
Phytochemical Profiling of Lavandula coronopifolia Poir. Aerial Parts Extract and Its Larvicidal, Antibacterial, and Antibiofilm Activity Against Pseudomonas aeruginosa

 and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Material, Extraction, and Preliminary Qualitative Analysis
2.2. HPLC-MS/MS
2.3. Molecular Networking Workflow Description
2.4. Larvicidal Assay
2.4.1. Insects
2.4.2. Bioassay
2.4.3. Preparation of Samples for Biochemical Assay
2.5. Microbiological Assay
2.5.1. Sample Collection and Identification of Isolated Bacteria
2.5.2. The Antimicrobial Susceptibility Testing
2.5.3. Minimum Inhibitory Concentration (MIC)
2.5.4. Minimal Bactericidal Concentration (MBC)
2.5.5. Biofilm Formation Assay and Quantification
2.5.6. Biofilm Inhibition Assay
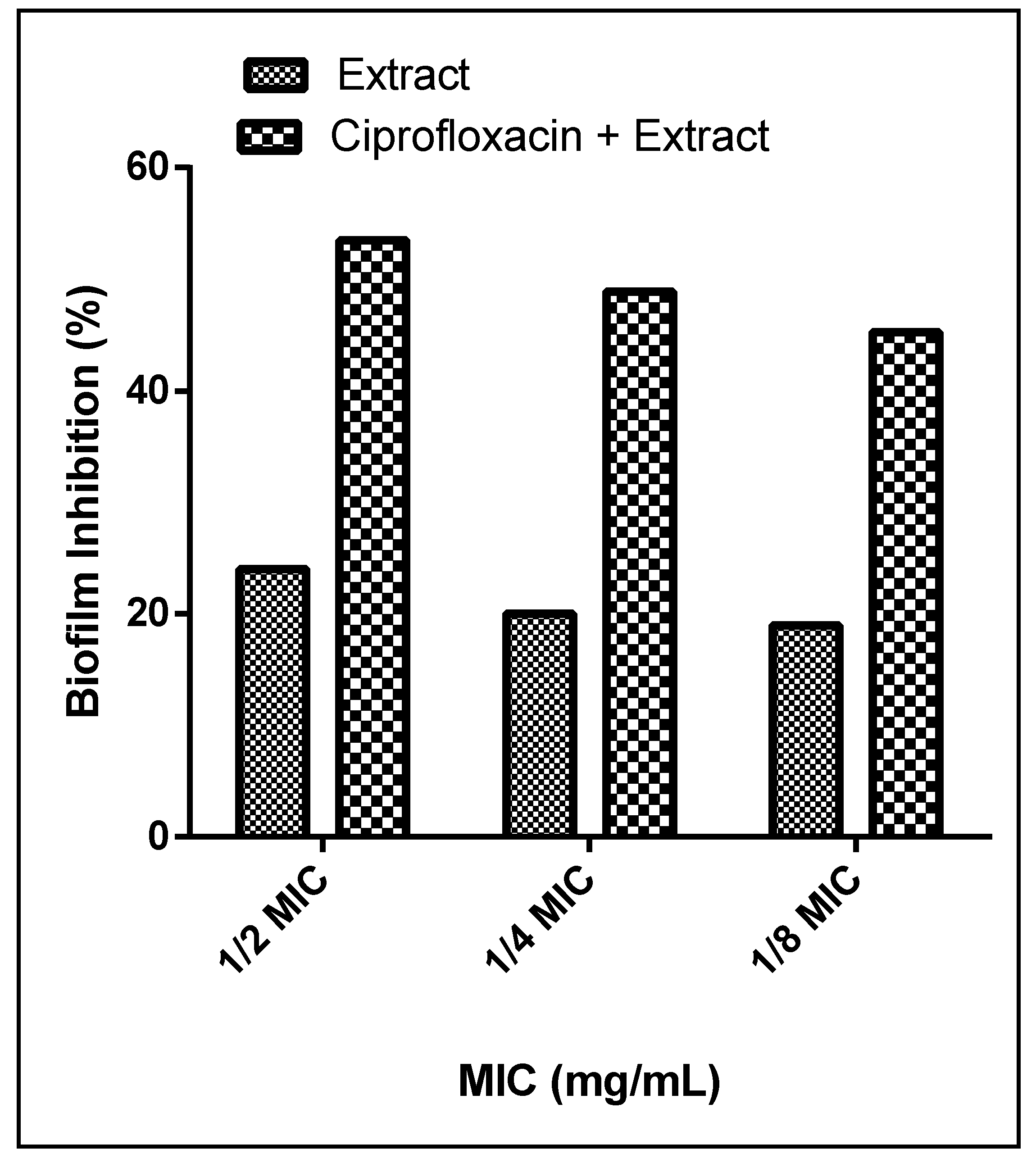
2.5.7. Combination of the Extract with Ciprofloxacin
2.6. Statistical Analysis
3. Results and Discussion
3.1. Phytochemical Screening and LC-MS/MS Profile of L. coronopifolia
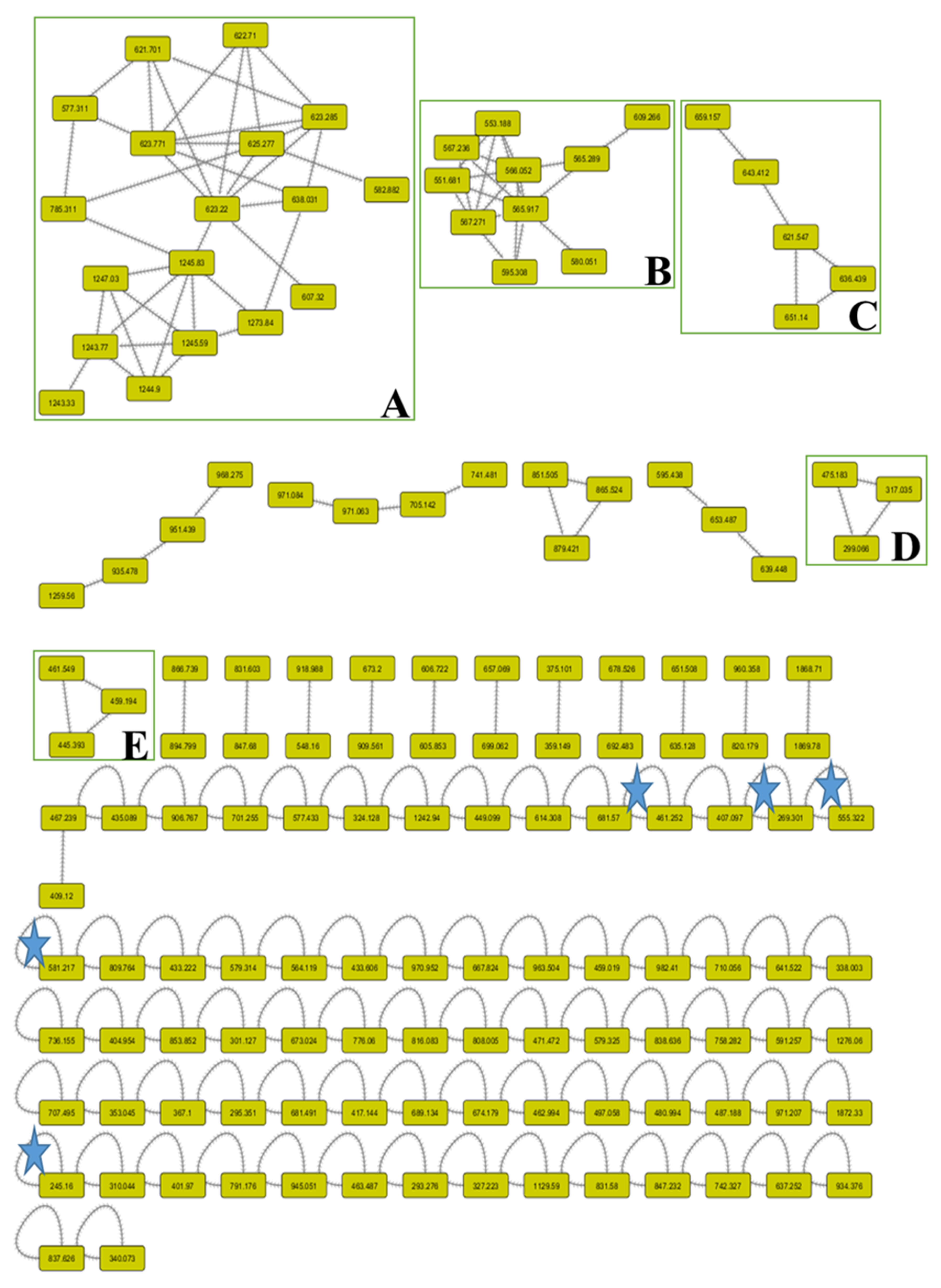
3.2. Molecular Networking (MNW) of L. coronopifolia Aerial Parts’ Metabolite Perception

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | tR (min) | [M–H]− | MS2 (m/z) | Tentatively Identified Compound | Ref. |
|---|---|---|---|---|---|
| 1 | 1.68 | 191 | 111, 173 | Quinic acid a | [38] |
| 2 | 1.75 | 133 | 89, 71, 115 | Malic acid a | [39] |
| 3 | 3.51 | 211 | 137, 179 | Caffeic acid derivative b | |
| 4 | 4.03 | 239 | 149, 179, 221 | Eucomic acid b | |
| 5 | 4.34 | 359 | 315, 197, 153 | Syringic acid 4-O-hexoside b | [38] |
| 6 | 4.61 | 237 | 115, 121, 137 | 2-(4-hydroxybenzyl)-malic acid b | |
| 7 | 5.03 | 375 | 125, 169, 213 | Sinapoyl trihydroxybenzoic acid b | |
| 8 | 6.31 | 475 | 197, 359 | Rosmarinic acid malate b | |
| 9 | 7.78 | 461 | 153, 315 | Protocatechuic acid rhamnosyl glucoside b | |
| 10 | 8.95 | 245 | 203, 186, 115 | N-Acetyltryptophan c | |
| 11 | 9.18 | 289 | 245 | Catechin d | [40] |
| 12 | 9.29 | 299 | 115, 133, 183 | Methyl trihydroxybenzoic acid malate b | |
| 13 | 13.54 | 385 | 223, 179 | Sinapic acid 3-O-glucoside e | |
| 14 | 13.69 | 387 | 433, 225, 179 | Dihydrosinapic acid hexoside e | |
| 15 | 14.51 | 387 | 433, 225, 179 | Dihydrosinapic acid hexoside e | |
| 16 | 15.61 | 387 | 433, 225, 179 | Dihydrosinapic acid hexoside e | |
| 17 | 17.57 | 389 | 227 | Resveratrol glucoside f | |
| 18 | 17.72 | 461 | 285 | Isoscutellarein-8-O-glucuronide #,g | |
| 19 | 18.25 | 359 | 197, 179, 161, 135 | Rosmarinic acid b | |
| 20 | 18.77 | 389 | 227 | Resveratrol glucoside f | |
| 21 | 19.88 | 461 | 285 | Luteolin-7-O-hexouronide g | [6] |
| 22 | 20.67 | 593 | 503, 473, 383, 353 | Apigenin di-C-hexoside g | [6] |
