
Nanoscale Material Heterogeneity of Glowworm Capture Threads Revealed by AFM

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Tapping Mode AFM and QNM Modulus Mapping of Glowworm Threads
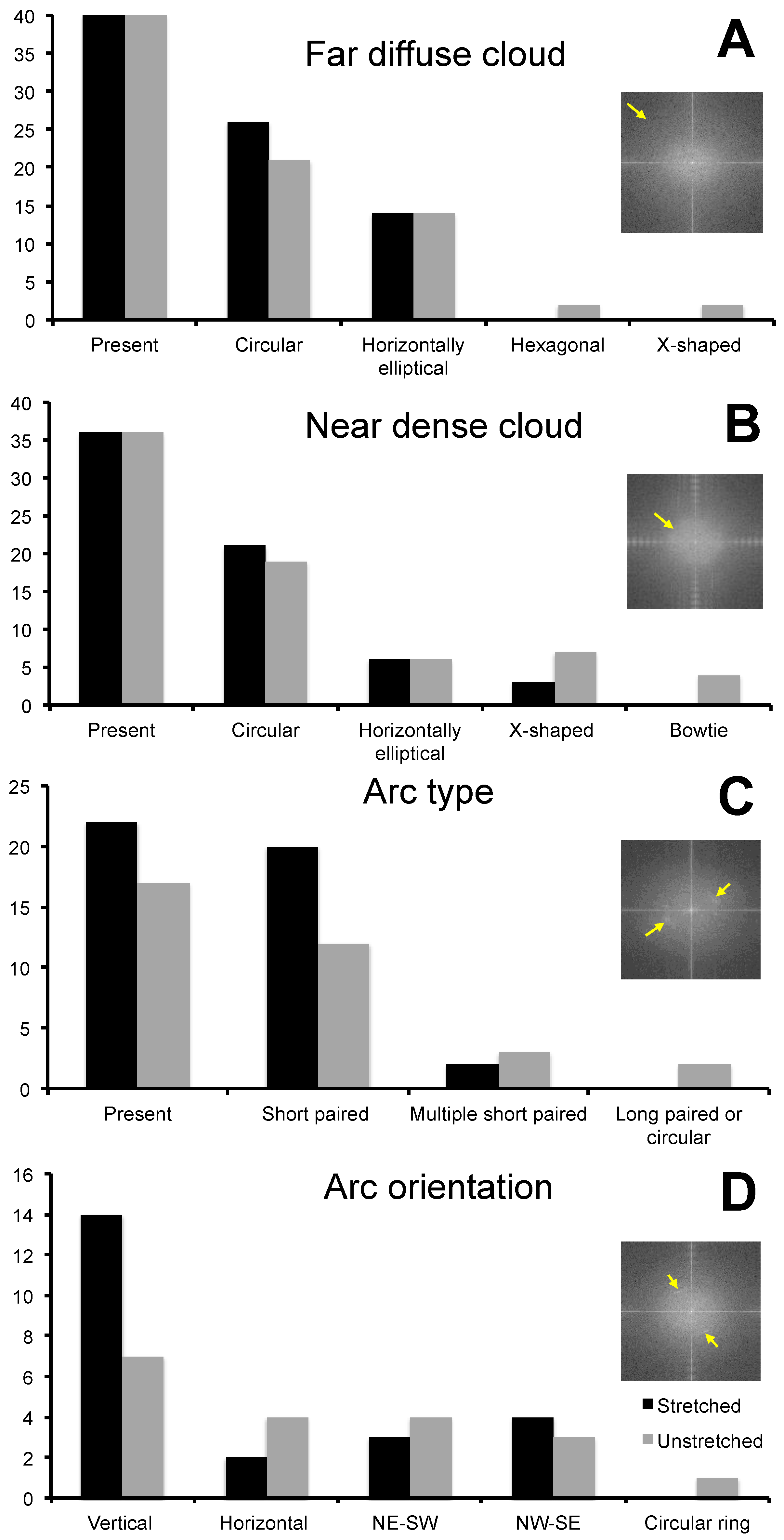
2.2. Fast Fourier Transform Analysis of Phase Angle Images of Glowworm Threads
3. Discussion
4. Materials and Methods
4.1. Capture Thread Collection
4.2. Atomic Force Microscopy
4.3. Fast Fourier Transform Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Gorb, S.N. Biological attachment devices: Exploring nature’s diversity for biomimetics. Philos. Trans. R. Soc. A 2008, 366, 1557–1574. [Google Scholar] [CrossRef] [PubMed]
- Von Byern, J.; Grunwald, I. Biological Adhesive Systems. From Nature to Technical and Medical Application; Springer: Wien, Austria, 2010; ISBN 9783709101414. [Google Scholar]
- Favi, P.M.; Yi, S.; Lenaghan, S.C.; Xia, L.; Zhang, M. Inspiration from the natural world: From bio-adhesives to bio-inspired adhesives. J. Adhes. Sci. Technol. 2014, 28, 290–319. [Google Scholar] [CrossRef]
- von Byern, J.; Müller, C.; Voigtländer, K.; Dorrer, V.; Marchetti–Deschmann, M.; Flammang, P.; Mayer, G. Examples of Bioadhesives for Defence and Predation. In Functional Surfaces in Biology III. Biologically-Inspired Systems; Gorb, S.N., Gorb, E.V., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; Volume 10, pp. 141–191. ISBN 9783319741444. [Google Scholar]
- Agnarsson, I.; Dhinojwala, A.; Sahni, V.; Blackledge, T.A. Spider silk as a novel high performance biomimetic muscle driven by humidity. J. Exp. Biol. 2009, 212, 1990–1994. [Google Scholar] [CrossRef] [Green Version]
- Sahni, V.; Blackledge, T.A.; Dhinojwala, A. A review on spider silk adhesion. J. Adhes. 2011, 87, 595–614. [Google Scholar] [CrossRef]
- Sahni, V.; Dhinojwala, A.; Opell, B.D.; Blackledge, T.A. Prey capture adhesives produced by orb-weaving spiders. In Biotechnology of Silk; Springer: Dordrecht, The Netherlands, 2014; pp. 203–217. ISBN 9789400771185. [Google Scholar]
- Piorkowski, D.; Liao, C.P.; Joel, A.C.; Wu, C.L.; Doran, N.; Blamires, S.J.; Pugno, N.M.; Tso, I.M. Adhesion of spider cribellate silk enhanced in high humidity by mechanical plasticization of the underlying fiber. J. Mech. Behav. Biomed. Mater. 2021, 114, 104200. [Google Scholar] [CrossRef]
- Piorkowski, D.; Blackledge, T.A.; Liao, C.P.; Doran, N.E.; Wu, C.L.; Blamires, S.J.; Tso, I.M. Humidity-dependent mechanical and adhesive properties of Arachnocampa tasmaniensis capture threads. J. Zool. 2018, 305, 256–266. [Google Scholar] [CrossRef]
- von Byern, J.; Chandler, P.; Merritt, D.; Adlassnig, W.; Stringer, I.; Meyer-Rochow, V.B.; Kovalev, A.; Dorrer, V.; Dimartino, S.; Marchetti-Deschmann, M.; et al. Biomechanical properties of fishing lines of the glowworm Arachnocampa luminosa (Diptera; Keroplatidae). Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Ashton, N.N.; Roe, D.R.; Weiss, R.B.; Cheatham, T.E., III; Stewart, R.J. Self-tensioning aquatic caddisfly silk: Ca2+-dependent structure, strength, and load cycle hysteresis. Biomacromolecules 2013, 14, 3668–3681. [Google Scholar] [CrossRef] [PubMed]
