
Antifeedant Effects and Repellent Activity of Loline Alkaloids from Endophyte-Infected Tall Fescue against Horn Flies, Haematobia irritans (Diptera: Muscidae)

 , ,
, , 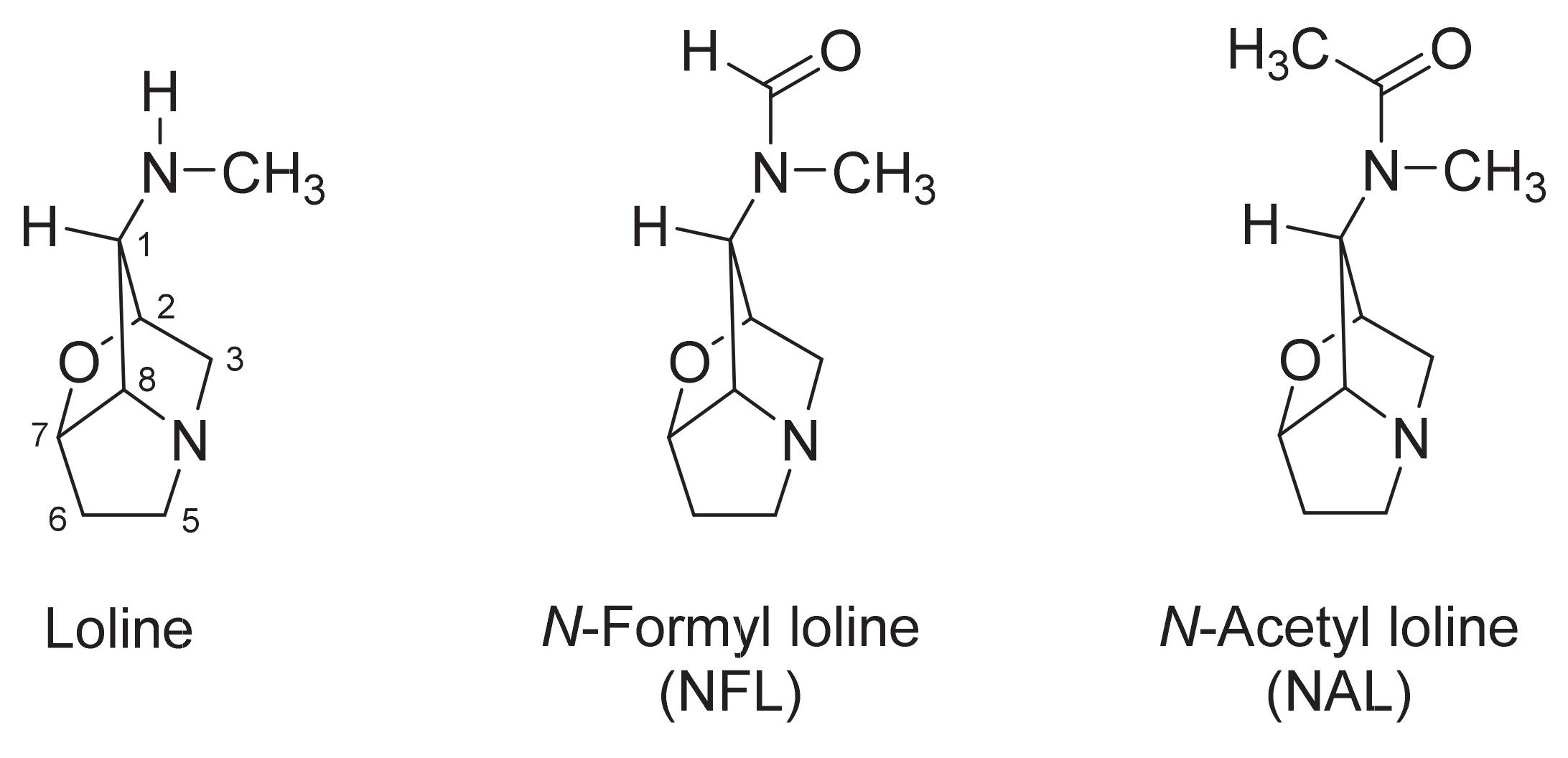{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
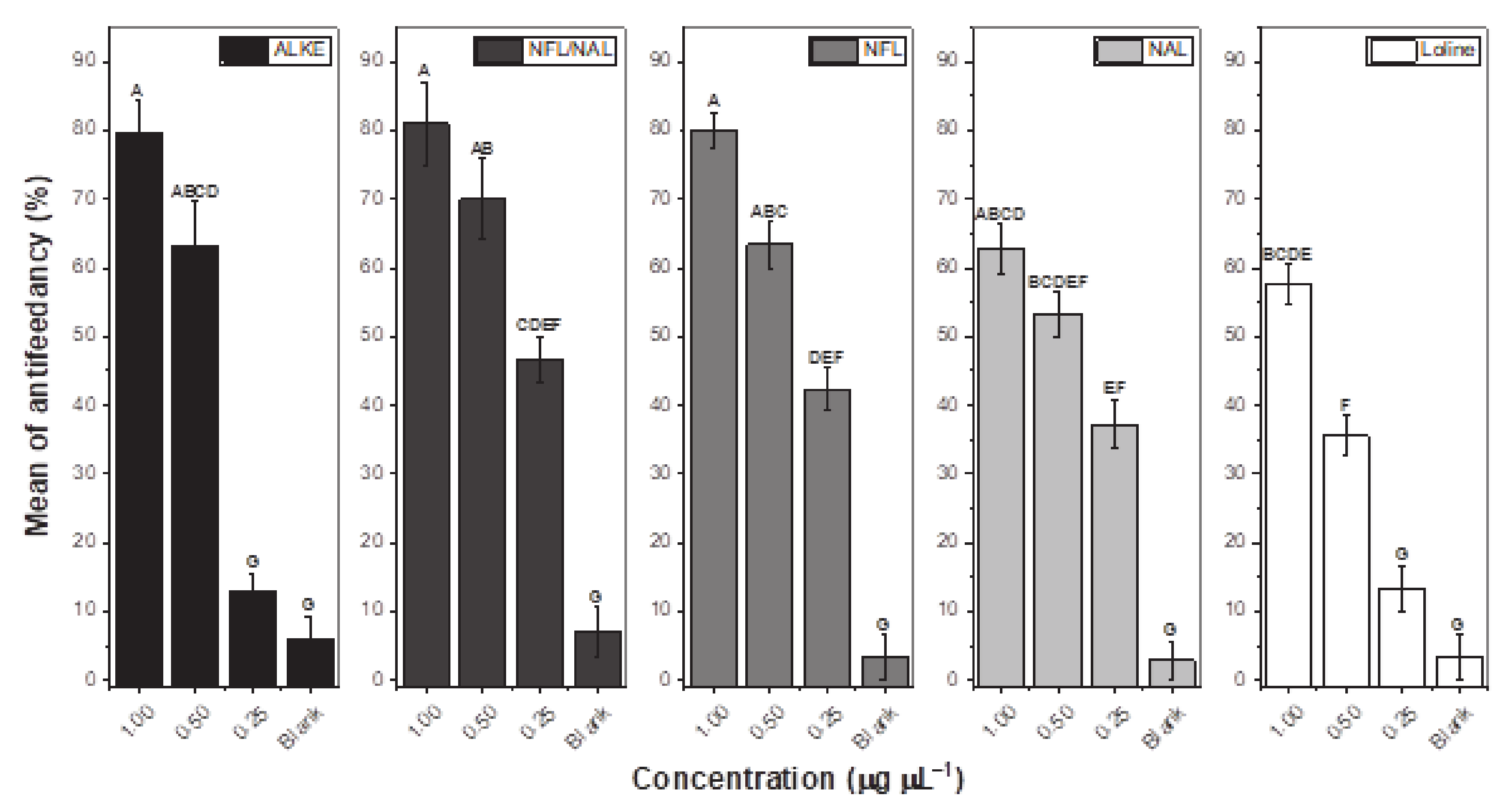
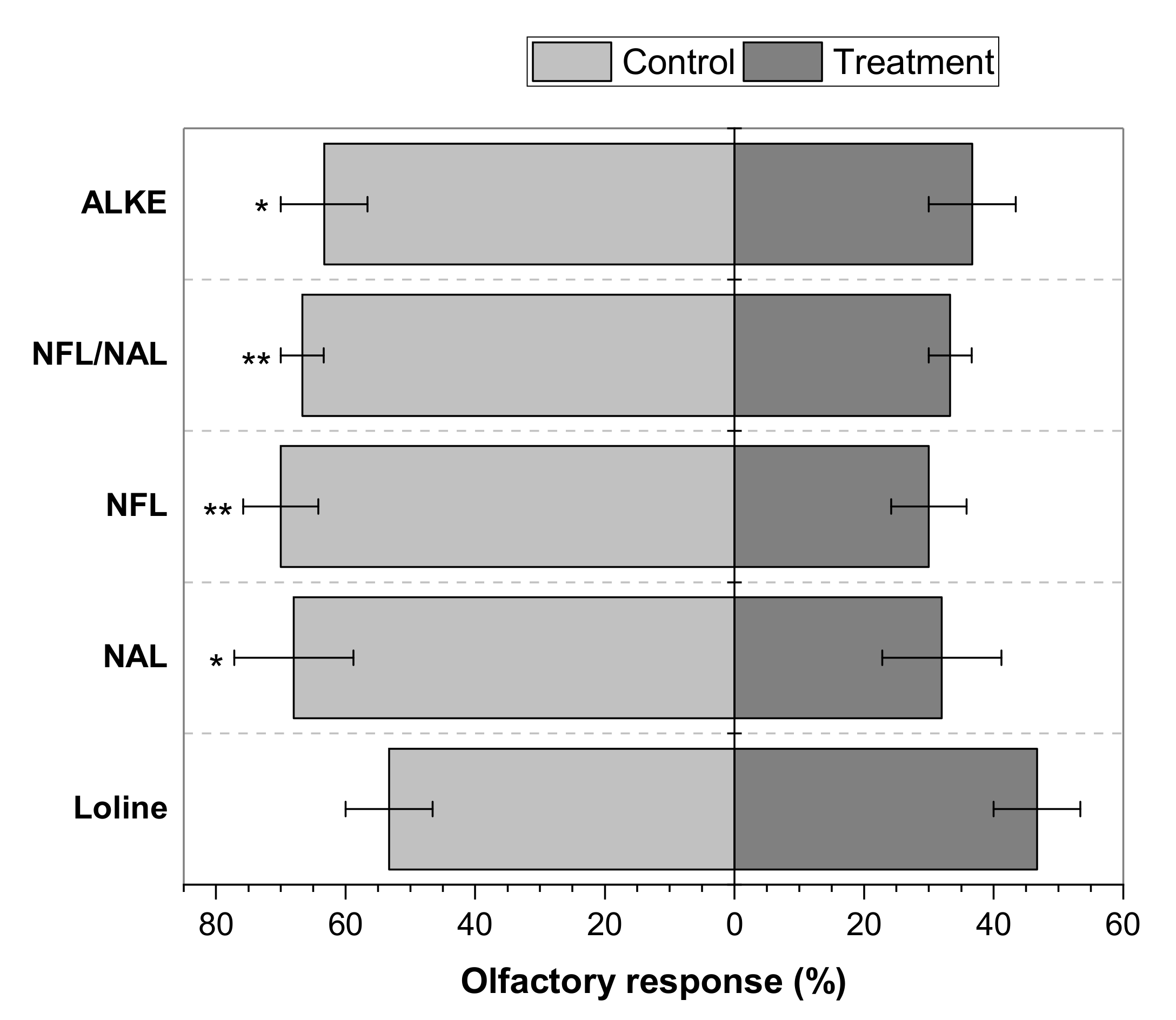
2. Results
3. Discussion
4. Materials and Methods
4.1. H. irritans Collection
4.2. Plant Material
4.3. Alkaloid Extract Obtention
4.4. N-Formyl Loline and N-Acetyl Loline Isolation
4.5. Loline Preparation
4.6. Analyses of Loline Alkaloids by GC/MS
4.7. Laboratory Feeding Bioassay
4.7.1. Blood Collection
4.7.2. Feeding Bioassay
4.8. Olfactometer Bioassays
4.9. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Cupp, E.W.; Cupp, M.S.; Ribeiro, J.M.; Kunz, S.E. Blood-feeding strategy of Haematobia irritans (Diptera: Muscidae). J. Med. Entomol. 1998, 35, 591–595. [Google Scholar] [CrossRef]
- Kuramochi, K. Survival, ovarian development and bloodmeal size for the horn fly Haematobia irritans irritans reared in vitro. Med. Vet. Entomol. 2000, 14, 201–206. [Google Scholar] [CrossRef]
- Mancebo, A.; Monzón, C.M.; Bulman, G.M. Haematobia irritans: Una actualización a diez años de su introducción en Argentina (Parte I). Vet. Argent. 2001, 18, 34–46. [Google Scholar]
- Mancebo, A.; Monzón, C.M.; Bulman, G.M. Haematobia irritans: Una actualización a diez años de su introducción en Argentina (Parte II). Vet. Argent. 2001, 18, 119–135. [Google Scholar]
- Foil, L.D.; Hogsette, J.A. Biology and control of tabanids, stable flies and horn flies. Rev. Sci. Tech. 1994, 13, 1125–1158. [Google Scholar] [CrossRef] [PubMed]
- Quiroz, R.H. Parasitología y Enfermedades Parasitarias de los Animales Domésticos, 3rd ed.; Limusa: Mexico City, Mexico, 2005. [Google Scholar]
- James, M.T.; Harwood, R.F. Herms’ Medical Entomology, 6th ed.; The McMillan Co: New York, NY, USA, 1969; pp. 275–277. [Google Scholar]
- Depner, K.R. The effect of temperature on development and diapause of the horn fly, Siphona irritans (L.) (Diptera: Muscidae). Can. Entomol. 1961, 10, 855–859. [Google Scholar] [CrossRef]
- Carballo, M.; Martínez, M. Hallazgo de Haematobia irritans en Uruguay. Veterinaria 1991, 27, 20–21. [Google Scholar]
- Luzuriaga, R.; Eddi, C.; Caracostantogolo, J.; Botto, E.; Pereira, J. Diagnosis of infestation of cattle by Haematobia irritans (L.) in Misiones, Argentina. Rev. Med. Vet. 1991, 72, 262–264. [Google Scholar]
