, C.H.; Binh, L.T.; Kasbohm, J. Chemical synthesis and antibacterial activity of novel-shaped silver nanoparticles. Int. Nano Lett. 2012, 2, 9. [Google Scholar] [CrossRef] [Green Version]
- Siritongsuk, P.; Hongsing, N.; Thammawithan, S.; Daduang, S.; Klaynongsruang, S.; Tuanyok, A.; Patramanon, R. Two-Phase Bactericidal Mechanism of Silver Nanoparticles against Burkholderia pseudomallei. PLoS ONE 2016, 11, e0168098. [Google Scholar] [CrossRef] [PubMed]
- Markowska, K.; Grudniak, A.M.; I Wolska, K. Silver nanoparticles as an alternative strategy against bacterial biofilms. Acta Biochim. Pol. 2013, 60, 523–530. [Google Scholar] [CrossRef] [Green Version]
- Fabrega, J.; Renshaw, J.C.; Lead, J.R. Interactions of Silver Nanoparticles withPseudomonas putidaBiofilms. Environ. Sci. Technol. 2009, 43, 9004–9009. [Google Scholar] [CrossRef]
- Kalishwaralal, K.; BarathManiKanth, S.; Pandian, S.R.K.; Deepak, V.; Gurunathan, S. Silver nanoparticles impede the biofilm formation by Pseudomonas aeruginosa and Staphylococcus epidermidis. Colloids Surf. B Biointerfaces 2010, 79, 340–344. [Google Scholar] [CrossRef]
- Martinez-Gutierrez, F.; Boegli, L.; Agostinho, A.; Sánchez, E.M.; Bach, H.; Ruiz, F.; James, G. Anti-biofilm activity of silver nanoparticles against different microorganisms. Biofouling 2013, 29, 651–660. [Google Scholar] [CrossRef]
- Zhang, X.-F.; Liu, Z.-G.; Shen, W.; Gurunathan, S. Silver Nanoparticles: Synthesis, Characterization, Properties, Applications, and Therapeutic Approaches. Int. J. Mol. Sci. 2016, 17, 1534. [Google Scholar] [CrossRef]
- Kim, T.Y.; Cha, S.-H.; Cho, S.; Park, Y. Tannic acid-mediated green synthesis of antibacterial silver nanoparticles. Arch. Pharmacal Res. 2016, 39, 465–473. [Google Scholar] [CrossRef]
- Myint, K.B.; Sing, L.C.; Wei, Z. Tannic Acid as Phytochemical Potentiator for Antibiotic Resistance Adaptation. APCBEE Procedia 2013, 7, 175–181. [Google Scholar] [CrossRef] [Green Version]
- Sharma, G.; Nam, J.-S.; Sharma, A.R.; Lee, S.-S. Antimicrobial Potential of Silver Nanoparticles Synthesized Using Medicinal Herb Coptidis rhizome. Molecules 2018, 23, 2268. [Google Scholar] [CrossRef] [Green Version]
- Moteriya, P.; Padalia, H.; Chanda, S. Characterization, synergistic antibacterial and free radical scavenging efficacy of silver nanoparticles synthesized using Cassia roxburghii leaf extract. J. Genet. Eng. Biotechnol. 2017, 15, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Danaei, M.; Dehghankhold, M.; Ataei, S.; Davarani, F.H.; Javanmard, R.; Dokhani, A.; Khorasani, S.; Mozafari, M.R. Impact of Particle Size and Polydispersity Index on the Clinical Applications of Lipidic Nanocarrier Systems. Pharmaceutics 2018, 10, 57. [Google Scholar] [CrossRef] [Green Version]
- Wypij, M.; Jędrzejewski, T.; Ostrowski, M.; Trzcińska, J.; Rai, M.; Golińska, P. Biogenic Silver Nanoparticles: Assessment of Their Cytotoxicity, Genotoxicity and Study of Capping Proteins. Molecules 2020, 25, 3022. [Google Scholar] [CrossRef]