- Fudge, D.S.; Gardner, K.H.; Forsyth, V.T.; Riekel, C.; Gosline, J.M. The mechanical properties of hydrated intermediate filaments: Insights from hagfish slime threads. Biophys. J. 2003, 85, 2015–2027. [Google Scholar] [CrossRef] [Green Version]
- Fudge, D.S.; Levy, N.; Chiu, S.; Gosline, J.M. Composition, morphology and mechanics of hagfish slime. J. Exp. Biol. 2005, 208, 4613–4625. [Google Scholar] [CrossRef] [Green Version]
- Blamires, S.J. Biomechanical costs and benefits of sit-and-wait foraging traps. Isr. J. Ecol. Evol. 2020, 66, 5–14. [Google Scholar] [CrossRef]
- Wolff, J.O.; Wells, D.; Reid, C.R.; Blamires, S.J. Clarity of objectives and working principles enhances the success of biomimetic programs. Bioinspir. Biomim. 2017, 12, 051001. [Google Scholar] [CrossRef] [PubMed]
- Amarpuri, G.; Chaurasia, V.; Jain, D.; Blackledge, T.A.; Dhinojwala, A. Ubiquitous distribution of salts and proteins in spider glue enhances spider silk adhesion. Sci. Rep. 2015, 5, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolff, J.O.; von Byern, J.; Piorkowski, D.; Fang, J.; Wang, X.; Adler, L.; Thomas, D.S.; Hook, J.M.; Blamires, S. Adhesive droplets of glowworm snares (Keroplatidae: Arachnocampa spp.) are a complex mix of organic compounds. Front. Mech. Eng. 2021, 7, 27. [Google Scholar] [CrossRef]
- Opell, B.D.; Hendricks, M.L. Adhesive recruitment by the viscous capture threads of araneoid orb-weaving spiders. J. Exp. Biol. 2007, 210, 553–560. [Google Scholar] [CrossRef] [Green Version]
- Meyer, A.; Pugno, N.M.; Cranford, S.W. Compliant threads maximize spider silk connection strength and toughness. J. R. Soc. Interface 2014, 11, 20140561. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Chang, Z.; Guo, H.Y.; Fang, W.; Li, Q.; Zhao, H.P.; Feng, X.Q.; Gao, H. Synergistic adhesion mechanisms of spider capture silk. J. R. Soc. Interface 2018, 15, 20170894. [Google Scholar] [CrossRef] [PubMed]
- Perea, G.B.; Riekel, C.; Guinea, G.V.; Madurga, R.; Daza, R.; Burghammer, M.; Hayashi, C.; Elices, M.; Plaza, G.R.; Pérez-Rigueiro, J. Identification and dynamics of polyglycine II nanocrystals in Argiope trifasciata flagelliform silk. Sci. Rep. 2013, 3, 1–6. [Google Scholar] [CrossRef]
- Edwards, F.W. A note on the “New Zealand glow-worm” (Diptera: Mycotophilidae). Ann. Mag. Nat. Hist. 1924, 9, 175–179. [Google Scholar] [CrossRef]
- Richards, A.M. Observations on the New Zealand glow-worm Arachnocampa luminosa (Skuse) 1890. Trans. Proc. R. Soc. N. Z. 1960, 8, 559–574. [Google Scholar]
- von Byern, J.; Dorrer, V.; Merritt, D.J.; Chandler, P.; Stringer, I.; Marchetti-Deschmann, M.; McNaughton, A.; Cyran, N.; Thiel, K.; Noeske, M.; et al. Characterization of the fishing lines in Titiwai (=Arachnocampa luminosa Skuse, 1890) from New Zealand and Australia. PLoS ONE 2016, 11, e0162687. [Google Scholar] [CrossRef]
