
Physiological and Transcriptomic Analysis Provide Insight into Low Temperature Enhancing Hypericin Biosynthesis in Hypericum perforatum


 and
and 
Abstract
:1. Introduction
2. Results
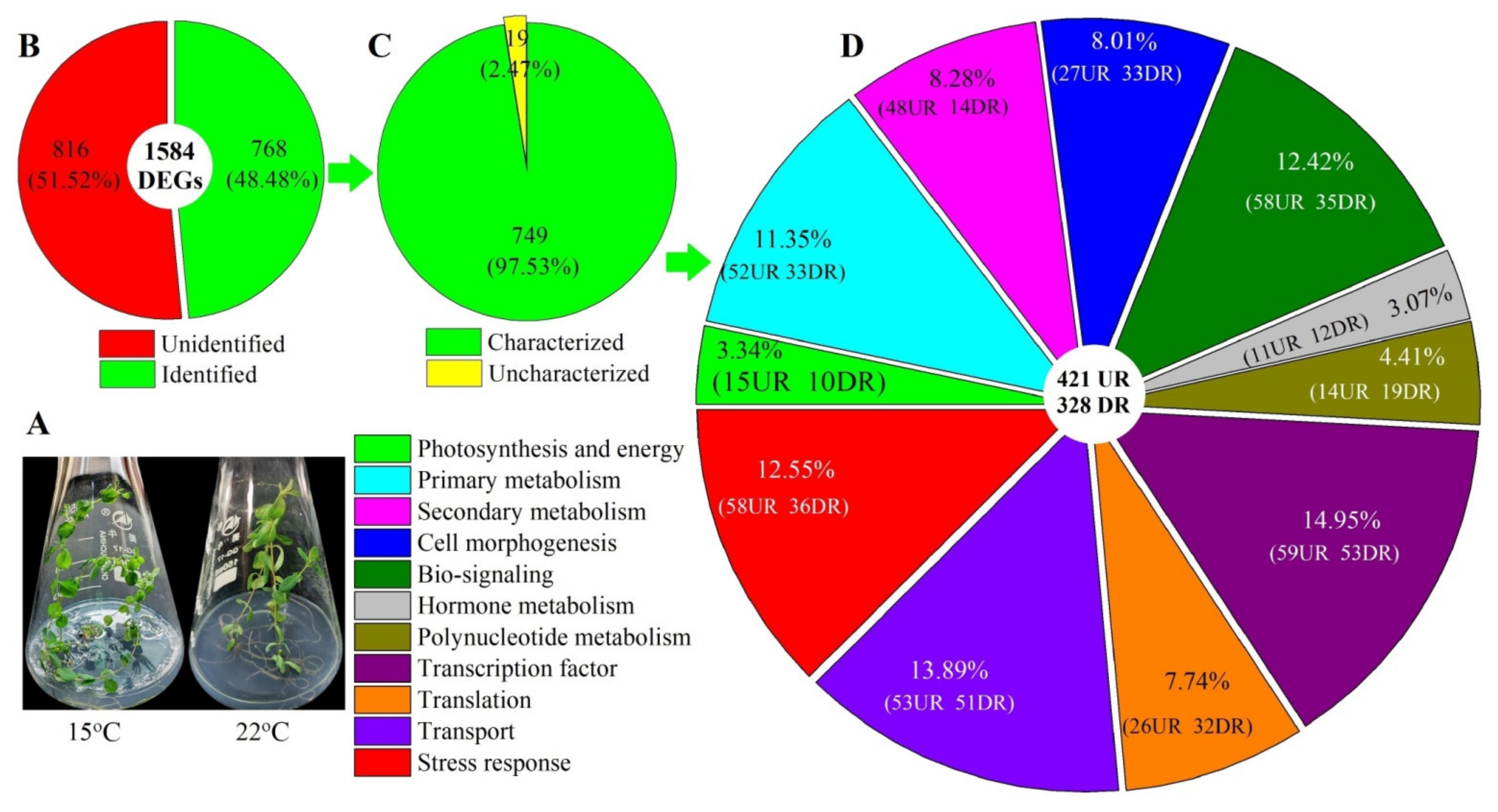
2.1. Effect of Low Temperature on Biomass, Dark Glands, Leaf Stomata and Hyp Accumulation
2.2. Global Gene Analysis
2.3. Biological Category of DEGs
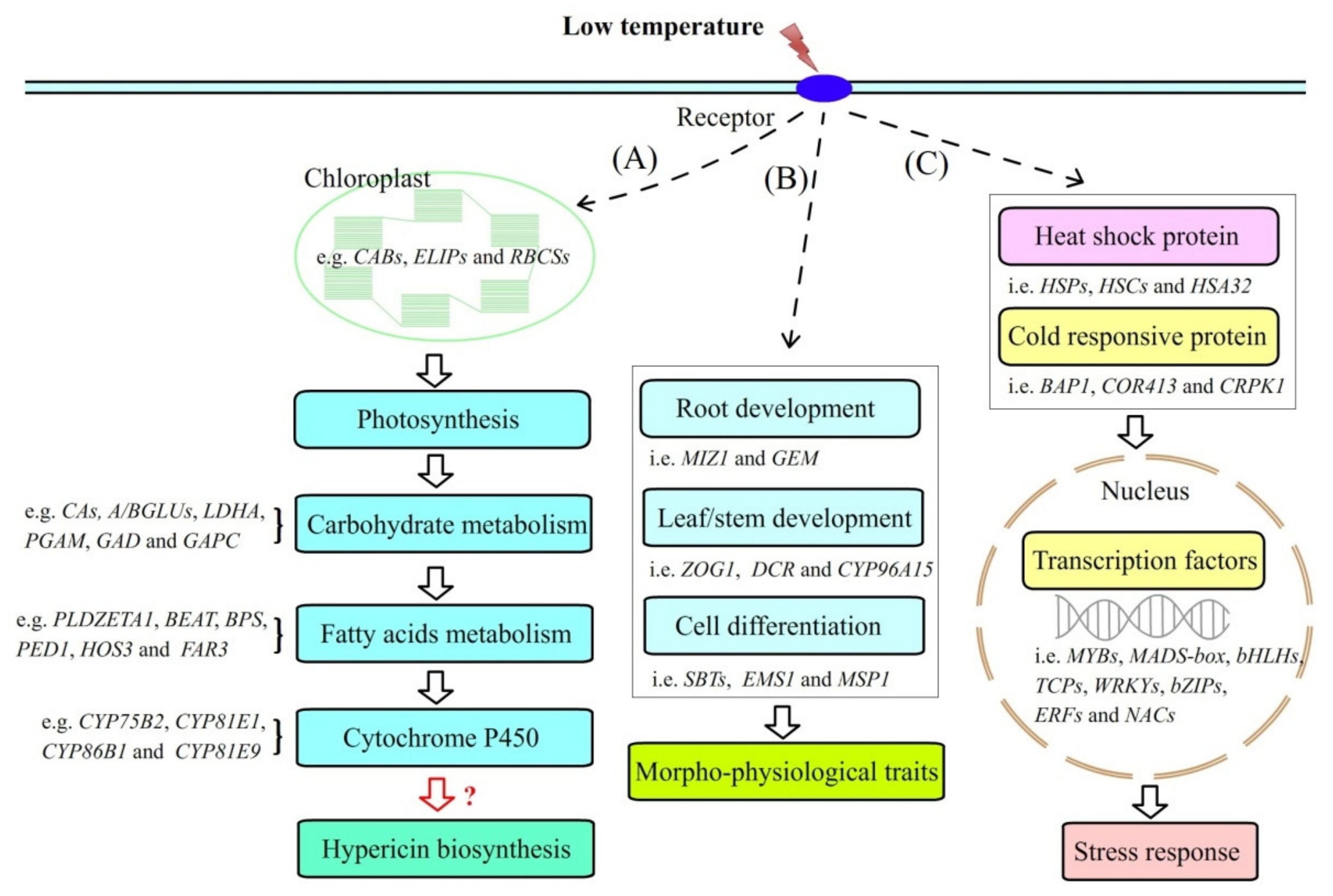
2.4. DEGs Involved in Hyp Biosynthesis
2.4.1. DEGs Involved in Photosynthesis
2.4.2. DEGs Involved in Carbohydrate Metabolism
2.4.3. DEGs Involved in Fatty Acids Metabolism
2.4.4. Differentially Expressed Cytochrome P450 (CYPs)
2.5. DEGs Involved in Morpho-Physiological Traits
2.6. DEGs Involved in Stress Response
2.6.1. DEGs Involved in Heat Shock Proteins (HSPs) and Cold-Responsive Proteins (CRPs)
2.6.2. Differentially Expressed Transcription Factors (TFs)
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Observation of Dark Gland and Leaf Stomata
4.3. Hyp HPLC Quantification
4.4. Total RNA Extraction and Illumina Sequencing
4.5. Sequence Filtration, Assembly, Unigene Expression Analysis and Basic Annotation
4.6. qRT-PCR Validation
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Bombardelli, E.; Morazzoni, P. Hypericum perforatum. Fitoterapia 1995, 66, 43–68. [Google Scholar]
- Carlo, G.D.; Borrelli, F.; Ernest, E.; Izzo, A.A. St. John’s Wort: Prozac from the plant kingdom. Trends. Pharmacol. Sci. 2001, 22, 292–297. [Google Scholar] [CrossRef]
- Vattikuti, U.M.R.; Ciddi, V. An overview on Hypericum perforatum. Linn. Nat. Prod. Rad. 2005, 4, 368–381. [Google Scholar]
- Miller, A.L. St. John’s Wort (Hypericum perforatum): Clinical effects on depression and other conditions. Altern. Med. Rev. A J. Clin. Ther. 1998, 3, 18–26. [Google Scholar]
- Laudahn, D. High dosed st.-john’s-wort-extract for treatment of mild and moderate depressions. Acta Hortic. 1993, 332, 265–272. [Google Scholar] [CrossRef]
- Velingkar, V.S.; Gupta, G.L.; Hegde, N.B. A current update on phytochemistry, pharmacology and herb–drug interactions of Hypericum perforatum. Phytochem. Rev. 2017, 16, 725–744. [Google Scholar] [CrossRef]
- Barnes, J.; Arnason, J.T.; Roufogalis, B.D. St John’s wort (Hypericum perforatum L.): Botanical, chemical, pharmacological and clinical advances. J. Pharm. Pharmacol. 2019, 71, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Sun, P.; Kang, T.; Xing, H.; Zhang, Z.; Yang, D.; Zhang, J.; Paré, P.W.; Li, M. Phytochemical Changes in Aerial Parts of Hypericum perforatum at Different Harvest Stages. Rec. Nat. Prod. 2019, 13, 1–9. [Google Scholar] [CrossRef]
