
Valonea Tannin: Tyrosinase Inhibition Activity, Structural Elucidation and Insights into the Inhibition Mechanism


Abstract
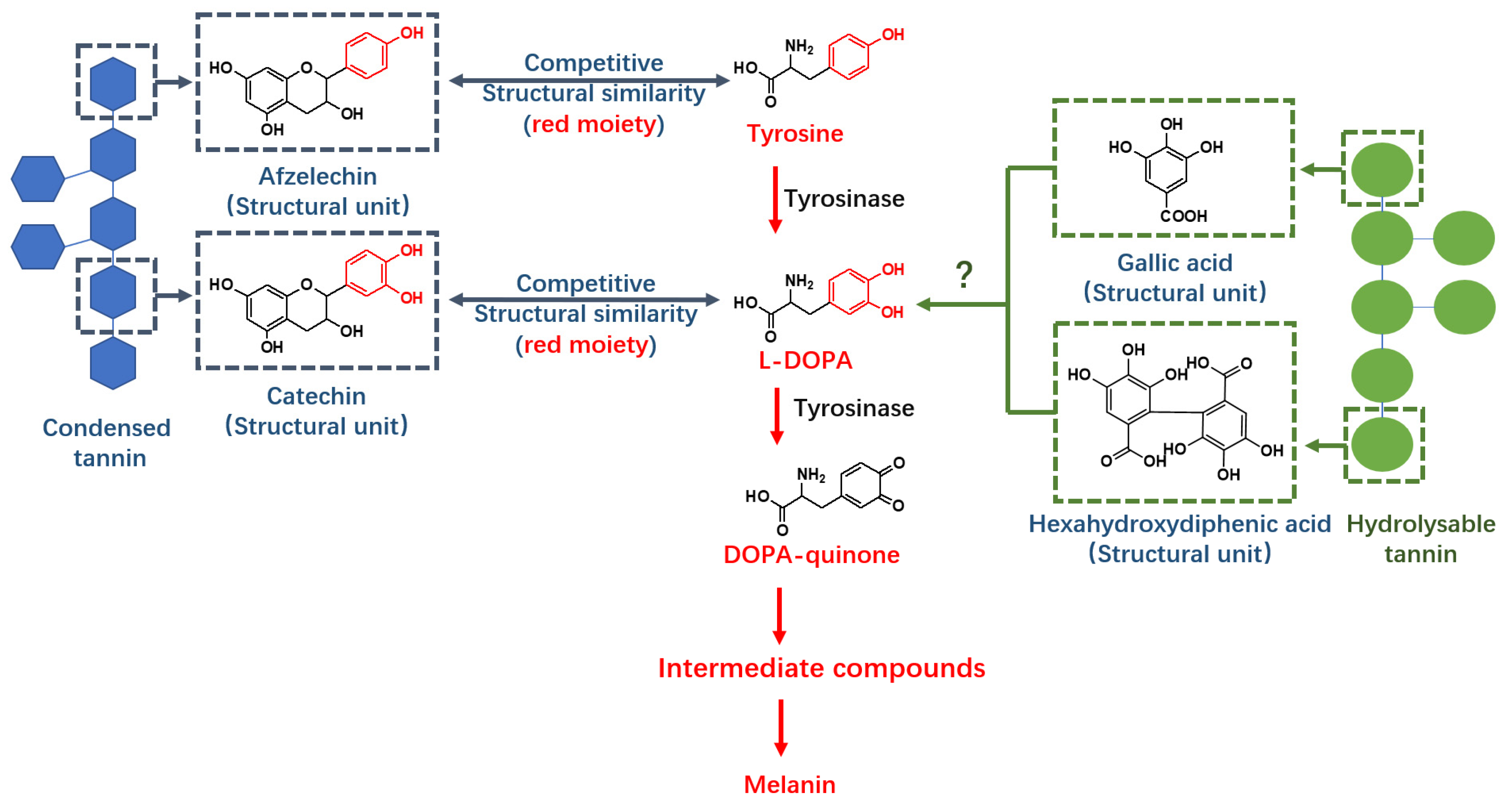
:1. Introduction
2. Results and Discussion
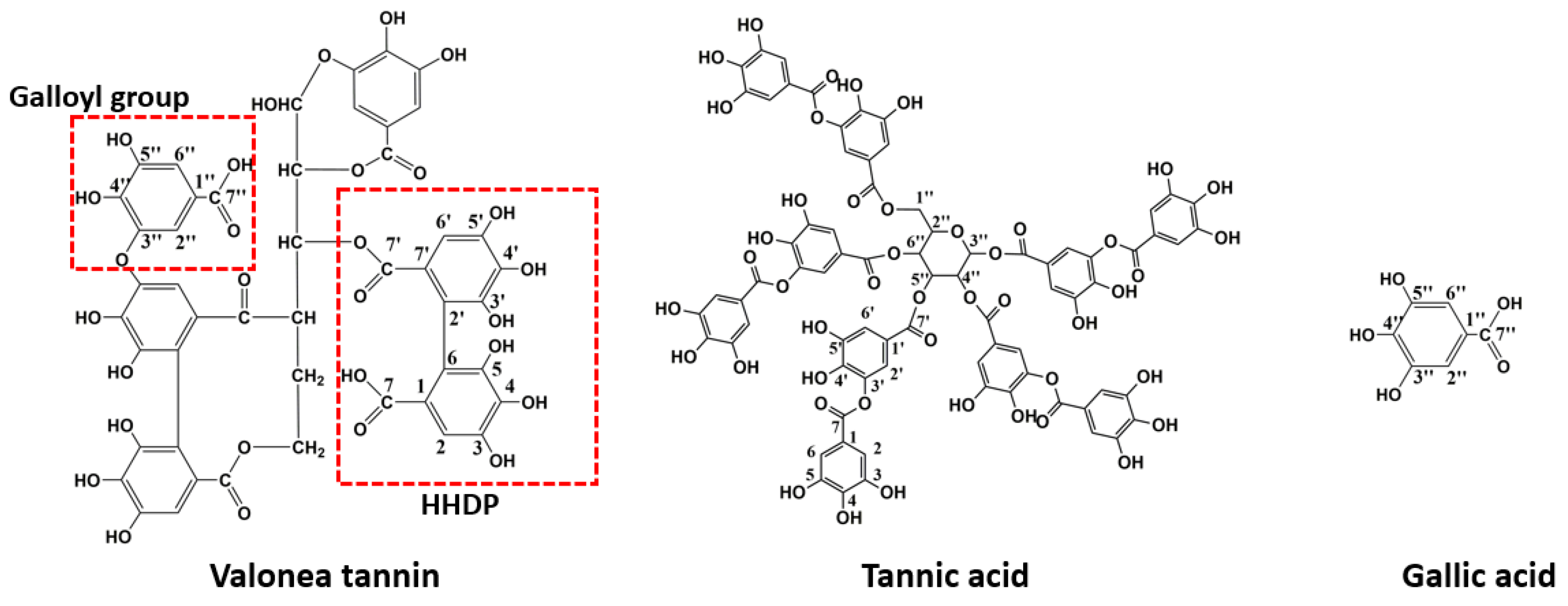
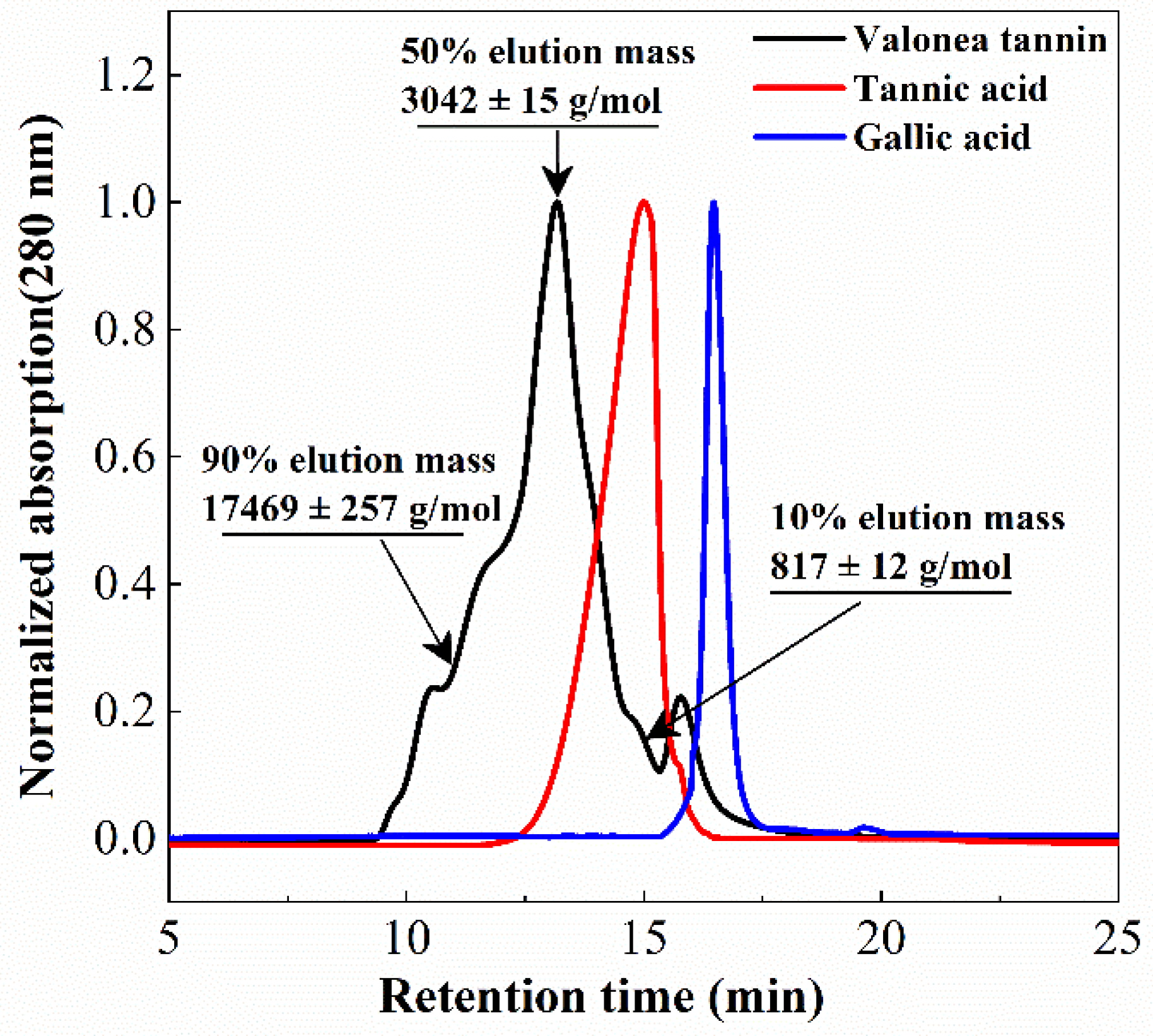
2.1. Structure Elucidation of Valonea Tannins
2.2. Tyrosinase Inhibition Activity of the Valonea Tannin
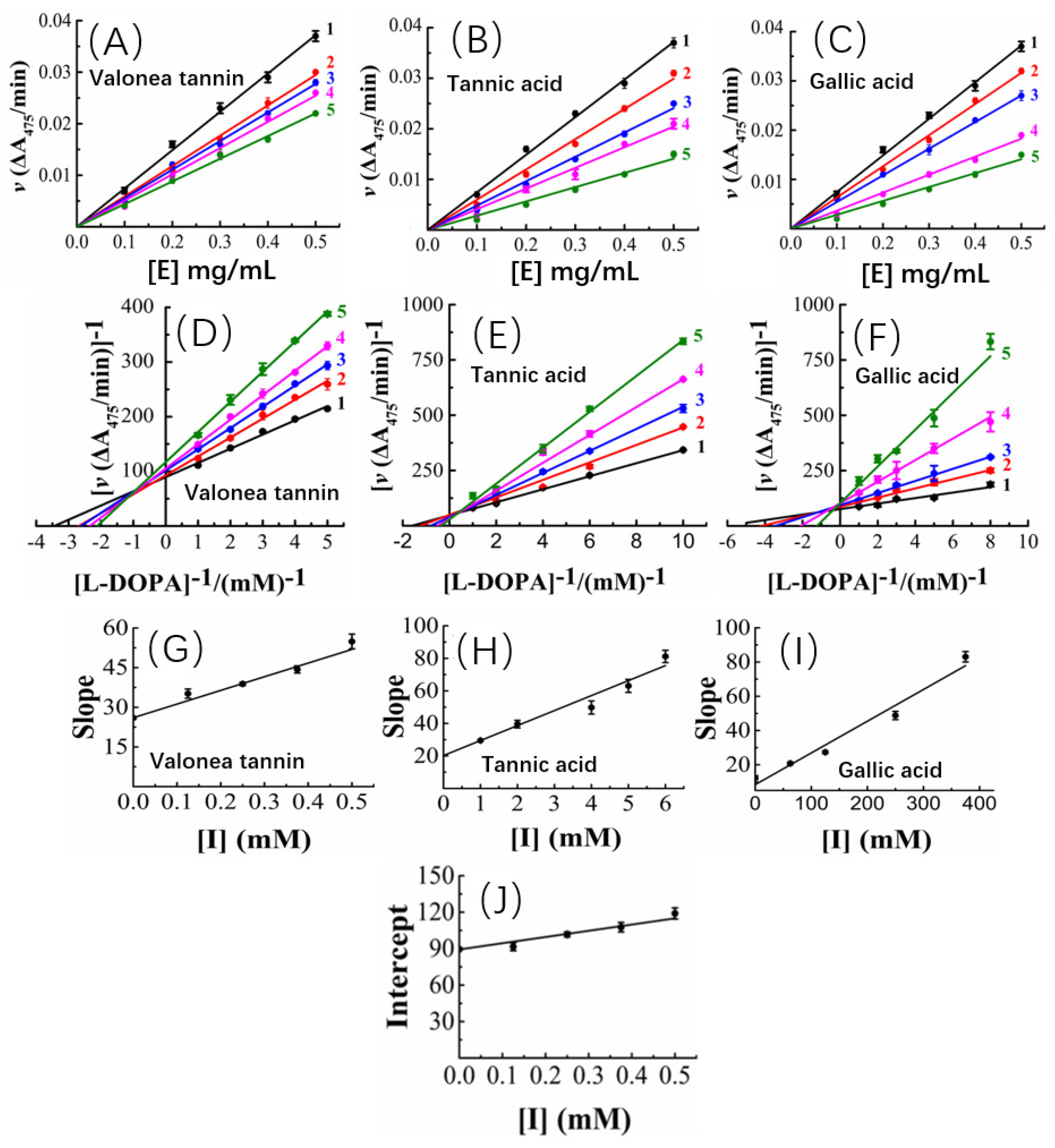
2.3. The Kinetic Study of Tyrosinase Inhibition Mechanism
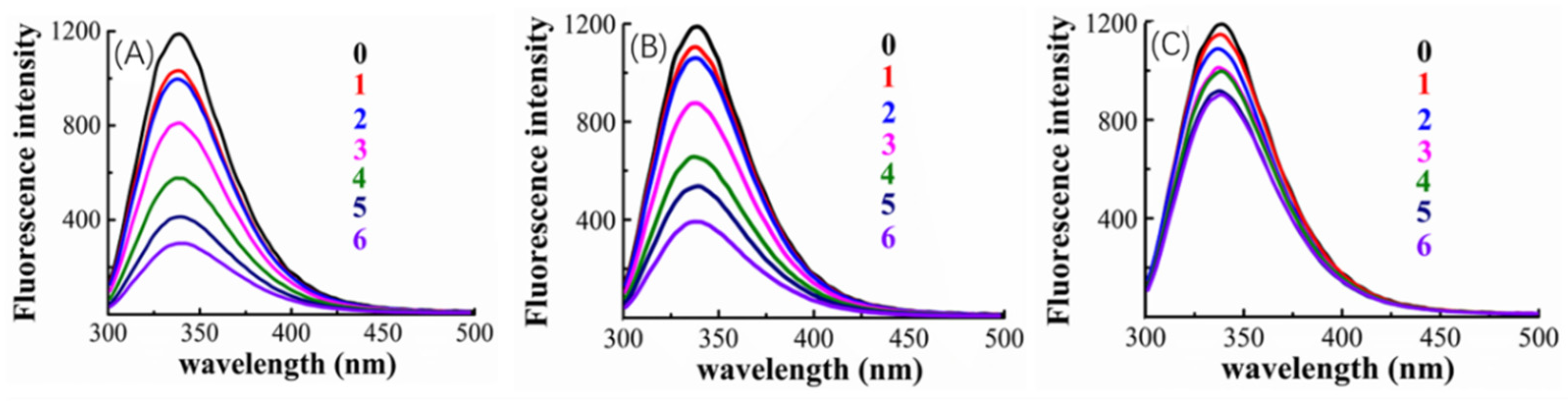
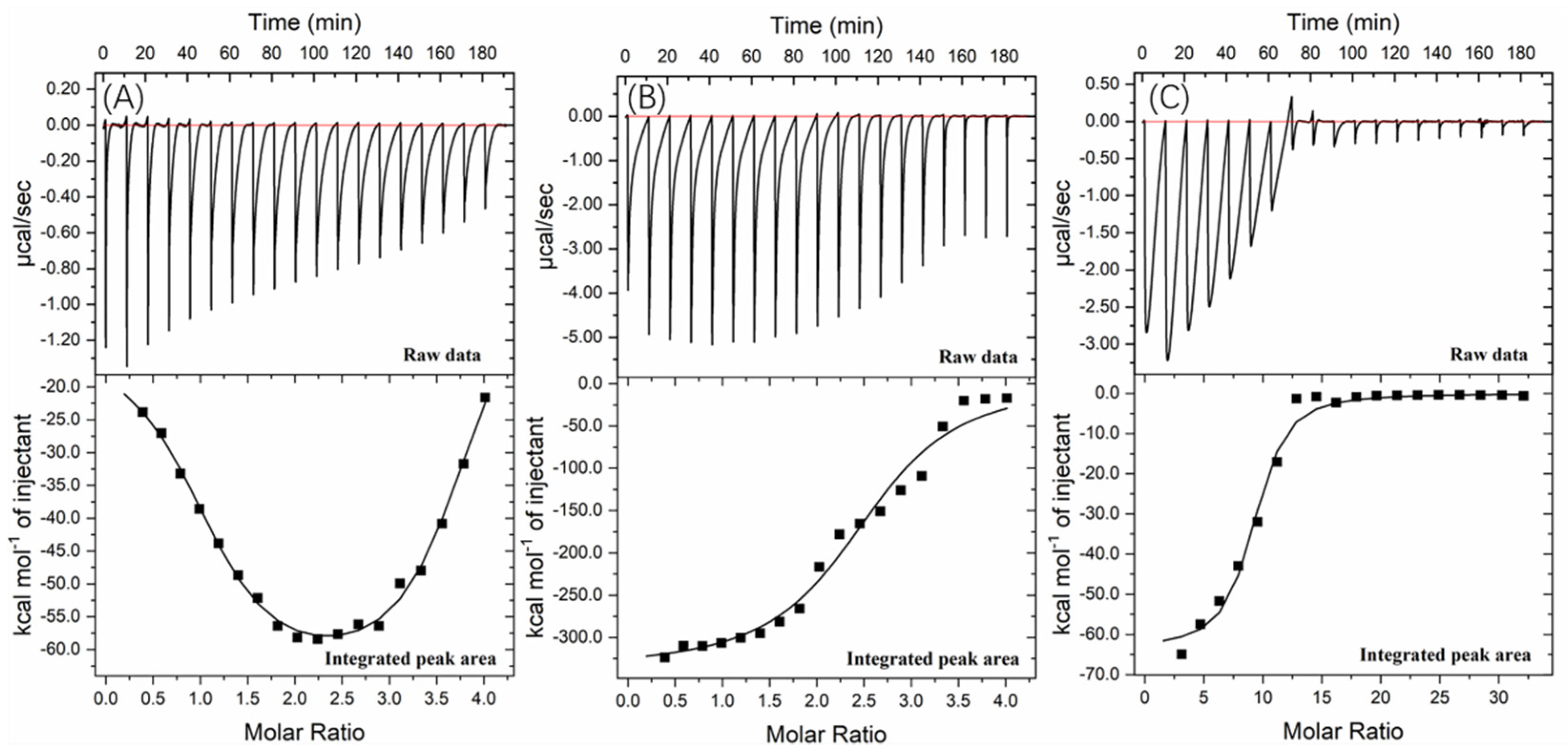
2.4. Fluorescence and Thermodynamic Characters of the Valonea Tannin-Tyrosinase Interaction
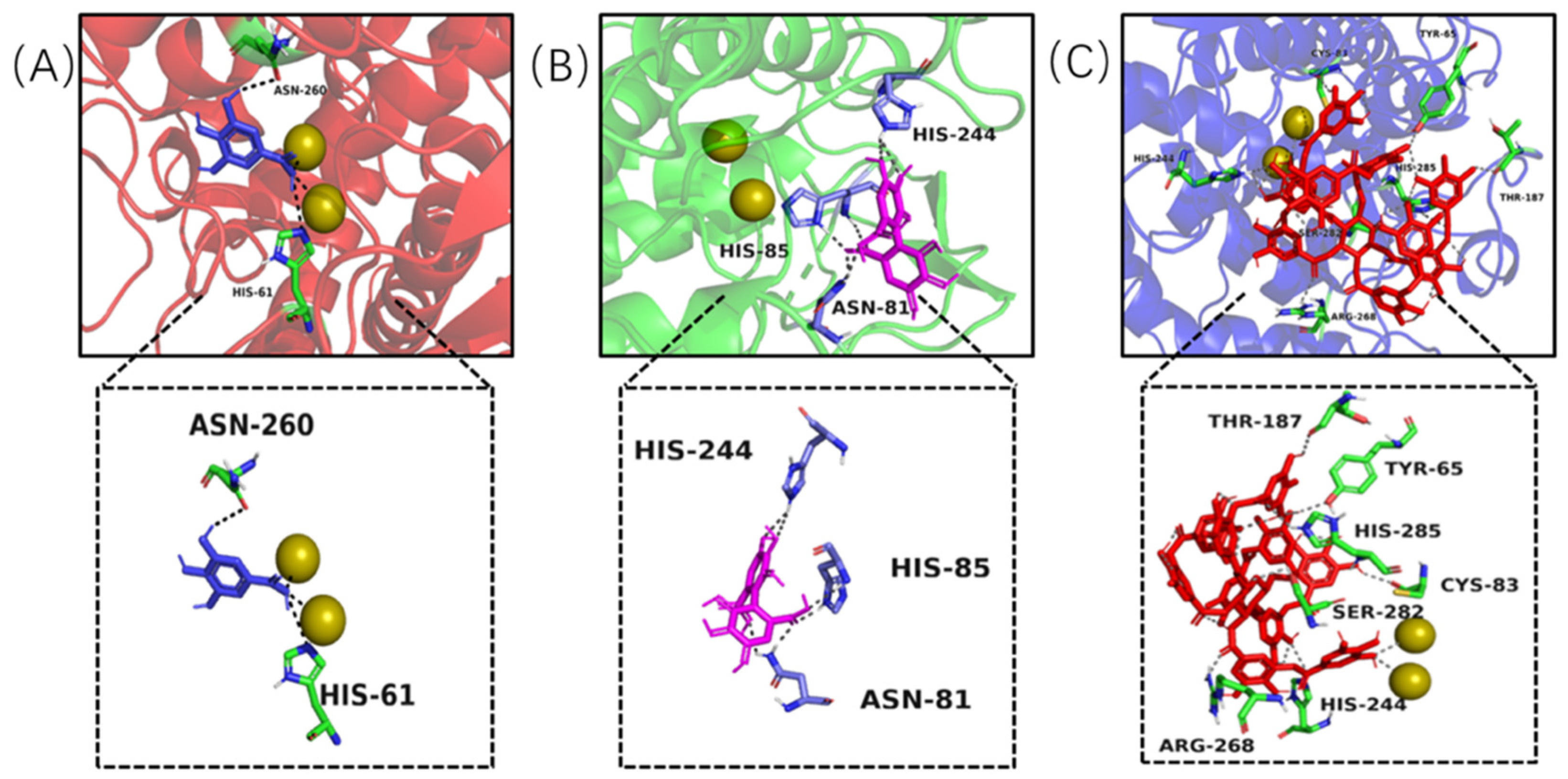
2.5. Molecular Docking Analysis
2.6. Copper Ion Chelating and Antioxidant Abilities
3. Materials and Methods
3.1. Extraction and Purification of Valonea Tannins
3.2. Characterization of Valonea Tannin Structure
3.3. Tyrosinase Inhibition Activity Assessment and Inhibition Type Analysis
3.4. Fluorescence Quenching Analysis of Tyrosinase in the Presence of Valonea Tannins
3.5. Isothermal Titration Calorimetry Analysis
3.6. In Silico Molecule Docking
3.7. Antioxidant Activity Analysis
3.8. Inductively Coupled Plasma-Optical Emission Spectroscopy (ICP-OES)
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Romani, A.; Ieri, F.; Turchetti, B.; Mulinacci, N.; Vincieri, F.; Buzzini, P. Analysis of condensed and hydrolysable tannins from commercial plant extracts. J. Pharm. Biomed. Anal. 2006, 41, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wang, W.; Mu, Y.; Zhang, J.; Zhang, S.; Li, J.; Zhang, W. Structural Properties and Copolycondensation Mechanism of Valonea Tannin-Modified Phenol-formaldehyde Resin. J. Polym. Environ. 2018, 26, 1297–1309. [Google Scholar] [CrossRef]
