
Anti-Inflammatory Activities of the Ethanol Extract of Prasiola japonica, an Edible Freshwater Green Algae, and Its Various Solvent Fractions in LPS-Induced Macrophages and Carrageenan-Induced Paw Edema via the AP-1 Pathway

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Results
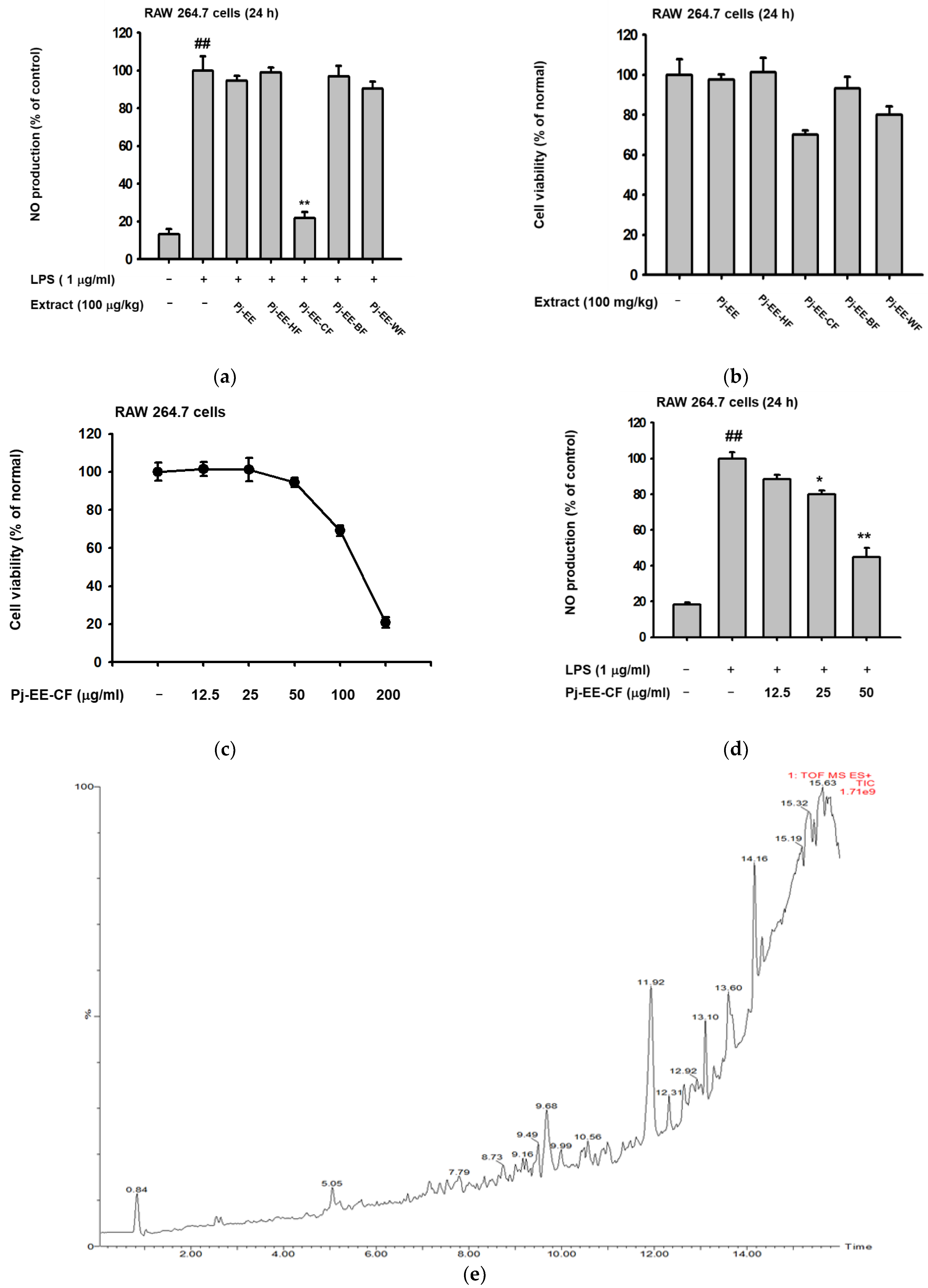
2.1. Effect of Pj-EE and Its Various Solvent Fractions on the NO Production of LPS-Induced Macrophages
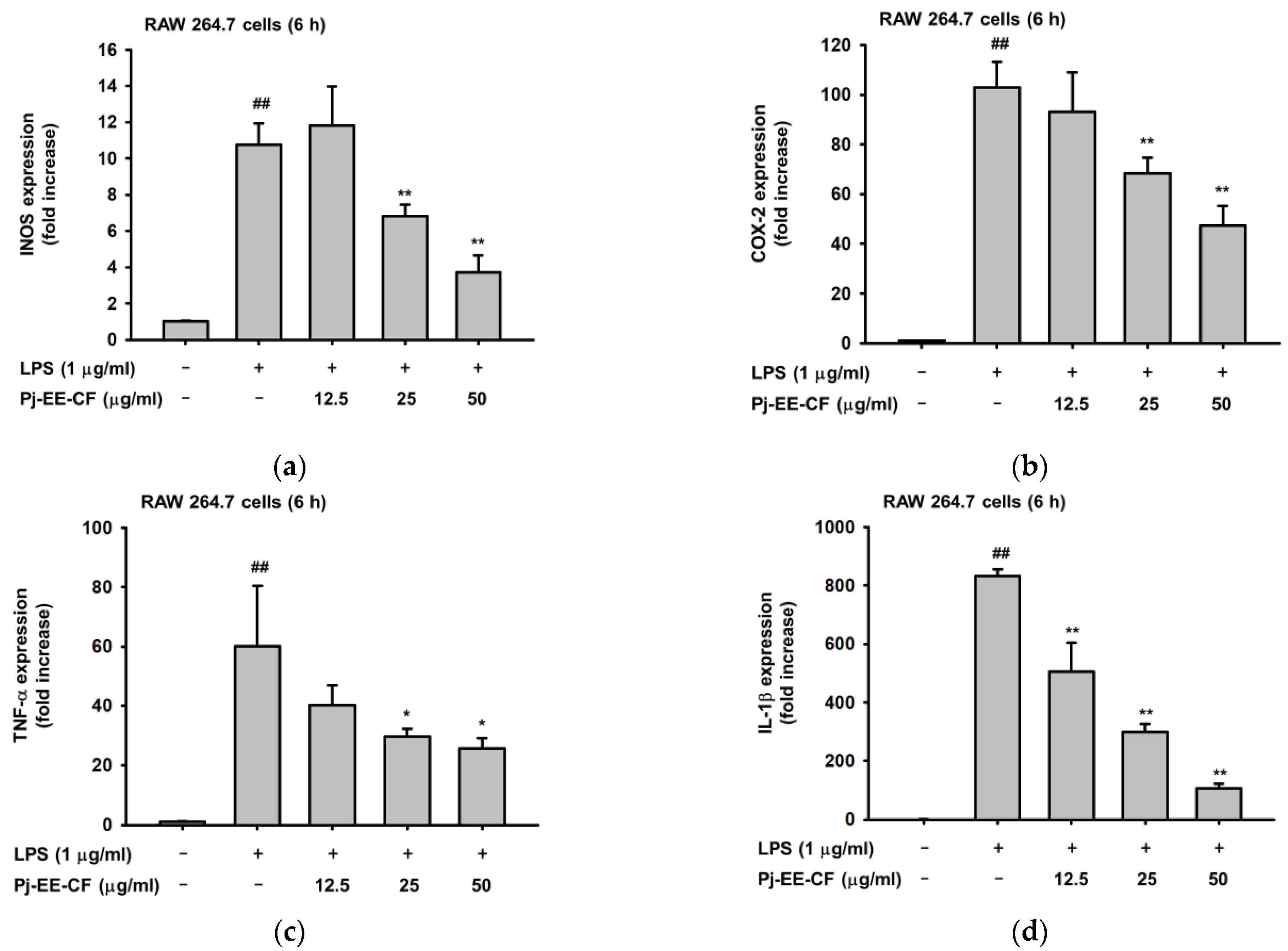
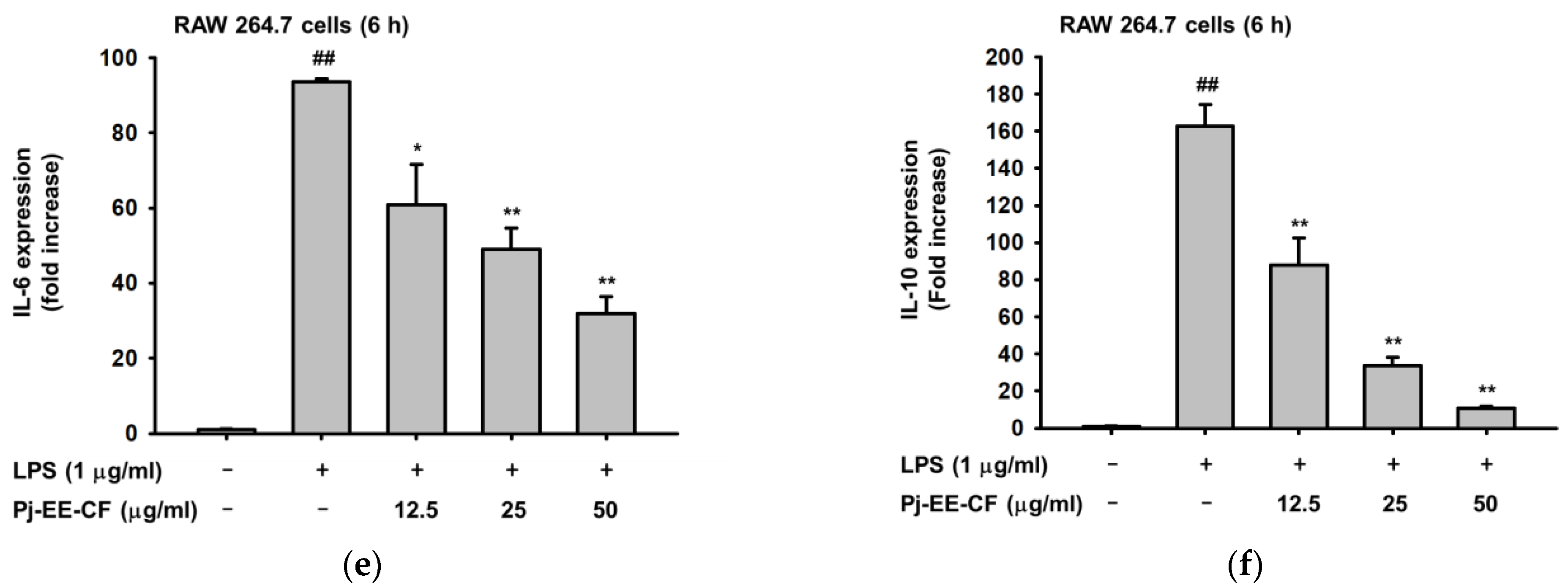
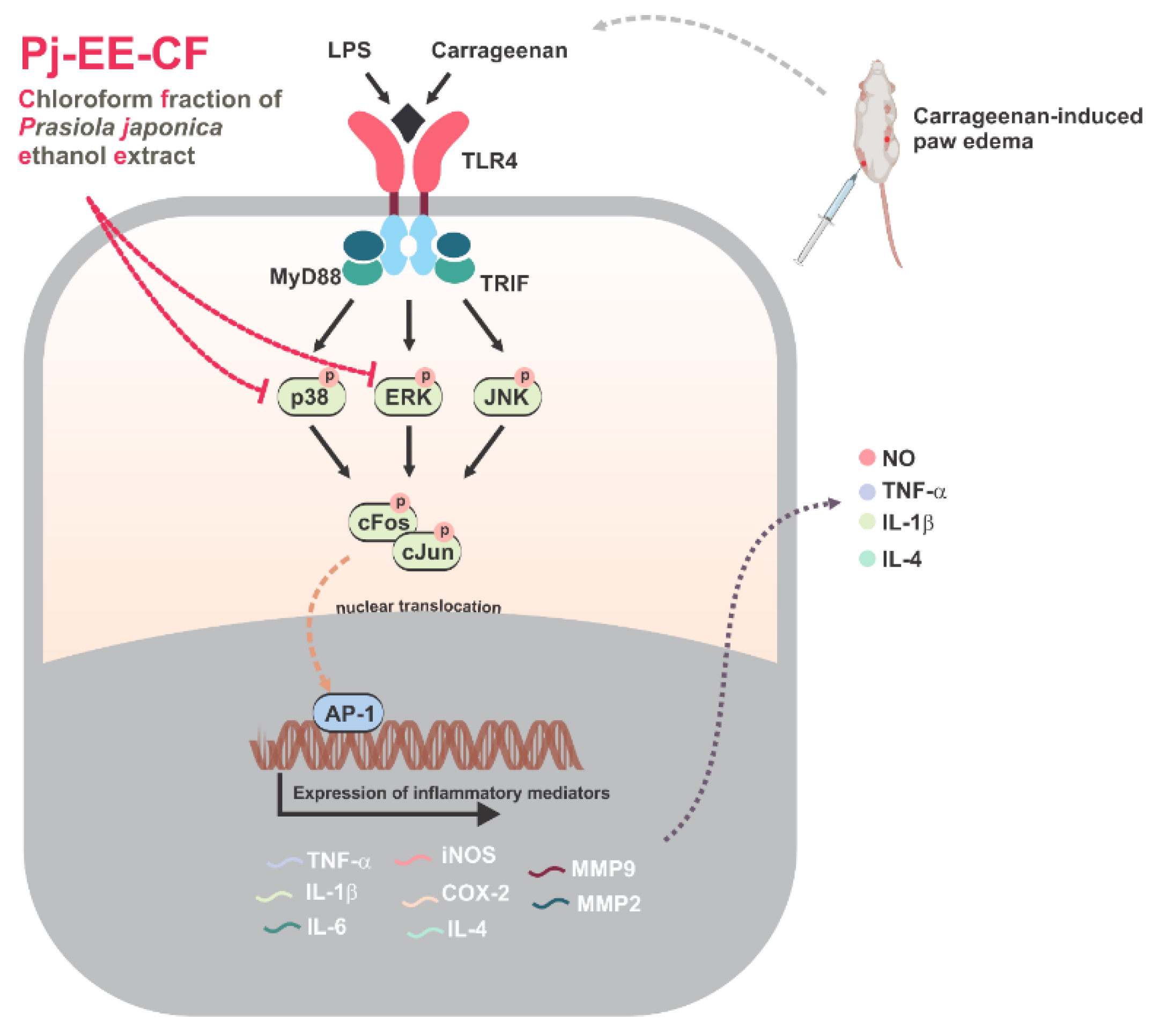
2.2. Effects of Pj-EE-CF on the Expression of Pro-Inflammatory Genes on LPS-Induced Macrophages
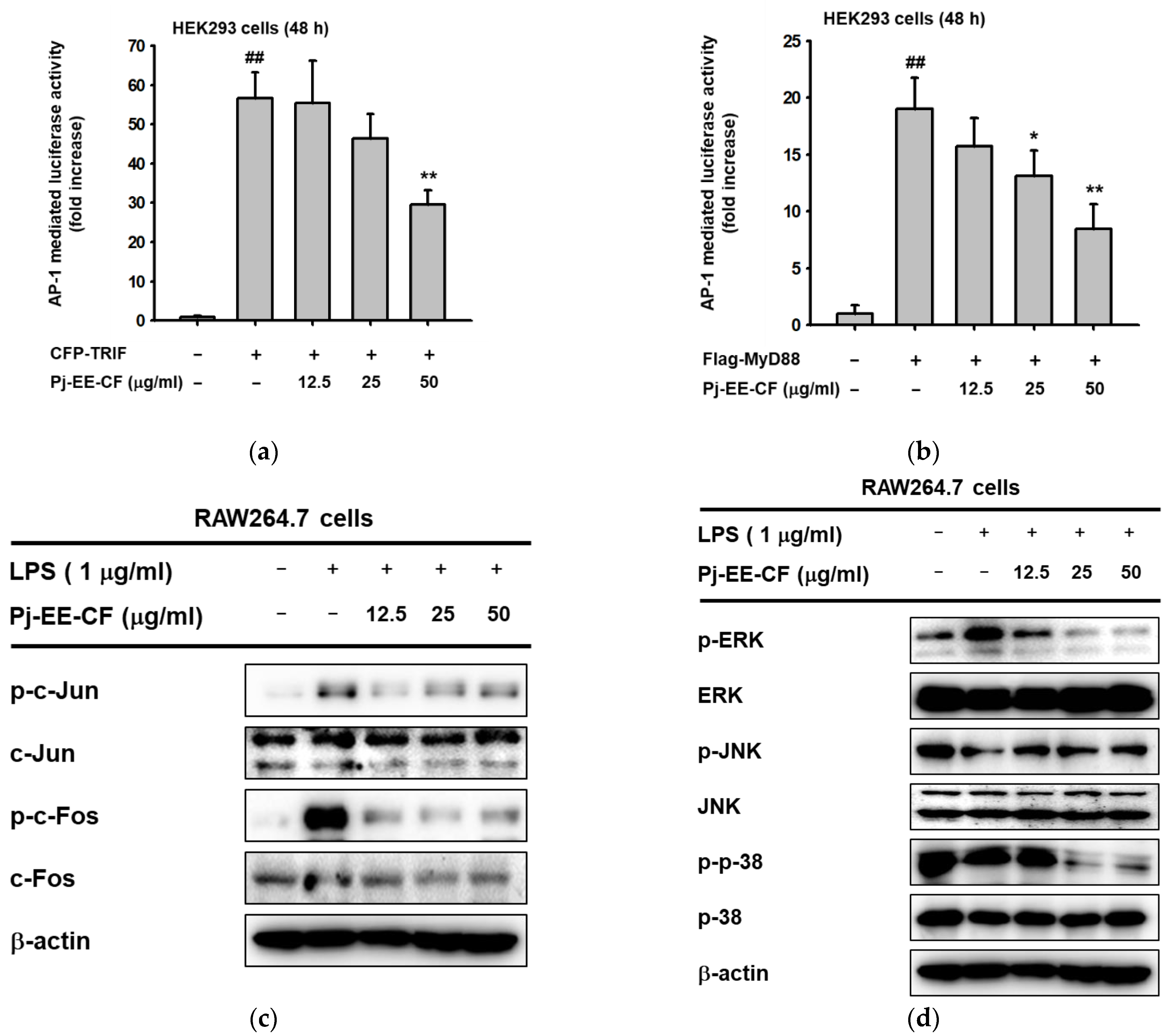
2.3. Effects of Pj-EE-CF on the Transcriptional Activation of AP-1 Signaling and the Upstream Signaling Molecules of AP-1 Activation
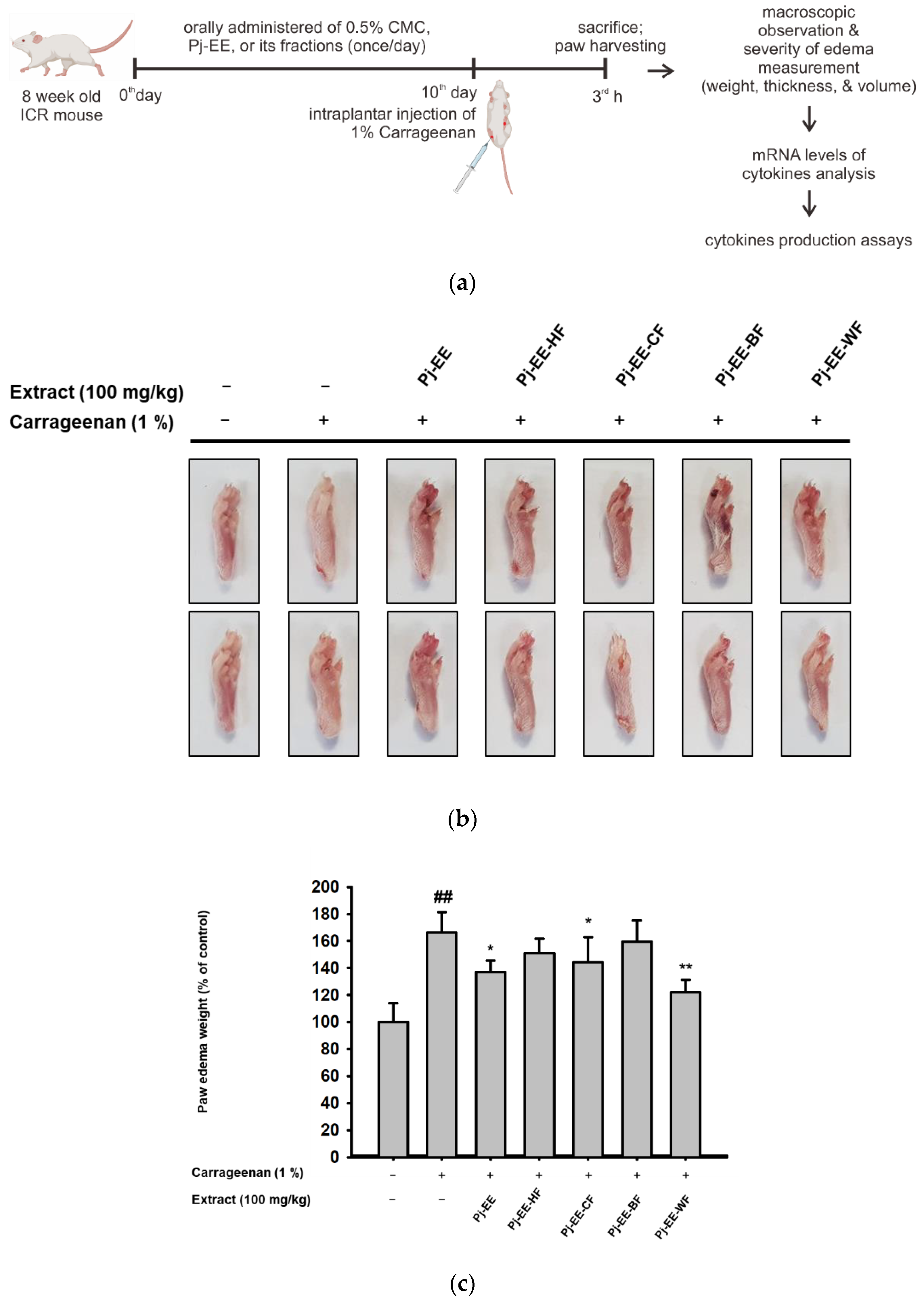
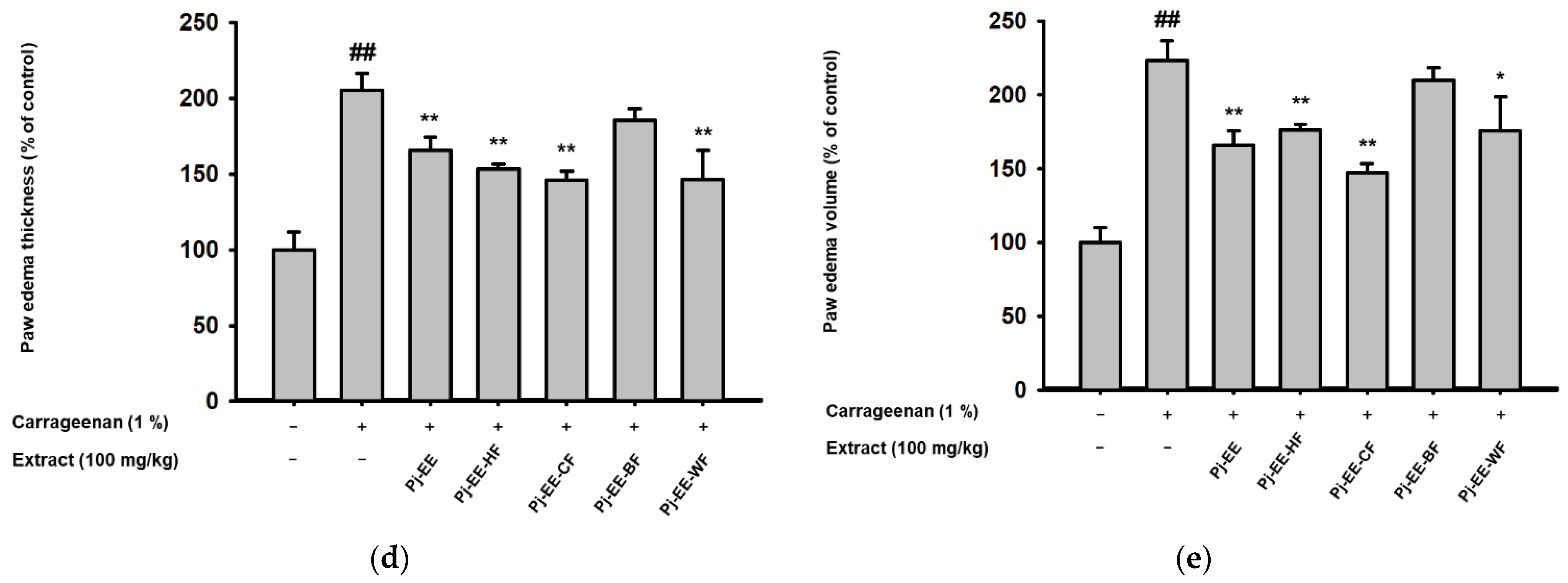
2.4. Effects of Pj-EE and Its Solvent Fractions on a Carrageenan-Induced Acute Paw Edema Model
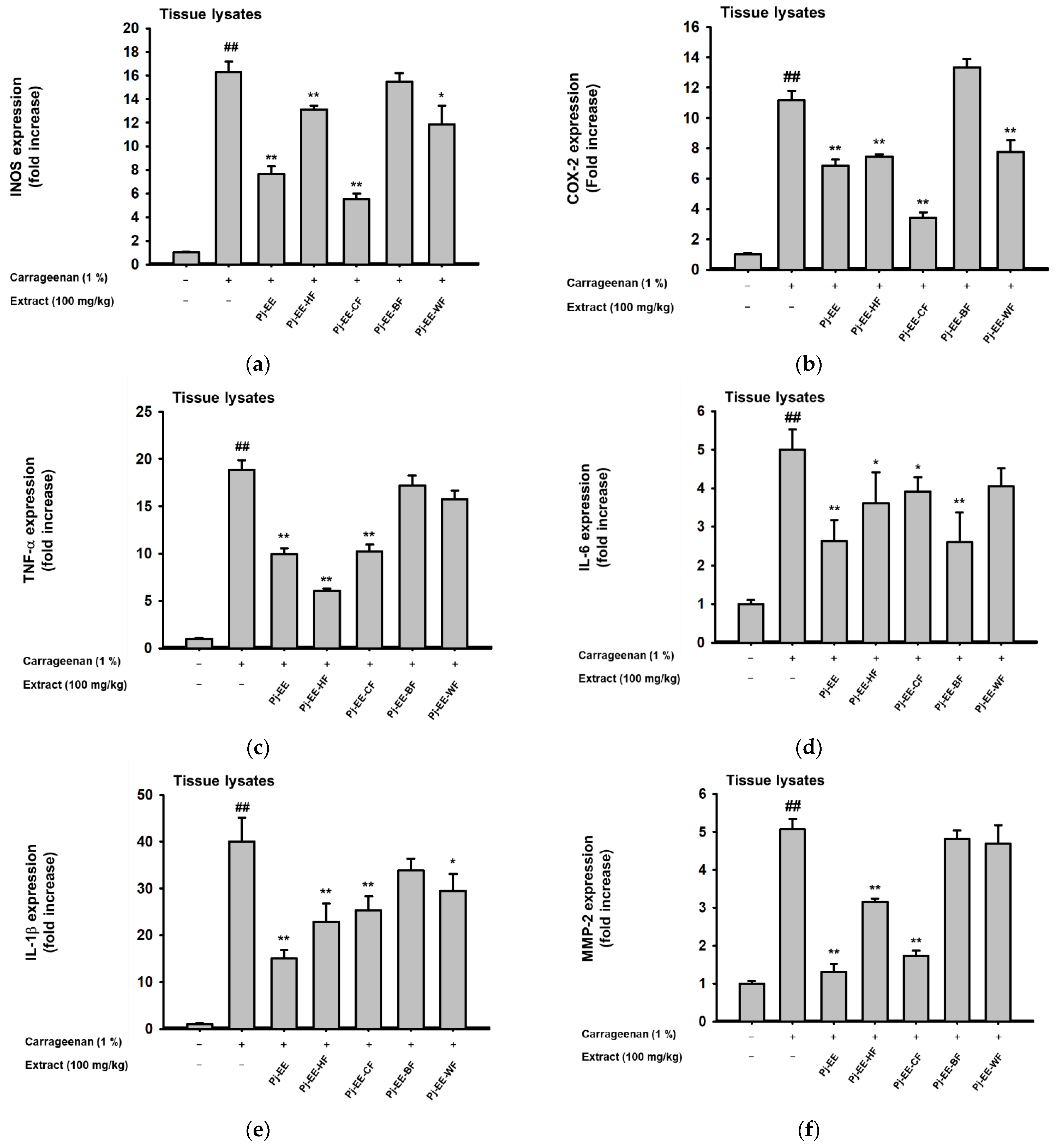
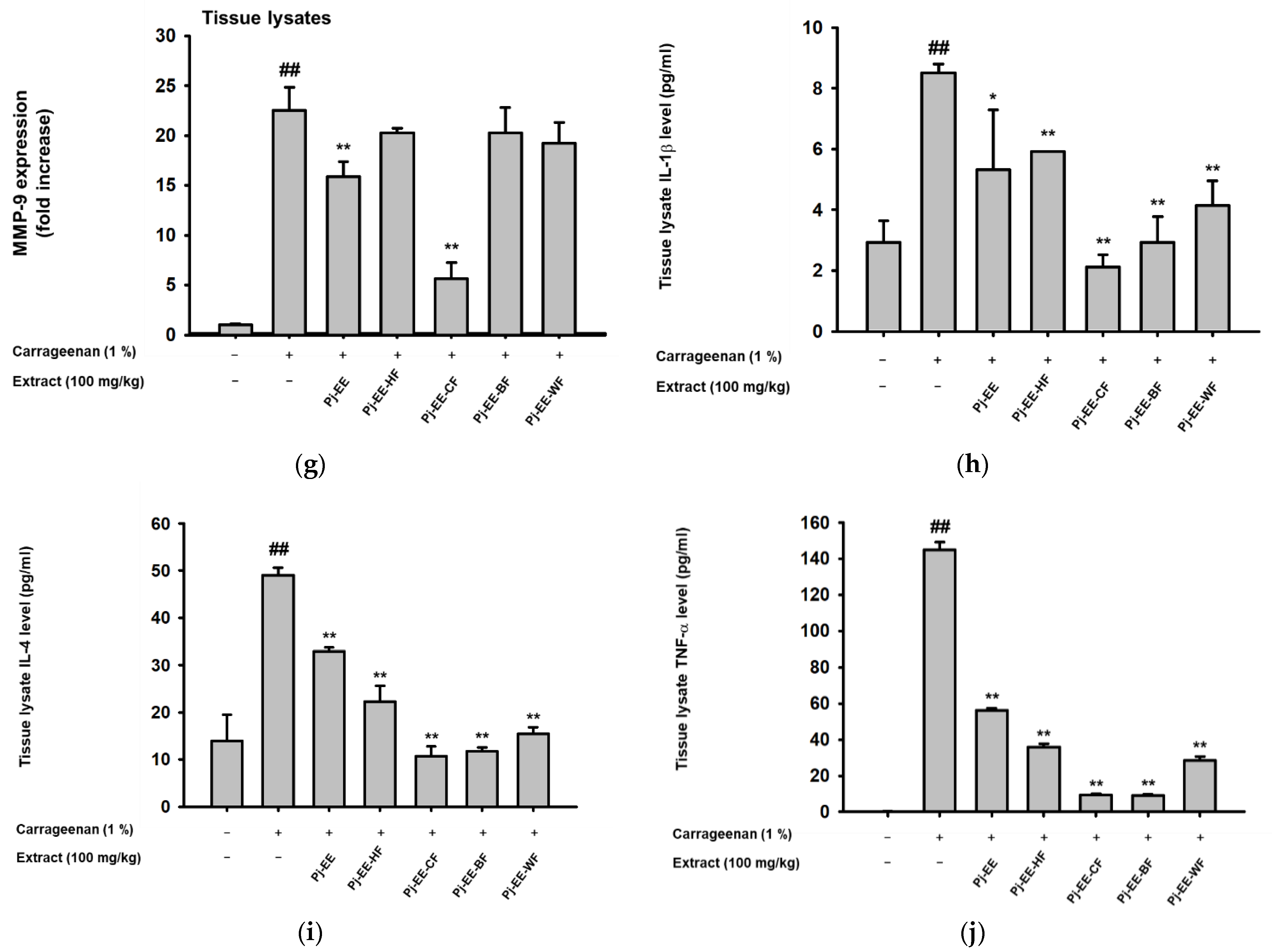
2.5. Effects of Pj-EE and Its Solvent Fractions on Pro-Inflammatory Genes and Cytokines Production in Carrageenan-Induced Acute Paw Edema Models
3. Discussion
4. Materials and Methods
4.1. Materials
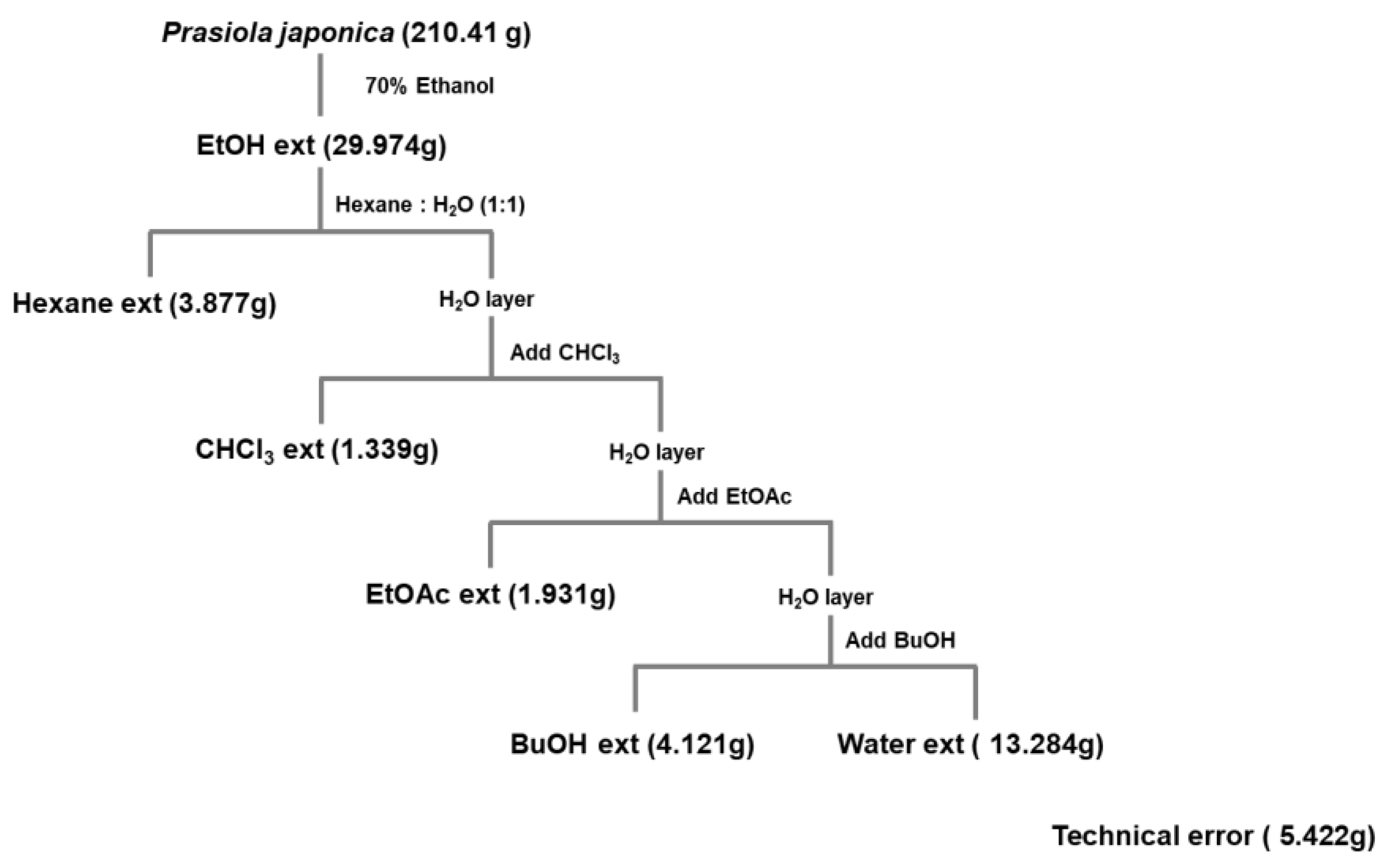
4.2. Pj-EE and Preparation of Its Solvent Fractions
4.3. Cell Culture and Treatment
4.4. Determination of NO and Cytokines Production
4.5. Cell Viability Assay
4.6. Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS)
4.7. mRNA Analysis by Quantitative Real-Time Polymerase Chain Reaction (qPCR)
4.8. Luciferase Reporter Gene Assay
4.9. Cell Lysate Extraction and Immunoblotting Analysis
4.10. Animals
4.11. Carrageenan-Induced Acute Paw Edema Mouse Model
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
| LPS | Lipopolysaccharide |
