
Dual Effect of Taxifolin on ZEB2 Cancer Signaling in HepG2 Cells


Abstract
:1. Introduction
2. Results
2.1. Quercetin and Taxifolin Are Not Toxic in Hep G2 Cells
2.2. Analysis of miRNAs Expression Profile in Affymetrix GeneChip™ miRNA 3.0 Arrays
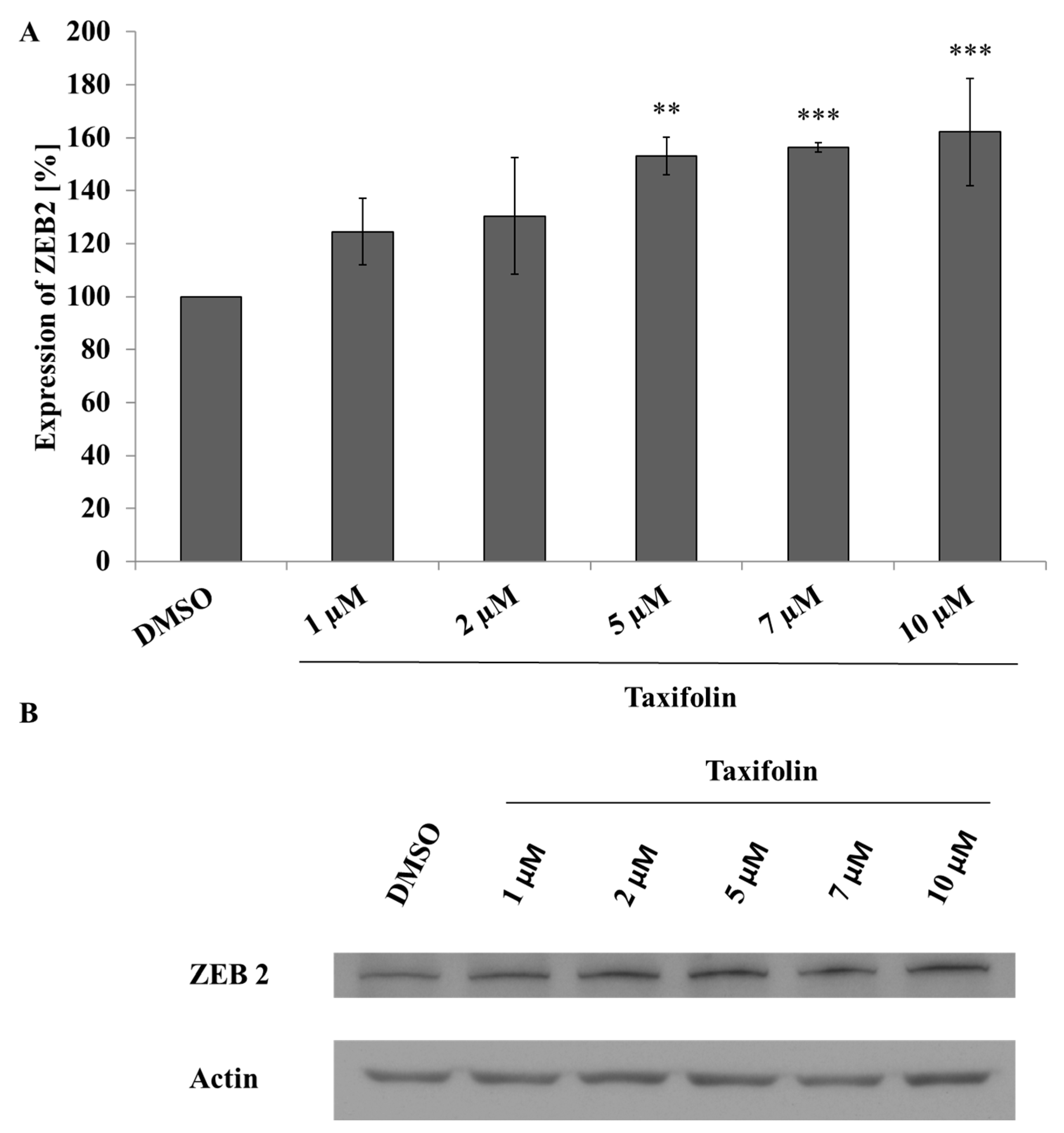
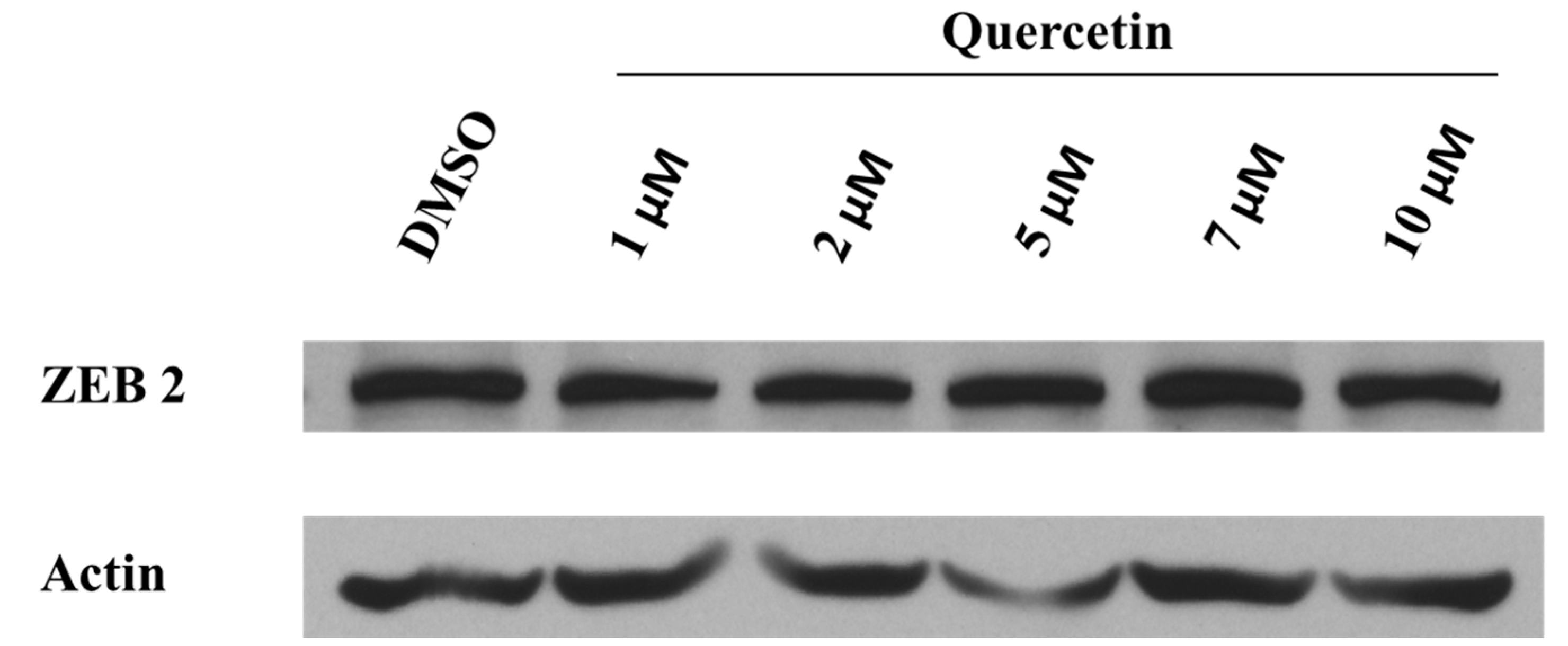
2.3. Taxifolin Modulates ZEB2 Expression in Hep G2 Cells But Not in Primary Cultures of Human Hepatocytes
2.4. ZEB2 Upregulation Did Not Cause the Epithelial to Mesenchymal Transition
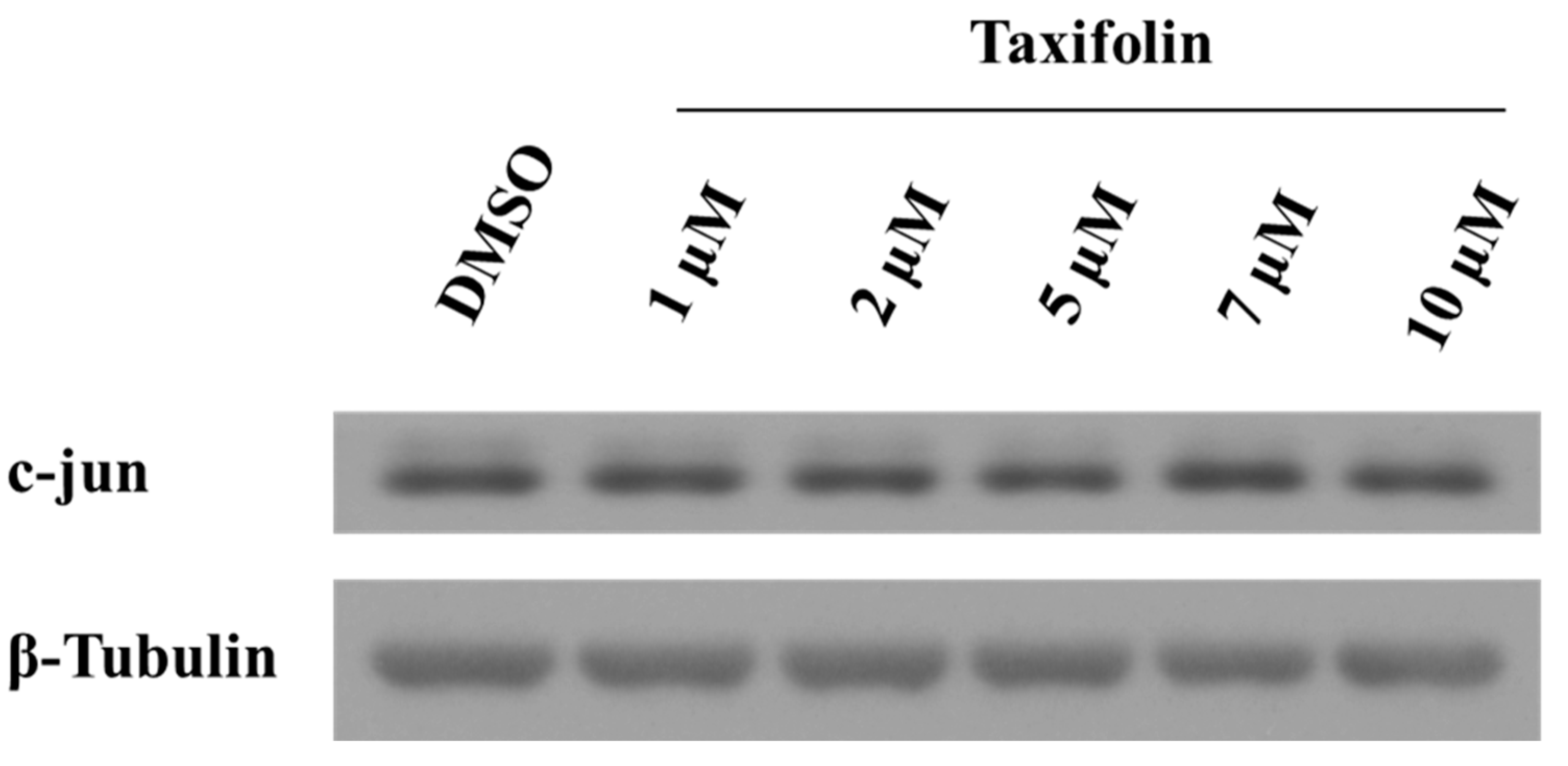
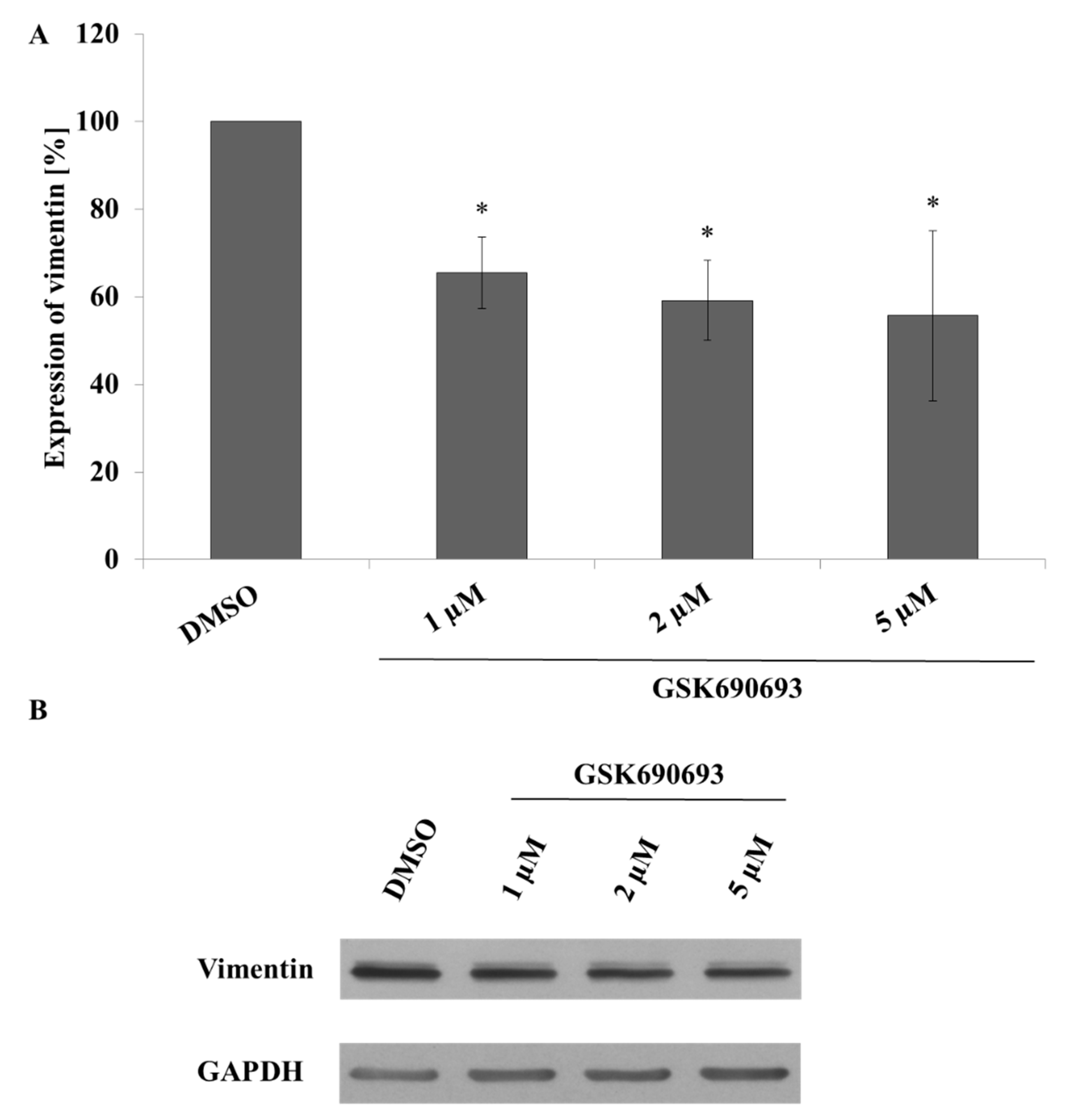
2.5. Taxifolin Negatively Regulates Vimentin Expression via Akt Dephosphorylation
2.6. Reduction of ZEB2 Expression by miR-377 Precursors Is Restored by Taxifolin in a Dose-Dependent Manner
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Culture Conditions
4.3. Primary Cultures of Human Hepatocytes
4.4. Cell Viability Assays
4.5. xCELLigence System
4.6. Bright-Field Microscopy
4.7. RNA Isolation
4.8. miRNA Array Analysis
4.9. RT-PCR
4.10. Immunodetection
4.11. Transfection
4.12. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | Mean Relative Expression Compared to Control | References | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Quercetin | Taxifolin | ||||||||
| Hep G2 | Human Hepatocytes | Hep G2 | Human Hepatocytes | ||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | ||
| hsa-miR-30a-5p | 0.92 | 0.05 | 0.95 | 0.05 | 0.89 | 0.24 | 0.95 | 0.09 | [27] |
| hsa-miR-30a-3p | 0.85 | 0.24 | 1.51 | 0.99 | 0.87 | 0.33 | 1.26 | 0.88 | [27] |
| hsa-miR-101-3p | 0.94 | 0.46 | 1.04 | 0.35 | 1.15 | 0.80 | 1.22 | 0.55 | [30] |
| hsa-miR-129-5p | 2.42 | 2.11 | 0.94 | 0.27 | 4.53 | 6.58 | 1.07 | 0.17 | [31] |
| hsa-miR-132-3p | 0.99 | 0.01 | 0.82 | 0.37 | 1.01 | 0.05 | 1.23 | 0.25 | [64] |
| hsa-miR-138-5p | 1.07 | 0.16 | 0.89 | 0.24 | 1.12 | 0.16 | 0.91 | 0.34 | [30] |
| hsa-miR-138-1-3p | 1.00 | 0.04 | 0.95 | 0.29 | 0.99 | 0.03 | 1.12 | 0.52 | [30] |
| hsa-miR-139-5p | 1.40 | 0.16 | 1.35 | 0.86 | 2.14 | 1.06 | 1.34 | 1.09 | [32] |
| hsa-miR-141-3p | 1.90 | 0.96 | 0.98 | 0.17 | 1.56 | 1.21 | 1.00 | 0.42 | [30] |
| hsa-miR-144-3p | 0.92 | 0.20 | 1.02 | 0.36 | 0.99 | 0.24 | 1.30 | 0.29 | [30] |
| hsa-miR-145-5p | 1.01 | 0.01 | 0.92 | 0.34 | 1.01 | 0.02 | 1.00 | 0.41 | [65] |
| hsa-miR-153 | 1.02 | 0.37 | 0.72 | 0.24 | 0.67 | 0.27 | 1.39 | 0.89 | [30] |
| hsa-miR-154-5p | 1.40 | 0.44 | 1.73 | 1.09 | 1.63 | 0.48 | 1.45 | 0.60 | [30] |
| hsa-miR-192-5p | 1.00 | 0.02 | 0.99 | 0.01 | 1.00 | 0.02 | 1.01 | 0.03 | [66] |
