
Immunological Analysis of Isothiocyanate-Modified α-Lactalbumin Using High-Performance Thin Layer Chromatography



Abstract
:1. Introduction
2. Results
2.1. HPTLC-Immunostaining (HPTLC-IS) of Intact Proteins and Its Validation
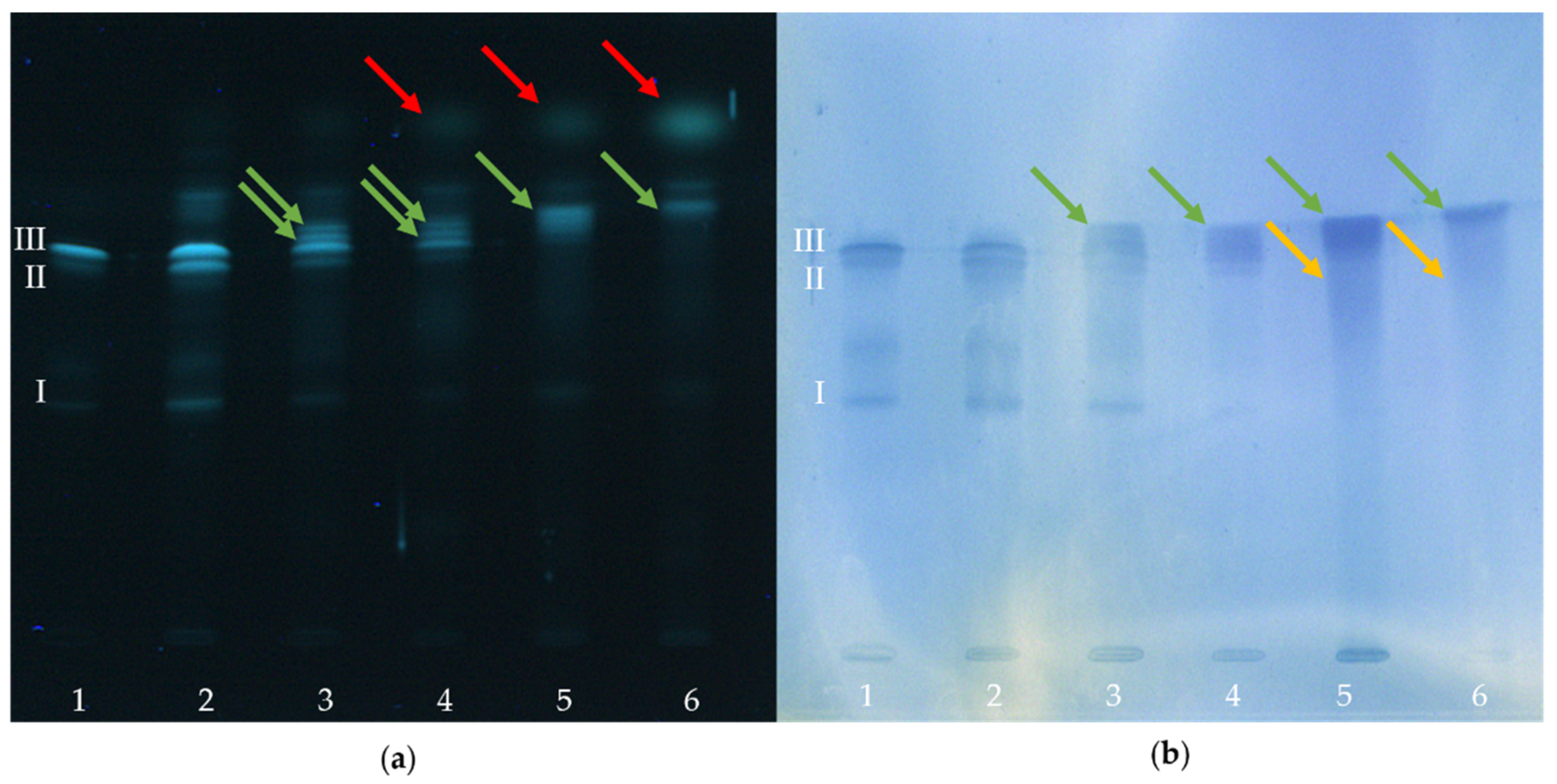
2.2. HPTLC-Immunostaining of BITC-Modified and Non-Modified α-Lactalbumin
2.3. HPTLC-Immunostaining of Tryptic Peptides
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Methods
4.2.1. Preparation of ITC Protein Conjugates
4.2.2. Tryptic Digestion of α-Lactalbumin
4.3. High-Performance Thin-Layer Chromatography-Immunostaining (HPTLC-IS)
4.3.1. Protein Derivatives
4.3.2. Tryptic Peptides
4.3.3. Protein-/Peptide-Specific Staining
4.3.4. High-Performance Thin-Layer Chromatography-Immunostaining (HPTLC-IS)
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Keppler, J.K.; Schwarz, K.; van der Goot, A.J. Covalent modification of food proteins by plant-based ingredients (polyphenols and organosulphur compounds): A commonplace reaction with novel utilization potential. Trends Food Sci. Technol. 2020, 101, 38–49. [Google Scholar] [CrossRef]
- Kühn, C.; von Oesen, T.; Hanschen, F.S.; Rohn, S. Determination of isothiocyanate-protein conjugates in milk and curd after adding garden cress (Lepidium sativum L.). Food Res. Int. 2018, 108, 621–627. [Google Scholar] [CrossRef]
- Rade-Kukic, K.; Schmitt, C.; Rawel, H.M. Formation of conjugates between β-lactoglobulin and allyl isothiocyanate: Effect on protein heat aggregation, foaming and emulsifying properties. Food Hydrocoll. 2011, 25, 694–706. [Google Scholar] [CrossRef]
- Keppler, J.K.; Koudelka, T.; Palani, K.; Tholey, A.; Schwarz, K. Interaction of β-Lactoglobulin with Small Hydrophobic Ligands—Influence of Covalent AITC Modification on β-LG Tryptic Cleavage. Food Biophys. 2014, 9, 349–358. [Google Scholar] [CrossRef]
- Keppler, J.K.; Martin, D.; Garamus, V.M.; Schwarz, K. Differences in binding behavior of (−)-epigallocatechin gallate to β-lactoglobulin heterodimers (AB) compared to homodimers (A) and (B). J. Mol. Recognit. 2015, 28, 656–666. [Google Scholar] [CrossRef]
- Keppler, J.K.; Martin, D.; Garamus, V.M.; Berton-Carabin, C.; Nipoti, E.; Coenye, T.; Schwarz, K. Functionality of whey proteins covalently modified by allyl isothiocyanate. Part 1 physicochemical and antibacterial properties of native and modified whey proteins at pH 2 to 7. Food Hydrocoll. 2017, 65, 130–143. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Lu, Y.; Xu, H.; Lin, D.; He, Z.; Wu, H.; Liu, L.; Wang, Z. Reducing the allergenic capacity of β-lactoglobulin by covalent conjugation with dietary polyphenols. Food Chem. 2018, 256, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Rawel, H.M.; Kroll, J.; Schröder, I. Reactions of isothiocyanates with food proteins: Influence on enzyme activity and tryptical degradation. Food/Nahrung 1998, 42, 197–199. [Google Scholar] [CrossRef]
- Kroll, J.; Noack, J.; Rawel, H.; Kroeck, R.; Proll, J. Chemical reactions of benzyl isothiocyanate with egg-white protein fractions. J. Sci. Food Agric. 1994, 65, 337–345. [Google Scholar] [CrossRef]
