
Basic Principles in the Design of Spider Silk Fibers


{kind=link}
{kind=link}
{kind=link}
{kind=link}
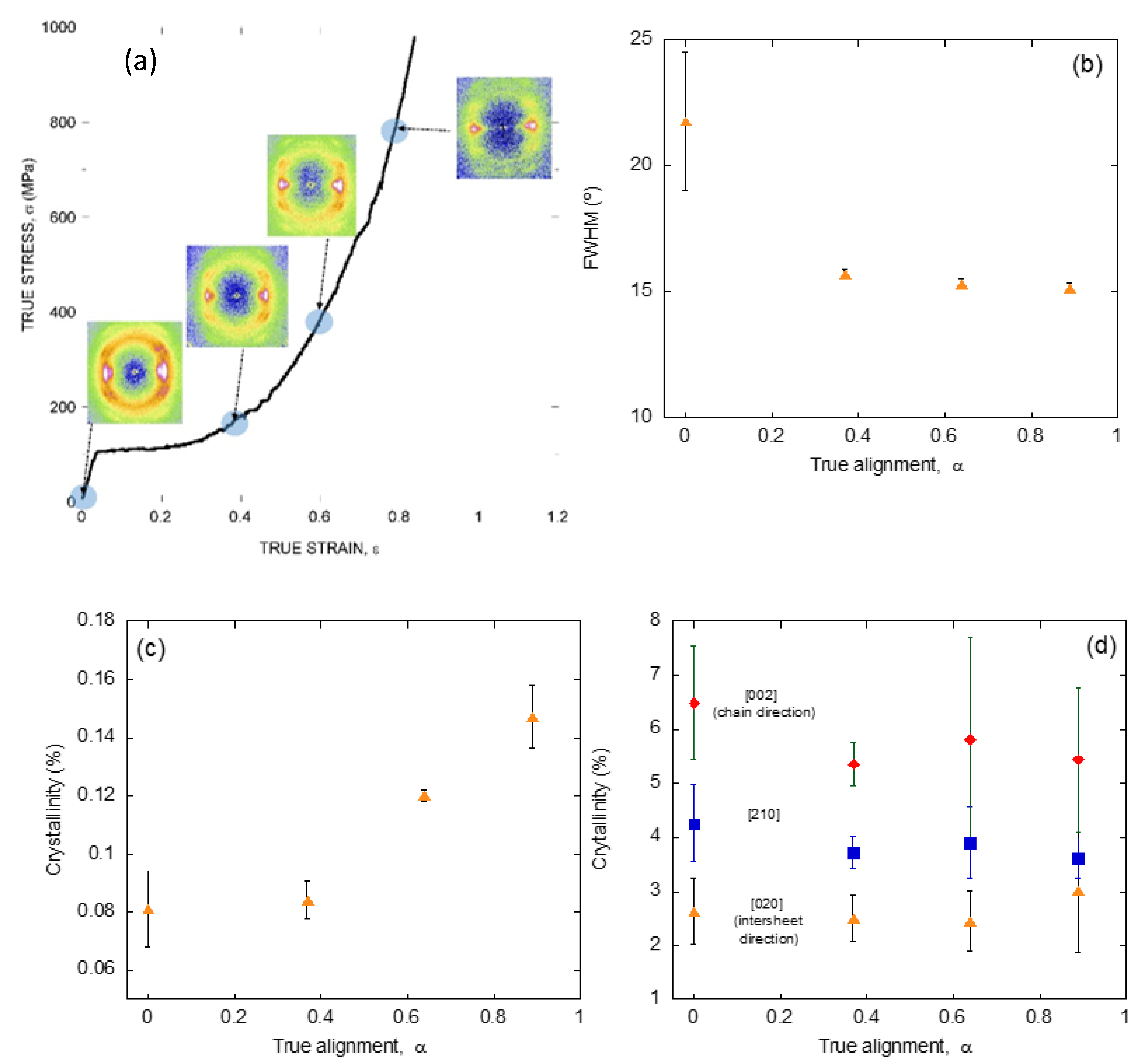{kind=link}
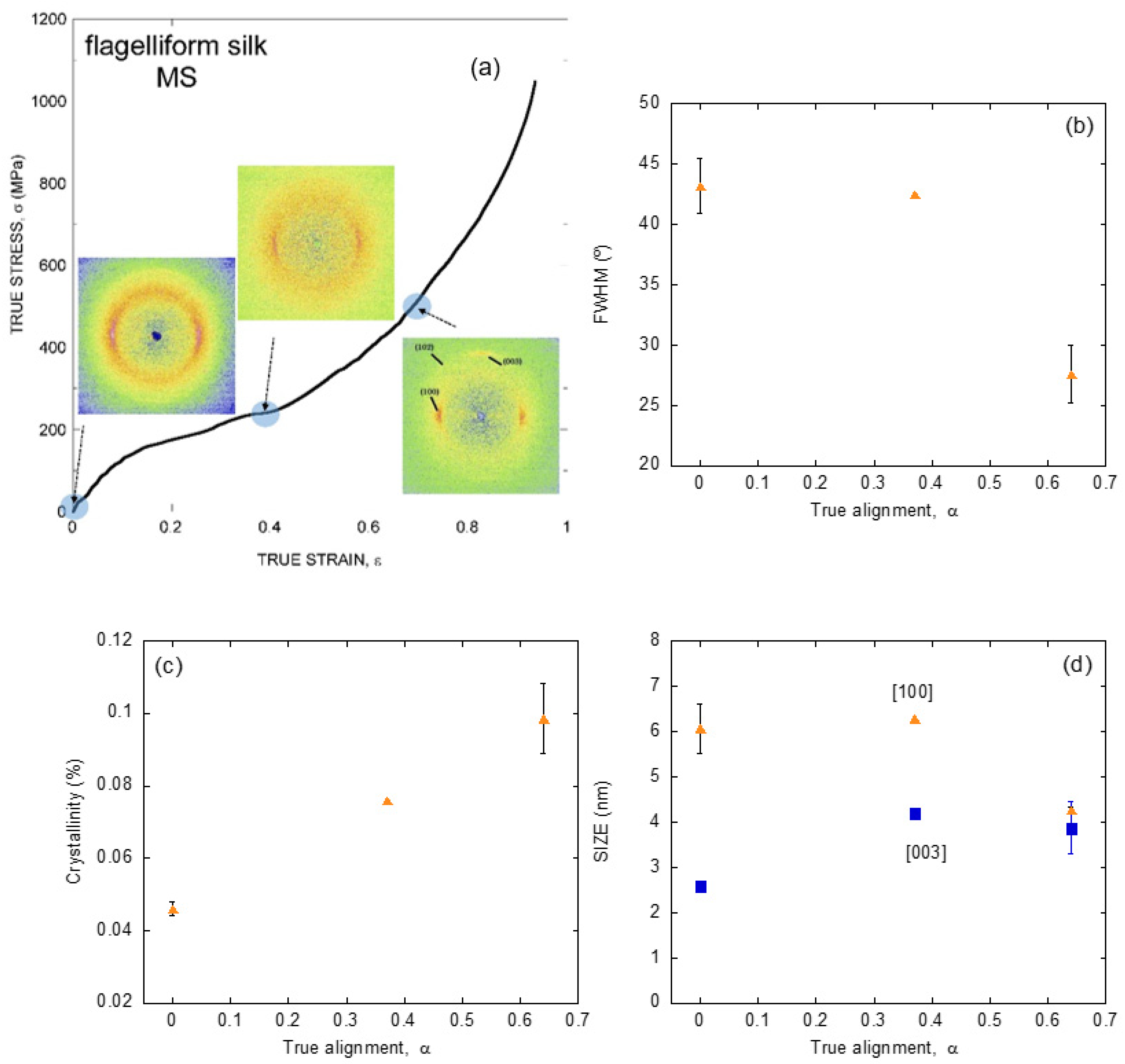{kind=link}
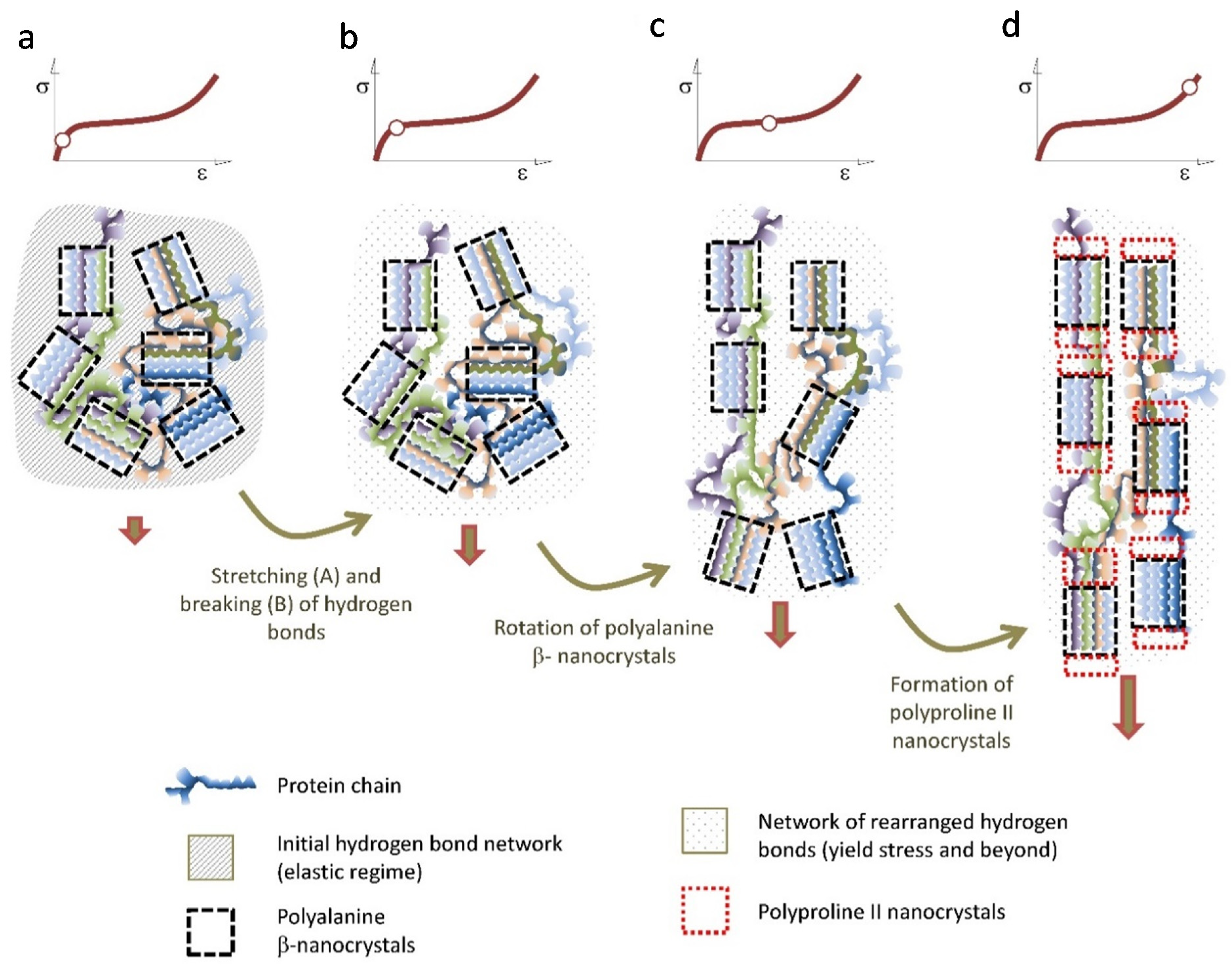{kind=link}
Abstract
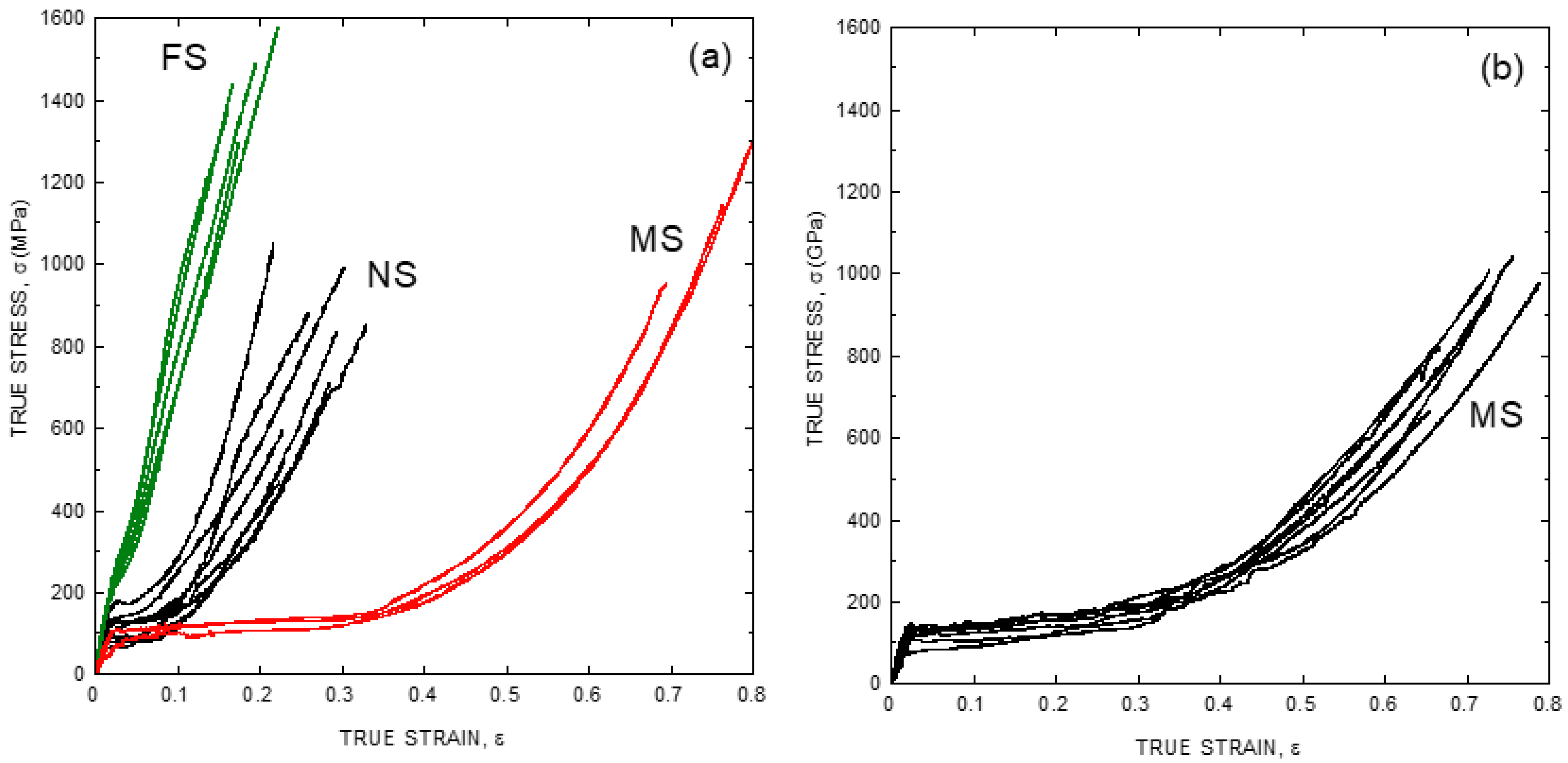
:1. The Singular Tensile Behavior of Spider Silk Fibers
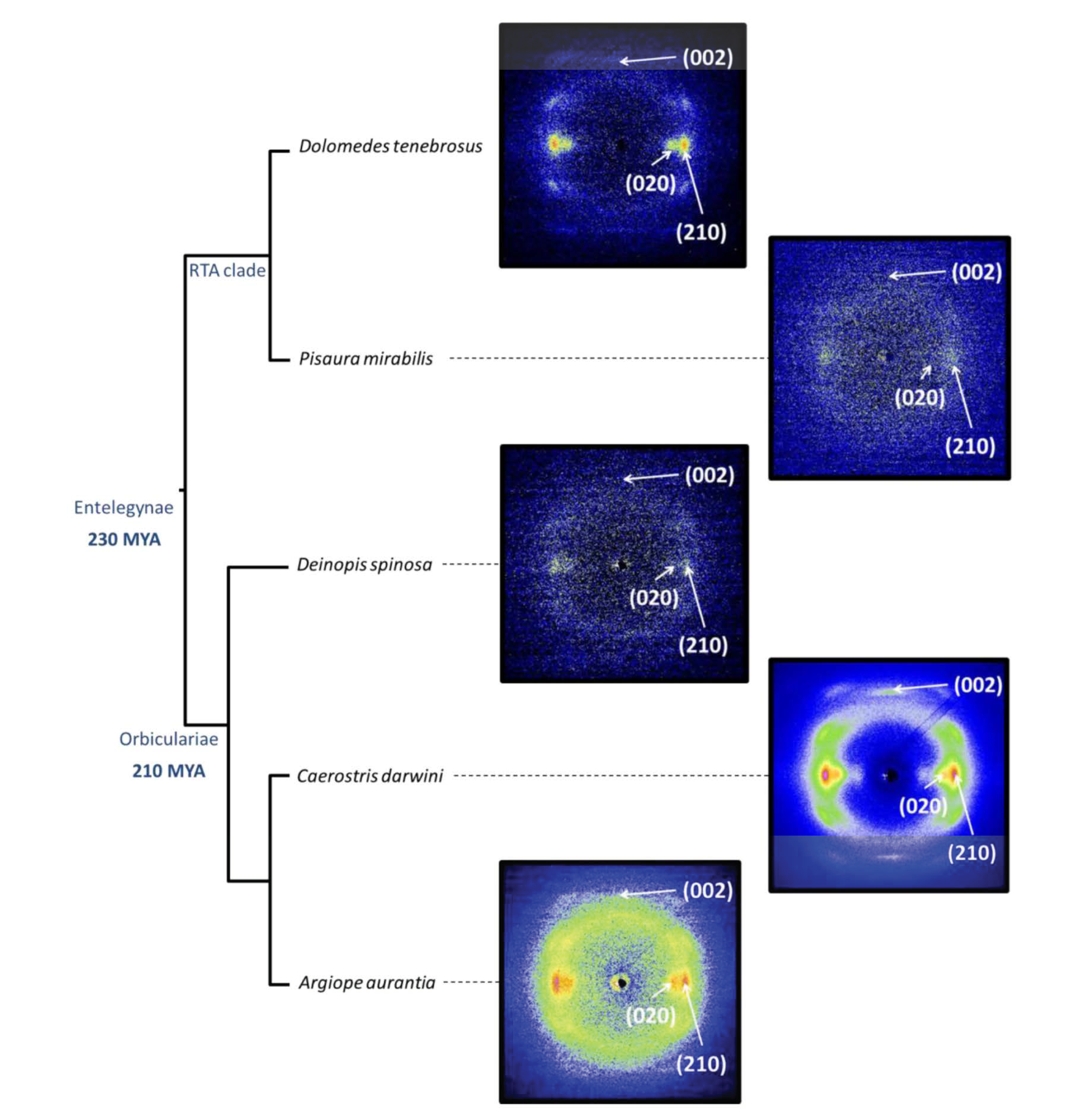
2. The Semicrystalline Organization of Spider Silk
3. Identifying the Microdeformation Mechanisms in Spider Silk
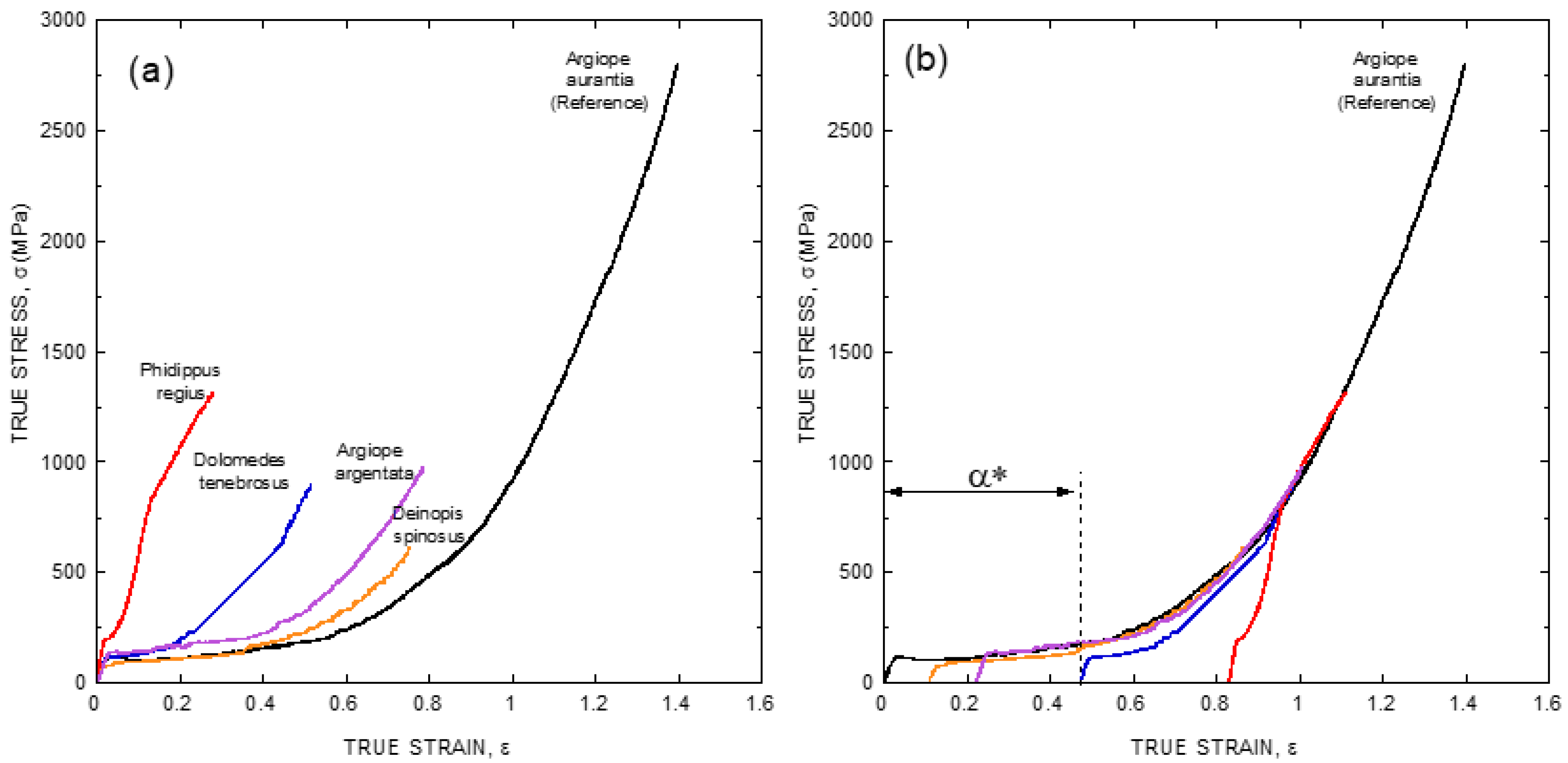
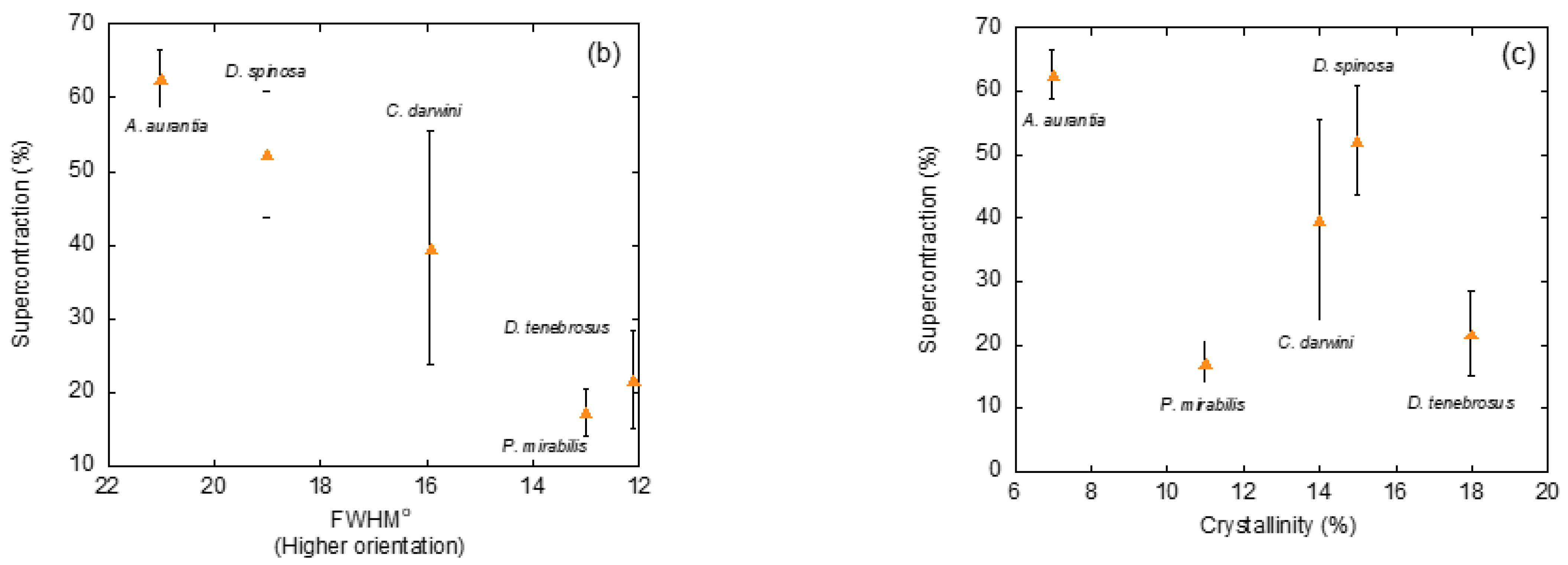
4. The Common Design Principles of Major Ampullate Gland Silks Spun by Entelegyane Spiders
5. Concluding Remarks
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gosline, J.M.; DeMont, M.; Denny, M.W. The structure and properties of spider silk. Endeavour 1986, 10, 37–43. [Google Scholar] [CrossRef]
- Kaplan, D.L.; Fossey, S.; Mello, C.M.; Arcidiacono, S.; Senecal, K.; Muller, W.; Stockwell, S.; Beckwitt, R.; Viney, C.; Kerkam, K. Biosynthesis and Processing of Silk Proteins. MRS Bull. 1992, 17, 41–47. [Google Scholar] [CrossRef]
- Bowen, C.H.; Dai, B.; Sargent, C.J.; Bai, W.; Ladiwala, P.; Feng, H.; Huang, W.; Kaplan, D.L.; Galazka, J.M.; Zhang, F. Recombinant Spidroins Fully Replicate Primary Mechanical Properties of Natural Spider Silk. Biomacromolecules 2018, 19, 3853–3860. [Google Scholar] [CrossRef] [Green Version]
- Work, R.W. The Force-Elongation Behavior of Web Fibers and Silks Forcibly Obtained from Orb-Web-Spinning Spiders. Text. Res. J. 1976, 46, 485–492. [Google Scholar] [CrossRef]
- Vollrath, F. General Properties of Some Spider Silks. ACS Symp. Ser. 1993, 544, 17–28. [Google Scholar] [CrossRef]
- Heim, M.; Keerl, D.; Scheibel, T. Spider Silk: From Soluble Protein to Extraordinary Fiber. Angew. Chem. Int. Ed. 2009, 48, 3584–3596. [Google Scholar] [CrossRef]
- Kiseleva, A.P.; Kiselev, G.O.; Nikolaeva, V.O.; Seisenbaeva, G.; Kessler, V.; Krivoshapkin, P.V.; Krivoshapkina, E.F. Hybrid Spider Silk with Inorganic Nanomaterials. Nanomatereials 2020, 10, 1853. [Google Scholar] [CrossRef]