| hsa-miR-200a-3p | 0.99 | 0.03 | 0.90 | 0.27 | 1.02 | 0.03 | 1.37 | 0.36 | [30] |
| hsa-miR-200b-3p | 1.01 | 0.02 | 1.03 | 0.49 | 0.99 | 0.05 | 0.98 | 0.19 | [30] |
| hsa-miR-200c-3p | 1.00 | 0.00 | 1.59 | 0.47 | 1.00 | 0.04 | 1.85 | 0.70 | [30] |
| hsa-miR-203 | 0.89 | 0.73 | 0.81 | 0.37 | 1.10 | 0.94 | 0.92 | 0.48 | [30] |
| hsa-miR-204 | 0.79 | 0.55 | 0.78 | 0.03 | 0.68 | 0.32 | 0.86 | 0.38 | [33] |
| hsa-miR-205-5p | 1.12 | 0.21 | 1.08 | 0.61 | 1.27 | 0.27 | 1.07 | 0.44 | [30] |
| hsa-miR-211 | 1.17 | 0.72 | 0.91 | 0.42 | 0.59 | 0.20 | 0.74 | 0.33 | [34] |
| hsa-miR-215 | 1.12 | 0.24 | 0.84 | 0.18 | 1.07 | 0.28 | 1.20 | 0.55 | [30] |
| hsa-miR-221-3p | 1.00 | 0.01 | 1.04 | 0.06 | 0.99 | 0.01 | 1.03 | 0.11 | [30] |
| hsa-miR-335-5p | 1.47 | 1.49 | 0.77 | 0.20 | 2.10 | 2.98 | 0.84 | 0.14 | [30] |
| hsa-miR-338-3p | 1.40 | 0.62 | 0.66 | 0.15 | 0.91 | 0.24 | 0.85 | 0.22 | [30] |
| hsa-miR-377-3p | 1.42 | 1.16 | 0.65 | 0.19 | 0.64 | 0.43 | 0.57 | 0.05 | [35] |
| hsa-miR-429 | 0.92 | 0.36 | 0.94 | 0.18 | 1.05 | 0.60 | 0.96 | 0.16 | [30] |
| hsa-miR-590-3p | 1.16 | 0.54 | 1.15 | 0.31 | 2.09 | 0.81 | 0.95 | 0.08 | [30] |
| hsa-miR-708-5p | 0.91 | 0.32 | 1.49 | 0.47 | 1.36 | 0.58 | 1.01 | 0.37 | [30] |
| hsa-miR-4782-3p | 1.47 | 0.55 | 0.75 | 0.21 | 1.73 | 0.96 | 1.06 | 0.37 | [30] |






References
- Marin, L.; Miguelez, E.M.; Villar, C.J.; Lombo, F. Bioavailability of dietary polyphenols and gut microbiota metabolism: Antimicrobial properties. Biomed. Res. Int. 2015, 2015, 905215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grosso, G.; Stepaniak, U.; Topor-Madry, R.; Szafraniec, K.; Pajak, A. Estimated dietary intake and major food sources of polyphenols in the Polish arm of the HAPIEE study. Nutrition 2014, 30, 1398–1403. [Google Scholar] [CrossRef] [Green Version]
- Ovaskainen, M.L.; Torronen, R.; Koponen, J.M.; Sinkko, H.; Hellstrom, J.; Reinivuo, H.; Mattila, P. Dietary intake and major food sources of polyphenols in Finnish adults. J. Nutr. 2008, 138, 562–566. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxid. Med. Cell Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasouli, H.; Farzaei, M.H.; Khodarahmi, R. Polyphenols and their benefits: A review. Int. J. Food Prop. 2017, 20, 1700–1741. [Google Scholar] [CrossRef] [Green Version]
- Tresserra-Rimbau, A.; Lamuela-Raventos, R.M.; Moreno, J.J. Polyphenols, food and pharma. Current knowledge and directions for future research. Biochem. Pharmacol. 2018, 156, 186–195. [Google Scholar] [CrossRef]