| MyD88 | Myeloid differentiation factor 88 |
| AP-1 | Activator protein-1 |
| MAPKs | Mitogen-activated protein kinases |
| JNK | c-Jun N-terminal kinase |
| RPMI 1640 | Roswell Park Memorial Institute 1640 |
| DMEM | Dulbecco’s modified Eagle’s medium |
| COX-2 | Cyclooxygenase-2 |
| MMP | Matrix metalloproteinase |
| IL-1β | Interleukin-1β |
| IL-6 | Interleukin 6 |
| TNF-α | Tumor necrosis factor alpha |
| MKK | Mitogen-activated protein kinase |
| iNOS | Inducible nitric oxide synthase |
References
- Kumar, H.; Kawai, T.; Akira, S. Pathogen recognition by the innate immune system. Int. Rev. Immunol. 2011, 30, 16–34. [Google Scholar] [CrossRef] [PubMed]
- Mogensen, T.H. Pathogen recognition and inflammatory signaling in innate immune defenses. Clin. Microbiol. Rev. 2009, 22, 240–273. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2017, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssens, S.; Beyaert, R. Role of toll-like receptors in pathogen recognition. Clin. Microbiol. Rev. 2003, 16, 637–646. [Google Scholar] [CrossRef] [Green Version]
- Kuzmich, N.N.; Sivak, K.V.; Chubarev, V.N.; Porozov, Y.B.; Savateeva-Lyubimova, T.N.; Peri, F. TLR4 signaling pathway modulators as potential therapeutics in inflammation and sepsis. Vaccines 2017, 5, 34. [Google Scholar] [CrossRef] [Green Version]
- Lopes, A.H.; Silva, R.L.; Fonseca, M.D.; Gomes, F.I.; Maganin, A.G.; Ribeiro, L.S.; Marques, L.M.M.; Cunha, F.Q.; Alves-Filho, J.C.; Zamboni, D.S.; et al. Molecular basis of carrageenan-induced cytokines production in macrophages. Cell Commun. Signal. 2020, 18, 141. [Google Scholar] [CrossRef]
- Yang, Y.; Kim, S.C.; Yu, T.; Yi, Y.S.; Rhee, M.H.; Sung, G.H.; Yoo, B.C.; Cho, J.Y. Functional roles of p38 mitogen-activated protein kinase in macrophage-mediated inflammatory responses. Mediat. Inflamm. 2014, 2014, 352371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aktan, F. iNOS-mediated nitric oxide production and its regulation. Life Sci. 2004, 75, 639–653. [Google Scholar] [CrossRef]