- Kroll, J.; Rawel, H.; Kröck, R.; Proll, J.; Schnaak, W. Interactions of isothiocyanates with egg white proteins. Food/Nahrung 1994, 38, 53–60. [Google Scholar] [CrossRef]
- Keppler, J.K.; Schwarz, K. Increasing the emulsifying capacity of whey proteins at acidic pH values through covalent modification with allyl isothiocyanate. Colloids Surf. A Physicochem. Eng. Asp. 2017, 522, 514–524. [Google Scholar] [CrossRef]
- Kühn, C.; Von Oesen, T.; Herz, C.; Schreiner, M.; Hanschen, F.S.; Lamy, E.; Rohn, S.; Kuehn, C. In Vitro Determination of Protein Conjugates in Human Cells by LC-ESI-MS/MS after Benzyl Isothiocyanate Exposure. J. Agric. Food Chem. 2018, 66, 6727–6733. [Google Scholar] [CrossRef] [PubMed]
- Beevi, S.S.; Mangamoori, L.N.; Dhand, V.; Ramakrishna, D.S. Isothiocyanate Profile and Selective Antibacterial Activity of Root, Stem, and Leaf Extracts Derived from Raphanus sativus L. Foodborne Pathog. Dis. 2009, 6, 129–136. [Google Scholar] [CrossRef]
- Lee, Y.M.; Seon, M.R.; Cho, H.J.; Kim, J.-S.; Park, J.H.Y. Benzyl isothiocyanate exhibits anti-inflammatory effects in murine macrophages and in mouse skin. J. Mol. Med. 2009, 87, 1251–1261. [Google Scholar] [CrossRef] [PubMed]
- Sofrata, A.; Santangelo, E.M.; Azeem, M.; Borg-Karlson, A.-K.; Gustafsson, A.; Pütsep, K. Benzyl Isothiocyanate, a Major Component from the Roots of Salvadora Persica Is Highly Active against Gram-Negative Bacteria. PLoS ONE 2011, 6, e23045. [Google Scholar] [CrossRef]
- Guzmán-Pérez, V.; Bumke-Vogt, C.; Schreiner, M.; Mewis, I.; Borchert, A.; Pfeiffer, A.F.H. Benzylglucosinolate Derived Isothiocyanate from Tropaeolum majus Reduces Gluconeogenic Gene and Protein Expression in Human Cells. PLoS ONE 2016, 11, e0162397. [Google Scholar] [CrossRef]
- Cejpek, K.; Valušek, J.; Velíšek, J. Reactions of Allyl Isothiocyanate with Alanine, Glycine, and Several Peptides in Model Systems. J. Agric. Food Chem. 2000, 48, 3560–3565. [Google Scholar] [CrossRef]
- Hanschen, F.S.; Brüggemann, N.; Brodehl, A.; Mewis, I.; Schreiner, M.; Rohn, S.; Kroh, L.W. Characterization of Products from the Reaction of Glucosinolate-Derived Isothiocyanates with Cysteine and Lysine Derivatives Formed in Either Model Systems or Broccoli Sprouts. J. Agric. Food Chem. 2012, 60, 7735–7745. [Google Scholar] [CrossRef]
- Zhang, Y.; Talalay, P. Anticarcinogenic activities of organic isothiocyanates: Chemistry and mechanisms. Cancer Res. 1994, 54, 1976–1981. [Google Scholar]
- Nciuc, N.S.T.Ă.; Râpeanu, G. An overview of bovine α -lactalbumin structure and functionality. Ann. Univ. Dunarea Jos Galati Fascicle VI Food Technol. 2010, 34, 82–93. [Google Scholar]
- Smithers, G.W. Whey and whey proteins—From ‘gutter-to-gold’. Int. Dairy J. 2008, 18, 695–704. [Google Scholar] [CrossRef]
- Smithers, G.W.; Ballard, F.J.; Copeland, A.D.; De Silva, K.J.; Dionysius, D.A.; Francis, G.L.; Goddard, C.; Grieve, P.A.; McIntosh, G.H.; Mitchell, I.R.; et al. New Opportunities from the Isolation and Utilization of Whey Proteins. J. Dairy Sci. 1996, 79, 1454–1459. [Google Scholar] [CrossRef]
- Kinsella, J.E.; Morr, C.V. Milk proteins: Physicochemical and functional properties. C R C Crit. Rev. Food Sci. Nutr. 1984, 21, 197–262. [Google Scholar] [CrossRef]