- Kusari, S.; Zuhlke, S.; Kosuth, J.; Cellarova, E.; Spiteller, M. Light-independent metabolomics of endophytic ThielaWia sub-thermophila provides insight into microbial hypericin biosynthesis. J. Nat. Prod. 2009, 72, 1825–1835. [Google Scholar] [CrossRef]
- Zobayed, S.M.A.; Afreen, F.; Kozai, E.G. Plant–environment interactions: Accumulation of hypericin in dark glands of Hypericum perforatum. Ann. Bot. 2006, 98, 793–804. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, P.; Altschmied, L.; Stark, P.; Rutten, T.; Guendel, A.; Scharfenberg, S.; Franke, K.; Baeumlein, H.; Wessjohann, L.; Koch, M.; et al. Discovery of key regulators of dark gland development and hypericin biosynthesis in St. John’s Wort (Hypericum perforatum). Plant Biotechnol. J. 2019, 17, 2299–2312. [Google Scholar] [CrossRef] [Green Version]
- Couceiro, M.; Afreen, F.; Zobayed, S.; Kozai, T. Variation in concentrations of major bioactive compounds of St. John’s wort: Effects of harvesting time, temperature and germplasm. Plant Sci. 2006, 170, 128–134. [Google Scholar] [CrossRef]
- Morshedloo, M.R.; Ebadi, A.; Maggi, F.; Fattahi, R.; Yazdani, M.J.D.; Jafari, M. Chemical characterization of the essential oil compositions from Iranian populations of Hypericum perforatum L. Ind. Crop. Prod. 2015, 76, 565–573. [Google Scholar] [CrossRef]
- Soták, M.; Czeranková, O.; Klein, D.; Jurčacková, Z.; Li, L.; Čellárová, E. Comparative Transcriptome Reconstruction of Four Hypericum Species Focused on Hypericin Biosynthesis. Front. Plant Sci. 2016, 7, 1039. [Google Scholar] [CrossRef] [Green Version]
- Ionescu, E.; Iordache, T.V.; Tebrencu, C.E.; Cretu, R.M.; Florea, A.M.; Radu, A.L.; Zaharia, A.; Sarbu, A. Evaluating the Content of Active Principles from Wild Hypericum perforatum L. in Various Harvesting Seasons. Rev. Chim. 2018, 69, 1892–1897. [Google Scholar] [CrossRef]
- Briskin, D.P.; Leroy, A.; Gawienowski, M. Influence of nitrogen on the production of hypericins by St. John’s wort. Plant Physiol. Biochem. 2000, 38, 413–420. [Google Scholar] [CrossRef]
- Gray, D.E.; Pallardy, S.G.; Garrett, H.E.; Rottinghaus, G.E. Effect of Acute Drought Stress and Time of Harvest on Phytochemistry and Dry Weight of St. John’s Wort Leaves and Flowers. Planta Medica 2003, 69, 1024–1030. [Google Scholar] [CrossRef]
- Brechner, M.L.; Albright, L.D.; Weston, L.A. Effects of UV-B on Secondary Metabolites of St. John’s Wort (Hypericum perforatum L.) Grown in Controlled Environments. Photochem. Photobiol. 2011, 87, 680–684. [Google Scholar] [CrossRef] [PubMed]