- Andersson, M.; Johansson, J.; Rising, A. Silk Spinning in Silkworms and Spiders. Int. J. Mol. Sci. 2016, 17, 1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keten, S.; Xu, Z.; Ihle, B.; Buehler, M.J. Nanoconfinement controls stiffness, strength and mechanical toughness of beta-sheet crystals in silk. Nat. Mater. 2010, 9, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Dunaway, D.L.; Thiel, B.L.; Viney, C. Tensile mechanical property evaluation of natural and epoxide-treated silk fibers. J. Appl. Polym. Sci. 1995, 58, 675–683. [Google Scholar] [CrossRef]
- Madsen, B.; Shao, Z.Z.; Vollrath, F. Variability in the mechanical properties of spider silks on three levels: Interspecific, intraspecific and intraindividual. Int. J. Biol. Macromol. 1999, 24, 301–306. [Google Scholar] [CrossRef]
- Blamires, S.J.; Tseng, Y.-H.; Wu, C.-L.; Toft, S.; Raubenheimer, D.; Tso, I.-M. Spider web and silk performance landscapes across nutrient space. Sci. Rep. 2016, 6, 26383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blamires, S.J.; Nobbs, M.; Martens, P.J.; Tso, I.-M.; Chuang, W.-T.; Chang, C.-K.; Sheu, H.-S. Multiscale mechanisms of nutritionally induced property variation in spider silks. PLoS ONE 2018, 13, e0192005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J.; Vienneau-Hathaway, J.; Dendev, E.; Lan, M.; Ayoub, N.A. The common house spider, Parasteatoda tepidariorum, maintains silk gene expression on sub-optimal diet. PLoS ONE 2020, 15, e0237286. [Google Scholar] [CrossRef] [PubMed]
- Swanson, B.O.; Blackledge, T.A.; Beltrán, J.; Hayashi, C.Y. Variation in the material properties of spider dragline silk across species. Appl. Phys. A 2006, 82, 213–218. [Google Scholar] [CrossRef]
- Blackledge, T.A.; Hayashi, C.Y. Silken toolkits: Biomechanics of silk fibers spun by the orb web spider Argiope argentata (Fabricius 1775). J. Exp. Biol. 2006, 209, 2452–2461. [Google Scholar] [CrossRef] [Green Version]