| 23 | 25.79 | 445 | 269, 175 | Apigenin-7-O-hexournide I @,g | [6] |
| 24 | 27.05 | 445 | 269, 175 | Apigenin-7-O-hexouronide II g | [6] |
| 25 | 28.62 | 623 | 477, 461, 315 | Hypolaetin 4′-O-methyl ether-O-hexoside-O-rhamnoside I g | |
| 26 | 33.57 | 623 | 477, 461, 315 | Hypolaetin-4′-O-methyl ether-O-hexoside-O-rhamnoside II g | |
| 27 | 32.30 | 623 | 179, 315, 461 | Isorhamnetin O-hexoside-O-rhamnoside g | |
| 28 | 34.98 | 607 | 461, 315, 299 | Hypolaetin di-O-rhamnoside g | |
| 29 | 35.08 | 447 | 285 | Luteolin-7-O-glucoside #,g | [6] |
| 30 | 37.23 | 637 | 491, 461, 315 | Hypolaetin 4′-O-methyl ether-8-glucuronide-O-rhamnoside g | |
| 31 | 38.80 | 459 | 283, 268 | Acacetin-O-hexouronic acid g | [41] |
| 32 | 41.04 | 577 | 269 | Apigenin-O-caffeoyl rhamnoside g | |
| 33 | 41.14 | 461 | 299, 283 | Methoxy leteolin-7-O-hexoside g | |
| 34 | 43.90 | 651 | 505, 475, 329 | Tricin-O-feruloyl rhamnoside g | |
| 35 | 48.62 | 621 | 459, 313 | Crismaritin-O-caffeoyl rhamnoside g | |
| 36 | 51.41 | 327 | 171, 229, 327 | Unknown | |
| 37 | 59.10 | 313 | 298, 284, 269 | Luteolin-7,3′-dimethyl ether g | [6] |
| 38 | 61.18 | 269 | 269, 151, 149 | Apigenin g | [6] |
| 39 | 62.63 | 555 | 403, 429, 327, 299 | Binaringenin methyl ether h | [37] |
| 40 | 69.07 | 551 | 457, 431, 389 | Methoxy amentoflavone h | |
| 41 | 69.42 | 553 | 458, 432, 390 | Dihydrobilobetin h | |
| 42 | 71.92 | 283 | 268, 133 | Acacetin g | [41] |
| 43 | 75.90 | 565 | 471, 389 | Dimethoxy amentoflavone h | |
| 44 | 78.77 | 609 | 577, 551, 489, 269 | Penta methoxy dihydro biapigenin h | |
| 45 | 81.37 | 579 | 533, 485, 389, 268 | Kayaflavone h | |
| 46 | 81.81 | 581 | 579, 535, 487 | Dihydrokayaflavone h |
3.3. 1H-NMR Analysis of L. coronopifolia Extract
3.4. Insecticidal Activity
3.5. Biochemical Activity
3.6. Microbiological Studies
3.6.1. Antimicrobial Susceptibility, MIC, and MBC
3.6.2. Biofilm Formation and Quantification Assay
3.6.3. Biofilm Inhibition Assay
3.6.4. Synergistic Activities
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Raju, P.; Arivalagan, P.; Natarajan, S. One-pot fabrication of multifunctional catechin@ ZIF-L nanocomposite: Assessment of antibiofilm, larvicidal and photocatalytic activities. J. Photochem. Photobiol. B Biol. 2020, 203, 111774. [Google Scholar] [CrossRef] [PubMed]
- Bagavan, A.; Rahuman, A.A. Evaluation of larvicidal activity of medicinal plant extracts against three mosquito vectors. Asian Pac. J. Trop. Med. 2011, 4, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Angajala, G.; Ramya, R.; Subashini, R. In-vitro anti-inflammatory and mosquito larvicidal efficacy of nickel nanoparticles phytofabricated from aqueous leaf extracts of Aegle marmelos Correa. Acta Trop. 2014, 135, 19–26. [Google Scholar] [CrossRef] [PubMed]