- Cheng, G.; Wei, L.; Xiurong, W.; Xiangzhen, L.; Shiguang, Z.; Songbin, F. IL-17 stimulates migration of carotid artery vascular smooth muscle cells in an MMP-9 dependent manner via p38 MAPK and ERK1/2-dependent NF-kappaB and AP-1 activation. Cell. Mol. Neurobiol. 2009, 29, 1161–1168. [Google Scholar] [CrossRef]
- Dinarello, C.A. Proinflammatory cytokines. Chest 2000, 118, 503–508. [Google Scholar] [CrossRef]
- Manicone, A.M.; McGuire, J.K. Matrix metalloproteinases as modulators of inflammation. Semin. Cell Dev. Biol. 2008, 19, 34–41. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Guo, J.; Feng, C.; Ke, Z.; Chen, L.; Pan, Y. The preoperative platelet-lymphocyte ratio versus neutrophil-lymphocyte ratio: Which is better as a prognostic factor in oral squamous cell carcinoma? Ther. Adv. Med. Oncol. 2016, 8, 160–167. [Google Scholar] [CrossRef] [Green Version]
- Park, S.H.; Choi, E.; Kim, S.; Kim, D.S.; Kim, J.H.; Chang, S.; Choi, J.S.; Park, K.J.; Roh, K.B.; Lee, J.; et al. Oxidative stress-protective and anti-melanogenic effects of loliolide and ethanol extract from fresh water green algae, Prasiola japonica. Int. J. Mol. Sci. 2018, 19, 2825. [Google Scholar] [CrossRef] [Green Version]
- Saini, D.K.; Pabbi, S.; Shukla, P. Cyanobacterial pigments: Perspectives and biotechnological approaches. Food Chem. Toxicol. 2018, 120, 616–624. [Google Scholar] [CrossRef] [PubMed]
- Drew, K.M.; Friedmann, I. Occurrence of motile gametes in Prasiola stipitata Suhr. Nature 1957, 180, 557–558. [Google Scholar] [CrossRef]
- Morphology and phylogenetic position of a freshwater Prasiola species (Prasiolales, Chlorophyta) in Korea. Algae 2015, 30, 197–205. [CrossRef] [Green Version]