- Meyer-Rochow, V.B. Glowworms: A review of Arachnocampa spp. and kin. Luminescence 2007, 22, 251–265. [Google Scholar] [CrossRef] [PubMed]
- Willis, R.E.; White, C.R.; Merritt, D.J. Using light as a lure is an efficient predatory strategy in Arachnocampa flava, an Australian glowworm. J. Comp. Physiol. B 2011, 181, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.A.; Weisman, S.; Trueman, H.E.; Merritt, D.J.; Sutherland, T.D. The other prey-capture silk: Fibres made by glow-worms (Diptera: Keroplatidae) comprise cross-β-sheet crystallites in an abundant amorphous fraction. Comp. Biochem. Physiol. B 2015, 187, 78–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucas, F.; Shaw, J.T.B.; Smith, S.G. Comparative studies of fibroins: I. The amino acid composition of various fibroins and its significance in relation to their crystal structure and taxonomy. J. Mol. Biol. 1960, 2, 339–349. [Google Scholar] [CrossRef]
- Jin, H.J.; Kaplan, D. Mechanism of silk processing in insects and spiders. Nature 2003, 424, 1057–1061. [Google Scholar] [CrossRef]
- Bauer, F.; Bertinetti, L.; Masic, A.; Scheibel, T. Dependence of mechanical properties of lacewing egg stalks on relative humidity. Biomacromolecules 2012, 13, 3730–3735. [Google Scholar] [CrossRef]
- Garcia, R.; Perez, R. Dynamic atomic force microscopy methods. Surf. Sci. Rep. 2002, 47, 197–302. [Google Scholar] [CrossRef]
- Proksch, R.; Kocun, M.; Hurley, D.; Viani, M.; Labuda, A.; Meinhold, W.; Bemis, J. Practical loss tangent imaging with amplitude-modulated atomic force microscopy. J. Appl. Phys. 2016, 119, 134901. [Google Scholar] [CrossRef]
- Magonov, S.N.; Elings, V.; Whangbo, M.H. Phase imaging and stiffness in tapping-mode atomic force microscopy. Surf. Sci. 1997, 375, L385–L391. [Google Scholar] [CrossRef]
- Bar, G.; Thomann, Y.; Brandsch, R.; Cantow, H.-J.; Whangbo, M.-H. Factors Affecting the Height and Phase Images in Tapping Mode Atomic Force Microscopy. Study of Phase-Separated Polymer Blends of Poly(ethene-co-styrene) and Poly(2,6-dimethyl-1,4-phenylene oxide). Langmuir 1997, 13, 3807–3812. [Google Scholar] [CrossRef]
- Holland, N.B.; Marchant, R.E. Individual plasma proteins detected on rough biomaterials by phase imaging AFM. J. Biomed. Mater. Res. 2000, 51, 307–315. [Google Scholar] [CrossRef]
- Ciccotti, M.; George, M.; Ranieri, V.; Wondraczek, L.; Marliere, C. Dynamic condensation of water at crack tips in fused silica glass. J. Noncryst. Solids 2008, 354, 564–568. [Google Scholar] [CrossRef] [Green Version]
- Bhaduri, S.; Mallick, S.; Shiradkar, N.; Kottantharayil, A. Characterization of reliability of anti-soiling coatings using tapping mode-AFM phase imaging. J. Renew. Sustain. Energy 2021, 13, 023702. [Google Scholar] [CrossRef]