- Ren, Z.; Yu, F.; Gao, H.; Chen, Z.; Peng, Y.; Liu, L. Selective Separation of Fluorite, Barite and Calcite with Valonea Extract and Sodium Fluosilicate as Depressants. Minerals 2017, 7, 24. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Wang, L. Developing a bio-based packaging film from soya by-products incorporated with valonea tannin. J. Clean. Prod. 2017, 143, 624–633. [Google Scholar] [CrossRef]
- Onem, E.; Gulumser, G.; Akay, S.; Yesil-Celiktas, O. Optimization of tannin isolation from acorn and application in leather processing. Ind. Crops Prod. 2014, 53, 16–22. [Google Scholar] [CrossRef]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin whitening agents: Medicinal chemistry perspective of tyrosinase inhibitors. J. Enzym. Inhib. Med. Chem. 2017, 32, 403–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iozumi, K.; Hoganson, G.E.; Pennella, R.; Everett, M.A.; Fuller, B.B. Role of Tyrosinase as the Determinant of Pigmentation in Cultured Human Melanocytes. J. Investig. Dermatol. 1993, 100, 806–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lianza, M.; Mandrone, M.; Chiocchio, I.; Tomasi, P.; Marincich, L.; Poli, F. Screening of ninety herbal products of commercial interest as potential ingredients for phytocosmetics. J. Enzym. Inhib. Med. Chem. 2020, 35, 1287–1291. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.T.H. Molecular design of tyrosinase inhibitors: A critical review of promising novel inhibitors from synthetic origins. Pure Appl. Chem. 2007, 79, 2277–2295. [Google Scholar] [CrossRef] [Green Version]
- Zolghadri, S.; Bahrami, A.; Khan, M.T.H.; Munoz-Munoz, J.; Garcia-Molina, F.; Garcia-Canovas, F.; Saboury, A.A. A comprehensive review on tyrosinase inhibitors. J. Enzym. Inhib. Med. Chem. 2019, 34, 279–309. [Google Scholar] [CrossRef] [Green Version]