- Cisternas, E.A. Mosca de los cuernos Haematobia irritans. Inf. INIA Remehue 1999, 11, 1–2. [Google Scholar]
- Fundación para la Innovación Agraria (FIA); BTA Consultores. Resultados y Lecciones en Control. Biológico de la Mosca de los Cuernos en bovinos con Extracto de Neem: Proyecto de Innovación en regiones Metropolitana, de Valparaíso, del Maule y del Biobío; Fundación Para La Innovación Agraria: La Reina, Chile, 2009; pp. 1–36. [Google Scholar]
- Schreiber, E.T.; Campbell, J.B.; Kunz, S.E.; Clanton, D.C.; Hudson, D.B. Effects of horn fly (Diptera: Muscidae) control on cows and gastrointestinal worm (Nematode: Trichostrongylidae) treatment for calves on cow and calf weight gains. J. Econ. Entomol. 1987, 80, 451–454. [Google Scholar] [CrossRef]
- DeRouen, S.M.; Foil, L.D.; MacKay, A.J.; Fraenke, D.E.; Sanson, D.W.; Wyatt, W.E. Effect of horn fly (Haematobia irritans) control on growth and reproduction of beef heifers. J. Econ. Entomol. 2003, 96, 1612–1616. [Google Scholar] [CrossRef]
- Campbell, J.B. Effect of horn fly control on cows as expressed by increased weaning weights of calves. J. Econ. Entomol. 1976, 69, 711–712. [Google Scholar] [CrossRef]
- Harvey, T.L.; Brethour, J.L. Effect of horn flies on weight gains of beef cattle. J. Econ. Entomol. 1979, 72, 516–518. [Google Scholar] [CrossRef] [PubMed]
- Steelman, C.D.; Brown, A.H., Jr.; Gbur, E.E.; Tolley, G. Interactive response of the horn fly (Diptera: Muscidae) and selected breeds of beef cattle. J. Econ. Entomol. 1991, 84, 1275–1282. [Google Scholar] [CrossRef] [PubMed]
- Foil, L.D.; Allison, M.; DeRouen, S.M.; Kimball, M.; Morrison, D.M.; Sanson, D.W.; Wyatt, W.E. Controlling horn flies. La Agric. 2000, 43, 21–23. [Google Scholar]
- Williams, J.D.; Sutherst, R.W.; Maywald, G.F.; Petherbridge, C.T. The southward spread of buffalo fly (Haematobia irritans exigua) in eastern Australia and its survival through a severe winter. Aust. Vet. J. 1985, 62, 367–369. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, R.L.; Metcalf, R.A. Destructive and Useful Insects, 5th ed.; McGraw-Hill: New York, NY, USA, 1993. [Google Scholar]
- Hawkins, J.A. The Horn Fly (Haematobia irritans); Merial Veterinary Bulletin No TSB-0-00028-FTB; Veterinary Professional Services, Merial: Iselin, NJ, USA, 2001. [Google Scholar]