- Permyakov, E.A.; Berliner, L.J. α-Lactalbumin: Structure and function. FEBS Lett. 2000, 473, 269–274. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, R.J.; Murray, B.A.; Walsh, D.J. Hypotensive Peptides from Milk Proteins. J. Nutr. 2004, 134, 980S–988S. [Google Scholar] [CrossRef] [Green Version]
- Markus, C.R.; Olivier, B.; Panhuysen, G.E.M.; Van Der Gugten, J.; Alles, M.S.; Tuiten, A.; Westenberg, H.G.M.; Fekkes, D.; Koppeschaar, H.F.; De Haan, E.E.H.F. The bovine protein α-lactalbumin increases the plasma ratio of tryptophan to the other large neutral amino acids, and in vulnerable subjects raises brain serotonin activity, reduces cortisol concentration, and improves mood under stress. Am. J. Clin. Nutr. 2000, 71, 1536–1544. [Google Scholar] [CrossRef] [Green Version]
- Sternhagen, L.G.; Allen, J.C. Growth rates of a human colon adenocarinoma cell line are regulated by the milk proteins alpha-lactalbumin. Adv. Exp. Med. Biol. 2001, 501, 115–120. [Google Scholar]
- Pellegrini, A.; Thomas, U.; Bramaz, N.; Hunziker, P.; Von Fellenberg, R. Isolation and identification of three bactericidal domains in the bovine α--lactalbumin molecule. Biochim. Biophys. Acta Gen. Subj. 1999, 1426, 439–448. [Google Scholar] [CrossRef]
- Cross, M.L.; Gill, H.S. Immunomodulatory Properties of Milk. Br. J. Nutr. 2000, 84, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Villa, C.; Costa, J.; Oliveira, M.B.P.P.; Mafra, I. Bovine Milk Allergens: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2017, 17, 137–164. [Google Scholar] [CrossRef] [Green Version]
- Paschke, A. Proteine als Lebensmittelallergene. Nachr. Chem. 2008, 56, 1005–1009. [Google Scholar] [CrossRef]
- Ilinskaya, A.N.; Dobrovolskaia, M.A. Understanding the immunogenicity and antigenicity of nanomaterials: Past, present and future. Toxicol. Appl. Pharmacol. 2016, 299, 70–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Yuan, S.; Huang, M.; Gao, J.; Wu, Z.; Tong, P.; Yang, A.; Chen, H. Identification of IgE and IgG epitopes on native Bos d 4 allergen specific to allergic children. Food Funct. 2016, 7, 2996–3005. [Google Scholar] [CrossRef] [PubMed]
- Gershoni, J.M.; Roitburd-Berman, A.; Siman-Tov, D.D.; Freund, N.T.; Weiss, Y. Epitope Mapping. BioDrugs 2007, 21, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Willison, L.A.N.; Zhang, Q.; Su, M.; Teuber, S.S.; Sathe, S.K.; Roux, K.H. Conformational epitope mapping of Pru du 6, a major allergen from almond nut. Mol. Immunol. 2013, 55, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Pomés, A. Relevant B Cell Epitopes in Allergic Disease. Int. Arch. Allergy Immunol. 2010, 152, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Sampson, H.A. Update on food allergy. J. Allergy Clin. Immunol. 2004, 113, 805–819. [Google Scholar] [CrossRef]
- Matissek, R.; Steiner, G.; Fischer, M. Lebensmittelanalytik; Bd. fünfte, vo; Springer: Berlin/Heidelberg, Germany, 2014; ISBN 978-3-642-34828-0. [Google Scholar]
- Davis, P.J.; Smales, C.M.; James, D.C. How can thermal processing modify the antigenicity of proteins? Allergy 2001, 56, 56–60. [Google Scholar] [CrossRef]