- Coppo, E.; Marchese, A. Antibacterial activity of polyphenols. Curr. Pharm. Biotechnol. 2014, 15, 380–390. [Google Scholar] [CrossRef]
- Gabrielova, E.; Kren, V.; Jaburek, M.; Modriansky, M. Silymarin component 2,3-dehydrosilybin attenuates cardiomyocyte damage following hypoxia/reoxygenation by limiting oxidative stress. Physiol. Res. 2015, 64, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Sunil, C.; Xu, B.J. An insight into the health-promoting effects of taxifolin (dihydroquercetin). Phytochemistry 2019, 166. [Google Scholar] [CrossRef]
- Milenkovic, D.; Deval, C.; Gouranton, E.; Landrier, J.F.; Scalbert, A.; Morand, C.; Mazur, A. Modulation of miRNA expression by dietary polyphenols in apoE deficient mice: A new mechanism of the action of polyphenols. PLoS ONE 2012, 7, e29837. [Google Scholar] [CrossRef]
- Del Follo-Martinez, A.; Banerjee, N.; Li, X.; Safe, S.; Mertens-Talcott, S. Resveratrol and quercetin in combination have anticancer activity in colon cancer cells and repress oncogenic microRNA-27a. Nutr. Cancer 2013, 65, 494–504. [Google Scholar] [CrossRef]
- Dostal, Z.; Modriansky, M. The effect of quercetin on microRNA expression: A critical review. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czech Repub. 2019, 163, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Gavrilas, L.I.; Ionescu, C.; Tudoran, O.; Lisencu, C.; Balacescu, O.; Miere, D. The Role of Bioactive Dietary Components in Modulating miRNA Expression in Colorectal Cancer. Nutrients 2016, 8, 590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V.; The, C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef]
- Griffiths-Jones, S.; Saini, H.K.; van Dongen, S.; Enright, A.J. miRBase: Tools for microRNA genomics. Nucleic Acids Res. 2008, 36, D154–D158. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, Y.; Akiyama, Y.; Yuasa, Y. Multiple-to-Multiple Relationships between MicroRNAs and Target Genes in Gastric Cancer. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.H.; Pan, X.P.; Cobb, G.P.; Anderson, T.A. microRNAs as oncogenes and tumor suppressors. Dev. Biol. 2007, 302, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Gebeshuber, C.A.; Zatloukal, K.; Martinez, J. miR-29a suppresses tristetraprolin, which is a regulator of epithelial polarity and metastasis. EMBO Rep. 2009, 10, 400–405. [Google Scholar] [CrossRef]
- Jiang, H.; Zhang, G.; Wu, J.H.; Jiang, C.P. Diverse roles of miR-29 in cancer (review). Oncol. Rep. 2014, 31, 1509–1516. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, D.M.; Medici, D. Signaling mechanisms of the epithelial-mesenchymal transition. Sci. Signal. 2014, 7, re8. [Google Scholar] [CrossRef] [Green Version]