- DeRouen, S.M.; Miller, J.E.; Foil, L.D. Control of horn flies (Haematobia irritans) and gastrointestinal nematodes and its relation with growth performance in stocker cattle. Prof. Anim. Sci. 2010, 26, 109–114. [Google Scholar] [CrossRef]
- Grisi, L.; Cerqueira, R.; de Souza, J.; Medeiros, A.; Andreotti, R.; Duarte, P.; Pérez, A.; Barros, J.; Silva, H. Reassessment of the potential economic impact of cattle parasites in Brazil. Braz. J. Vet. Parasitol. 2014, 23, 150–156. [Google Scholar] [CrossRef] [Green Version]
- Velasco, R.; González, J.; Morales, G.; Ortega, E. Daño económico y costos de control en bovinos mosca de los cuernos. Inf. Agropecu. Bioleche INIA Quilamapu 2001, 14, 4–7. [Google Scholar]
- Lanuza, F.; Paredes, E.; Sievers, G.; Cortázar, J.M. Manejo sanitario y principales enfermedades de los bovinos de carne. In Producción y Manejo de Carne Bovina en Chile; Catrileo, A., Ed.; Instituto de Investigaciones Agropecuarias: Temuco, Chile, 2005; pp. 513–550. [Google Scholar]
- Guglielmone, A.A.; Castelli, M.E.; Idiart, J.; Fisher, W.F.; Volpogni, M.M.; Quaino, O.; Anziani, O.S.; Flores, S.G.; Warnke, O. Skin lesions and cattle hide damage from Haematobia irritans in cattle. Med. Vet. Entomol. 1999, 13, 323–328. [Google Scholar] [CrossRef]
- Barros, A.T.M.; Ottea, J.; Sanson, D.; Foil, L.D. Horn fly (Diptera: Muscidae) resistance to organophosphate insecticides. Vet. Parasitol. 2001, 96, 243–256. [Google Scholar] [CrossRef]
- Oyarzún, M.P.; Quiroz, A.; Birkett, M.A. Insecticide resistance in the horn fly: Alternative control strategies. Med. Vet. Entomol. 2008, 22, 188–202. [Google Scholar] [CrossRef]
- Oyarzún, M.P.; Li, Y.; Figueroa, C.C. High levels of insecticide resistance in introduced horn fly (Diptera: Muscidae) populations and implications for management. J. Econ. Entomol. 2011, 104, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Parra, L.; Rojas, C.; Catrileo, A.; Galdames, R.; Mutis, A.; Birkett, M.A.; Quiroz, A. Differences in the fly-load of Haematobia irritans (Diptera: Muscidae) on cattle is modified by endophyte infection of pastures. Electron. J. Biotechnol. 2013, 16, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Parra, L.; Mutis, A.; Chacón, M.; Lizama, M.; Rojas, C.; Catrileo, A.; Rubilar, O.; Tortella, G.; Birkett, M.A.; Quiroz, A. Horn fly larval survival in cattle dung is reduced by endophyte infection of tall fescue pasture. Pest. Manag. Sci. 2016, 72, 1328–1334. [Google Scholar] [CrossRef]