- Naw, M.W.D.; Hara, Y. Morphology and molecular phylogeny of Prasiola sp. (Prasiolales, Chlorophyta) from Myanmar. Phycol. Res. 2002, 50, 175–182. [Google Scholar] [CrossRef]
- Da Woom, S.; Hee Jung, K.; Su Kil, J.; Mansig, J.; Seong Soo, J. Screening of functional components derived from fresh water laver, Prasiola japonica, and its pharmacological properties. J. Biomed. Res. 2013, 14, 83–90. [Google Scholar]
- Choi, E.; Yi, Y.-S.; Lee, J.; Park, S.H.; Kim, S.; Hossain, M.A.; Jang, S.; Choi, Y.I.; Park, K.J.; Kim, D.S.; et al. Anti-apoptotic and anti-inflammatory activities of edible fresh water algae Prasiola japonica in UVB-irradiated skin keratinocytes. Am. J. Chin. Med. 2019, 47, 1853–1868. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.Y.; Park, S.H.; Lim, H.Y.; Jang, S.G.; Park, K.J.; Kim, D.S.; Kim, J.H.; Cho, J.Y. In vivo anti-inflammatory effects of Prasiola japonica ethanol extract. J. Funct. Foods 2021, 80, 104440. [Google Scholar] [CrossRef]
- Aziz, N.; Son, Y.J.; Cho, J.Y. Thymoquinone suppresses IRF-3-mediated expression of Type I interferons via suppression of TBK1. Int. J. Mol. Sci. 2018, 19, 1355. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.P.; Son, K.H.; Chang, H.W.; Kang, S.S. Anti-inflammatory plant flavonoids and cellular action mechanisms. J. Pharmacol. Sci. 2004, 96, 229–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahmawati, L.; Aziz, N.; Oh, J.; Hong, Y.H.; Woo, B.Y.; Hong, Y.D.; Manilack, P.; Souladeth, P.; Jung, J.H.; Lee, W.S.; et al. Cissus subtetragona Planch. ameliorates inflammatory responses in LPS-induced macrophages, HCl/EtOH-induced fastritis, and LPS-induced lung injury via attenuation of Src and TAK1. Molecules 2021, 26, 6073. [Google Scholar] [CrossRef]