- Vandewalle, C.; Van Roy, F.; Berx, G. The role of the ZEB family of transcription factors in development and disease. Cell Mol. Life Sci. 2009, 66, 773–787. [Google Scholar] [CrossRef]
- Nieto, M.A.; Huang, R.Y.; Jackson, R.A.; Thiery, J.P. Emt: 2016. Cell 2016, 166, 21–45. [Google Scholar] [CrossRef] [Green Version]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Tang, Z.Y.; He, Y.; Liu, L.F.; Li, D.J.; Chen, X. miRNA-205 Is a Candidate Tumor Suppressor that Targets ZEB2 in Renal Cell Carcinoma. Oncol. Res. Treat. 2014, 37, 658–664. [Google Scholar] [CrossRef] [PubMed]
- Li, L.N.; Li, W.L. Epithelial-mesenchymal transition in human cancer: Comprehensive reprogramming of metabolism, epigenetics, and differentiation. Pharmacol. Ther. 2015, 150, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Di Gennaro, A.; Damiano, V.; Brisotto, G.; Armellin, M.; Perin, T.; Zucchetto, A.; Guardascione, M.; Spaink, H.P.; Doglioni, C.; Snaar-Jagalska, B.E.; et al. A p53/miR-30a/ZEB2 axis controls triple negative breast cancer aggressiveness. Cell Death Differ. 2018. [Google Scholar] [CrossRef]
- Long, Z.H.; Bai, Z.G.; Song, J.N.; Zheng, Z.; Li, J.; Zhang, J.; Cai, J.; Yao, H.W.; Wang, J.; Yang, Y.C.; et al. miR-141 Inhibits Proliferation and Migration of Colorectal Cancer SW480 Cells. Anticancer Res. 2017, 37, 4345–4352. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Zhang, Z.; Liu, Z.; Qiu, B.; Liu, K.; Dong, G. MicroRNA-335 inhibits invasion and metastasis of colorectal cancer by targeting ZEB2. Med. Oncol. 2014, 31, 982. [Google Scholar] [CrossRef]
- Chou, C.H.; Shrestha, S.; Yang, C.D.; Chang, N.W.; Lin, Y.L.; Liao, K.W.; Huang, W.C.; Sun, T.H.; Tu, S.J.; Lee, W.H.; et al. miRTarBase update 2018: A resource for experimentally validated microRNA-target interactions. Nucleic Acids Res. 2018, 46, D296–D302. [Google Scholar] [CrossRef]
- Xiao, L.; Zhou, X.; Liu, F.; Hu, C.; Zhu, X.; Luo, Y.; Wang, M.; Xu, X.; Yang, S.; Kanwar, Y.S.; et al. MicroRNA-129-5p modulates epithelial-to-mesenchymal transition by targeting SIP1 and SOX4 during peritoneal dialysis. Lab. Investig. 2015, 95, 817–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, S.; Wang, L.; Zhang, H.; Min, Y.; Lou, Y.; Sun, H.; Jiang, Y.; Zhang, W.; Liang, A.; Guo, Y.; et al. miR-139-5p suppresses cancer cell migration and invasion through targeting ZEB1 and ZEB2 in GBM. Tumour Biol. 2015, 36, 6741–6749. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Zhou, Y.J.; Yang, Z.C.; Chen, B.Y.; Huang, W.N.; Liu, Y.Y.; Zhang, Y. MiR-204/ZEB2 axis functions as key mediator for MALAT1-induced epithelial-mesenchymal transition in breast cancer. Tumor Biol. 2017, 39. [Google Scholar] [CrossRef]