- Germ, M.; Stibilj, V.; Kreft, S.; Gaberščik, A.; Kreft, I. Flavonoid, tannin and hypericin concentrations in the leaves of St. John’s wort (Hypericum perforatum L.) are affected by UV-B radiation levels. Food Chem. 2010, 122, 471–474. [Google Scholar] [CrossRef]
- Tavakoli, F.; Rafieiolhossaini, M.; Ravash, R.; Ebrahimi, M. UV-B radiation and low temperature promoted hypericin biosyn-thesis in adventitious root culture of Hypericum perforatum. Plant Signal. Behav. 2020, 15, 1764184. [Google Scholar] [CrossRef]
- Najafabadi, A.S.; Khanahmadi, M.; Ebrahimi, M.; Moradi, K.; Behroozi, P.; Noormohammadi, N. Effect of different quality of light on growth and production of secondary metabolites in adventitious root cultivation of Hypericum perforatum. Plant Signal. Behav. 2019, 14, 1640561. [Google Scholar] [CrossRef]
- Yao, Y.Y.; Kang, T.L.; Jin, L.; Liu, Z.H.; Zhang, Z.; Xing, H.; Sun, P.; Li, M.F. Temperature-dependent growth and hypericin biosynthesis in Hypericum perforatum. Plant Physiol. Biochem. 2019, 139, 613–619. [Google Scholar] [CrossRef]
- Zobayed, S.; Afreen, F.; Kozai, T. Temperature stress can alter the photosynthetic efficiency and secondary metabolite concentrations in St. John’s wort. Plant Physiol. Biochem. 2005, 43, 977–984. [Google Scholar] [CrossRef]
- Bok, J.W.; Chiang, Y.-M.; Szewczyk, E.; Reyes-Dominguez, Y.; Davidson, A.D.; Sanchez, J.F.; Lo, H.-C.; Watanabe, K.; Strauss, J.; Oakley, B.R.; et al. Chromatin-level regulation of biosynthetic gene clusters. Nat. Chem. Biol. 2009, 5, 462–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, Y.-M.; Szewczyk, E.; Davidson, A.D.; Entwistle, R.; Keller, N.P.; Wang, C.C.C.; Oakley, B.R. Characterization of the Aspergillus nidulans Monodictyphenone Gene Cluster. Appl. Environ. Microbiol. 2010, 76, 2067–2074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzo, P.; Altschmied, L.; Ravindran, B.M.; Rutten, T.; D’Auria, J.C. The Biochemical and Genetic Basis for the Biosynthesis of Bioactive Compounds in Hypericum Perforatum, L. One of the Largest Medicinal Crops in Europe. Genes 2020, 11, 1210. [Google Scholar] [CrossRef]
- Brunáková, K.; Petijová, L.; Zámecník, J.; Turecková, V.; Cellárová, E. The role of ABA in the freezing injury avoidance in two Hypericum species differing in frost tolerance and potential to synthesize hypericins. Plant. Cell. Tiss. Org. 2015, 122, 45–56. [Google Scholar] [CrossRef]
