
A Review of Twenty Years of Research on the Regulation of Signaling Pathways by Natural Products in Breast Cancer

 ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
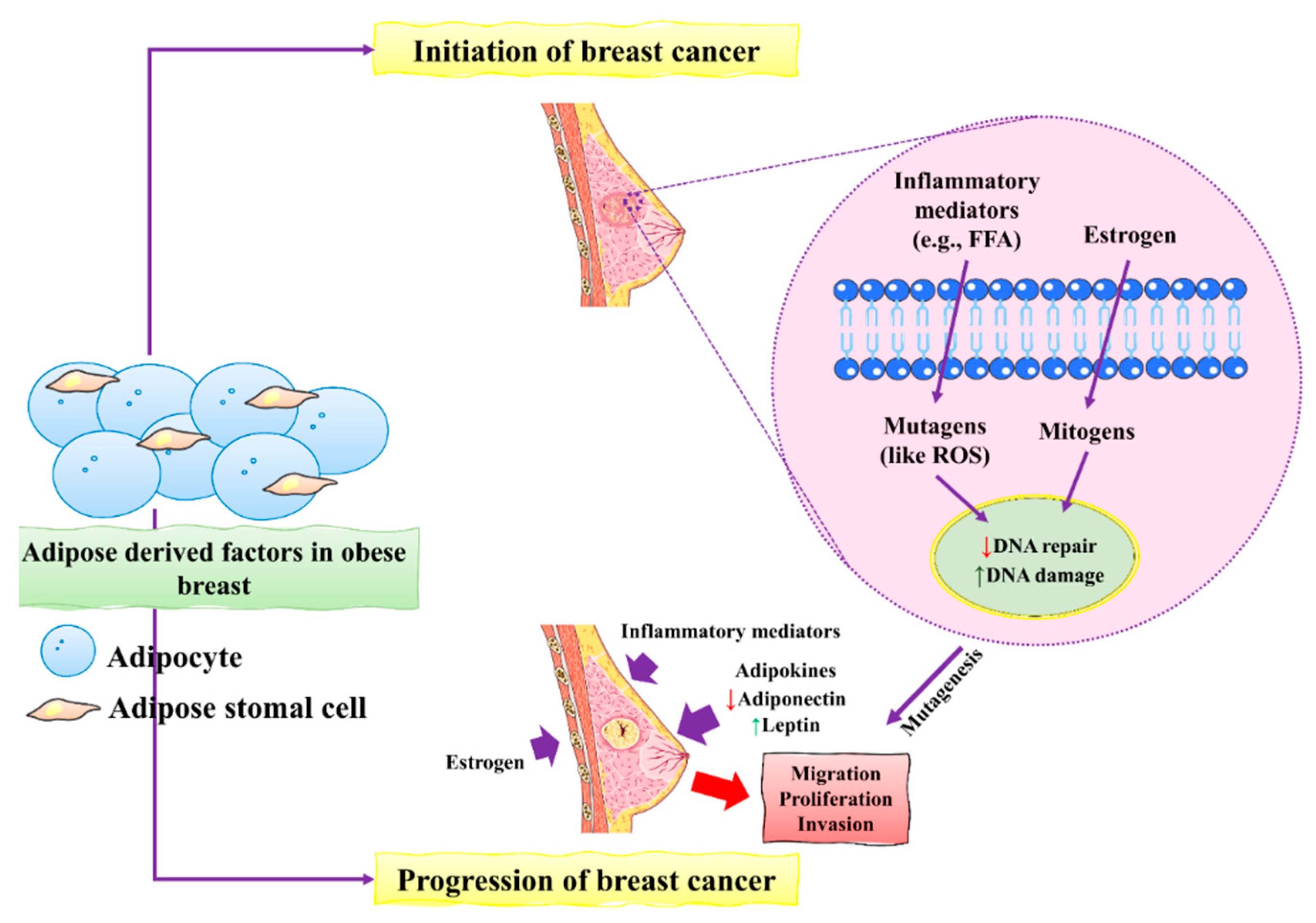
2. Pathogenesis of BC
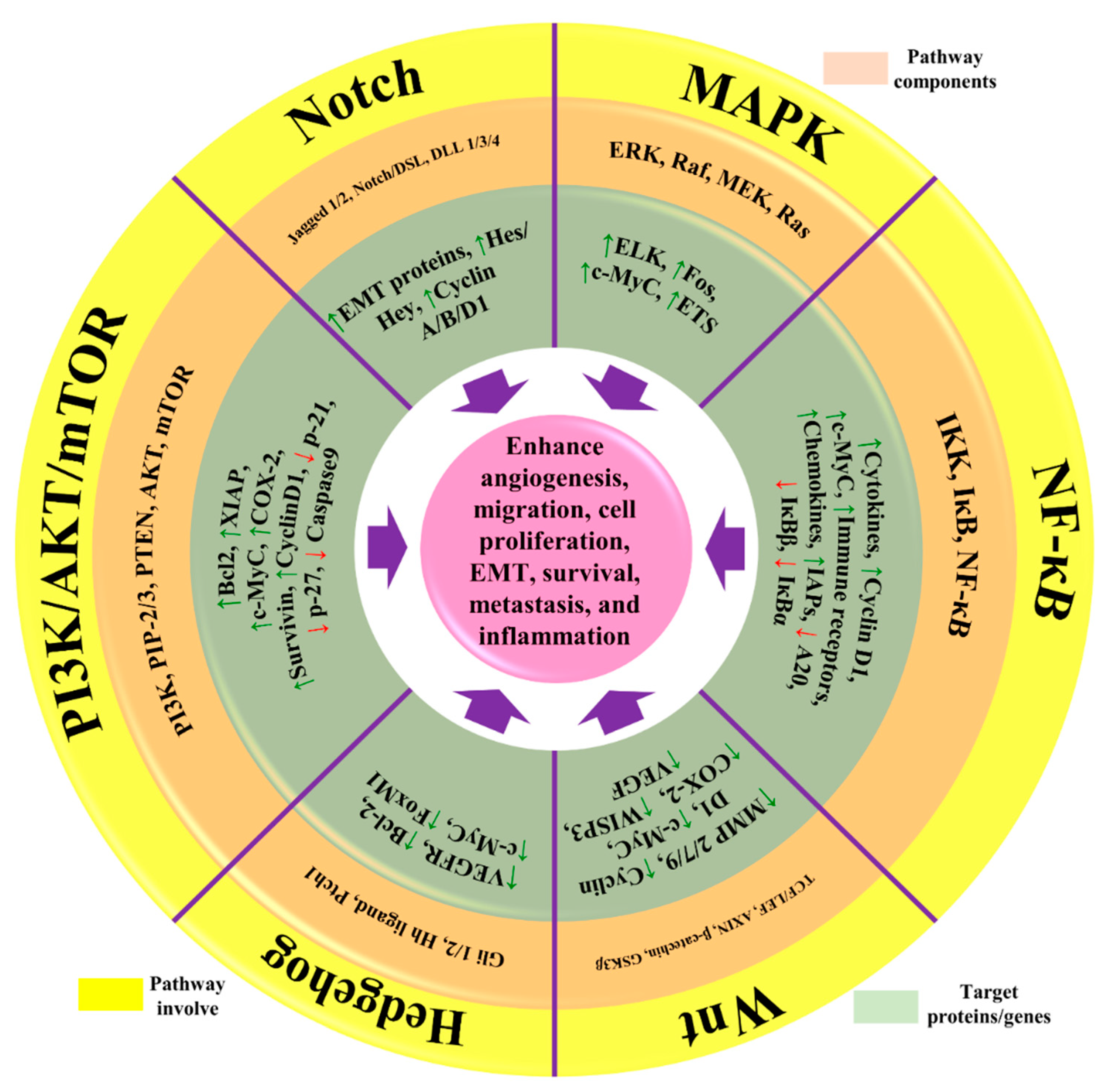
3. Downstream Regulation of Signaling Pathways by Natural Products
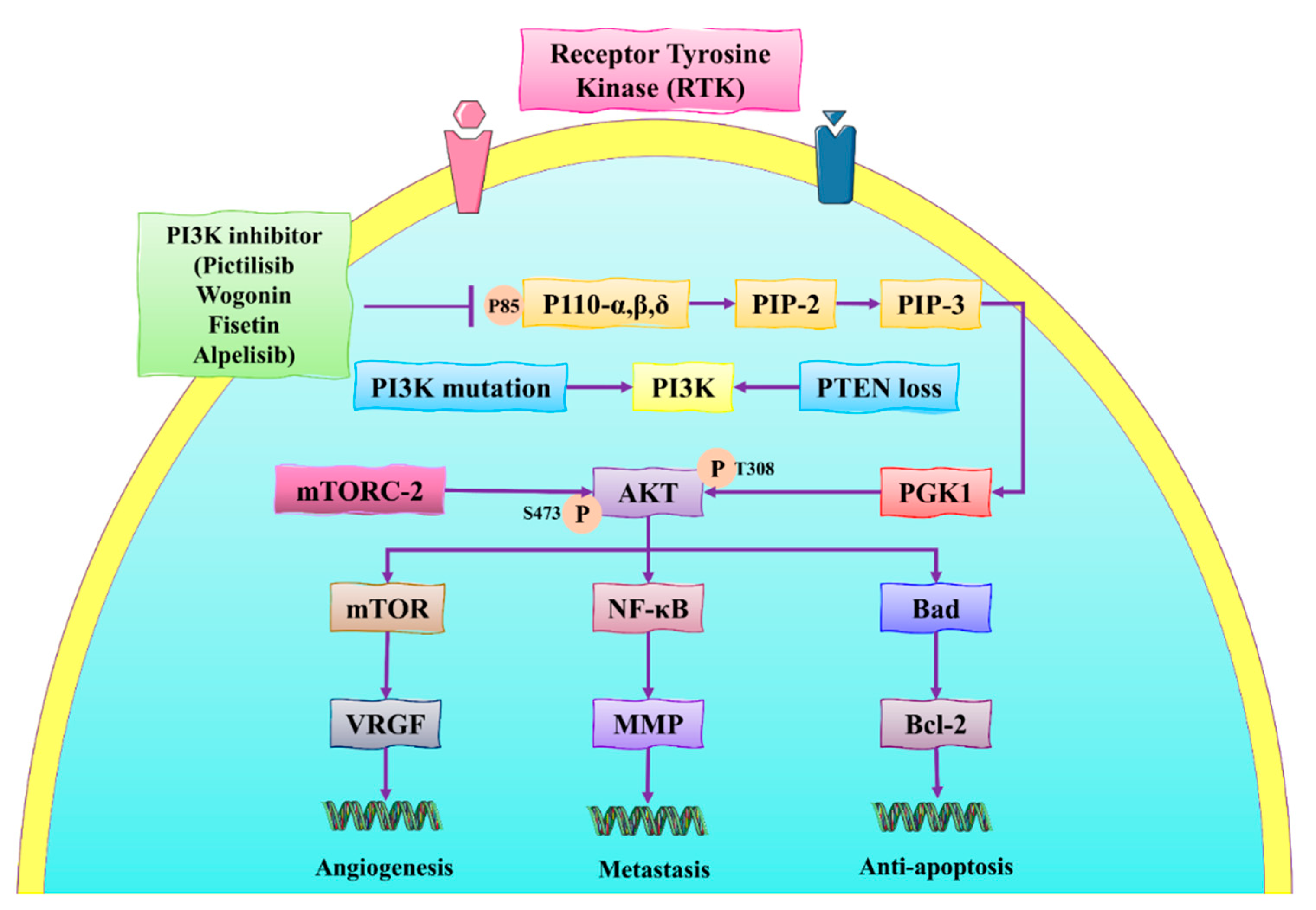
3.1. PI3K/Akt/mTOR Signaling Pathway
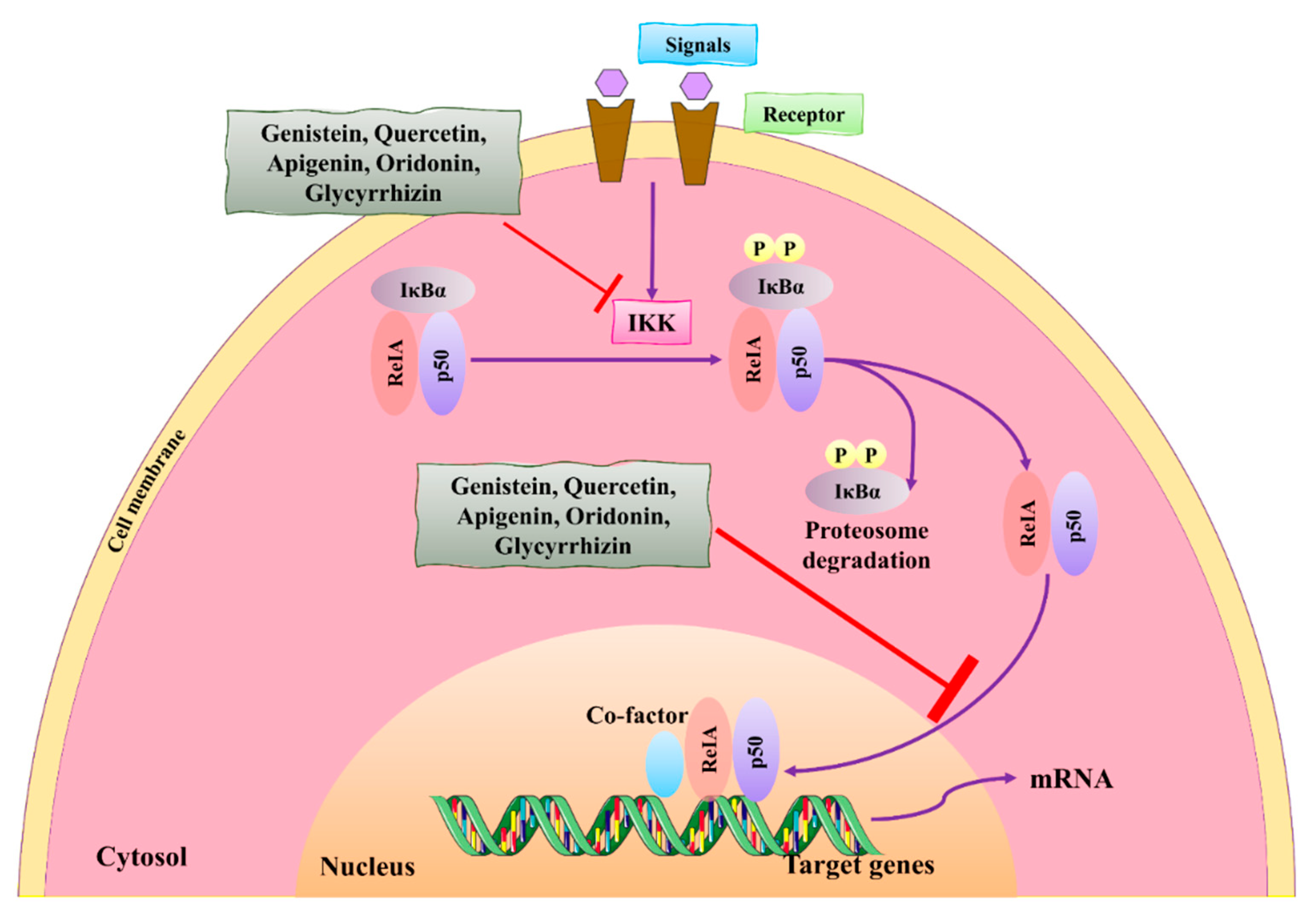
3.2. NF-κB Signaling Pathway
3.3. MAPK/ERK Signaling Pathway
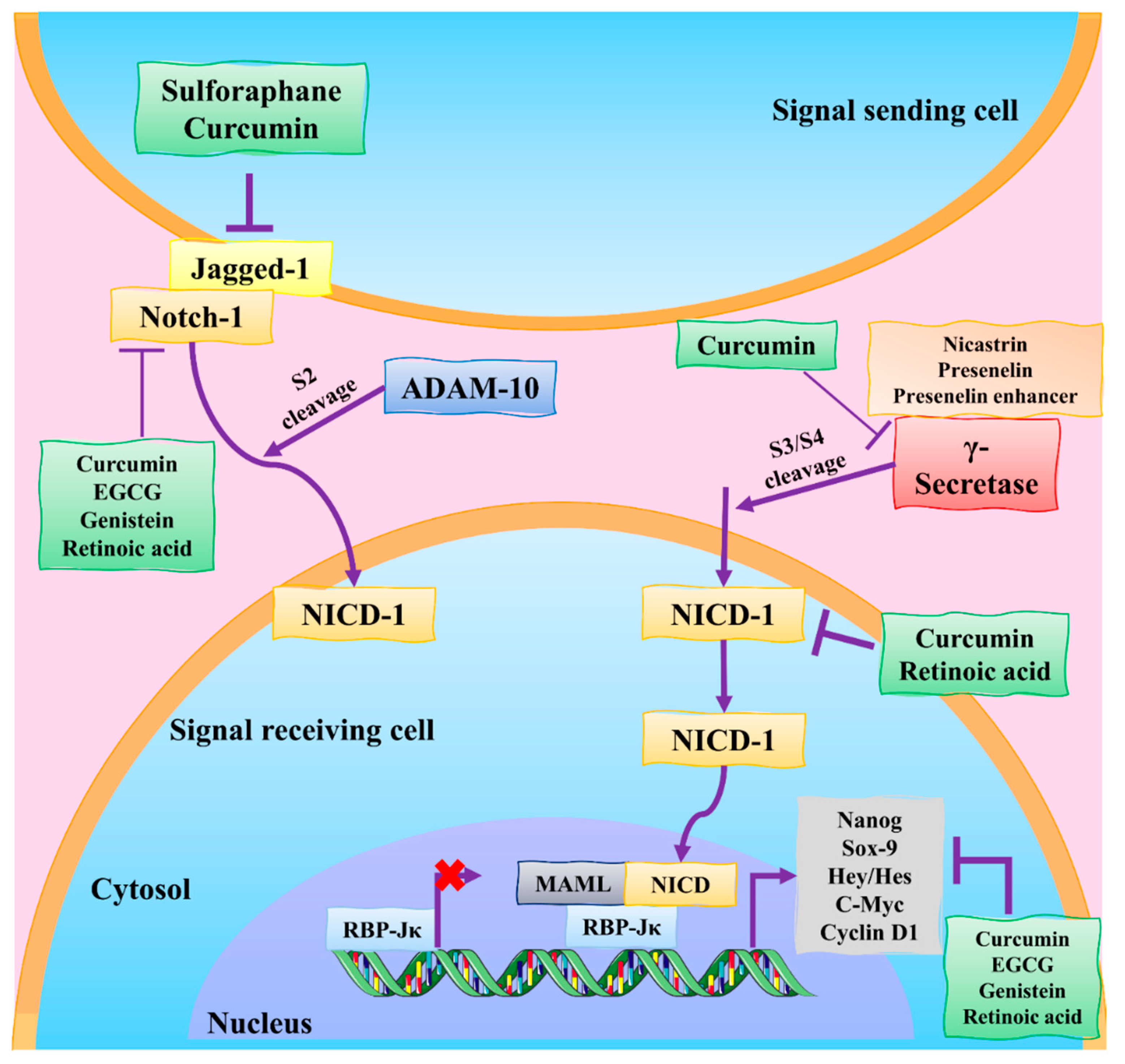
3.4. Notch Signaling Pathway
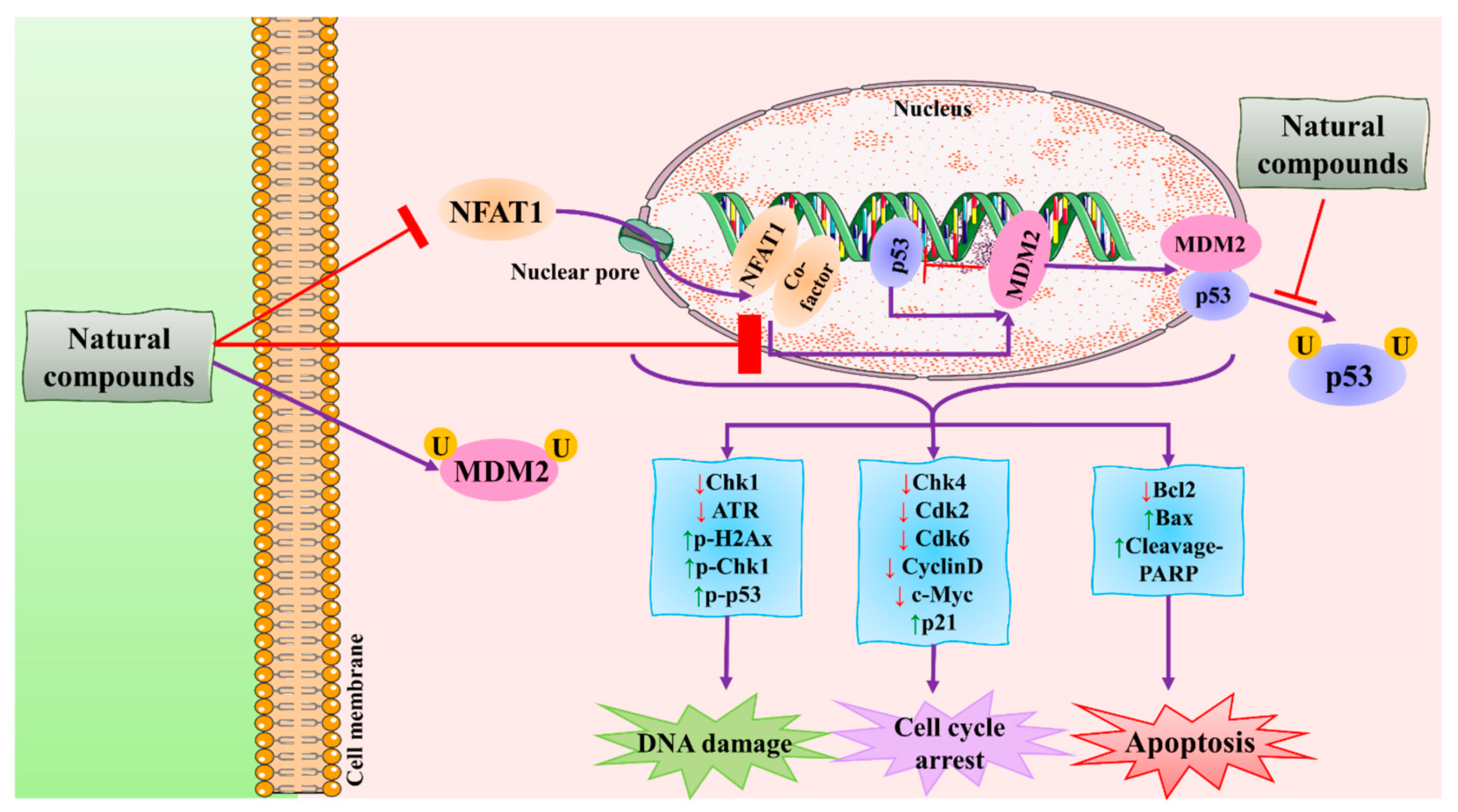
3.5. MDM2 Signaling Pathway
4. Synergistic Approach of Natural Products against BC
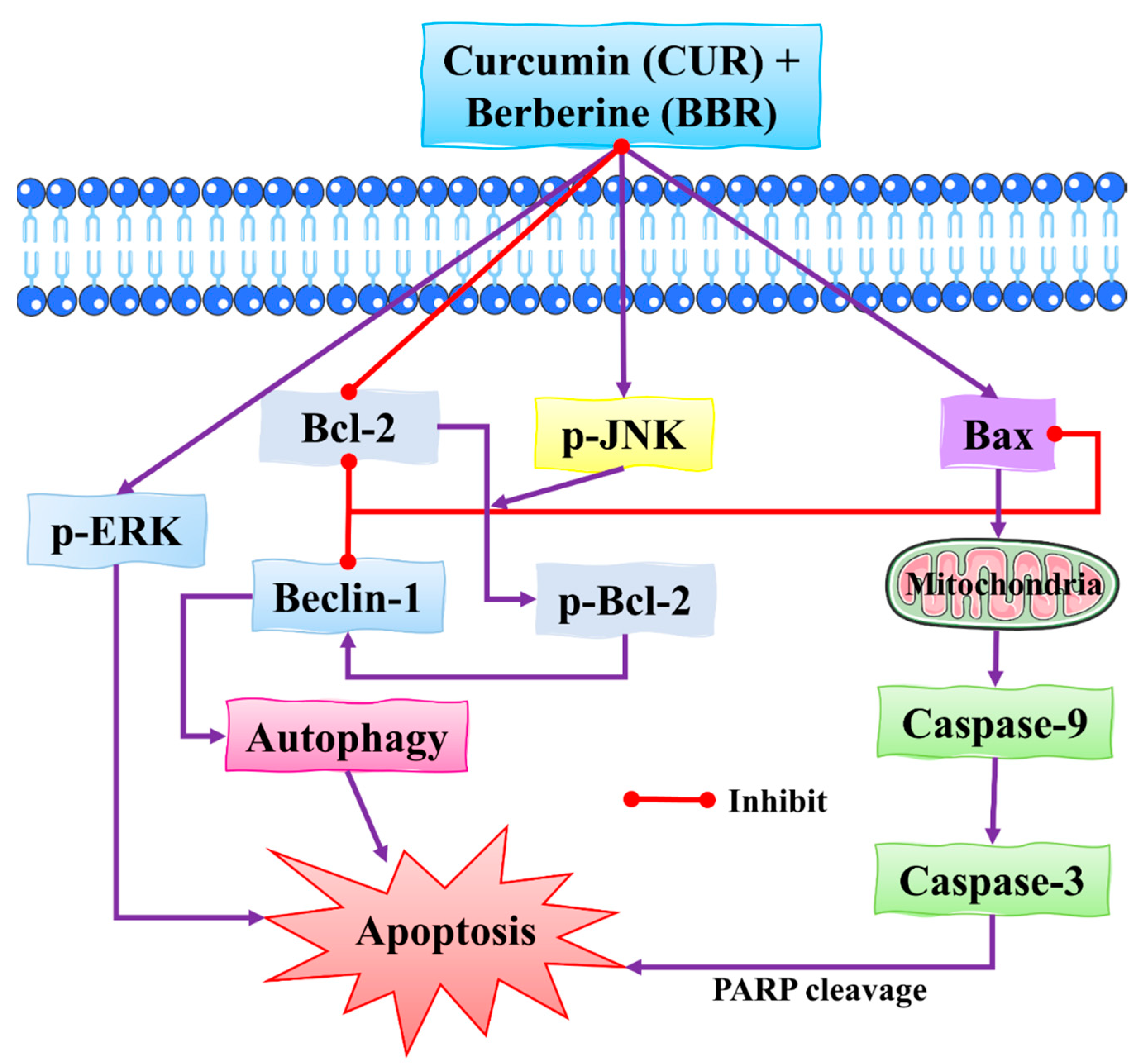
4.1. Synergistic Effects of Curcumin and Berberine