- Blamires, S.J.; Blackledge, T.A.; Tso, I.-M. Physicochemical Property Variation in Spider Silk: Ecology, Evolution, and Synthetic Production. Annu. Rev. Entomol. 2017, 62, 443–460. [Google Scholar] [CrossRef]
- Work, R.W.; Emerson, P.D. An Apparatus and Technique for the Forcible Silking of Spiders. J. Arachnol. 1982, 10, 1–10. [Google Scholar]
- Pérez-Rigueiro, J.; Elices, M.; Llorca, J.; Viney, C. Tensile properties of Argiope trifasciata drag line silk obtained from the spider’s web. J. Appl. Polym. Sci. 2001, 82, 2245–2251. [Google Scholar] [CrossRef]
- Garrido, M.; Elices, M.; Viney, C.; Pérez-Rigueiro, J. Active control of spider silk strength: Comparison of drag line spun on vertical and horizontal surfaces. Polymer 2002, 43, 1537–1540. [Google Scholar] [CrossRef]
- Work, R.W. Dimensions, Birefringences, and Force-Elongation Behavior of Major and Minor Ampullate Silk Fibers from Orb-Web-Spinning Spiders—Effects of Wetting on these Properties. Text. Res. J. 1977, 47, 650–662. [Google Scholar] [CrossRef]
- Gosline, J.M.; Denny, M.W.; Demont, M.E. Spider silk as rubber. Nat. Cell Biol. 1984, 309, 551–552. [Google Scholar] [CrossRef]
- Gosline, J.M.; A Guerette, P.; Ortlepp, C.S.; Savage, K.N. The mechanical design of spider silks: From fibroin sequence to mechanical function. J. Exp. Biol. 1999, 202, 3295–3303. [Google Scholar] [PubMed]
- Savage, K.N.; Guerette, P.A.; Gosline, J.M. Supercontraction stress in spider webs. Biomacromolecules 2004, 5, 275–679. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Rigueiro, J.; Plaza, G.R.; Torres, F.G.; Hijar, A.; Hayashi, C.; Perea, G.; Elices, M.; Guinea, G. Supercontraction of dragline silk spun by lynx spiders (Oxyopidae). Int. J. Biol. Macromol. 2010, 46, 555–557. [Google Scholar] [CrossRef]
- Boutry, C.; Blackledge, T.A. Evolution of supercontraction in spider silk: Structure-function relationship from tarantulas to orb-weavers. J. Exp. Biol. 2010, 213, 3505–3514. [Google Scholar] [CrossRef] [Green Version]