- Weiss, R.M.; Del Fabbro, E.; Kolisang, P. Cosmetic ochronosis caused by bleaching creams containing 2% hydroquinone. South Afr. Med J. 1990, 77, 373. [Google Scholar]
- A Ibrahim, Z.; Gheida, S.F.; El Maghraby, G.M.; E Farag, Z. Evaluation of the efficacy and safety of combinations of hydroquinone, glycolic acid, and hyaluronic acid in the treatment of melasma. J. Cosmet. Dermatol. 2015, 14, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Schofield, P.; Mbugua, D.; Pell, A. Analysis of condensed tannins: A review. Anim. Feed. Sci. Technol. 2001, 91, 21–40. [Google Scholar] [CrossRef]
- Sarikurkcu, C.; Kocak, M.S.; Tepe, B.; Uren, M.C. An alternative antioxidative and enzyme inhibitory agent from Turkey: Robinia pseudoacacia L. Ind. Crops Prod. 2015, 78, 110–115. [Google Scholar] [CrossRef]
- Chai, W.-M.; Shi, Y.; Feng, H.-L.; Xu, L.; Xiang, Z.-H.; Gao, Y.-S.; Chen, Q.-X. Structure Characterization and Anti-tyrosinase Mechanism of Polymeric Proanthocyanidins Fractionated from Kiwifruit Pericarp. J. Agric. Food Chem. 2014, 62, 6382–6389. [Google Scholar] [CrossRef] [PubMed]
- Chai, W.-M.; Wang, R.; Wei, M.-K.; Zou, Z.-R.; Deng, R.-G.; Liu, W.-S.; Peng, Y.-Y. Proanthocyanidins Extracted from Rhododendron pulchrum Leaves as Source of Tyrosinase Inhibitors: Structure, Activity, and Mechanism. PLoS ONE 2015, 10, e0145483. [Google Scholar] [CrossRef]
- Manosroi, A.; Jantrawut, P.; Akazawa, H.; Akihisa, T.; Manosroi, J. Biological activities of phenolic compounds isolated from galls of Terminalia chebula Retz. (Combretaceae). Nat. Prod. Res. 2010, 24, 1915–1926. [Google Scholar] [CrossRef] [PubMed]
- Teng, B.; Hayasaka, Y.; Smith, P.A.; Bindon, K.A. Effect of Grape Seed and Skin Tannin Molecular Mass and Composition on the Rate of Reaction with Anthocyanin and Subsequent Formation of Polymeric Pigments in the Presence of Acetaldehyde. J. Agric. Food Chem. 2019, 67, 8938–8949. [Google Scholar] [CrossRef]