- Maillard, A.P.F.; Gonçalves, S.; Santos, N.C.; De Mishima, B.A.L.; Dalmasso, P.R.; Hollmann, A. Studies on interaction of green silver nanoparticles with whole bacteria by surface characterization techniques. Biochim. Biophys. Acta Biomembr. 2019, 1861, 1086–1092. [Google Scholar] [CrossRef]
- Arya, S.S.; Sharma, M.M.; Das, R.K.; Rookes, J.; Cahill, D.; Lenka, S.K. Vanillin mediated green synthesis and application of gold nanoparticles for reversal of antimicrobial resistance in Pseudomonas aeruginosa clinical isolates. Heliyon 2019, 5, e02021. [Google Scholar] [CrossRef] [Green Version]
- Christena, L.R.; Mangalagowri, V.; Pradheeba, P.; Ahmed, K.B.A.; Shalini, B.I.S.; Vidyalakshmi, M.; Anbazhagan, V.; Subramanian, N.S. Copper nanoparticles as an efflux pump inhibitor to tackle drug resistant bacteria. RSC Adv. 2015, 5, 12899–12909. [Google Scholar] [CrossRef]
- Behdad, R.; Pargol, M.; Mirzaie, A.; Karizi, S.Z.; Noorbazargan, H.; Akbarzadeh, I. Efflux pump inhibitory activity of biologically synthesized silver nanoparticles against multidrug-resistant Acinetobacter baumannii clinical isolates. J. Basic Microbiol. 2020, 60, 494–507. [Google Scholar] [CrossRef] [PubMed]
- Fatima, F.; Verma, S.R.; Pathak, N.; Bajpai, P. Extracellular mycosynthesis of silver nanoparticles and their microbicidal activity. J. Glob. Antimicrob. Resist. 2016, 7, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Rai, M.; Deshmukh, S.; Ingle, A.; Gade, A. Silver nanoparticles: The powerful nanoweapon against multidrug-resistant bacteria. J. Appl. Microbiol. 2012, 112, 841–852. [Google Scholar] [CrossRef]
- Cao, Y.; Zheng, R.; Ji, X.; Liu, H.; Xie, R.; Yang, W. Syntheses and Characterization of Nearly Monodispersed, Size-Tunable Silver Nanoparticles over a Wide Size Range of 7–200 nm by Tannic Acid Reduction. Langmuir 2014, 30, 3876–3882. [Google Scholar] [CrossRef] [PubMed]
- Cataldo, F.; Angelini, G. A Green Synthesis of Colloidal Silver Nanoparticles and Their Reaction with Ozone. Eur. Chem. Bull. 2013, 2, 700–705. [Google Scholar]
- Ahmad, T. Reviewing the Tannic Acid Mediated Synthesis of Metal Nanoparticles. J. Nanotechnol. 2014, 2014, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Chaturvedi, V.K.; Yadav, N.; Rai, N.K.; Ellah, N.H.A.; Bohara, R.A.; Rehan, I.F.; Marraiki, N.; Batiha, G.E.-S.; Hetta, H.F.; Singh, P. Pleurotus sajor-caju-Mediated Synthesis of Silver and Gold Nanoparticles Active against Colon Cancer Cell Lines: A New Era of Herbonanoceutics. Molecules 2020, 25, 3091. [Google Scholar] [CrossRef]
- Abudalo, M.A.; Al-Mheidat, I.R.; Al-Shurafat, A.W.; Grinham, C.; Oyanedel-Craver, V. Synthesis of silver nanoparticles using a modified Tollens’ method in conjunction with phytochemicals and assessment of their antimicrobial activity. PeerJ 2019, 7, e6413. [Google Scholar] [CrossRef] [Green Version]
- Ajitha, B.; Reddy, Y.A.K.; Reddy, P.S.; Jeon, H.-J.; Ahn, C.W. Role of capping agents in controlling silver nanoparticles size, antibacterial activity and potential application as optical hydrogen peroxide sensor. RSC Adv. 2016, 6, 36171–36179. [Google Scholar] [CrossRef]