- Li, Z.; Luo, Y.; Feng, L. Effects of Maillard reaction conditions on the antigenicity of α-lactalbumin and β-lactoglobulin in whey protein conjugated with maltose. Eur. Food Res. Technol. 2011, 233, 387–394. [Google Scholar] [CrossRef]
- Jiménez-Saiz, R.; Belloque, J.; Molina, E.; López-Fandiño, R. Human Immunoglobulin E (IgE) Binding to Heated and Glycated Ovalbumin and Ovomucoid before and after in Vitro Digestion. J. Agric. Food Chem. 2011, 59, 10044–10051. [Google Scholar] [CrossRef]
- Bu, G.; Luo, Y.; Chen, F.; Liu, K.; Zhu, T. Milk processing as a tool to reduce cow’s milk allergenicity: A mini-review. Dairy Sci. Technol. 2013, 93, 211–223. [Google Scholar] [CrossRef] [Green Version]
- Verhoeckx, K.C.M.; Vissers, Y.M.; Baumert, J.L.; Faludi, R.; Feys, M.; Flanagan, S.; Herouet-Guicheney, C.; Holzhauser, T.; Shimojo, R.; van der Bolt, N.; et al. Food processing and allergenicity. Food Chem. Toxicol. 2015, 80, 223–240. [Google Scholar] [CrossRef]
- Teodorowicz, M.; Van Neerven, J.; Savelkoul, H. Food Processing: The Influence of the Maillard Reaction on Immunogenicity and Allergenicity of Food Proteins. Nutrients 2017, 9, 835. [Google Scholar] [CrossRef]
- Mills, E.N.C.; Sancho, A.I.; Rigby, N.M.; Jenkins, J.A.; Mackie, A.R. Impact of food processing on the structural and allergenic properties of food allergens. Mol. Nutr. Food Res. 2009, 53, 963–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paschke, A.; Besler, M. Stability of bovine allergens during food processing. Ann. Allergy Asthma Immunol. 2002, 89, 16–20. [Google Scholar] [CrossRef]
- Fiocchi, A.; Bouygue, G.R.; Sarratud, T.; Terracciano, L.; Martelli, A.; Restani, P. Clinical tolerance of processed foods. Ann. Allergy Asthma Immunol. 2004, 93, S38–S46. [Google Scholar] [CrossRef]
- Biller, J.; Morschheuser, L.; Riedner, M.; Rohn, S. Development of optimized mobile phases for protein separation by high performance thin layer chromatography. J. Chromatogr. A 2015, 1415, 146–154. [Google Scholar] [CrossRef]
- Morlock, G.; Schwack, W. Hyphenations in planar chromatography. J. Chromatogr. A 2010, 1217, 6600–6609. [Google Scholar] [CrossRef]
- Morlock, G.; Schwack, W. Coupling of planar chromatography to mass spectrometry. TrAC Trends Anal. Chem. 2010, 29, 1157–1171. [Google Scholar] [CrossRef]
- Morlock, G.E.; Klingelhöfer, I. Liquid Chromatography-Bioassay-Mass Spectrometry for Profiling of Physiologically Active Food. Anal. Chem. 2014, 86, 8289–8295. [Google Scholar] [CrossRef] [PubMed]
- Morschheuser, L.; Mink, K.; Horst, R.; Kallinich, C.; Rohn, S. Immunological analysis of food proteins using high-performance thin-layer chromatography-immunostaining. J. Chromatogr. A 2017, 1526, 157–166. [Google Scholar] [CrossRef]
- Meisen, I.; Mormann, M.; Müthing, J. Thin-layer chromatography, overlay technique and mass spectrometry: A versatile triad advancing glycosphingolipidomics. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2011, 1811, 875–896. [Google Scholar] [CrossRef]