- El-Garf, I.; Grayer, R.J.; Kite, G.C.; Veitch, N.C. Hypolaetin 8-O-glucuronide and related flavonoids from Lavandula coronopifolia and L. pubescens. Biochem. Syst. Ecol. 1999, 27, 843–846. [Google Scholar] [CrossRef]
- The Plant List (2013). Version 1.1. Available online: http://www.theplantlist.org/ (accessed on 1 January 2021).
- Contreras, M.D.; Algieri, F.; Rodriguez-Nogales, A.; Gálvez, J.; Segura-Carretero, A. Phytochemical profiling of anti-inflammatory Lavandula extracts via RP–HPLC–DAD–QTOF–MS and–MS/MS: Assessment of their qualitative and quantitative differences. Electrophoresis 2018, 39, 1284–1293. [Google Scholar] [CrossRef] [PubMed]
- Harborne, J.B.; Williams, C.A. Phytochemistry of the Genus Lavandula; CRC Press: London, UK, 2002. [Google Scholar]
- Lis-Balchin, M. Lavender: The Genus Lavandula; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- El-Gendi, O.D.; Kusano, A.; Kusano, G. Two new triterpenic glucosidates from Lavandula coronipifolia in Egypt. Nat. Med. 2000, 54, 38–41. [Google Scholar]
- Elsbaey, M.; Mwakalukwa, R.; Shimizu, K.; Miyamoto, T. Pentacylic triterpenes from Lavandula coronopifolia: Structure related inhibitory activity on α-glucosidase. Nat. Prod. Res. 2019, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Messaoud, C.; Chograni, H.; Boussaid, M. Chemical composition and antioxidant activities of essential oils and methanol extracts of three wild Lavandula L. species. Nat. Prod. Res. 2012, 26, 1976–1984. [Google Scholar] [CrossRef]
- Hassan, W.; El Gamal, A.; El-Sheddy, E.; Al-Oquil, M.; Farshori, N. The chemical composition and antimicrobial activity of the essential oil of Lavandula coronopifolia growing in Saudi Arabia. J. Chem. Pharm. Res. 2014, 6, 604–615. [Google Scholar]
- Farshori, N.N.; Al-Sheddi, E.S.; Al-Oqail, M.M.; Hassan, W.H.; Al-Khedhairy, A.A.; Musarrat, J.; Siddiqui, M.A. Hepatoprotective potential of Lavandula coronopifolia extracts against ethanol induced oxidative stress-mediated cytotoxicity in HepG2 cells. Toxicol. Ind. Health 2015, 31, 727–737. [Google Scholar] [CrossRef]
- Ait Said, L.; Zahlane, K.; Ghalbane, I.; El Messoussi, S.; Romane, A.; Cavaleiro, C.; Salgueiro, L. Chemical composition and antibacterial activity of Lavandula coronopifolia essential oil against antibiotic-resistant bacteria. Nat. Prod. Res. 2015, 29, 582–585. [Google Scholar] [CrossRef]
- El-mekkawy, S.; Shahat, A.A.; Alqahtani, A.S.; Alsaid, M.S.; Abdelfattah, M.A.; Ullah, R.; Emam, M.; Yasri, A.; Sobeh, M. A Polyphenols-Rich Extract from Moricandia sinaica Boiss. Exhibits Analgesic, Anti-Inflammatory and Antipyretic Activities In Vivo. Molecules 2020, 25, 5049. [Google Scholar] [CrossRef] [PubMed]