- Orlowski, P.; Tomaszewska, E.; Ranoszek-Soliwoda, K.; Gniadek, M.; Labedz, O.; Malewski, T.; Nowakowska, J.; Chodaczek, G.; Celichowski, G.; Grobelny, J.; et al. Tannic Acid-Modified Silver and Gold Nanoparticles as Novel Stimulators of Dendritic Cells Activation. Front. Immunol. 2018, 9, 1115. [Google Scholar] [CrossRef] [PubMed]
- Morones, J.R.; Elechiguerra, J.L.; Camacho, A.; Holt, K.; Kouri, J.B.; Ramírez, J.T.; Yacaman, M.J. The bactericidal effect of silver nanoparticles. Nanotechnology 2005, 16, 2346–2353. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Ge, C.; Zhang, Y.; Ma, W.; Su, X.; Chen, L.; Li, S.; Wang, L.; Mu, X.; Xu, Y. Tannic acid-modified silver nanoparticles for enhancing anti-biofilm activities and modulating biofilm formation. Biomater. Sci. 2020, 8, 4852–4860. [Google Scholar] [CrossRef]
- Lu, Z.; Rong, K.; Li, J.; Yang, H.; Chen, R. Size-dependent antibacterial activities of silver nanoparticles against oral anaerobic pathogenic bacteria. J. Mater. Sci. Mater. Med. 2013, 24, 1465–1471. [Google Scholar] [CrossRef]
- Raza, M.A.; Kanwal, Z.; Rauf, A.; Sabri, A.N.; Riaz, S.; Naseem, S. Size- and Shape-Dependent Antibacterial Studies of Silver Nanoparticles Synthesized by Wet Chemical Routes. Nanomaterials 2016, 6, 74. [Google Scholar] [CrossRef] [Green Version]
- A Skomorokhova, E.; Sankova, T.P.; A Orlov, I.; Savelev, A.N.; Magazenkova, D.N.; Pliss, M.G.; Skvortsov, A.N.; Sosnin, I.M.; A Kirilenko, D.; Grishchuk, I.V.; et al. Size-Dependent Bioactivity of Silver Nanoparticles: Antibacterial Properties, Influence on Copper Status in Mice, and Whole-Body Turnover. Nanotechnol. Sci. Appl. 2020, 13, 137–157. [Google Scholar] [CrossRef]
- Feng, Q.L.; Chen, G.Q.; Cui, F.Z.; Kim, T.N.; Kim, J.O. A Mechanistic Study of the Antibacterial Effect of Silver Ions on Escherichia Coli and Staphylococcus Aureus. J. Biomed. Mater. Res. 2000, 52, 662–668. [Google Scholar] [CrossRef]
- Ipe, D.S.; Kumar, P.T.S.; Love, R.M.; Hamlet, S.M. Silver Nanoparticles at Biocompatible Dosage Synergistically Increases Bacterial Susceptibility to Antibiotics. Front. Microbiol. 2020, 11, 1074. [Google Scholar] [CrossRef]
- Bakhtiari-Sardari, A.; Mashreghi, M.; Eshghi, H.; Behnam-Rasouli, F.; Lashani, E.; Shahnavaz, B. Comparative evaluation of silver nanoparticles biosynthesis by two cold-tolerant Streptomyces strains and their biological activities. Biotechnol. Lett. 2020, 42, 1985–1999. [Google Scholar] [CrossRef] [PubMed]
- Shahmiri, M.; Enciso, M.; Adda, C.G.; Smith, B.J.; Perugini, M.A.; Mechler, A. Membrane Core-Specific Antimicrobial Action of Cathelicidin LL-37 Peptide Switches Between Pore and Nanofibre Formation. Sci. Rep. 2016, 6, 38184. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, K.; Okumura, K.; Isogai, H.; Isogai, E. The Human Cathelicidin Antimicrobial Peptide LL-37 and Mimics are Potential Anticancer Drugs. Front. Oncol. 2015, 5, 144. [Google Scholar] [CrossRef] [Green Version]