- Park, B.S.; Lee, J.-O. Recognition of lipopolysaccharide pattern by TLR4 complexes. Exp. Mol. Med. 2013, 45, e66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, Y.-S.; Cho, J.Y.; Kim, D. Cerbera manghas methanol extract exerts anti-inflammatory activity by targeting c-Jun N-terminal kinase in the AP-1 pathway. J. Ethnopharmacol. 2016, 193, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Mansouri, M.T.; Hemmati, A.A.; Naghizadeh, B.; Mard, S.A.; Rezaie, A.; Ghorbanzadeh, B. A study of the mechanisms underlying the anti-inflammatory effect of ellagic acid in carrageenan-induced paw edema in rats. Indian J. Pharmacol. 2015, 47, 292–298. [Google Scholar] [CrossRef] [Green Version]
- Winter, C.A.; Risley, E.A.; Nuss, G.W. Carrageenin-induced edema in hind paw of the rat as an assay for antiiflammatory drugs. Proc. Soc. Exp. Biol. Med. Soc. Exp. Biol. Med. 1962, 111, 544–547. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Maitituersun, A.; Li, C.; Li, Q.; Xu, F.; Liu, T. Evaluation on analgesic and anti-inflammatory activities of total flavonoids from Juniperus sabina. Evid. -Based Complementary Altern. Med. Ecam 2018, 2018, 7965306. [Google Scholar] [CrossRef] [Green Version]
- Hämäläinen, M.; Nieminen, R.; Vuorela, P.; Heinonen, M.; Moilanen, E. Anti-inflammatory effects of flavonoids: Genistein, kaempferol, quercetin, and daidzein inhibit STAT-1 and NF-kappaB activations, whereas flavone, isorhamnetin, naringenin, and pelargonidin inhibit only NF-kappaB activation along with their inhibitory effect on iNOS expression and NO production in activated macrophages. Mediat. Inflamm. 2007, 2007, 45673. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Park, J.G.; Lee, J.; Yang, W.S.; Park, G.W.; Kim, H.G.; Yi, Y.S.; Baek, K.S.; Sung, N.Y.; Hossen, M.J.; et al. The dietary flavonoid kaempferol mediates anti-inflammatory responses via the Src, Syk, IRAK1, and IRAK4 molecular targets. Mediat. Inflamm. 2015, 2015, 904142. [Google Scholar] [CrossRef]