- Stan, G.; King, S.W. Atomic force microscopy for nanoscale mechanical property characterization. J. Vac. Sci. Technol. B 2020, 38, 060801. [Google Scholar] [CrossRef]
- Lo, Y.S.; Huefner, N.D.; Chan, W.S.; Dryden, P.; Hagenhoff, B.; Beebe, T.P. Organic and inorganic contamination on commercial AFM cantilevers. Langmuir 1999, 15, 6522–6526. [Google Scholar] [CrossRef]
- Du, N.; Liu, X.Y.; Narayanan, J.; Li, L.; Lim, M.L.M.; Li, D. Design of superior spider silk: From nanostructure to mechanical properties. Biophys. J. 2006, 91, 4528–4535. [Google Scholar] [CrossRef] [Green Version]
- Schäfer, A.; Vehoff, T.; Glišović, A.; Salditt, T. Spider silk softening by water uptake: An AFM study. Eur. Biophys. J. 2008, 37, 197–204. [Google Scholar] [CrossRef]
- Strzelecki, J.; Strzelecka, J.; Mikulska, K.; Tszydel, M.; Balter, A.; Nowak, W. Nanomechanics of new materials—AFM and computer modelling studies of trichoptera silk. Open Phys. 2011, 9, 482–491. [Google Scholar] [CrossRef]
- Wang, Q.; Ling, S.; Yao, Q.; Li, Q.; Hu, D.; Dai, Q.; Weitz, D.A.; Kaplan, D.L.; Buehler, M.J.; Zhang, Y. Observations of 3 nm silk nanofibrils exfoliated from natural silkworm silk fibers. ACS Mater. Lett. 2020, 2, 153–160. [Google Scholar] [CrossRef]
- Zhong, J.; Ma, M.; Li, W.; Zhou, J.; Yan, Z.; He, D. Self-assembly of regenerated silk fibroin from random coil nanostructures to antiparallel β-sheet nanostructures. Biopolymers 2014, 101, 1181–1192. [Google Scholar] [CrossRef]
- Zhong, J.; Liu, X.; Wei, D.; Yan, J.; Wang, P.; Sun, G.; He, D. Effect of incubation temperature on the self-assembly of regenerated silk fibroin: A study using AFM. Int. J. Biol. Macromol. 2015, 76, 195–202. [Google Scholar] [CrossRef]
- Wang, M.; Jin, H.J.; Kaplan, D.L.; Rutledge, G.C. Mechanical properties of electrospun silk fibers. Macromolecules 2004, 37, 6856–6864. [Google Scholar] [CrossRef]
- Wang, Z.; Cang, Y.; Kremer, F.; Thomas, E.L.; Fytas, G. Determination of the Complete Elasticity of Nephila pilipes Spider Silk. Biomacromolecules 2020, 21, 1179–1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sponner, A.; Vater, W.; Monajembashi, S.; Unger, E.; Grosse, F.; Weisshart, K. Composition and hierarchical organisation of a spider silk. PLoS ONE 2007, 2, e998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riekel, C.; Burghammer, M.; Dane, T.G.; Ferrero, C.; Rosenthal, M. Nanoscale structural features in major ampullate spider silk. Biomacromolecules 2017, 18, 231–241. [Google Scholar] [CrossRef]
- Holland, G.P.; Lewis, R.V.; Yarger, J.L. WISE NMR characterization of nanoscale heterogeneity and mobility in supercontracted Nephila clavipes spider dragline silk. J. Am. Chem. Soc. 2004, 126, 5867–5872. [Google Scholar] [CrossRef] [PubMed]
- Sapede, D.; Seydel, T.; Forsyth, V.T.; Koza, M.M.; Schweins, R.; Vollrath, F.; Riekel, C. Nanofibrillar structure and molecular mobility in spider dragline silk. Macromolecules 2005, 38, 8447–8453. [Google Scholar] [CrossRef]