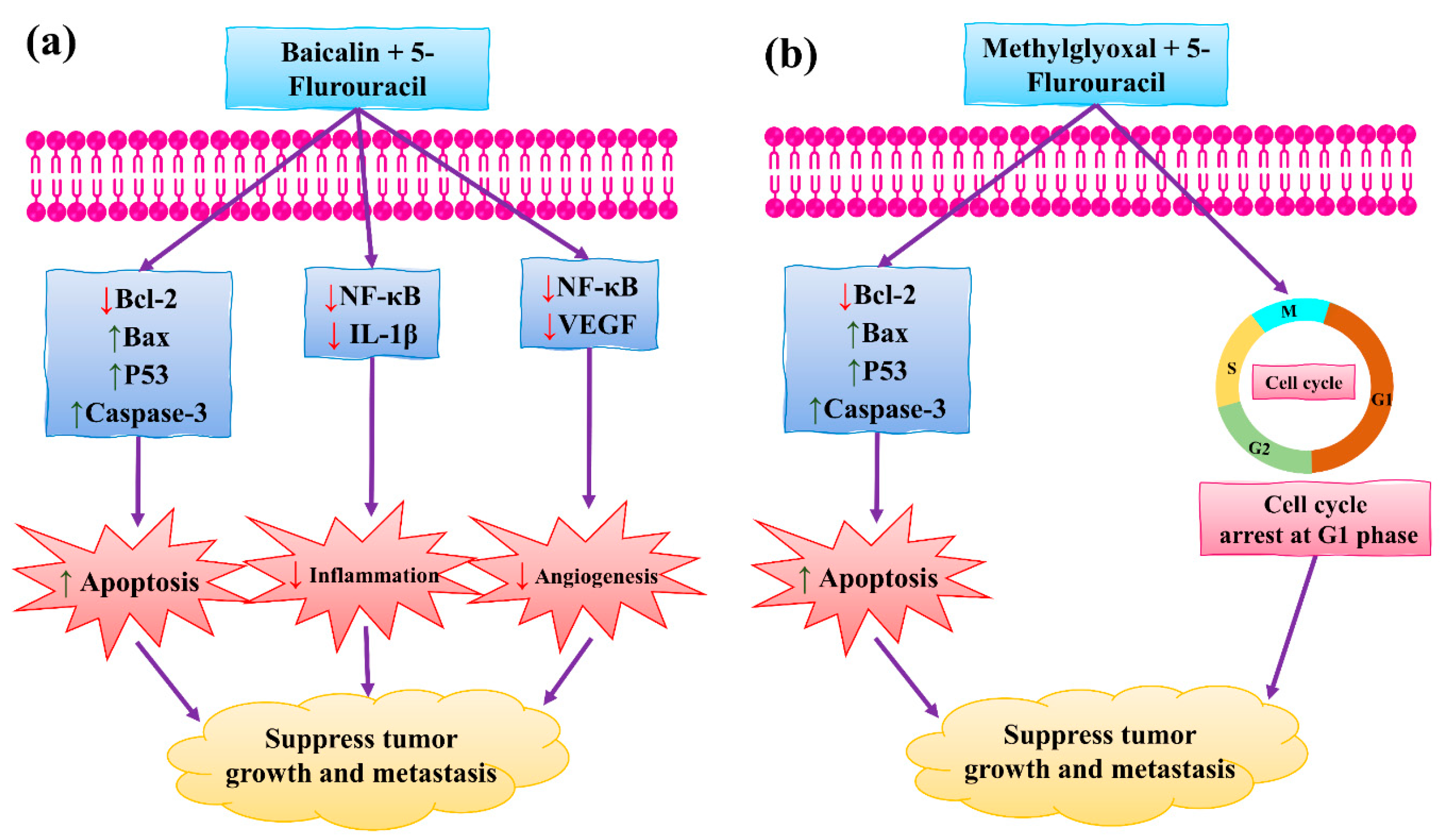
4.2. Synergistic Effects of Baicalin and Methylglyoxal with 5-Fluorouracil
4.3. Synergistic Effects of Resveratrol and Salinomycin
5. Recent Discoveries and Developments of Natural Products against BC
5.1. Novel Verminoside and Their Derivatives
5.2. Novel Phloroglucinol and Derivatives
5.3. Role of Viridiflorol against BC
5.4. Effect of Vulpinic Acid against BC
5.5. Action of Genistein against BC
6. Clinical Trials
7. Current Challenges and Future Perspectives
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Abbreviation | Explanation |
| ADAM-10 | A disintegrin and metalloproteinase domain-containing protein 10 |
| ADR | Adverse drug reactions |
| Akt | Protein kinase B |
| AP | Apigenin |
| BA | Baicalin |
| Bad | Bcl-2 antagonist of cell death |
| Bax | Bcl-2-associated X protein |