- Anantharaman, A.; Rizvi, M.S.; Sahal, D. Synergy with Rifampin and Kanamycin Enhances Potency, Kill Kinetics, and Selectivity of De Novo-Designed Antimicrobial Peptides. Antimicrob. Agents Chemother. 2010, 54, 1693–1699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foerster, S.; Unemo, M.; Hathaway, L.J.; Low, N.; Althaus, C.L. Time-kill curve analysis and pharmacodynamic modelling for in vitro evaluation of antimicrobials against Neisseria gonorrhoeae. BMC Microbiol. 2016, 16, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorachit, M.; Lam, K.; Jayanetra, P.; Costerton, J.W. Electron Microscopy of the Mode of Growth of Pseudomonas Psedomallei in Vitro and in Vivo. J. Trop. Med. Hyg. 1995, 98, 379–391. [Google Scholar]
- Sawasdidoln, C.; Taweechaisupapong, S.; Sermswan, R.W.; Tattawasart, U.; Tungpradabkul, S.; Wongratanacheewin, S. Growing Burkholderia pseudomallei in Biofilm Stimulating Conditions Significantly Induces Antimicrobial Resistance. PLoS ONE 2010, 5, e9196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palanisamy, N.K.; Ferina, N.; Amirulhusni, A.N.; Mohd-Zain, Z.; Hussaini, J.; Ping, L.J.; Durairaj, R. Antibiofilm properties of chemically synthesized silver nanoparticles found against Pseudomonas aeruginosa. J. Nanobiotechnol. 2014, 12, 2. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Yu, T.; Xu, P.; Xu, X.; Ji, S.; Gao, W.; Shi, L. Role of Efflux Pumps in the in vitro Development of Ciprofloxacin Resistance in Listeria monocytogenes. Front. Microbiol. 2018, 9, 2350. [Google Scholar] [CrossRef]
- Anbazhagan, P.V.; Thavitiki, P.R.; Varra, M.; Annamalai, L.; Putturu, R.; Lakkineni, V.R.; Pesingi, P.K. Evaluation of efflux pump activity of multidrug-resistant Salmonella Typhimurium isolated from poultry wet markets in India. Infect. Drug Resist. 2019, 12, 1081–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podnecky, N.L.; Rhodes, K.A.; Schweizer, H.P. Efflux pump-mediated drug resistance in Burkholderia. Front. Microbiol. 2015, 6, 305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pathania, R.; Sharma, A.; Gupta, V.K. Efflux pump inhibitors for bacterial pathogens: From bench to bedside. Indian J. Med Res. 2019, 149, 129–145. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, A.K.; Chakraborty, R.; Basu, T. Mechanism of antibacterial activity of copper nanoparticles. Nanotechnology 2014, 25, 135101. [Google Scholar] [CrossRef]
- Mishra, M.; Kumar, S.; Majhi, R.K.; Goswami, L.; Goswami, C.; Mohapatra, H. Antibacterial Efficacy of Polysaccharide Capped Silver Nanoparticles Is Not Compromised by AcrAB-TolC Efflux Pump. Front. Microbiol. 2018, 9, 823. [Google Scholar] [CrossRef] [PubMed]
- Nallathamby, P.D.; Lee, K.J.; Desai, T.; Xu, X.-H.N. Study of the Multidrug Membrane Transporter of Single LivingPseudomonas aeruginosaCells Using Size-Dependent Plasmonic Nanoparticle Optical Probes. Biochemistry 2010, 49, 5942–5953. [Google Scholar] [CrossRef] [Green Version]
- Salas-Orozco, M.; Niño-Martínez, N.; Martínez-Castañón, G.-A.; Méndez, F.T.; Jasso, M.E.C.; Ruiz, F. Mechanisms of Resistance to Silver Nanoparticles in Endodontic Bacteria: A Literature Review. J. Nanomater. 2019, 2019, 1–11. [Google Scholar] [CrossRef]