- Jiang, G.B.; Wen, L.; Deng, W.P.; Jian, Z.Y.; Zheng, H.M. Regulatory role of miR-211-5p in hepatocellular carcinoma metastasis by targeting ZEB2. Biomed. Pharmacother. 2017, 90, 806–812. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Hu, Y.; Wang, J. MicroRNA-377 targets zinc finger E-box-binding homeobox 2 to inhibit cell proliferation and invasion of cervical cancer. Oncol. Res. 2018. [Google Scholar] [CrossRef]
- Vandewalle, C.; Comijn, J.; De Craene, B.; Vermassen, P.; Bruyneel, E.; Andersen, H.; Tulchinsky, E.; Van Roy, F.; Berx, G. SIP1/ZEB2 induces EMT by repressing genes of different epithelial cell-cell junctions. Nucleic Acids Res. 2005, 33, 6566–6578. [Google Scholar] [CrossRef]
- Alam, M.N.; Almoyad, M.; Huq, F. Polyphenols in Colorectal Cancer: Current State of Knowledge including Clinical Trials and Molecular Mechanism of Action. Biomed. Res. Int. 2018, 2018, 4154185. [Google Scholar] [CrossRef] [Green Version]
- Bindels, S.; Mestdagt, M.; Vandewalle, C.; Jacobs, N.; Volders, L.; Noel, A.; van Roy, F.; Berx, G.; Foidart, J.M.; Gilles, C. Regulation of vimentin by SIP1 in human epithelial breast tumor cells. Oncogene 2006, 25, 4975–4985. [Google Scholar] [CrossRef] [Green Version]
- Du, K.; Montminy, M. CREB is a regulatory target for the protein kinase Akt/PKB. J. Biol. Chem. 1998, 273, 32377–32379. [Google Scholar] [CrossRef] [Green Version]
- Conquer, J.A.; Maiani, G.; Azzini, E.; Raguzzini, A.; Holub, B.J. Supplementation with quercetin markedly increases plasma quercetin concentration without effect on selected risk factors for heart disease in healthy subjects. J. Nutr. 1998, 128, 593–597. [Google Scholar] [CrossRef]
- Graefe, E.U.; Wittig, J.; Mueller, S.; Riethling, A.K.; Uehleke, B.; Drewelow, B.; Pforte, H.; Jacobasch, G.; Derendorf, H.; Veit, M. Pharmacokinetics and bioavailability of quercetin glycosides in humans. J. Clin. Pharmacol. 2001, 41, 492–499. [Google Scholar] [CrossRef]
- Russo, M.; Spagnuolo, C.; Tedesco, I.; Bilotto, S.; Russo, G.L. The flavonoid quercetin in disease prevention and therapy: Facts and fancies. Biochem. Pharmacol. 2012, 83, 6–15. [Google Scholar] [CrossRef]