| BBR | Berberine; BC: breast cancer |
| Bcl-2 | B-cell lymphoma 2 |
| CDK | Cyclin-dependent kinase |
| Chk | Checkpoint kinase |
| CUR | Curcumin |
| DATS | Diallyl trisulfide |
| EGCG | Epigallocatechin-3-gallate |
| EGF(R) | Epidermal growth factor (receptor) |
| ER | Estrogen receptor |
| ERK | Extracellular signal-regulated kinase |
| 5-FU | 5-Fluorouracil |
| HCT | Hematocrit |
| IKK | IκB kinase |
| IL-1β | Interleukin 1 beta |
| IDS | Increase in drug sensitivity |
| JNK | C-Jun N-terminal kinase |
| Lin A | Lineariifolianoid A |
| MAML | Mastermind-like protein |
| MDM2 | Mouse double minute 2 homolog |
| MEK | Mitogen-activated protein kinase kinase |
| MCF-7 | Michigan Cancer Foundation 7 |
| MDR1 | Multidrug resistance 1 |
| MG | Methylglyoxal |
| MMP | Matrix metalloprotein |
| mTOR | Mammalian target of rapamycin |
| mTORC-2 | mTOR Complex 2 |
| MRP1 | Multidrug resistance protein 1 |
| NFAT1 | Nuclear factor of activated T cells 1 |
| NF-κB | Nuclear factor kappa B |
| NICD | Notch receptor intracellular domain |
| p53 | Inducible gene 3 |
| PARP | Poly (ADP-ribose) polymerase |
| PDK1/2 | 3-phosphoinositide dependent kinase-1/2 |
| PI3K | Phosphatidylinositol 3-kinase |
| PIP-3 | Phosphatidylinositol (3,4,5)-trisphosphate |
| PPD | Protopanaxadiol |
| PTEN | Phosphatase and tensin homolog |
| RBP-Jκ | Recombinant signal binding protein for Immunoglobulin kappa J region |
| RSVL | Resveratrol |
| SAL | Salinomycin |
| TNBC | Triple-negative breast cancer |
| VEGF | Vascular endothelial growth factor |
| VOA | Voacamine |
| VS | Verminoside |
| WG | Wogonin |
References