- Elices, M.; Pérez-Rigueiro, J.; Plaza, G.R.; Guinea, G.V. Recovery in spider silk fibers. J. Appl. Polym. Sci. 2004, 92, 3537–3541. [Google Scholar] [CrossRef]
- Guinea, G.V.; Elices, M.; Pérez-Rigueiro, J.; Plaza, G.R. Stretching of supercontracted fibers: A link between spinning and the variability of spider silk. J. Exp. Biol. 2005, 208, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Madurga, R.; Plaza, G.R.; Blackledge, T.A.; Guinea, G.; Elices, M.; Pérez-Rigueiro, J. Material properties of evolutionary diverse spider silks described by variation in a single structural parameter. Sci. Rep. 2016, 6, 18991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guinea, G.V.; Pérez-Rigueiro, J.; Plaza, G.R.; Elices, M. Volume constancy during stretching of spider silk. Biomacromolecules 2006, 7, 2173–2177. [Google Scholar] [CrossRef]
- Garrote, J.; Ruiz, V.; Troncoso, O.P.; Torres, F.G.; Arnedo, M.; Elices, M.; Guinea, G.V.; Pérez-Rigueiro, J. Application of the Spider Silk Standardization Initiative ((SI)-I-3) methodology to the characterization of major ampullate gland silk fibers spun by spiders from Pantanos de Villa wetlands (Lima, Peru). J. Mech. Behav. Biomed. Mater. 2020, 111, 104023. [Google Scholar] [CrossRef]
- Babb, P.L.; Lahens, N.F.; Correa-Garhwal, S.M.; Nicholson, D.N.; Kim, E.J.; Hogenesch, J.B.; Kuntner, M.; Higgins, L.; Hayashi, C.Y.; Agnarsson, L.H.I.; et al. The Nephila clavipes genome highlights the diversity of spider silk genes and their complex expression. Nat. Genet. 2017, 49, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Craig, H.C.; Piorkowski, D.; Nakagawa, S.; Kasumovic, M.M.; Blamires, S.J. Meta-analysis reveals materiomic relationships in major ampullate silk across the spider phylogeny. J. R. Soc. Interface 2020, 17, 20200471. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Lewis, R.V. Structure of a protein superfiber: Spider dragline silk. Proc. Natl. Acad. Sci. USA 1990, 87, 7120–7124. [Google Scholar] [CrossRef] [Green Version]