- Dakal, T.C.; Kumar, A.; Majumdar, R.S.; Yadav, V. Mechanistic Basis of Antimicrobial Actions of Silver Nanoparticles. Front. Microbiol. 2016, 7, 1831. [Google Scholar] [CrossRef] [Green Version]
- Dobias, J.; Bernier-Latmani, R. Silver Release from Silver Nanoparticles in Natural Waters. Environ. Sci. Technol. 2013, 47, 4140–4146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Third Informational Supplement. CLSI document M100-S23 (ISBN 1-56238-866-5 [Electronic]). Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2013. [Google Scholar]
- Irazazabal, L.N.; Porto, W.F.; Fensterseifer, I.C.; Alves, E.S.; Matos, C.O.; Menezes, A.C.; Felício, M.R.; Gonçalves, S.; Santos, N.C.; Ribeiro, S.M.; et al. Fast and potent bactericidal membrane lytic activity of PaDBS1R1, a novel cationic antimicrobial peptide. Biochim. Biophys. Acta Biomembr. 2019, 1861, 178–190. [Google Scholar] [CrossRef]
- Silveira, Z.D.S.; Macêdo, N.S.; Dos Santos, J.F.S.; De Freitas, T.S.; Barbosa, C.R.D.S.; Júnior, D.L.D.S.; Muniz, D.F.; De Oliveira, L.C.C.; Júnior, J.P.S.; Da Cunha, F.A.B.; et al. Evaluation of the Antibacterial Activity and Efflux Pump Reversal of Thymol and Carvacrol against Staphylococcus aureus and Their Toxicity in Drosophila melanogaster. Molecules 2020, 25, 2103. [Google Scholar] [CrossRef]
- Teh, C.H.; Nazni, W.A.; Nurulhusna, A.H.; Norazah, A.; Lee, H.L. Determination of antibacterial activity and minimum inhibitory concentration of larval extract of fly via resazurin-based turbidometric assay. BMC Microbiol. 2017, 17, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sengyee, S.; Saiprom, N.; Paksanont, S.; Limmathurotsakul, D.; Wuthiekanun, V.; Chantratita, N. Susceptibility of Clinical Isolates of Burkholderia pseudomallei to a Lipid A Biosynthesis Inhibitor. Am. J. Trop. Med. Hyg. 2017, 97, 62–67. [Google Scholar] [CrossRef]
- Pankey, G.A.; Ashcraft, D.S. The detection of synergy between meropenem and polymyxin B against meropenem-resistant Acinetobacter baumannii using Etest® and time-kill assay. Diagn. Microbiol. Infect. Dis. 2009, 63, 228–232. [Google Scholar] [CrossRef]
- Kunyanee, C.; Kamjumphol, W.; Taweechaisupapong, S.; Kanthawong, S.; Wongwajana, S.; Wongratanacheewin, S.; Hahnvajanawong, C.; Chareonsudjai, S. Burkholderia pseudomallei Biofilm Promotes Adhesion, Internalization and Stimulates Proinflammatory Cytokines in Human Epithelial A549 Cells. PLoS ONE 2016, 11, e0160741. [Google Scholar] [CrossRef] [Green Version]
- Lemos, A.S.O.; Campos, L.M.; Melo, L.; Guedes, M.C.M.R.; Oliveira, L.G.; Silva, T.P.; Melo, R.C.N.; Rocha, V.N.; Aguiar, J.A.K.; Apolônio, A.C.M.; et al. Antibacterial and Antibiofilm Activities of Psychorubrin, a Pyranonaphthoquinone Isolated From Mitracarpus frigidus (Rubiaceae). Front. Microbiol. 2018, 9, 724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elbehiry, A.; Al-Dubaib, M.; Marzouk, E.; Moussa, I. Antibacterial effects and resistance induction of silver and gold nanoparticles againstStaphylococcus aureus-induced mastitis and the potential toxicity in rats. Microbiol. Open 2019, 8, e00698. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibacterial Agents (µg/mL) | Escherichia coli (strain O157:H7) | Burkholderia pseudomallei (strain 1026b) | ||