- El-Garawani, I.; Emam, M.; Elkhateeb, W.; El-Seedi, H.; Khalifa, S.; Oshiba, S.; Abou-Ghanima, S.; Daba, G. In Vitro Antigenotoxic, Antihelminthic and Antioxidant Potentials Based on the Extracted Metabolites from Lichen, Candelariella vitellina. Pharmaceutics 2020, 12, 477. [Google Scholar] [CrossRef] [PubMed]
- El Raey, M.A.; El-Hagrassi, A.M.; Osman, A.F.; Darwish, K.M.; Emam, M. Acalypha wilkesiana flowers: Phenolic profiling, cytotoxic activity of their biosynthesized silver nanoparticles and molecular docking study for its constituents as Topoisomerase-I inhibitors. Biocatal. Agric. Biotechnol. 2019, 20, 101243. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerberg, E.J.; Barnard, D.R.; Ward, R.A. Manual for Mosquito Rearing and Experimental Techniques; American Mosquito Control Association Inc.: Mount Laurel, NJ, USA, 1994. [Google Scholar]
- World Health Organization. Guidelines for Laboratory and Field Testing of Mosquito Larvicides; World Health Organization: Geneva, Switzerland, 2005. [Google Scholar]
- Amin, T. Biochemical and Physiological Studies of Some Insect Growth Regulators on the Cotton Leafworm Spodoptera littoralis (Boisd.). Ph.D. Thesis, Faculty of Science, Cairo University, Giza, Egypt, 1998. [Google Scholar]
- Simpson, D.; Bull, D.; Lindquist, D. A semimicrotechnique for the estimation of cholinesterase activity in boll weevils. Ann. Entomol. Soc. Am. 1964, 57, 367–371. [Google Scholar] [CrossRef]
- Van Asperen, K. A study of housefly esterases by means of a sensitive colorimetric method. J. Insect Physiol. 1962, 8, 401–414. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases the first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Hansen, L.; Hodgson, E. Biochemical characteristics of insect microsomes: N-and O-demethylation. Biochem. Pharmacol. 1971, 20, 1569–1578. [Google Scholar] [CrossRef]
- El-Sayed, N.R.; Samir, R.; Abdel-Hafez, J.M.; Ramadan, M.A. Olive Leaf Extract Modulates Quorum Sensing Genes and Biofilm Formation in Multi-Drug Resistant Pseudomonas aeruginosa. Antibiotics 2020, 9, 526. [Google Scholar] [CrossRef]
- Cooper, K. Theory of antibiotic inhibition zones in agar media. Nature 1955, 176, 510–511. [Google Scholar] [CrossRef]
- Moideen, M.M.J.; Raffick, M.M. Antibacterial, antifungal activity and phytochemical analysis of Sanseviveria roxburghiana root. IJ Phytopharm. 2012, 3, 21–26. [Google Scholar]
- Briceland, L.L.; Pasko, M.; Mylotte, J. Serum bactericidal rate as measure of antibiotic interactions. Antimicrob. Agents Chemother. 1987, 31, 679–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wikler, M.A. Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically: Approved standard. CLSI (NCCLS) 2006, 26, M7-A7. [Google Scholar]