- Velada, I.; Ragonezi, C.; Arnholdt-Schmitt, B.; Cardoso, H. Reference Genes Selection and Normalization of Oxidative Stress Responsive Genes upon Different Temperature Stress Conditions in Hypericum perforatum L. PLoS ONE 2014, 9, e115206. [Google Scholar] [CrossRef]
- Bais, H.P.; Vepachedu, R.; Lawrence, C.B.; Stermitz, F.R.; Vivanco, J.M. Molecular and Biochemical Characterization of an Enzyme Responsible for the Formation of Hypericin in St. John’s Wort (Hypericum perforatum L.). J. Biol. Chem. 2003, 278, 32413–32422. [Google Scholar] [CrossRef] [Green Version]
- Košuth, J.; Smelcerovic, A.; Borsch, T.; Zuehlke, S.; Karppinen, K.; Spiteller, M.; Hohtola, A.; Čellárová, E. The hyp-1 gene is not a limiting factor for hypericin biosynthesis in the genus Hypericum. Funct. Plant Biol. 2011, 38, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.; Kim, Y.-J.; Sukweenadhi, J.; Rahimi, S.; Kwon, W.-S.; Yang, D.-C. Molecular characterization of 5-chlorophyll a/b-binding protein genes from Panax ginseng Meyer and their expression analysis during abiotic stresses. Photosynth. Res. 2016, 54, 446–458. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology; Prue, D.V., Fosket, D.E., Amasino, R., Eds.; The control of flowering; Sinauer Associates Inc.: Sunderland, MA, USA, 2010. [Google Scholar]
- Ludwig, M.; Burnell, J.N. Molecular comparison of carbonic anhydrase from Flaveria species demonstrating different photo-synthetic pathways. Plant Mol. Biol. 1995, 29, 353–365. [Google Scholar] [CrossRef]
- Hondred, D.; Hanson, A.D. Hypoxically inducible barley lactate dehydrogenase: cDNA cloning and molecular analysis. Proc. Natl. Acad. Sci. USA 1990, 87, 7300–7304. [Google Scholar] [CrossRef] [Green Version]
- Jedrzejas, M.J. Structure, function, and evolution of phosphoglycerate mutases: Comparison with fructose-2,6-bisphosphatase, acid phosphatase, and alkaline phosphatase. Prog. Biophys. Mol. Biol. 2000, 73, 263–287. [Google Scholar] [CrossRef]
- Singhania, R.R.; Patel, A.K.; Sukumaran, R.K.; Larroche, C.; Pandey, A. Role and significance of beta-glucosidases in the hydrolysis of cellulose for bioethanol production. Bioresour. Technol. 2013, 127, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Yap, K.L.; Yuan, T.; Mal, T.K.; Vogel, H.J.; Ikura, M. Structural Basis for Simultaneous Binding of Two Carboxy-terminal Peptides of Plant Glutamate Decarboxylase to Calmodulin. J. Mol. Biol. 2003, 328, 193–204. [Google Scholar] [CrossRef]
- Shih, M.C.; Lazar, G.; Goodman, H.M. Evidence in favor of the symbiotic origin of chloroplasts: Primary structure and evolution of tobacco glyceraldehyde-3-phosphate dehydrogenases. Cell 1986, 47, 73–80. [Google Scholar] [CrossRef]