- Gatesy, J.; Hayashi, C.; Motriuk, D.; Woods, J.; Lewis, R. Extreme diversity, conservation, and convergence of spider silk fibroin sequences. Science 2001, 291, 2603–2605. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Rigueiro, J.; Ruiz, V.; Cenis, J.L.; Elices, M.; Guinea, G.V. Lessons from Spider and Silkworm Silk Guts. Front. Mater. 2020, 7, 46. [Google Scholar] [CrossRef]
- Warwicker, J. The Crystal Structure of Silk Fibroins. Trans. Faraday Soc. 1954, 50, 1011–1012. [Google Scholar] [CrossRef]
- Warwicker, J.O. Comparative Studies of Fibroins 2. Crystal Structures of various Fibroins. J. Mol. Biol. 1960, 2, 350–362. [Google Scholar] [CrossRef]
- Marsh, R.E.; Corey, R.B.; Pauling, L. An investigation of the structure of silk fibroin. Biochim. Biophys. Acta BBA Bioenerg. 1955, 16, 1–34. [Google Scholar] [CrossRef]
- Biology Analysis Group; Xia, Q.; Zhou, Z.; Lu, C.; Cheng, D.; Dai, F.-Y.; Liu, B.; Zhao, P.; Zha, X.; Cheng, T.; et al. A Draft Sequence for the Genome of the Domesticated Silkworm (Bombyx mori). Science 2004, 306, 1937–1940. [Google Scholar] [CrossRef] [Green Version]
- Asakura, T.; Suzuki, Y.; Nakazawa, Y.; Yazawa, K.; Holland, G.P.; Yarger, J.L. Silk structure studied with nuclear magnetic resonance. Prog. Nucl. Magn. Reson. Spectrosc. 2013, 69, 23–68. [Google Scholar] [CrossRef] [PubMed]
- Holland, G.P.; Creager, M.S.; Jenkins, J.E.; Lewis, R.V.; Yarger, J.L. Determining Secondary Structure in Spider Dragline Silk by Carbon−Carbon Correlation Solid-State NMR Spectroscopy. J. Am. Chem. Soc. 2008, 130, 9871–9877. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, J.E.; Creager, M.S.; Butler, E.B.; Lewis, R.V.; Yarger, J.L.; Holland, G.P. Holland, Solid-state NMR evidence for elastin-like beta-turn structure in spider dragline silk. Chem. Commun. 2010, 46, 6714–6716. [Google Scholar] [CrossRef]
- Yang, Z.; Liivak, O.; Seidel, A.; LaVerde, G.; Zax, D.B.; Jelinski, L.W. Supercontraction and Backbone Dynamics in Spider Silk: 13C and2H NMR Studies. J. Am. Chem. Soc. 2000, 122, 9019–9025. [Google Scholar] [CrossRef]