- Keten, S.; Buehler, M.J. Nanostructure and molecular mechanics of spider dragline silk protein assemblies. J. R. Soc. Interface 2010, 7, 1709–1721. [Google Scholar] [CrossRef] [Green Version]
- Heim, M.; Römer, L.; Scheibel, T. Hierarchical structures made of proteins. The complex architecture of spider webs and their constituent silk proteins. Chem. Soc. Rev. 2010, 39, 156–164. [Google Scholar] [CrossRef]
- Yarger, J.L.; Cherry, B.R.; van der Vaart, A. Uncovering the structure–function relationship in spider silk. Nat. Rev. Mater. 2018, 3, 1–11. [Google Scholar] [CrossRef]
- Blamires, S.J.; Blackledge, T.A.; Tso, I.M. Physicochemical property variation in spider silk: Ecology, evolution, and synthetic production. Annu. Rev. Entomol. 2017, 62, 443–460. [Google Scholar] [CrossRef] [PubMed]
- Hutter, J.L.; Bechhoefer, J. Calibration of atomic-force microscope tips. Rev. Sci. Instrum. 1993, 64, 1868–1873. [Google Scholar] [CrossRef] [Green Version]
- Sneddon, I.N. Fourier Transforms; McGraw-Hill: New York, NY, USA, 1951; ISBN 0486685225. [Google Scholar]
- Young, T.J.; Monclus, M.A.; Burnett, T.L.; Broughton, W.R.; Ogin, S.L.; Smith, P.A. The use of the PeakForceTM quantitative nanomechanical mapping AFM-based method for high-resolution Young’s modulus measurement of polymers. Meas. Sci. Technol. 2011, 22, 125703. [Google Scholar] [CrossRef] [Green Version]
- Pittenger, B.; Erina, N.; Su, C. Quantitative Mechanical Property Mapping at the Nanoscale with PeakForce QNM. Application Note #128 Veeco Instruments Inc. 2010. Available online: http://www.bruker.com/fileadmin/user_upload/8-PDF-Docs/SurfaceAnalysis/AFM/ApplicationNotes/AN128-RevB0-Quantitative_Mechanical_Property_Mapping_at_the_Nanoscale_with_PeakForceQNM-AppNote.pdf (accessed on 9 August 2018).
- Rasband, W.S. ImageJ, Version 1.51s; U. S. National Institutes of Health: Bethesda, MD, USA, 2018. Available online: https://imagej.nih.gov/ij/ (accessed on 5 January 2021).






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piorkowski, D.; He, B.-C.; Blamires, S.J.; Tso, I.-M.; Kane, D.M. Nanoscale Material Heterogeneity of Glowworm Capture Threads Revealed by AFM. Molecules 2021, 26, 3500. https://doi.org/10.3390/molecules26123500
Piorkowski D, He B-C, Blamires SJ, Tso I-M, Kane DM. Nanoscale Material Heterogeneity of Glowworm Capture Threads Revealed by AFM. Molecules. 2021; 26(12):3500. https://doi.org/10.3390/molecules26123500
Chicago/Turabian StylePiorkowski, Dakota, Bo-Ching He, Sean J. Blamires, I-Min Tso, and Deborah M. Kane. 2021. "Nanoscale Material Heterogeneity of Glowworm Capture Threads Revealed by AFM" Molecules 26, no. 12: 3500. https://doi.org/10.3390/molecules26123500
APA StylePiorkowski, D., He, B. -C., Blamires, S. J., Tso, I. -M., & Kane, D. M. (2021). Nanoscale Material Heterogeneity of Glowworm Capture Threads Revealed by AFM. Molecules, 26(12), 3500. https://doi.org/10.3390/molecules26123500