- Li, M.; Welti, R.; Wang, X. Quantitative profiling of Arabidopsis polar glycerolipids in response to phosphorus starvation. Roles of phospholipases D zeta1 and D zeta2 in phosphatidylcholine hydrolysis and digalactosyldiacylglycerol accumulation in phosphorus-starved plants. Plant Physiol. 2006, 142, 750–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudareva, N.; D’Auria, J.C.; Nam, K.H.; Raguso, R.A.; Pichersky, E. Acetyl-CoA: Benzylalcohol acetyltransferase-an enzyme involved in floral scent production in Clarkia breweri. Plant J. 1998, 14, 297–304. [Google Scholar] [CrossRef] [Green Version]
- Klundt, T.; Bocola, M.; Lütge, M.; Beuerle, T.; Liu, B.; Beerhues, L. A Single Amino Acid Substitution Converts Benzophenone Synthase into Phenylpyrone Synthase. J. Biol. Chem. 2009, 284, 30957–30964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Germain, V.; Rylott, E.L.; Larson, T.R.; Sherson, S.M.; Bechtold, N.; Carde, J.-P.; Bryce, J.H.; Graham, I.A.; Smith, S.M. Requirement for 3-ketoacyl-CoA thiolase-2 in peroxisome development, fatty acid beta-oxidation and breakdown of triacylglyc-erol in lipid bodies of Arabidopsis seedlings. Plant J. 2001, 28, 1–12. [Google Scholar] [CrossRef]
- Ehlting, J.; Hamberger, B.; Million-Rousseau, R.; Werck-Reichhart, D. Cytochromes P450 in phenolic metabolism. Phytochem. Rev. 2006, 5, 239–270. [Google Scholar] [CrossRef]
- Lau, W.; Sattely, E.S. Six enzymes from mayapple that complete the biosynthetic pathway to the etoposide aglycone. Science 2015, 349, 1224–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, J.V.; Kim, K.-W.; Lee, C.; Costa, M.A.; May, G.D.; Crow, J.A.; Davin, L.B.; Lewis, N.G. Next Generation Sequencing in Predicting Gene Function in Podophyllotoxin Biosynthesis. J. Biol. Chem. 2013, 288, 466–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, D.R. Plant cytochrome P450s from moss to poplar. Phytochem. Rev. 2006, 5, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Sun, P.; Kang, T.; Xing, H.; Yang, D.; Zhang, J.; Paré, P.W. Mapping podophyllotoxin biosynthesis and growth-related transcripts with high elevation in Sinopodophyllum hexandrum. Ind. Crop. Prod. 2018, 124, 510–518. [Google Scholar] [CrossRef]