- Rajnochova Svobodova, A.; Rysava, A.; Psotova, M.; Kosina, P.; Zalesak, B.; Ulrichova, J.; Vostalova, J. The Phototoxic Potential of the Flavonoids, Taxifolin and Quercetin. Photochem. Photobiol. 2017, 93, 1240–1247. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, R.A.; Delic, S.; Herrmann, H.; Snider, N.T. Vimentin on the move: New developments in cell migration. F1000Res 2018, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Y.; Shen, S.; Verma, I.M. NF-kappaB, an active player in human cancers. Cancer Immunol. Res. 2014, 2, 823–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McElwee, M.K.; Song, M.O.; Freedman, J.H. Copper activation of NF-kappaB signaling in HepG2 cells. J. Mol. Biol. 2009, 393, 1013–1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilles, C.; Polette, M.; Mestdagt, M.; Nawrocki-Raby, B.; Ruggeri, P.; Birembaut, P.; Foidart, J.M. Transactivation of vimentin by beta-catenin in human breast cancer cells. Cancer Res. 2003, 63, 2658–2664. [Google Scholar] [CrossRef] [Green Version]
- Razak, S.; Afsar, T.; Ullah, A.; Almajwal, A.; Alkholief, M.; Alshamsan, A.; Jahan, S. Taxifolin, a natural flavonoid interacts with cell cycle regulators causes cell cycle arrest and causes tumor regression by activating Wnt/ beta -catenin signaling pathway. BMC Cancer 2018, 18, 1043. [Google Scholar] [CrossRef] [Green Version]
- Manigandan, K.; Manimaran, D.; Jayaraj, R.L.; Elangovan, N.; Dhivya, V.; Kaphle, A. Taxifolin curbs NF-kappaB-mediated Wnt/beta-catenin signaling via up-regulating Nrf2 pathway in experimental colon carcinogenesis. Biochimie 2015, 119, 103–112. [Google Scholar] [CrossRef]
- Manning, B.D.; Toker, A. AKT/PKB Signaling: Navigating the Network. Cell 2017, 169, 381–405. [Google Scholar] [CrossRef] [Green Version]
- Carruba, G.; Cervello, M.; Miceli, M.D.; Farruggio, R.; Notarbartolo, M.; Virruso, L.; Giannitrapani, L.; Gambino, R.; Montalto, G.; Castagnetta, L. Truncated form of beta-catenin and reduced expression of wild-type catenins feature HepG2 human liver cancer cells. Ann. N. Y. Acad. Sci. 1999, 886, 212–216. [Google Scholar] [CrossRef]
- Wei, W.; Jin, J.; Schlisio, S.; Harper, J.W.; Kaelin, W.G., Jr. The v-Jun point mutation allows c-Jun to escape GSK3-dependent recognition and destruction by the Fbw7 ubiquitin ligase. Cancer Cell 2005, 8, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Szymonowicz, K.; Oeck, S.; Malewicz, N.M.; Jendrossek, V. New Insights into Protein Kinase B/Akt Signaling: Role of Localized Akt Activation and Compartment-Specific Target Proteins for the Cellular Radiation Response. Cancers 2018, 10, 78. [Google Scholar] [CrossRef] [Green Version]