- O’Toole, G.A.; Kolter, R. Initiation of biofilm formation in Pseudomonas fluorescens WCS365 proceeds via multiple, convergent signalling pathways: A genetic analysis. Mol. Microbiol. 1998, 28, 449–461. [Google Scholar] [CrossRef]
- O’Toole, G.A. Microtiter dish biofilm formation assay. J. Vis. Exp. 2011, 47, e2437. [Google Scholar] [CrossRef] [PubMed]
- Stepanović, S.; Vuković, D.; Hola, V.; Bonaventura, G.D.; Djukić, S.; Ćirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. Apmis 2007, 115, 891–899. [Google Scholar] [CrossRef]
- Stepanović, S.; Vuković, D.; Dakić, I.; Savić, B.; Švabić-Vlahović, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Duncan, D.B. Multiple range and multiple F tests. Biometrics 1955, 11, 1–42. [Google Scholar] [CrossRef]
- Vukics, V.; Guttman, A. Structural characterization of flavonoid glycosides by multi-stage mass spectrometry. Mass Spectrom. Rev. 2010, 29, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Chen, B.; Zhang, Y.; Ou, H.; Li, Y.; Li, S.; Shi, P.; Lin, X. Analysis of the total biflavonoids extract from Selaginella doederleinii by HPLC-QTOF-MS and its in vitro and in vivo anticancer effects. Molecules 2017, 22, 325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Tao, S.; Zhang, S. Characterization and quantification of polyphenols and triterpenoids in thinned young fruits of ten pear varieties by UPLC-Q TRAP-MS/MS. Molecules 2019, 24, 159. [Google Scholar] [CrossRef] [Green Version]
- Spínola, V.; Pinto, J.; Castilho, P.C. Identification and quantification of phenolic compounds of selected fruits from Madeira Island by HPLC-DAD–ESI-MSn and screening for their antioxidant activity. Food Chem. 2015, 173, 14–30. [Google Scholar] [CrossRef]
- Yuzuak, S.; Ballington, J.; Xie, D.-Y. HPLC-qTOF-MS/MS-based profiling of flavan-3-ols and dimeric proanthocyanidins in berries of two muscadine grape hybrids FLH 13-11 and FLH 17-66. Metabolites 2018, 8, 57. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Wang, L.; Dai, P.; Zeng, X.; Qi, X.; Zhu, L.; Yan, T.; Wang, Y.; Lu, L.; Hu, M. A combined strategy of mass fragmentation, post-column cobalt complexation and shift in ultraviolet absorption spectra to determine the uridine 5′-diphospho-glucuronosyltransferase metabolism profiling of flavones after oral administration of a flavone mixture in rats. J. Chromatogr. A 2015, 1395, 116–128. [Google Scholar] [PubMed]
- Helmy, N.; Bakr, R.F.; Nawwar, G.A.; Helmy, O.M. Biochemical effects of some agricultural waste extracts against Culex pipiens (Diptera: Culicidae). Egypt. Acad. J. Biol. Sci. C Physiol. Mol. Biol. 2010, 2, 75–81. [Google Scholar] [CrossRef]