- A Kennedy, J.; Taylor, A.W. Analysis of proanthocyanidins by high-performance gel permeation chromatography. J. Chromatogr. A 2003, 995, 99–107. [Google Scholar] [CrossRef]
- Xu, M.; Zha, Z.-J.; Qin, X.-L.; Zhang, X.-L.; Yang, C.-R.; Zhang, Y.-J. Phenolic Antioxidants from the Whole Plant of Phyllanthus urinaria. Chem. Biodivers. 2007, 4, 2246–2252. [Google Scholar] [CrossRef]
- Ozgunay, H.; Sari, O.; Tozan, M. Molecular investigation of valonea tannin. J. Am. Leather Chem. Assoc. 2007, 102, 154–157. [Google Scholar]
- Abdalla, S.; Pizzi, A.; Bahabri, F.; Ganash, A. Analysis of Valonia Oak (Quercus aegylops) Acorn Tannin and Wood Adhesives Application. BioResources 2015, 10, 7165–7177. [Google Scholar] [CrossRef] [Green Version]
- Chung, K.-T.; Wong, T.Y.; Wei, C.-I.; Huang, Y.-W.; Lin, Y. Tannins and Human Health: A Review. Crit. Rev. Food Sci. Nutr. 1998, 38, 421–464. [Google Scholar] [CrossRef]
- Chang, T.-S. An Updated Review of Tyrosinase Inhibitors. Int. J. Mol. Sci. 2009, 10, 2440–2475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldrop, G.L. A qualitative approach to enzyme inhibition. Biochem. Mol. Biol. Educ. 2009, 37, 11–15. [Google Scholar] [CrossRef]
- Si, C.-L.; Wu, L.; Shen, T.; Huang, X.-F.; Du, Z.-G.; Ren, X.-D.; Luo, X.-G.; Hu, W.-C. Recovery of Low-molecular Weight Galloyltannins from Agricultural Residue of Juglans sigillata Dode Seed Husks and their Tyrosinase Inhibitory Effect. BioResources 2014, 9, 2226–2236. [Google Scholar] [CrossRef] [Green Version]
- Whiteley, C.G. Enzyme kinetics: Partial and complete uncompetitive inhibition. Biochem. Educ. 2000, 28, 144–147. [Google Scholar] [CrossRef]
- Kubo, I.; Kinst-Hori, I.; Nihei, K.-I.; Soria, F.; Takasaki, M.; Calderón, J.S.; Céspedes, C.L. Tyrosinase Inhibitors from Galls of Rhus javanica Leaves and Their Effects on Insects. Zeitschrift für Naturforschung C 2003, 58, 719–725. [Google Scholar] [CrossRef]