- Hoveland, C.S. Economic importance of Acremonium endophytes. Agric. Ecosyst. Environ. 1993, 44, 3–14. [Google Scholar] [CrossRef]
- Schardl, C.L.; Leuchtmann, A.; Spiering, M.J. Symbioses of grasses with seedborne fungal endophytes. Annu Rev. Plant. Biol. 2004, 55, 315–340. [Google Scholar] [CrossRef]
- Potter, D.A.; Tyler, J.T.; Redmond, C.T.; Schardl, C.L.; Panaccione, D.G. Contribution of ergot alkaloids to suppression of a grassfeeding caterpillar assessed with gene knockout endophytes in perennial ryegrass. Entomol. Exp. Appl. 2008, 126, 138–147. [Google Scholar] [CrossRef]
- Panaccione, D.; Beaulieu, W.; Cook, D. Bioactive alkaloids in vertically transmitted fungal endophytes. Funct. Ecol. 2014, 28, 299–314. [Google Scholar] [CrossRef] [Green Version]
- Schardl, C.L.; Young, C.A.; Faulkner, J.R.; Florea, S.; Pan, J. Chemotypic diversity of epichloae, fungal symbionts of grasses. Fungal Ecol. 2011, 5, 331–344. [Google Scholar] [CrossRef]
- Showler, A.T. Botanically based repellent and insecticidal effects against horn flies and stable flies (Diptera: Muscidae). J. Integr. Pest. Manag. 2017, 15, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Dougherty, C.T.; Knapp, F.W.; Bush, L.P.; Maul, J.E.; van Willigen, J. Mortality of horn fly (Diptera: Muscidae) larvae in bovine dung supplemented with loline alkaloids from tall fescue. J. Med. Entomol. 1998, 35, 798–803. [Google Scholar] [CrossRef]
- Dougherty, C.T.; Knapp, F.W.; Bush, L.P. Mortality of horn fly larvae (Diptera: Muscidae) in bovine dung supplemented with ergotamine and N-formyl loline. J. Med. Entomol. 1999, 36, 73–77. [Google Scholar] [CrossRef]
- Faeth, S.H. Are endophytic fungi defensive plant mutualists? Oikos 2002, 98, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Nelli, M.R.; Scheerer, J.R. Synthesis of Peramine, an Anti-insect Defensive Alkaloid Produced by Endophytic Fungi of Cool Season Grasses. J. Nat. Prod. 2016, 79, 1189–1192. [Google Scholar] [CrossRef] [PubMed]
- Schardl, C.L.; Grossman, R.B.; Nagabhyru, P.; Faulkner, J.R.; Mallik, U.P. Loline alkaloids: Currencies of mutualism. Phytochemistry 2007, 68, 980–996. [Google Scholar] [CrossRef] [PubMed]