- Jha, V.; Devkar, S.; Gharat, K.; Kasbe, S.; Matharoo, D.K.; Pendse, S.; Bhosale, A.; Bhargava, A. Screening of Phytochemicals as Potential Inhibitors of Breast Cancer using Structure Based Multitargeted Molecular Docking Analysis. Phytomed. Plus 2022, 2, 100227. [Google Scholar] [CrossRef]
- Nassif, A.B.; Talib, M.A.; Nasir, Q.; Afadar, Y.; Elgendy, O. Breast cancer detection using artificial intelligence techniques: A systematic literature review. Artif. Intell. Med. 2022, 127, 102276. [Google Scholar] [CrossRef] [PubMed]
- Rieder, V.; Salama, M.; Glöckner, L.; Muhr, D.; Berger, A.; Tea, M.K.; Pfeiler, G.; Rappaport-Fuerhauser, C.; Gschwantler-Kaulich, D.; Weingartshofer, S. Effect of lifestyle and reproductive factors on the onset of breast cancer in female BRCA 1 and 2 mutation carriers. Mol. Genet. Genom. Med. 2016, 4, 172–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sisti, J.S.; Collins, L.C.; Beck, A.H.; Tamimi, R.M.; Rosner, B.A.; Eliassen, A.H. Reproductive risk factors in relation to molecular subtypes of breast cancer: Results from the nurses’ health studies. Int. J. Cancer 2016, 138, 2346–2356. [Google Scholar] [CrossRef]
- Ma, H.; Ursin, G.; Xu, X.; Lee, E.; Togawa, K.; Duan, L.; Lu, Y.; Malone, K.E.; Marchbanks, P.A.; McDonald, J.A. Reproductive factors and the risk of triple-negative breast cancer in white women and African-American women: A pooled analysis. Breast Cancer Res. 2017, 19, 6. [Google Scholar] [CrossRef] [Green Version]
- Campeau, P.M.; Foulkes, W.D.; Tischkowitz, M.D. Hereditary breast cancer: New genetic developments, new therapeutic avenues. Hum. Genet. 2008, 124, 31–42. [Google Scholar] [CrossRef]