- Brugliera, F.; Barri-Rewell, G.; Holton, T.A.; Mason, J.G. Isolation and characterization of a flavonoid 3′-hydroxylase cDNA clone corresponding to the Ht1 locus of Petunia hybrid. Plant J. 1999, 19, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Akashi, T.; Aoki, T.; Ayabe, S.-I. CYP81E1, a cytochrome P450 cDNA of licorice (Glycyrrhiza echinata L.), encodes isoflavone 2’-hydroxylase. Biochem. Biophys. Res. Commun. 1998, 251, 67–70. [Google Scholar] [CrossRef]
- Greer, S.; Wen, M.; Bird, D.; Wu, X.; Samuels, L.; Kunst, L.; Jetter, R. The Cytochrome P450 Enzyme CYP96A15 Is the Midchain Alkane Hydroxylase Responsible for Formation of Secondary Alcohols and Ketones in Stem Cuticular Wax of Arabidopsis. Plant Physiol. 2007, 145, 653–667. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.-Z.; Wang, G.-F.; Speal, B.; Ma, H. The Excess Microsporocytes1 gene encodes a putative leucine-rich repeat receptor protein kinase that controls somatic and reproductive cell fates in the Arabidopsis anther. Genes Dev. 2002, 16, 2021–2031. [Google Scholar] [CrossRef] [Green Version]
- Caro, E.; Castellano, M.M.; Gutierrez, C. A chromatin link that couples cell division to root epidermis patterning in Arabidopsis. Nat. Cell Biol. 2007, 447, 213–217. [Google Scholar] [CrossRef]
- Moriwaki, T.; Miyazawa, Y.; Kobayashi, A.; Uchida, M.; Watanabe, C.; Fujii, N.; Takahashi, H. Hormonal Regulation of Lateral Root Development in Arabidopsis Modulated by MIZ1 and Requirement of GNOM Activity for MIZ1 Function. Plant Physiol. 2011, 157, 1209–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, R.; Liu, J.X.; Howell, S.H. Proteolytic processing of a precursor protein for a growth-promoting peptide by a sub-tilisin serine protease in Arabidopsis. Plant J. 2008, 56, 219–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, R.C.; Mok, M.C.; Mok, D.W.S. Isolation of a cytokinin gene, ZOG1, encoding zeatin O-glucosyltransferase from Phaseolus lunatus. Proc. Natl. Acad. Sci. USA 1999, 96, 284–289. [Google Scholar] [CrossRef] [Green Version]
- Lindquist, S.; Craig, A.E. The Heat-Shock Proteins. Annu. Rev. Genet. 1988, 22, 631–677. [Google Scholar] [CrossRef] [PubMed]
- Vierling, E. The roles of heat shock proteins in plants. Ann. Rev. Plant. Physiol. Mol. Biol. 1991, 42, 579–620. [Google Scholar] [CrossRef]
- Che, S.; Zhang, L.; Zhang, Z.; Lin, X.Z. Expression analysis of heat-shock protein gene Hsp845 in the Antarctic psychrotrophic bacterium Psychrobacter sp. G under temperature and salinity stress. Adv. Polar. Sci. 2016, 27, 14–20. [Google Scholar]
- Liu, Z.-M.; Zhu, X.-L.; Lu, J.; Cai, W.-J.; Ye, Y.-P.; Lv, Y.-P. Effect of high temperature stress on heat shock protein expression and antioxidant enzyme activity of two morphs of the mud crab Scylla paramamosain. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2018, 223, 10–17. [Google Scholar] [CrossRef]