- Popay, A. Insect herbivory and defensive mutualisms between plants and fungi. In Defensive Mutualism in Microbial Symbiosis; White, J., Jr., Torres, M., Eds.; CRC: Boca Raton, FL, USA, 2009; pp. 347–366. [Google Scholar]
- Finch, S.; Munday, J.; Munday, R.; Kerby, J. Short-term toxicity studies of loline alkaloids in mice. Food Chem. Toxicol. 2016, 94, 243–249. [Google Scholar] [CrossRef]
- Yates, S.G.; Fenster, J.C.; Bartelt, R.J. Assay of tall fescue seed extracts, fractions, and alkaloids using the large milkweed bug. J. Agric. Food Chem. 1989, 37, 354–357. [Google Scholar] [CrossRef]
- Siegel, M.R.; Latch, G.C.M.; Bush, L.P.; Fannin, F.F.; Rowan, D.D.; Tapper, B.A.; Bacon, C.W.; Johnson, M.C. Fungal endophyte-infected grasses: Alkaloid accumulation and aphid response. J. Chem. Ecol. 1990, 16, 3301–3315. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, H.H.; Siegel, M.R.; Blankenship, J.D.; Mallory, A.C.; Bush, L.P.; Schardl, C.L. Contribution of fungal loline alkaloids to protection from aphids in a grass-endophyte mutualism. Mol. Plant. Microbe. Interact. 2000, 13, 1027–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riedell, W.E.; Kieckhefer, R.E.; Petroski, R.J.; Powell, R.G. Naturally occurring and synthetic loline alkaloid derivatives: Insect feeding behavior modification and toxicity. J. Entomol. Sci. 1991, 26, 122–129. [Google Scholar] [CrossRef]
- Lehtonen, P.; Helander, M.; Wink, M.; Sporer, F.; Saikkonen, K. Transfer of endophyte-origin defensive alkaloids from a grass to a hemiparasitic plant. Ecol. Lett. 2005, 8, 1256–1263. [Google Scholar] [CrossRef]
- Patterson, C.G.; Potter, D.A.; Fannin, F.F. Feeding deterrency of alkaloids from endophyte-infected grasses to Japanese beetle grubs. Entomol. Exp. Appl. 1991, 61, 285–289. [Google Scholar] [CrossRef]
- Bultman, T.L.; Borowicz, K.L.; Schneble, R.M.; Coudron, T.A.; Bush, L.P. Effect of a fungal endophyte on the growth and survival of two Euplectrus parasitoids. Oikos 1997, 78, 170–176. [Google Scholar] [CrossRef]
- Westendorf, M.L.; Mitchell, G.E.; Tucker, R.E., Jr.; Bush, L.P.; Petroski, R.J.; Powell, R.G. In Vitro and in Vivo Ruminal and Physiological Responses to Endophyte-Infected Tall Fescue. J. Dairy Sci. 1993, 76, 555–563. [Google Scholar] [CrossRef]