- Asakura, T.; Tasei, Y.; Aoki, A.; Nishimura, A. Mixture of Rectangular and Staggered Packing Arrangements of Polyalanine Region in Spider Dragline Silk in Dry and Hydrated States as Revealed by 13C NMR and X-ray Diffraction. Macromolecules 2018, 51, 1058–1068. [Google Scholar] [CrossRef]
- Riekel, C.; Bränden, C.; Craig, C.; Ferrero, C.; Heidelbach, F.; Müller, M. Aspects of X-ray diffraction on single spider fibers. Int. J. Biol. Macromol. 1999, 24, 179–186. [Google Scholar] [CrossRef]
- Madurga, R.; Blackledge, T.A.; Perea, B.; Plaza, G.R.; Riekel, C.; Burghammer, M.; Elices, M.; Guinea, G.; Pérez-Rigueiro, J. Persistence and variation in microstructural design during the evolution of spider silk. Sci. Rep. 2015, 5, 14820. [Google Scholar] [CrossRef] [Green Version]
- Plaza, G.R.; Pérez-Rigueiro, J.; Riekel, C.; Perea, G.B.; Agulló-Rueda, F.; Burghammer, M.; Guinea, G.V.; Elices, M. Relationship between microstructure and mechanical properties in spider silk fibers: Identification of two regimes in the microstructural changes. Soft Matter 2012, 8, 6015–6026. [Google Scholar] [CrossRef] [Green Version]
- Work, R.W.; Morosoff, N. A Physico-Chemical Study of the Supercontraction of Spider Major Ampullate Silk Fibers. Text. Res. J. 1982, 52, 349–356. [Google Scholar] [CrossRef]
- Termonia, Y. Molecular Modeling of Spider Silk Elasticity. Macromolecules 1994, 27, 7378–7381. [Google Scholar] [CrossRef]
- Termonia, Y. Molecular modelling of the stress/strainbehaviour of spider dragline. In Structural Biological Materials; Elices, M., Ed.; Pergamon Press: Amsterdam, The Netherlands, 2000; pp. 335–349. [Google Scholar]
- Guinea, G.V.; Elices, M.; Plaza, G.R.; Perea, G.B.; Daza, R.; Riekel, C.; Agulló-Rueda, F.; Hayashi, C.; Zhao, Y.; Pérez-Rigueiro, J. Minor Ampullate Silks from Nephila and Argiope Spiders: Tensile Properties and Microstructural Characterization. Biomacromolecules 2012, 13, 2087–2098. [Google Scholar] [CrossRef]