- Van de Weert, M.; Stella, L. Fluorescence quenching and ligand binding: A critical discussion of a popular methodology. J. Mol. Struct. 2011, 998, 144–150. [Google Scholar] [CrossRef]
- Chai, W.-M.; Wei, M.-K.; Wang, R.; Deng, R.-G.; Zou, Z.-R.; Peng, Y.-Y. Avocado Proanthocyanidins as a Source of Tyrosinase Inhibitors: Structure Characterization, Inhibitory Activity, and Mechanism. J. Agric. Food Chem. 2015, 63, 7381–7387. [Google Scholar] [CrossRef] [PubMed]
- Deaville, E.R.; Green, R.J.; Mueller-Harvey, I.; Willoughby, A.I.; Frazier, R.A. Hydrolyzable Tannin Structures Influence Relative Globular and Random Coil Protein Binding Strengths. J. Agric. Food Chem. 2007, 55, 4554–4561. [Google Scholar] [CrossRef]
- Patil, S.; Sistla, S.; Jadhav, J. Interaction of small molecules with human tyrosinase: A surface plasmon resonance and molecular docking study. Int. J. Biol. Macromol. 2016, 92, 1123–1129. [Google Scholar] [CrossRef] [PubMed]
- McRae, J.M.; Kennedy, J.A. Wine and Grape Tannin Interactions with Salivary Proteins and Their Impact on Astringency: A Review of Current Research. Molecules 2011, 16, 2348–2364. [Google Scholar] [CrossRef] [Green Version]
- McRae, J.M.; Falconer, R.J.; Kennedy, J.A. Thermodynamics of Grape and Wine Tannin Interaction with Polyproline: Impiications for Red Wine Astringency. J. Agric. Food Chem. 2010, 58, 12510–12518. [Google Scholar] [CrossRef]
- Haslam, E. Natural Polyphenols (Vegetable Tannins) as Drugs: Possible Modes of Action. J. Nat. Prod. 1996, 59, 205–215. [Google Scholar] [CrossRef]
- Noh, H.; Lee, S.J.; Jo, H.-J.; Choi, H.W.; Hong, S.; Kong, K.-H. Histidine residues at the copper-binding site in human tyrosinase are essential for its catalytic activities. J. Enzym. Inhib. Med. Chem. 2020, 35, 726–732. [Google Scholar] [CrossRef] [PubMed]