- Crawford, R.R.; Potukuchi, P.K.; Schuetz, E.G.; Schuetz, J.D. Beyond Competitive Inhibition: Regulation of ABC Transporters by Kinases and Protein-Protein Interactions as Potential Mechanisms of Drug-Drug Interactions. Drug Metab. Dispos. Biol. Fate Chem. 2018, 46, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Li, H.; Pi, Y.; Li, Z.; Jin, S. Cardioprotective effect of IGF-1 against myocardial ischemia/reperfusion injury through activation of PI3K/Akt pathway in rats in vivo. J. Int. Med. Res. 2019, 47, 3886–3897. [Google Scholar] [CrossRef] [Green Version]
- Manning, B.D.; Cantley, L.C. AKT/PKB signaling: Navigating downstream. Cell 2007, 129, 1261–1274. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.S.; Rosenblatt, K.; Huang, K.L.; Lahat, G.; Brobey, R.; Bolshakov, S.; Nguyen, T.; Ding, Z.; Belousov, R.; Bill, K.; et al. Vimentin is a novel AKT1 target mediating motility and invasion. Oncogene 2011, 30, 457–470. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Gu, N.; Xue, C.; Li, B.R. Plant flavonoid taxifolin inhibits the growth, migration and invasion of human osteosarcoma cells. Mol. Med. Rep. 2018, 17, 3239–3245. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Xu, W.W.; Lam, A.K.Y.; Wang, Y.; Hu, H.F.; Guan, X.Y.; Qin, Y.R.; Saremi, N.; Tsao, S.W.; He, Q.Y.; et al. Significance of PI3K/AKT signaling pathway in metastasis of esophageal squamous cell carcinoma and its potential as a target for anti-metastasis therapy. Oncotarget 2017, 8, 38755–38766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sieuwerts, A.M.; Klijn, J.G.; Peters, H.A.; Foekens, J.A. The MTT tetrazolium salt assay scrutinized: How to use this assay reliably to measure metabolic activity of cell cultures in vitro for the assessment of growth characteristics, IC50-values and cell survival. Eur J. Clin. Chem. Clin. Biochem. 1995, 33, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Kollinerova, S.; Dostal, Z.; Modriansky, M. MicroRNA hsa-miR-29b potentiates etoposide toxicity in HeLa cells via down-regulation of Mcl-1. Toxicol. Vitr. 2017, 40, 289–296. [Google Scholar] [CrossRef]
- Zheng, Y.B.; Luo, H.P.; Shi, Q.; Hao, Z.N.; Ding, Y.; Wang, Q.S.; Li, S.B.; Xiao, G.C.; Tong, S.L. miR-132 inhibits colorectal cancer invasion and metastasis via directly targeting ZEB2. World J. Gastroenterol. 2014, 20, 6515–6522. [Google Scholar] [CrossRef]
- Zhou, D.D.; Wang, X.; Wang, Y.; Xiang, X.J.; Liang, Z.C.; Zhou, Y.; Xu, A.; Bi, C.H.; Zhang, L. MicroRNA-145 inhibits hepatic stellate cell activation and proliferation by targeting ZEB2 through Wnt/beta-catenin pathway. Mol. Immunol. 2016, 75, 151–160. [Google Scholar] [CrossRef]