- Survay, N.S.; Upadhyaya, C.P.; Kumar, B.; Young, K.E.; Yoon, D.-Y.; Park, S.-W. New genera of flavonols and flavonol derivatives as therapeutic molecules. J. Korean Soc. Appl. Biol. Chem. 2011, 54, 1–18. [Google Scholar] [CrossRef]
- Park, I.; Goo, D.; Nam, H.; Wickramasuriya, S.S.; Lee, K.; Zimmerman, N.P.; Smith, A.H.; Rehberger, T.G.; Lillehoj, H.S. Effects of dietary maltol on innate immunity, gut health, and growth performance of broiler chickens challenged with Eimeria maxima. Front. Vet. Sci. 2021, 8, 508. [Google Scholar] [CrossRef]
- Liao, W.; Liu, Z.; Zhang, T.; Sun, S.; Ye, J.; Li, Z.; Mao, L.; Ren, J. Enhancement of anti-inflammatory properties of nobiletin in macrophages by a nano-emulsion preparation. J. Agric. Food Chem. 2018, 66, 91–98. [Google Scholar] [CrossRef]
- Jeong, J.W.; Lee, H.H.; Han, M.H.; Kim, G.Y.; Kim, W.J.; Choi, Y.H. Anti-inflammatory effects of genistein via suppression of the toll-like receptor 4-mediated signaling pathway in lipopolysaccharide-stimulated BV2 microglia. Chem. -Biol. Interact. 2014, 212, 30–39. [Google Scholar] [CrossRef]
- Wang, Z.; Hao, W.; Hu, J.; Mi, X.; Han, Y.; Ren, S.; Jiang, S.; Wang, Y.; Li, X.; Li, W. Maltol improves APAP-induced hepatotoxicity by inhibiting oxidative stress and inflammation response via NF-κB and PI3K/Akt signal pathways. Antioxidants 2019, 8, 395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, X.; Jin, D.; Zhang, Y.; Zhou, R.; Zhang, Y.; Lian, F. Systematic elucidation of the mechanism of Sappan lignum in the treatment of diabetic peripheral neuropathy based on network pharmacology. Evid. -Based Complementary Altern. Med. 2021, 2021, 5528018. [Google Scholar] [CrossRef] [PubMed]
- Anand, K.K.; Sharma, M.L.; Singh, B.; Ghatak, B.J. Antiinflamatory, antipyretic & analgesic properties of bavachinin-a flavanone isolated from seeds of Psoralea corylifolia Linn. (Babchi). Indian J. Exp. Biol. 1978, 16, 1216–1217. [Google Scholar]