- Ramsden, C.A.; Riley, P.A. Mechanistic studies of tyrosinase suicide inactivation. Arkivoc 2010, 2010, 260–274. [Google Scholar] [CrossRef]
- Ros, J.R.; Rodríguez-López, J.N.; García-Cánovas, F. Effect of l-ascorbic acid on the monophenolase activity of tyrosinase. Biochem. J. 1993, 295, 309–312. [Google Scholar] [CrossRef] [Green Version]
- Teng, B.; Petrie, P.; Smith, P.; Bindon, K. Comparison of water addition and early-harvest strategies to decrease alcohol concentration in Vitis vinifera cv. Shiraz wine: Impact on wine phenolics, tannin composition and colour properties. Aust. J. Grape Wine Res. 2020, 26, 158–171. [Google Scholar] [CrossRef]
- Jelesarov, I.; Bosshard, H.R. Isothermal titration calorimetry and differential scanning calorimetry as complementary tools to investigate the energetics of biomolecular recognition. J. Mol. Recognit. 1999, 12, 3–18. [Google Scholar] [CrossRef]
- Heitz, M.P.; Rupp, J.W. Determining mushroom tyrosinase inhibition by imidazolium ionic liquids: A spectroscopic and molecular docking study. Int. J. Biol. Macromol. 2018, 107, 1971–1981. [Google Scholar] [CrossRef] [PubMed]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Zeng, X.; Du, Z.; Ding, X.; Jiang, W. Characterization of the direct interaction between apple condensed tannins and cholesterol in vitro. Food Chem. 2020, 309, 125762. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Shift (ppm) | Assignment | Note |
|---|---|---|
| Valonea tannin | ||
| 164.41 | C7″ | Carbonyl C=O, HHDP |
| 163.98 | C7 | Carbonyl C=O, galloyl |
| 158.50 | C7′ | Carbonyl C=O, HHDP |
| 144.67 | C3 | Phenolic –OH, galloyl |
| 143.65 | C5, C3′, C5′, C3″, C5″ | Phenolic –OH, galloyl and HHDP |
| 135.92 | C4, C4′, C4″ | Phenolic –OH, galloyl and HHDP |
| 123.88 | C1, C1′, C1″ | Phenolic carbons, galloyl and HHDP |