- Morschheuser, L.; Mükusch, S.; Riedner, M.; Seitz, H.; Rohn, S. High-performance thin-layer chromatography as a fast screening tool for phosphorylated peptides. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2016, 1008, 198–205. [Google Scholar] [CrossRef]
- Widmer, V.; Handloser, D.; Reich, E. Quantitative HPTLC Analysis of Artemisinin in Dried Artemisia annua L.: A Practical Approach. J. Liq. Chromatogr. Relat. Technol. 2007, 30, 2209–2219. [Google Scholar] [CrossRef]
- Reim, V.; Rohn, S. Characterization of saponins in peas (Pisum sativum L.) by HPTLC coupled to mass spectrometry and a hemolysis assay. Food Res. Int. 2015, 76, 3–10. [Google Scholar] [CrossRef]
- Morschheuser, L.; Wessels, H.; Pille, C.; Fischer, J.; Hünniger, T.; Fischer, M.; Paschke-Kratzin, A.; Rohn, S. HPTLC-aptastaining—Innovative protein detection system for high-performance thin-layer chromatography. Sci. Rep. 2016, 6, 26665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsen, J.V.; Ong, S.-E.; Mann, M. Trypsin Cleaves Exclusively C-terminal to Arginine and Lysine Residues. Mol. Cell. Proteom. 2004, 3, 608–614. [Google Scholar] [CrossRef] [Green Version]
- Vanaman, T.C.; Brew, K.; Hill, R.L. The Disulfide Bonds of Bovine α-Lactalbumin. J. Biol. Chem. 1970, 245, 4583–4590. [Google Scholar] [CrossRef]
- Järvinen, K.M.; Chatchatee, P.; Bardina, L.; Beyer, K.; Sampson, H.A. IgE and IgG binding epitopes on β-lactalbumin and β-lactoglobulin in cow’s milk allergy. Int. Arch. Allergy Immunol. 2001, 126, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Wal, J.-M. Bovine milk allergenicity. Ann. Allergy Asthma Immunol. 2004, 93, S2–S11. [Google Scholar] [CrossRef]
- Monaci, L.; Tregoat, V.; Van Hengel, A.J.; Anklam, E. Milk allergens, their characteristics and their detection in food: A review. Eur. Food Res. Technol. 2006, 223, 149–179. [Google Scholar] [CrossRef]
- Maynard, F.; Jost, R.; Wal, J.-M. Human IgE Binding Capacity of Tryptic Peptides from Bovine α-Lactalbumin. Int. Arch. Allergy Immunol. 1997, 113, 478–488. [Google Scholar] [CrossRef]
- Farrell, H.M.; Jimenez-Flores, R.; Bleck, G.T.; Brown, E.M.; Butler, J.E.; Creamer, L.K.; Hicks, C.L.; Hollar, C.M.; Ng-Kwai-Hang, K.F.; Swaisgood, H.E. Nomenclature of the Proteins of Cows’ Milk—Sixth Revision. J. Dairy Sci. 2004, 87, 1641–1674. [Google Scholar] [CrossRef] [Green Version]
- Keppler, J.K.; Koudelka, T.; Palani, K.; Stuhldreier, M.C.; Temps, F.; Tholey, A.; Schwarz, K. Characterization of the covalent binding of allyl isothiocyanate toβ-lactoglobulin by fluorescence quenching, equilibrium measurement, and mass spectrometry. J. Biomol. Struct. Dyn. 2014, 32, 1103–1117. [Google Scholar] [CrossRef]
- Kawakishi, S.; Kaneko, T. Interaction of proteins with allyl isothiocyanate. J. Agric. Food Chem. 1987, 35, 85–88. [Google Scholar] [CrossRef]
- Maynard, F.; Chatel, J.-M.; Wal, J.-M. Immunological IgE Cross-Reactions of Bovine and Humanα-Lactalbumins in Cow’s Milk Allergic Patients. Food Agric. Immunol. 1999, 11, 179–189. [Google Scholar] [CrossRef]
- Bu, G.; Luo, Y.; Zheng, Z.; Zheng, H. Effect of heat treatment on the antigenicity of bovine α-lactalbumin and β-lactoglobulin in whey protein isolate. Food Agric. Immunol. 2009, 20, 195–206. [Google Scholar] [CrossRef]