- Kim, T.; Veronese, A.; Pichiorri, F.; Lee, T.J.; Jeon, Y.J.; Volinia, S.; Pineau, P.; Marchio, A.; Palatini, J.; Suh, S.S.; et al. p53 regulates epithelial-mesenchymal transition through microRNAs targeting ZEB1 and ZEB2. J. Exp. Med. 2011, 208, 875–883. [Google Scholar] [CrossRef] [PubMed]







| miRNA | Mean Relative Expression Compared to Control | References | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Quercetin | Taxifolin | ||||||||
| Hep G2 | Human Hepatocytes | Hep G2 | Human Hepatocytes | ||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | ||
| hsa-miR-129-5p | 2.42 | 2.11 | 0.94 | 0.27 | 4.53 | 6.58 | 1.07 | 0.17 | [31] |
| hsa-miR-139-5p | 1.40 | 0.16 | 1.35 | 0.86 | 2.14 | 1.06 | 1.34 | 1.09 | [32] |
| hsa-miR-141-3p | 1.90 | 0.96 | 0.98 | 0.17 | 1.56 | 1.21 | 1.00 | 0.42 | [30] |
| hsa-miR-153 | 1.02 | 0.37 | 0.72 | 0.24 | 0.67 | 0.27 | 1.39 | 0.89 | [30] |
| hsa-miR-154-5p | 1.40 | 0.44 | 1.73 | 1.09 | 1.63 | 0.48 | 1.45 | 0.60 | [30] |
| hsa-miR-200c-3p | 1.00 | 0.00 | 1.59 | 0.47 | 1.00 | 0.04 | 1.85 | 0.70 | [30] |
| hsa-miR-204 | 0.79 | 0.55 | 0.78 | 0.03 | 0.68 | 0.32 | 0.86 | 0.38 | [33] |
| hsa-miR-211 | 1.17 | 0.72 | 0.91 | 0.42 | 0.59 | 0.20 | 0.74 | 0.33 | [34] |
| hsa-miR-335-5p | 1.47 | 1.49 | 0.77 | 0.20 | 2.10 | 2.98 | 0.84 | 0.14 | [30] |
| hsa-miR-377-3p | 1.42 | 1.16 | 0.65 | 0.19 | 0.64 | 0.43 | 0.57 | 0.05 | [35] |
| hsa-miR-590-3p | 1.16 | 0.54 | 1.15 | 0.31 | 2.09 | 0.81 | 0.95 | 0.08 | [30] |
| hsa-miR-4782-3p | 1.47 | 0.55 | 0.75 | 0.21 | 1.73 | 0.96 | 1.06 | 0.37 | [30] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dostal, Z.; Sebera, M.; Srovnal, J.; Staffova, K.; Modriansky, M. Dual Effect of Taxifolin on ZEB2 Cancer Signaling in HepG2 Cells. Molecules 2021, 26, 1476. https://doi.org/10.3390/molecules26051476
Dostal Z, Sebera M, Srovnal J, Staffova K, Modriansky M. Dual Effect of Taxifolin on ZEB2 Cancer Signaling in HepG2 Cells. Molecules. 2021; 26(5):1476. https://doi.org/10.3390/molecules26051476
Chicago/Turabian StyleDostal, Zdenek, Martin Sebera, Josef Srovnal, Katerina Staffova, and Martin Modriansky. 2021. "Dual Effect of Taxifolin on ZEB2 Cancer Signaling in HepG2 Cells" Molecules 26, no. 5: 1476. https://doi.org/10.3390/molecules26051476
APA StyleDostal, Z., Sebera, M., Srovnal, J., Staffova, K., & Modriansky, M. (2021). Dual Effect of Taxifolin on ZEB2 Cancer Signaling in HepG2 Cells. Molecules, 26(5), 1476. https://doi.org/10.3390/molecules26051476