| 114.89 | C2′ | Phenolic C-C bridges, HHDP |
| 113.63 | C2″ | Phenolic C-C bridges, HHDP |
| 109.61 | C2, C6 | Phenolic carbons, Galloyl |
| 70.04 | C-5‴ | Open glucose form |
| 62.71 | C-3‴, C-4‴, C-6‴ | Open glucose form |
| 57.40 | C-2‴ | Open glucose form |
| Gallic acid | ||
| 169.1 | C7 | Carbonyl C=O |
| 144.9 | C3, C5 | Phenolic –OH |
| 138.2 | C4 | Phenolic –OH |
| 120.5 | C1 | Phenolic carbons |
| 109.0 | C2, C6 | Phenolic carbons |
| Tannic acid | ||
| 169.7 | C7 | Carbonyl C=O |
| 165.4 | C7′ | Carbonyl C=O |
| 144.9 | C3, C5, C3′, C5′ | Phenolic –OH |
| 139.2 | C4 | Phenolic –OH |
| 138.0 | C4′ | Phenolic –OH |
| 121.3 | C1, C1′ | Phenolic carbons |
| 109.1 | C2, C6, C2′, C6′ | Phenolic carbons |
| 92.5 | C1″ | Closed glucoside |
| 71.9 | C2″ | Closed glucoside |
| 67.1 | C4″ | Closed glucoside |
| 66.2 | C6″ | Closed glucoside |
| 63.0 | C5″ | Closed glucoside |
| Sample | Valonea Tannin | Tannic Acid | Gallic Acid | Hydroquinone |
|---|---|---|---|---|
| IC50 (mM) 1 | 1.15 ± 0.37 | 4.00 ± 0.10 | 389.56 ± 4.77 | 1809.38 ± 5.74 |
| IC50 (g/L) | 3.50 ± 0.11 | 6.80 ± 0.17 | 66.23 ± 0.81 | 199.21 ± 0.63 |
| Fold 2 | -- | 3.4 | 339 |
| Valonea Tannin | Tannic Acid | Gallic Acid | |
|---|---|---|---|
| Inhibition mechanism | Reversible | Reversible | Reversible |
| Inhibition type | Competitive-noncompetitive mixed | Competitive | Competitive |
| Inhibition constants (mM) 2 | KIS = 1.68 ± 0.18 | KI = 1.97 ± 0.32 | KI = 56.58 ± 3.65 |
| KI = 0.51 ± 0.05 |
| Valonea Tannin | Tannic Acid | Gallic Acid | |
|---|---|---|---|
| Maximum emitting wavelength (nm) | 340 | 340 | 340 |
| Quenching efficiency (%) 2 | 74.3 ± 0.2 | 66.5 ± 0.4 | 24.2 ± 0.4 |
| Kq (×1013) | 2.85 ± 0.00 | 1.94 ± 0.00 | 0.28 ± 0.00 |
| Quenching type | Static | Static | Static |
| Linkage type | Non-covalent | Non-covalent | Non-covalent |