- Dris, D.; Tine-Djebbar, F.; Soltani, N. Lavandula dentata essential oils: Chemical composition and larvicidal activity against Culiseta longiareolata and Culex pipiens (Diptera: Culicidae). Afr. Entomol. 2017, 25, 387–394. [Google Scholar] [CrossRef]
- Traboulsi, A.F.; Taoubi, K.; El-Haj, S.; Bessiere, J.M.; Rammal, S. Insecticidal properties of essential plant oils against the mosquito Culex pipiens molestus (Diptera: Culicidae). Pest Manag. Sci. 2002, 58, 491–495. [Google Scholar] [CrossRef]
- Waliwitiya, R.; Nicholson, R.A.; Kennedy, C.J.; Lowenberger, C.A. The synergistic effects of insecticidal essential oils and piperonyl butoxide on biotransformational enzyme activities in Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2012, 49, 614–623. [Google Scholar] [CrossRef]
- Huang, C.-S.; Lii, C.-K.; Lin, A.-H.; Yeh, Y.-W.; Yao, H.-T.; Li, C.-C.; Wang, T.-S.; Chen, H.-W. Protection by chrysin, apigenin, and luteolin against oxidative stress is mediated by the Nrf2-dependent up-regulation of heme oxygenase 1 and glutamate cysteine ligase in rat primary hepatocytes. Arch. Toxicol. 2013, 87, 167–178. [Google Scholar] [CrossRef]
- Oppenoorth, F.; Rupes, V.; ElBashir, S.; Houx, N.; Voerman, S. Glutathione-dependent degradation of parathion and its significance for resistance in the housefly. Pestic. Biochem. Physiol. 1972, 2, 262–269. [Google Scholar] [CrossRef]
- Motoyama, N.; Dauterman, W. Interstrain comparison of glutathione-dependent reactions in susceptible and resistant houseflies. Pestic. Biochem. Physiol. 1975, 5, 489–495. [Google Scholar] [CrossRef]
- Gershenzon, J.; Fontana, A.; Burow, M.; Wittstock, U.; Degenhardt, J. Mixtures of plant secondary metabolites: Metabolic origins and ecological benefits. In the Ecology of Plant Secondary Metabolites: From Genes to Global Processes; Cambridge University Press: Cambridge, UK, 2012; Volume 56. [Google Scholar]
- Salunke, B.; Kotkar, H.; Mendki, P.; Upasani, S.; Maheshwari, V. Efficacy of flavonoids in controlling Callosobruchus chinensis (L.) (Coleoptera: Bruchidae), a post-harvest pest of grain legumes. Crop Prot. 2005, 24, 888–893. [Google Scholar] [CrossRef]
- Djeridane, A.; Yousfi, M.; Nadjemi, B.; Maamri, S.; Djireb, F.; Stocker, P. Phenolic extracts from various Algerian plants as strong inhibitors of porcine liver carboxylesterase. J. Enzym. Inhib. Med. Chem. 2006, 21, 719–726. [Google Scholar] [CrossRef] [PubMed]