- Espinoza, J.; Ungerfeld, E.; Escobar, P.; Piñeira, J.; Mejías, J.; Quiroz, A. Loline alkaloids presence in blood serum, urine and feces of steers fed with tall fescue (Festuca arundinacea) infected by endophytic fungus Neotyphodium coenophialum. In 6th International Workshop in Advances in Science and Technology of Bioresources, Pucón, Chile, 29–30 November–01 December 2017; Reyes, M., Luengo, A., Eds.; Universidad de La Frontera: Temuco, Chile, 2017; p. 94. [Google Scholar]
- Showler, A.T.; Harlien, J.L. Effects of the botanical compound p-anisaldehyde on horn fly (Diptera: Muscidae) repellency, mortality, and reproduction. J. Med. Entomol. 2018, 55, 183–192. [Google Scholar] [CrossRef] [Green Version]
- Showler, A.T.; Harlien, J.L.; Perez de Léon, A.A. Effects of Laboratory Grade Limonene and a Commercial Limonene-Based Insecticide on Haematobia irritans irritans (Muscidae: Diptera): Deterrence, Mortality, and Reproduction. J. Med. Entomol. 2019, 56, 1064–1070. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.J.; Brewer, G.J.; Boxler, D.J.; Friesen, K.; Taylor, D.B. Comparisons of antifeedancy and spatial repellency of three natural product repellents against horn flies, Haematobia irritans (Diptera:Muscidae). Pest. Manag. Sci. 2015, 71, 1553–1560. [Google Scholar] [CrossRef] [Green Version]
- Mullens, B.A.; Reifenrath, W.G.; Butler, S.M. Laboratory trials of fatty acids as repellents or antifeedants against house flies, horn flies and stable flies (Diptera: Muscidae). Pest. Manag. Sci. 2009, 65, 1360–1366. [Google Scholar] [CrossRef]
- Oyarzún, M.P.; Palma, R.; Alberti, E.; Hormazabal, H.; Pardo, F.; Birkett, M.A.; Quiroz, A. Olfactory Response of Haematobia irritans (Diptera: Muscidae) ton Cattle-Derived Volatile Compounds. Med. Entomol. 2009, 46, 1320–1326. [Google Scholar] [CrossRef] [Green Version]
- Klauck, V.; Pazinato, R.; Radavelli, W.M.; Volpato, A.; Stefani, L.M.; Santos, R.C.; Vaucher, R.A.; Boligon, A.A.; Athayde, M.S.; da Silva, A.S. In vitro repellent effect of tea tree (Melaleuca alternifolia) and andiroba (Carapa guianensis) oils on Haematobia irritans and Chrysomya megacephala flies. Trop. Biomed. 2015, 32, 160–166. [Google Scholar]
- Klauck, V.; Pazinato, R.; Stefani, L.M.; Santos, R.C.; Vaucher, R.A.; Baldissera, M.D.; Raffin, R.; Boligon, A.; Athayde, M.; Baretta, D.; et al. Insecticidal and repellent effects of tea tree and andiroba oils on flies associated with livestock. Med. Vet. Entomol. 2014, 28, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Marlatt, C.L. Report of the Entomologist; Technical Report No. 1927-28; USDA: Washington, DC, USA, 1928. [Google Scholar]