- Davey, B. Antigene. In Immunologie; Birkhäuser: Basel, Switzerland, 1991; pp. 106–124. [Google Scholar]
- Sélo, I.; Clément, G.; Bernard, H.; Chatel, J.M.; Créminon, C.; Peltre, G.; Wal, J.M. Allergy to bovine β-lactoglobulin: Specificity of human IgE to tryptic peptides. Clin. Exp. Allergy 1999, 29, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Hanschen, F.S.; Lamy, E.; Schreiner, M.; Rohn, S. Reactivity and Stability of Glucosinolates and Their Breakdown Products in Foods. Angew. Chem. Int. Ed. 2014, 53, 11430–11450. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, D.A.; Nesbit, J.B.; Hurlburt, B.K.; Cheng, H.; Maleki, S.J. Processing Can Alter the Properties of Peanut Extract Preparations. J. Agric. Food Chem. 2010, 58, 1138–1143. [Google Scholar] [CrossRef]
- Maleki, S.J.; Hurlburt, B.K. Structural and functional alterations in major peanut allergens caused by thermal processing. J. Assoc. Off. Anal. Chem. 2005, 87, 1475–1479. [Google Scholar]
- Jiménez-Saiz, R.; Benedé, S.; Molina, E.; López-Expósito, I. Effect of Processing Technologies on the Allergenicity of Food Products. Crit. Rev. Food Sci. Nutr. 2015, 55, 1902–1917. [Google Scholar] [CrossRef] [Green Version]
- Paschke, A. Aspects of food processing and its effect on allergen structure. Mol. Nutr. Food Res. 2009, 53, 959–962. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, A.S. Effects of protein aggregates: An immunologic perspective. AAPS J. 2006, 8, E501–E507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doke, S.; Nakamura, R.; Torii, S. Allergenicity of food proteins interacted with oxidized lipids in soybean-sensitive individuals. Agric. Biol. Chem. 1989, 53, 1231–1235. [Google Scholar] [CrossRef] [Green Version]
- Stanic-Vucinic, D.; Cirkovic-Velickovic, T. The modifications of bovine β-lactoglobulin: Effects on its structural and functional properties. J. Serb. Chem. Soc. 2013, 78, 445–461. [Google Scholar] [CrossRef]
- Bleumink, E.; Berrens, L. Synthetic approaches to the biological activity of beta-lactoglobulin in human allergy to cows’ milk. Nature 1966, 212, 541–543. [Google Scholar] [CrossRef]
- Lu, Y.; Li, S.; Xu, H.; Zhang, T.; Lin, X.; Wu, X. Effect of Covalent Interaction with Chlorogenic Acid on the Allergenic Capacity of Ovalbumin. J. Agric. Food Chem. 2018, 66, 9794–9800. [Google Scholar] [CrossRef]
- Xu, H.; Zhang, T.; Lu, Y.; Lin, X.; Hu, X.; Liu, L.; He, Z.; Wu, X. Effect of chlorogenic acid covalent conjugation on the allergenicity, digestibility and functional properties of whey protein. Food Chem. 2019, 298, 125024. [Google Scholar] [CrossRef]
- Wijesinha-Bettoni, R.; Gao, C.; Jenkins, J.A.; Mackie, A.R.; Wilde, P.J.; Mills, E.N.C.; Smith, L.J. Heat Treatment of Bovine α-Lactalbumin Results in Partially Folded, Disulfide Bond Shuffled States with Enhanced Surface Activity. Biochemistry 2007, 46, 9774–9784. [Google Scholar] [CrossRef]
- FDA. Bioanalytical Method Validation Guidance for Industry; US Department of Health and Human Services Food and Drug Administration Center for Drug Evaluation and Research and Center for Veterinary Medicine: Rockville, MD, USA, 2018; pp. 1–41.