- Zhou, M.; Shen, C.; Wu, L.; Tang, K.; Lin, J. CBF-dependent signaling pathway: A key responder to low temperature stress in plants. Crit. Rev. Biotechnol. 2010, 31, 186–192. [Google Scholar] [CrossRef]
- Liu, Z.; Jia, Y.; Ding, Y.; Shi, Y.; Li, Z.; Guo, Y.; Gong, Z.; Yang, S. Plasma Membrane CRPK1-Mediated Phosphorylation of 14-3-3 Proteins Induces Their Nuclear Import to Fine-Tune CBF Signaling during Cold Response. Mol. Cell 2017, 66, 117–128.e5. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Yang, S.; Li, Y.; Hua, J. The Arabidopsis BAP1 and BAP2 Genes Are General Inhibitors of Programmed Cell Death. Plant Physiol. 2007, 145, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Zhou, A.; Sun, H.; Feng, S.; Zhou, M.; Gong, S.; Wang, J.; Zhang, S. A novel cold-regulated gene from Phlox subulata, PsCor413im1, enhances low temperature tolerance in Arabidopsis. Biochem. Biophys. Res. Commun. 2018, 495, 1688–1694. [Google Scholar] [CrossRef] [PubMed]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB transcription factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Kitajima, S.; Koyama, T.; Ohme-Takagi, M.; Shinshi, H.; Sato, F. Characterization of Gene Expression of NsERFs, Transcription Factors of Basic PR Genes from Nicotiana sylvestris. Plant Cell Physiol. 2000, 41, 817–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakano, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Genome-Wide Analysis of the ERF Gene Family in Arabidopsis and Rice. Plant Physiol. 2006, 140, 411–432. [Google Scholar] [CrossRef] [Green Version]
- Vannini, C.; Locatelli, F.; Bracale, M.; Magnani, E.; Marsoni, M.; Osnato, M.; Mattana, M.; Baldoni, E.; Coraggio, I. Overex-pression of the rice Osmyb4 gene increases chilling and freezing tolerance of Arabidopsis thaliana plants. Plant J. 2004, 37, 115–127. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Z.; Kang, J.; Kang, D.; Gu, H.; Qin, G. AtMYB14 Regulates Cold Tolerance in Arabidopsis. Plant Mol. Biol. Rep. 2013, 31, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Faria, J.A.Q.A.; Reis, P.A.B.; Reis, M.T.B.; Rosado, G.L.; Pinheiro, G.L.; Mendes, G.C.; Fontes, E.P.B. The NAC domain-containing protein, GmNAC6, is a downstream component of the ER stress- and osmotic stress-induced NRP-mediated cell-death signaling pathway. BMC Plant Biol. 2011, 11, 129. [Google Scholar] [CrossRef] [Green Version]