- Stanbury, R.E.; Goodhue, L.D. New fly repellents in dairy sprays. Agric. Chem. 1960, 15, 43–44. [Google Scholar]
- Heimerdinger, A.; Olivo, C.J.; Sobczak, M.F.; Gabbi, A.M.; Scaravelli, L.F.B.; Pereira, L.E.T.; Piuco, M.A. Utilization of Phytotherapetic Products on the Control of Haematobia Irritans in Dairy Cows Contaminated Naturally; International Society for Animal Hygiene: Saint Malo, France, 2004; p. 255. [Google Scholar]
- Milián, C.A.F. Evaluación del Efecto Repelente del té de Limón (Cymbopogon citratus) Utilizado a Diferentes Diluciones Contra la Mosca los Cuernos (Haematobia irritans) en Ganado Bovino de Doble Propósito Durante el Mes de Entere de 2008 en Salamá, Baja Verapaz. Licentiate Thesis, Universidad de San Carlos de Guatemala, Excuela de Medicina Veterinaria, Antigua, Guatemala, August 2009. [Google Scholar]
- Ramirez, A.M.; Carillo, A.C.; Molano, C.R. Preliminary screening of ethanolic extracts of five medicinal plants against Haematobia irritans L. (Diptera: Muscidae). Rev. UDCA Actual. Divulg. Cient. 2009, 12, 69–78. [Google Scholar] [CrossRef]
- Juan, L.W.; Lucia, A.; Zerba, E.N.; Harrand, L.; Marco, M.; Masuh, H.M. Chemical composition and fumigant toxicity of the essential oils from 16 species of Eucalyptus against Haematobia irritans (Diptera: Muscidae) adults. J. Econ. Entomol. 2011, 104, 1087–1092. [Google Scholar] [CrossRef]
- Lachance, S.; Grange, F. Repellency effectiveness of seven plant essential oils, sunflower oil and natural insecticides against horn flies on pastured dairy cows and heifers. Med. Vet. Entomol. 2014, 28, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Mullens, B.A.; Watson, D.W.; Gerry, A.C.; Sandelin, B.A.; Soto, D.; Rawls, D.; Denning, S.; Guisewite, L.; Cammack, J. Field trials of fatty acids and geraniol applied to cattle for suppression of horn flies, Haematobia irritans (Diptera: Muscidae), with observations on fly defensive behaviors. Vet. Parasitol. 2017, 245, 14–28. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.W.; Strickland, J.R.; Waller, J.C.; Fribourg, H.A.; Linnabary, R.D.; Abney, L.K. Endophytic fungal toxin effect on adrenergic receptors in lateral saphenous veins (cranial branch) of cattle grazing tall fescue. J. Anim. Sci. 1998, 76, 2853–2856. [Google Scholar] [CrossRef]