- Zeng, K.W.; Yu, Q.; Song, F.J.; Liao, L.X.; Zhao, M.B.; Dong, X.; Jiang, Y.; Tu, P.F. Deoxysappanone B, a homoisoflavone from the Chinese medicinal plant Caesalpinia sappan L., protects neurons from microglia-mediated inflammatory injuries via inhibition of IκB kinase (IKK)-NF-κB and p38/ERK MAPK pathways. Eur. J. Pharm. 2015, 748, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.G.; Kim, M.Y.; Cho, J.Y. Alisma canaliculatum ethanol extract suppresses inflammatory responses in LPS-stimulated macrophages, HCl/EtOH-induced gastritis, and DSS-triggered colitis by targeting Src/Syk and TAK1 activities. J. Ethnopharmacol. 2018, 219, 202–212. [Google Scholar] [CrossRef]
- Misko, T.P.; Schilling, R.J.; Salvemini, D.; Moore, W.M.; Currie, M.G. A Fluorometric assay for the measurement of nitrite in biological samples. Anal. Biochem. 1993, 214, 11–16. [Google Scholar] [CrossRef]
- Gerlier, D.; Thomasset, N. Use of MTT colorimetric assay to measure cell activation. J. Immunol. Methods 1986, 94, 57–63. [Google Scholar] [CrossRef]
- Hong, Y.H.; Kim, J.H.; Cho, J.Y. Ranunculus bulumei methanol extract exerts anti-inflammatory activity by targeting Src/Syk in NF-κB signaling. Biomolecules 2020, 10, 546. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PCR Type | Genes Name | Sequence (5′-3′) | |
|---|---|---|---|
| q PCR | GAPDH | Forward | GAAGGTCGGTGTGAACGGAT |
| Reverse | AGTGATGGCATGGACTGTGG | ||
| iNOS | Forward | CAAGAGAACGGAGAACGGAGA | |
| Reverse | GATGGACCCCAAGCAAGACT | ||
| COX-2 | Forward | TGAGTACCGCAAACGCTTCT | |
| Reverse | TGGGAGGCACTTGCATTGAT | ||
| TNF-α | Forward | TTGACCTCAGCGCTGAGTTG | |
| Reverse | CCTGTAGCCCACGTCGTAGC | ||
| IL-1β | Forward | CAGGATGAGGACATGAGCACC | |
| Reverse | CTCTGCAGACTCAAACTCCAC | ||
| IL-6 | Forward | GCCTTCTTGGGACTGATGCT | |
| Reverse | TGGAAATTGGGGTAGGAAGGAC | ||
| IL-4 | Forward | GGTCTCAACCCCCAGCTAGT | |
| Reverse | GCCCATGATCTCTCTCAAGT | ||