| Ka (L/mol) ×105 | 2.23 ± 0.00 | 1.32 ± 0.00 | 0.73 ± 0.00 |
| Valonea Tannin | Tannic Acid | Gallic Acid | |
|---|---|---|---|
| ΔH (×104 cal/mol) | −1.26 ± 0.44 (site 1) | −33.62 ± 1.11 | −6.34 ± 0.23 |
| −6.66 ± 0.22 (site 2) | |||
| TΔS (×104 cal/mol/deg) | −0.33 (site 1) | −32.79 | −5.67 |
| −5.90 (site 2) | |||
| K (×105) | 70.6 ± 3.0 (site 1) | 2.0 ± 0.5 | 0.9 ± 0.3 |
| 4.3 ± 0.8 (site 2) | |||
| n | 1.02 ± 0.04 (site 1) | 2.50 ± 0.05 | 8.75 ± 0.21 |
| 2.70 ± 0.05 (site 2) |
| Cu2+ Chelating (%) | DPPH· (IC50 mM) | ABTS·+ (IC50 mM) | |
|---|---|---|---|
| Valonea tannin | 56.61 ± 0.66 | 0.051 ± 0.002 | 0.040 ± 0.000 |
| Tannic acid | 56.24 ± 0.10 | 0.043 ± 0.001 | 0.043 ± 0.002 |
| Gallic acid | 52.17 ± 0.53 | 0.322 ± 0.004 | 0.253 ± 0.001 |
| Ascorbic acid | - | 0.683 ± 0.009 | 0.713 ± 0.012 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Liu, Y.; He, X.; Teng, B.; McRae, J.M. Valonea Tannin: Tyrosinase Inhibition Activity, Structural Elucidation and Insights into the Inhibition Mechanism. Molecules 2021, 26, 2747. https://doi.org/10.3390/molecules26092747
Liu J, Liu Y, He X, Teng B, McRae JM. Valonea Tannin: Tyrosinase Inhibition Activity, Structural Elucidation and Insights into the Inhibition Mechanism. Molecules. 2021; 26(9):2747. https://doi.org/10.3390/molecules26092747
Chicago/Turabian StyleLiu, Jiaman, Yuqing Liu, Xiaofeng He, Bo Teng, and Jacqui M. McRae. 2021. "Valonea Tannin: Tyrosinase Inhibition Activity, Structural Elucidation and Insights into the Inhibition Mechanism" Molecules 26, no. 9: 2747. https://doi.org/10.3390/molecules26092747
APA StyleLiu, J., Liu, Y., He, X., Teng, B., & McRae, J. M. (2021). Valonea Tannin: Tyrosinase Inhibition Activity, Structural Elucidation and Insights into the Inhibition Mechanism. Molecules, 26(9), 2747. https://doi.org/10.3390/molecules26092747