- Barbosa Filho, J.M.; Medeiros, K.C.P.; Diniz, M.d.F.F.; Batista, L.M.; Athayde-Filho, P.F.; Silva, M.S.; da Cunha, E.V.; Almeida, J.R.; Quintans-Júnior, L.J. Natural products inhibitors of the enzyme acetylcholinesterase. Rev. Bras. Farmacogn. 2006, 16, 258–285. [Google Scholar] [CrossRef] [Green Version]
- Praveena, A.; Sanjayan, K. Inhibition of acetylcholinesterase in three insects of economic importance by linalool, a monoterpene phytochemical. Insect Pest Manag. A Curr. Scenar. 2011, 340–345. [Google Scholar]
- Galvão, S.S.L.; Monteiro, A.; Siqueira, E.P.; Bomfim, M.R.Q.; Dias-Souza, M.V.; Ferreira, G.F.; Denadai, A.M.L.; Santos, Á.R.; Lúcia dos Santos, V.; Souza-Fagundes, E.M. Annona glabra flavonoids act as antimicrobials by binding to Pseudomonas aeruginosa cell walls. Front. Microbiol. 2016, 7, 2053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostafa, I.; Abbas, H.A.; Ashour, M.L.; Yasri, A.; El-Shazly, A.M.; Wink, M.; Sobeh, M. Polyphenols from Salix tetrasperma impair virulence and inhibit Quorum sensing of Pseudomonas aeruginosa. Molecules 2020, 25, 1341. [Google Scholar] [CrossRef] [Green Version]
- Nain, Z.; Mansur, F.J.; Syed, S.B.; Islam, M.A.; Azakami, H.; Islam, M.R.; Karim, M.M. Inhibition of biofilm formation, quorum sensing and other virulence factors in Pseudomonas aeruginosa by polyphenols of Gynura procumbens leaves. J. Biomol. Struct. Dyn. 2020, 1–15. [Google Scholar] [CrossRef]
- Koley, T.K.; Khan, Z.; Oulkar, D.; Banerjee, T.; Singh, A.; Karkute, S.G.; Banerjee, K. Coupling the high-resolution LC–MS characterisation of the phenolic compounds with the antimicrobial and antibiofilm properties of helencha (Enydra fluctuans Lour.). J. Food Sci. Technol. 2021, 1–11. [Google Scholar] [CrossRef]
- Mrabti, H.N.; Bouyahya, A.; Ed-Dra, A.; Kachmar, M.R.; Mrabti, N.N.; Benali, T.; Shariati, M.A.; Ouahbi, A.; Doudach, L.; Faouzi, M.E.A. Polyphenolic profile and biological properties of Arbutus unedo root extracts. Eur. J. Integr. Med. 2021, 42, 101266. [Google Scholar] [CrossRef]
- Ofokansi, K.C.; Attama, A.A.; Uzor, P.F.; Ovri, M.O. Evaluation of the combined antimicrobial activity of the leaf extract of Phyllantus muellerianus with ciprofloxacin. J. Pharm Technol. Drug Res. 2013, 2, 16. [Google Scholar] [CrossRef] [Green Version]
- Kumar, L.; Chhibber, S.; Harjai, K. Zingerone inhibit biofilm formation and improve antibiofilm efficacy of ciprofloxacin against Pseudomonas aeruginosa PAO1. Fitoterapia 2013, 90, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Grau, S.; Echeverria-Esnal, D.; Gómez-Zorrilla, S.; Navarrete-Rouco, M.E.; Masclans, J.R.; Espona, M.; Gracia-Arnillas, M.P.; Duran, X.; Comas, M.; Horcajada, J.P. Evolution of Antimicrobial Consumption During the First Wave of COVID-19 Pandemic. Antibiotics 2021, 10, 132. [Google Scholar] [CrossRef]