|---|---|---|---|---|
| MIC 2 | MBC 3 | MIC | MBC | |
| LL-37 | 32 | 32 | >64 | >64 |
| LL-31 | 32 | 64 | >64 | >64 |
| AgNPs | 16 | 16 | 32 | 32 |
| AgNO3 | 4 | 4 | 16 | 16 |
| Tannic acid (TA) | >512 | >512 | >512 | >512 |
| Ceftazidime (CAZ) 1 | 2 | 2 | 2 | 2 |
| Antibacterial Agents (µg/mL) | B. pseudomallei (Strains) | |||||
|---|---|---|---|---|---|---|
| 1026b 1 | H777 2 | 316c 3 | ||||
| MIC | MBC | MIC | MBC | MIC | MBC | |
| AgNPs | 32 | 32 | 64 | 128 | 64 | 128 |
| AgNO₃ | 16 | 16 | 16 | 16 | 16 | 16 |
| Tannic acid (TA) | >512 | >512 | >512 | >512 | >512 | >512 |
| Ceftazidime (CAZ) | 2 | 2 | 4 | 4 | 64 | 128 |
| Bp Strain | Agents Used for Evaluation as an EPI | MIC Before Passage | MIC After 30th Passage | ||||||
|---|---|---|---|---|---|---|---|---|---|
| MIC of Agent (µg/mL) | MIC of EtBr (µg/mL) | Reduction (n-fold) in EtBr MIC | MIC of Agent (µg/mL) | MIC of EtBr (µg/mL) | Reduction (n-fold) in EtBr MIC | ||||
| Without Agent | With Agent | Without Agent | With Agent | ||||||
| 1026b | Ceftazidime | 2 | 128 | 32 | 4 | 2 | 128 | 32 | 4 |
| AgNPs | 32 | 128 | 8 | 16 | 32 | 128 | 8 | 16 | |
| H777 | Ceftazidime | 4 | 128 | 32 | 4 | 4 | 128 | 32 | 4 |
| AgNPs | 64 | 128 | 16 | 8 | 64 | 128 | 16 | 8 | |
| 316c | Ceftazidime | 64 | 128 | 32 | 4 | 64 | 128 | 32 | 4 |
| AgNPs | 64 | 128 | 32 | 4 | 64 | 128 | 32 | 4 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Srichaiyapol, O.; Thammawithan, S.; Siritongsuk, P.; Nasompag, S.; Daduang, S.; Klaynongsruang, S.; Kulchat, S.; Patramanon, R. Tannic Acid-Stabilized Silver Nanoparticles Used in Biomedical Application as an Effective Antimelioidosis and Prolonged Efflux Pump Inhibitor against Melioidosis Causative Pathogen. Molecules 2021, 26, 1004. https://doi.org/10.3390/molecules26041004
Srichaiyapol O, Thammawithan S, Siritongsuk P, Nasompag S, Daduang S, Klaynongsruang S, Kulchat S, Patramanon R. Tannic Acid-Stabilized Silver Nanoparticles Used in Biomedical Application as an Effective Antimelioidosis and Prolonged Efflux Pump Inhibitor against Melioidosis Causative Pathogen. Molecules. 2021; 26(4):1004. https://doi.org/10.3390/molecules26041004
Chicago/Turabian StyleSrichaiyapol, Oranee, Saengrawee Thammawithan, Pawinee Siritongsuk, Sawinee Nasompag, Sakda Daduang, Sompong Klaynongsruang, Sirinan Kulchat, and Rina Patramanon. 2021. "Tannic Acid-Stabilized Silver Nanoparticles Used in Biomedical Application as an Effective Antimelioidosis and Prolonged Efflux Pump Inhibitor against Melioidosis Causative Pathogen" Molecules 26, no. 4: 1004. https://doi.org/10.3390/molecules26041004
APA StyleSrichaiyapol, O., Thammawithan, S., Siritongsuk, P., Nasompag, S., Daduang, S., Klaynongsruang, S., Kulchat, S., & Patramanon, R. (2021). Tannic Acid-Stabilized Silver Nanoparticles Used in Biomedical Application as an Effective Antimelioidosis and Prolonged Efflux Pump Inhibitor against Melioidosis Causative Pathogen. Molecules, 26(4), 1004. https://doi.org/10.3390/molecules26041004