| MMP-2 | Forward | GTCCCTACCGAGTCTCTTCT | |
| Reverse | TTTTTAAGTTTCCGCTTCTG | ||
| MMP-9 | Forward | GCCACTTGTCGGCGATAAGG | |
| Reverse | CACTGTCCACCCCTCAGAGC | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahmawati, L.; Park, S.H.; Kim, D.S.; Lee, H.P.; Aziz, N.; Lee, C.Y.; Kim, S.A.; Jang, S.G.; Kim, D.S.; Cho, J.Y. Anti-Inflammatory Activities of the Ethanol Extract of Prasiola japonica, an Edible Freshwater Green Algae, and Its Various Solvent Fractions in LPS-Induced Macrophages and Carrageenan-Induced Paw Edema via the AP-1 Pathway. Molecules 2022, 27, 194. https://doi.org/10.3390/molecules27010194
Rahmawati L, Park SH, Kim DS, Lee HP, Aziz N, Lee CY, Kim SA, Jang SG, Kim DS, Cho JY. Anti-Inflammatory Activities of the Ethanol Extract of Prasiola japonica, an Edible Freshwater Green Algae, and Its Various Solvent Fractions in LPS-Induced Macrophages and Carrageenan-Induced Paw Edema via the AP-1 Pathway. Molecules. 2022; 27(1):194. https://doi.org/10.3390/molecules27010194
Chicago/Turabian StyleRahmawati, Laily, Sang Hee Park, Dong Seon Kim, Hwa Pyoung Lee, Nur Aziz, Chae Young Lee, Seung A Kim, Seok Gu Jang, Dong Sam Kim, and Jae Youl Cho. 2022. "Anti-Inflammatory Activities of the Ethanol Extract of Prasiola japonica, an Edible Freshwater Green Algae, and Its Various Solvent Fractions in LPS-Induced Macrophages and Carrageenan-Induced Paw Edema via the AP-1 Pathway" Molecules 27, no. 1: 194. https://doi.org/10.3390/molecules27010194
APA StyleRahmawati, L., Park, S. H., Kim, D. S., Lee, H. P., Aziz, N., Lee, C. Y., Kim, S. A., Jang, S. G., Kim, D. S., & Cho, J. Y. (2022). Anti-Inflammatory Activities of the Ethanol Extract of Prasiola japonica, an Edible Freshwater Green Algae, and Its Various Solvent Fractions in LPS-Induced Macrophages and Carrageenan-Induced Paw Edema via the AP-1 Pathway. Molecules, 27(1), 194. https://doi.org/10.3390/molecules27010194