- Tiwari Pandey, A.; Pandey, I.; Zamboni, P.; Gemmati, D.; Kanase, A.; Singh, A.V.; Singh, M.P. Traditional Herbal Remedies with a Multifunctional Therapeutic Approach as an Implication in COVID-19 Associated Co-Infections. Coatings 2020, 10, 761. [Google Scholar] [CrossRef]






| Extract (µg/mL) | 24 h Post Treatment | 48 h Post Treatment | 72 h Post Treatment |
|---|---|---|---|
| LC25 (* F.l. at 95%) | 20.054 (15.20–24.85) | 11.274 (7.55–15.05) | 8.668 (5.29–12.21) |
| LC50(* F.l. at 95%) | 52.74 (44.304–62.95) | 34.07 (27.43–41.41) | 29.076 (22.58–36.062) |
| LC95(* F.l. at 95%) | 557.50 (374.11–975.18) | 505.44 (326.011–953.94) | 556.28 (341.87–1151.47) |
| Slope ± SE | 1.61 ± 0.147 | 1.40 ± 0.141 | 1.28 ± 0.138 |
| χ2 a | 6.4544 | 1.9897 | 0.5856 |
| Probability (P) | 0.0915 | 0.5745 | 0.8997 |
| P. aeruginosa Isolates | Zone of Inhibition (mm) | Extract | ||
|---|---|---|---|---|
| Extract | Ciprofloxacin | MIC | MBC | |
| mg/mL | ||||
| C1 | 20 | 1748 | 0.3125 | 1.25 |
| C2 | 20 | 40 | 0.3125 | 1.25 |
| C3 | 20 | 50 | 0.3125 | 1.25 |
| C4 | 28 | 35 | 0.3125 | 1.25 |
| ATCC (12924) | 26 | 22 | 0.1562 | 1.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Emam, M.; Abdel-Haleem, D.R.; Salem, M.M.; Abdel-Hafez, L.J.M.; Latif, R.R.A.; Farag, S.M.; Sobeh, M.; El Raey, M.A. Phytochemical Profiling of Lavandula coronopifolia Poir. Aerial Parts Extract and Its Larvicidal, Antibacterial, and Antibiofilm Activity Against Pseudomonas aeruginosa. Molecules 2021, 26, 1710. https://doi.org/10.3390/molecules26061710
Emam M, Abdel-Haleem DR, Salem MM, Abdel-Hafez LJM, Latif RRA, Farag SM, Sobeh M, El Raey MA. Phytochemical Profiling of Lavandula coronopifolia Poir. Aerial Parts Extract and Its Larvicidal, Antibacterial, and Antibiofilm Activity Against Pseudomonas aeruginosa. Molecules. 2021; 26(6):1710. https://doi.org/10.3390/molecules26061710
Chicago/Turabian StyleEmam, Mahmoud, Doaa R. Abdel-Haleem, Maha M. Salem, Lina Jamil M. Abdel-Hafez, Rasha R. Abdel Latif, Shaimaa Mahmoud Farag, Mansour Sobeh, and Mohamed A. El Raey. 2021. "Phytochemical Profiling of Lavandula coronopifolia Poir. Aerial Parts Extract and Its Larvicidal, Antibacterial, and Antibiofilm Activity Against Pseudomonas aeruginosa" Molecules 26, no. 6: 1710. https://doi.org/10.3390/molecules26061710
APA StyleEmam, M., Abdel-Haleem, D. R., Salem, M. M., Abdel-Hafez, L. J. M., Latif, R. R. A., Farag, S. M., Sobeh, M., & El Raey, M. A. (2021). Phytochemical Profiling of Lavandula coronopifolia Poir. Aerial Parts Extract and Its Larvicidal, Antibacterial, and Antibiofilm Activity Against Pseudomonas aeruginosa. Molecules, 26(6), 1710. https://doi.org/10.3390/molecules26061710