- Vollrath, F.; Edmonds, D.T. Modulation of the mechanical properties of spider silk by coating with water. Nat. Cell Biol. 1989, 340, 305–307. [Google Scholar] [CrossRef]
- Vollrath, F.; Edmonds, D. Elastic Properties of Spiders Capture Silk—Reply. Naturwissenschaften 1995, 82, 379–380. [Google Scholar]
- Opell, B.D.; Karinshak, S.E.; Sigler, M.A. Humidity affects the extensibility of an orb-weaving spider’s viscous thread droplets. J. Exp. Biol. 2011, 214, 2988–2993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Opell, B.D.; Hendricks, M.L. Adhesive recruitment by the viscous capture threads of araneoid orb-weaving spiders. J. Exp. Biol. 2007, 210, 553–560. [Google Scholar] [CrossRef] [Green Version]
- Guinea, G.V.; Cerdeira, M.; Plaza, G.R.; Elices, M.; Pérez-Rigueiro, J. Recovery in Viscid Line Fibers. Biomacromolecules 2010, 11, 1174–1179. [Google Scholar] [CrossRef] [PubMed]
- Perea, G.B.; Riekel, C.; Guinea, G.V.; Madurga, R.; Daza, R.; Burghammer, M.; Hayashi, C.M.; Elices, M.; Plaza, G.R.; Pérez-Rigueiro, J. Identification and dynamics of polyglycine II nanocrystals in Argiope trifasciata flagelliform silk. Sci. Rep. 2013, 3, 3061. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, M.-E.; Lefèvre, T.; Pézolet, M. Conformation and Orientation of Proteins in Various Types of Silk Fibers Produced by Nephila clavipes Spiders. Biomacromolecules 2009, 10, 2945–2953. [Google Scholar] [CrossRef]
- Gosline, J.M.; Pollak, C.C.; Guerette, P.A.; Cheng, A.; Demont, M.E.; Denny, M.W. Elastomeric Network Models for the Frame and Viscid Silks from the Orb Web of the Spider Araneus diadematus. ACS Symp. Ser. 1993, 544, 328–341. [Google Scholar] [CrossRef]
- Gosline, J.; Lillie, M.; Carrington, E.; Guerette, P.; Ortlepp, C.; Savage, K. Elastic proteins: Biological roles and mechanical properties. Philos. Trans. R. Soc. B Biol. Sci. 2002, 357, 121–132. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Sponner, A.; Porter, D.; Vollrath, F. Proline and Processing of Spider Silks. Biomacromolecules 2008, 9, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Blackledge, T.A.; Pérez-Rigueiro, J.; Plaza, G.R.; Perea, B.; Navarro, A.; Guinea, G.V.; Elices, M. Sequential origin in the high performance properties of orb spider dragline silk. Sci. Rep. 2012, 2, 782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nova, A.; Keten, S.; Pugno, N.M.; Redaelli, A.; Buehler, M.J. Molecular and Nanostructural Mechanisms of Deformation, Strength and Toughness of Spider Silk Fibrils. Nano Lett. 2010, 10, 2626–2634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keten, S.; Buehler, M.J. Nanostructure and molecular mechanics of spider dragline silk protein assemblies. J. R. Soc. Interface 2010, 7, 1709–1721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perea, G.B.; Solanas, C.; Plaza, G.R.; Guinea, G.V.; Jorge, I.; Vázquez, J.; Mateos, J.M.P.; Marí-Buyé, N.; Elices, M.; Pérez-Rigueiro, J. Unexpected behavior of irradiated spider silk links conformational freedom to mechanical performance. Soft Matter 2015, 11, 4868–4878. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Rigueiro, J.; Elices, M.; Plaza, G.R.; Guinea, G.V. Similarities and Differences in the Supramolecular Organization of Silkworm and Spider Silk. Macromolecules 2007, 40, 5360–5365. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Rigueiro, J.; Elices, M.; Plaza, G.R.; Guinea, G.V. Basic Principles in the Design of Spider Silk Fibers. Molecules 2021, 26, 1794. https://doi.org/10.3390/molecules26061794
Pérez-Rigueiro J, Elices M, Plaza GR, Guinea GV. Basic Principles in the Design of Spider Silk Fibers. Molecules. 2021; 26(6):1794. https://doi.org/10.3390/molecules26061794
Chicago/Turabian StylePérez-Rigueiro, José, Manuel Elices, Gustavo R. Plaza, and Gustavo V. Guinea. 2021. "Basic Principles in the Design of Spider Silk Fibers" Molecules 26, no. 6: 1794. https://doi.org/10.3390/molecules26061794
APA StylePérez-Rigueiro, J., Elices, M., Plaza, G. R., & Guinea, G. V. (2021). Basic Principles in the Design of Spider Silk Fibers. Molecules, 26(6), 1794. https://doi.org/10.3390/molecules26061794