- Solomons, R.N.; Oliver, J.W.; Linnabary, R.D. Reactivity of dorsal pedal vein of cattle to selected alkaloids associated with Acremonium coenophialum-infected fescue grass. Am. J. Vet. Res. 1989, 50, 235–238. [Google Scholar] [PubMed]
- Rudolph, W.; Remane, D.; Wissenbach, D.K.; Peters, F.T. Liquid chromatography-mass spectrometry-based determination of ergocristine, ergocryptine, ergotamine, ergovaline, hypoglycin A, lolitrem B, methylene cyclopropyl acetic acid carnitine, N-acetylloline, N-formylloline, paxilline, and peramine in equine hair. J. Chromatogr. B 2019, 1117, 127–135. [Google Scholar] [CrossRef]
- Belanger, F.C. A rapid seedling screening method for determination of fungal endophyte viability. Crop. Sci. 1996, 36, 460–462. [Google Scholar] [CrossRef]
- Saha, D.C.; Jackson, M.A.; Johnson-Cicalese, J.M. A rapid staining method for detection of endophytic fungi in turf and forage grasses. Phytopathology 1988, 78, 237–239. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Espinoza, J.; Chacón-Fuentes, M.; Quiroz, A.; Bardehle, L.; Escobar-Bahamondes, P.; Ungerfeld, E. Antifeedant Effects and Repellent Activity of Loline Alkaloids from Endophyte-Infected Tall Fescue against Horn Flies, Haematobia irritans (Diptera: Muscidae). Molecules 2021, 26, 817. https://doi.org/10.3390/molecules26040817
Espinoza J, Chacón-Fuentes M, Quiroz A, Bardehle L, Escobar-Bahamondes P, Ungerfeld E. Antifeedant Effects and Repellent Activity of Loline Alkaloids from Endophyte-Infected Tall Fescue against Horn Flies, Haematobia irritans (Diptera: Muscidae). Molecules. 2021; 26(4):817. https://doi.org/10.3390/molecules26040817
Chicago/Turabian StyleEspinoza, Javier, Manuel Chacón-Fuentes, Andrés Quiroz, Leonardo Bardehle, Paul Escobar-Bahamondes, and Emilio Ungerfeld. 2021. "Antifeedant Effects and Repellent Activity of Loline Alkaloids from Endophyte-Infected Tall Fescue against Horn Flies, Haematobia irritans (Diptera: Muscidae)" Molecules 26, no. 4: 817. https://doi.org/10.3390/molecules26040817
APA StyleEspinoza, J., Chacón-Fuentes, M., Quiroz, A., Bardehle, L., Escobar-Bahamondes, P., & Ungerfeld, E. (2021). Antifeedant Effects and Repellent Activity of Loline Alkaloids from Endophyte-Infected Tall Fescue against Horn Flies, Haematobia irritans (Diptera: Muscidae). Molecules, 26(4), 817. https://doi.org/10.3390/molecules26040817