- Rushton, P.J.; Somssich, I.E.; Ringler, P.; Shen, Q.J. WRKY transcription factors. Trends Plant Sci. 2010, 15, 247–258. [Google Scholar] [CrossRef]
- Li, M.; Lv, M.; Yang, D.; Wei, J.; Xing, H.; Paré, P.W. Temperature-regulated anatomical and gene-expression changes in Sinopodophyllum hexandrum seedlings. Ind. Crop. Prod. 2020, 152, 112479. [Google Scholar] [CrossRef]
- Chi, J.; Franklin, M. Determination of hypericin in plasma by high-performance liquid chromatography. J. Chromatogr. B Biomed. Sci. Appl. 1999, 724, 195–198. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNAseq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Brief Bioinform. 2009, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Brief Bioinform. 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [Green Version]
- Willems, E.; Leyns, L.; Vandesompele, J. Standardization of real-time PCR gene expression data from independent biological replicates. Anal. Biochem. 2008, 379, 127–129. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temp. | Density (mm2) | Opening Percentage (%) | Length (μm) | Width (μm) | Ratio of Length/Width |
|---|---|---|---|---|---|
| 15 °C | 22.53 ± 2.28 * | 91.59 ± 0.08 * | 13.27 ± 1.40 | 5.13 ± 1.01 * | 2.69 ± 0.58 * |
| 22 °C | 19.80 ± 2.39 | 66.21 ± 0.05 | 13.42 ± 1.31 | 4.44 ± 1.23 | 3.20 ± 0.78 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, H.; Li, J.; Chen, S.; Sun, P.; Xing, H.; Yang, D.; Zhang, X.; Li, M.; Wei, J. Physiological and Transcriptomic Analysis Provide Insight into Low Temperature Enhancing Hypericin Biosynthesis in Hypericum perforatum. Molecules 2021, 26, 2294. https://doi.org/10.3390/molecules26082294
Su H, Li J, Chen S, Sun P, Xing H, Yang D, Zhang X, Li M, Wei J. Physiological and Transcriptomic Analysis Provide Insight into Low Temperature Enhancing Hypericin Biosynthesis in Hypericum perforatum. Molecules. 2021; 26(8):2294. https://doi.org/10.3390/molecules26082294
Chicago/Turabian StyleSu, Hongyan, Jie Li, Sijin Chen, Ping Sun, Hua Xing, Delong Yang, Xiaona Zhang, Mengfei Li, and Jianhe Wei. 2021. "Physiological and Transcriptomic Analysis Provide Insight into Low Temperature Enhancing Hypericin Biosynthesis in Hypericum perforatum" Molecules 26, no. 8: 2294. https://doi.org/10.3390/molecules26082294
APA StyleSu, H., Li, J., Chen, S., Sun, P., Xing, H., Yang, D., Zhang, X., Li, M., & Wei, J. (2021). Physiological and Transcriptomic Analysis Provide Insight into Low Temperature Enhancing Hypericin Biosynthesis in Hypericum perforatum. Molecules, 26(8), 2294. https://doi.org/10.3390/molecules26082294