- Kromidas, S.; Klinkner, R.; Mertens, R. Methodenvalidierung im analytischen Labor. Nachr. Chem. Tech. Lab. 1995, 43, 669–676. [Google Scholar] [CrossRef]
- Wadhwa, M.; Knezevic, I.; Kang, H.-N.; Thorpe, R. Immunogenicity assessment of biotherapeutic products: An overview of assays and their utility. Biologicals 2015, 43, 298–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowatzke, W.L.; Oliver, K.G.; Cho, C.Y.; Rallabhandi, P.; Garber, E.A.E. Single-Laboratory Validation of the Multiplex xMAP Food Allergen Detection Assay with Incurred Food Samples. J. Agric. Food Chem. 2019, 67, 484–498. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Synonym | Value | RSD |
|---|---|---|
| Limit of detection | 0.040 µg | 0.74% |
| Limit of quantification | 0.177 µg | 3.27% |
| Accuracy | 99.93% | 4.27% |
| Precision | 8.55% | - |
| Coefficient of determination | 0.956 | 1.52% |
| Mass | Position | Peptide Sequence |
|---|---|---|
| 618.35 | 20–24 | EQLTK |
| 653.31 | 25–29 | CEVFR |
| 389.24 | 30–32 | ELK |
| 375.22 | 33–35 | DLK |
| 4654.15 | 36–77 | GYGGVSLPEWVCTTFHTSGYDTQAIVQNNDSTEYGLFQINNK |
| 549.29 | 78–81 | IWCK |
| 1889.78 | 82–98 | DDQNPHSSNICNISCDK |
| 1642.73 | 99–112 | FLDDDLTDDIMCVK |
| 147.11 | 113–113 | K |
| 488.31 | 114–117 | ILDK |
| 1220.65 | 118–127 | VGINYWLAHK |
| 650.32 | 128–133 | ALCSEK |
| 1034.50 | 134–141 | LDQWLCEK |
| 132.10 | 142–142 | L |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spöttel, J.; Brockelt, J.; Badekow, S.; Rohn, S. Immunological Analysis of Isothiocyanate-Modified α-Lactalbumin Using High-Performance Thin Layer Chromatography. Molecules 2021, 26, 1842. https://doi.org/10.3390/molecules26071842
Spöttel J, Brockelt J, Badekow S, Rohn S. Immunological Analysis of Isothiocyanate-Modified α-Lactalbumin Using High-Performance Thin Layer Chromatography. Molecules. 2021; 26(7):1842. https://doi.org/10.3390/molecules26071842
Chicago/Turabian StyleSpöttel, Jenny, Johannes Brockelt, Svenja Badekow, and Sascha Rohn. 2021. "Immunological Analysis of Isothiocyanate-Modified α-Lactalbumin Using High-Performance Thin Layer Chromatography" Molecules 26, no. 7: 1842. https://doi.org/10.3390/molecules26071842
APA StyleSpöttel, J., Brockelt, J., Badekow, S., & Rohn, S. (2021). Immunological Analysis of Isothiocyanate-Modified α-Lactalbumin Using High-Performance Thin Layer Chromatography. Molecules, 26(7), 1842. https://doi.org/10.3390/molecules26071842