- Moulder, S.; Hortobagyi, G. Advances in the treatment of breast cancer. Clin. Pharmacol. Ther. 2008, 83, 26–36. [Google Scholar] [CrossRef]
- Cazzaniga, M.; Bonanni, B. Breast cancer chemoprevention: Old and new approaches. J. Biomed. Biotechnol. 2012, 2012, 985620. [Google Scholar] [CrossRef]
- Mocanu, M.-M.; Nagy, P.; Szöllősi, J. Chemoprevention of breast cancer by dietary polyphenols. Molecules 2015, 20, 22578–22620. [Google Scholar] [CrossRef] [Green Version]
- Abotaleb, M.; Kubatka, P.; Caprnda, M.; Varghese, E.; Zolakova, B.; Zubor, P.; Opatrilova, R.; Kruzliak, P.; Stefanicka, P.; Büsselberg, D. Chemotherapeutic agents for the treatment of metastatic breast cancer: An update. Biomed. Pharmacother. 2018, 101, 458–477. [Google Scholar] [CrossRef]
- Akram, M.; Iqbal, M.; Daniyal, M.; Khan, A.U. Awareness and current knowledge of breast cancer. Biol. Res. 2017, 50, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeSantis, C.; Ma, J.; Bryan, L.; Jemal, A. Breast cancer statistics, 2013. CA Cancer J. Clin. 2014, 64, 52–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Wu, Y.; Cong, W.; Hu, M.; Li, X.; Zhou, C. Experience of women with breast cancer undergoing chemotherapy: A systematic review of qualitative research. Qual. Life Res. 2021, 30, 1249–1265. [Google Scholar] [CrossRef] [PubMed]
- Sartaj, A.; Baboota, S.; Ali, J. Assessment of Combination Approaches of Phytoconstituents with Chemotherapy for the Treatment of Breast Cancer: A Systematic Review. Curr. Pharm. Des. 2021, 27, 4630–4648. [Google Scholar] [CrossRef]
- Yap, K.M.; Sekar, M.; Fuloria, S.; Wu, Y.S.; Gan, S.H.; Rani, N.N.I.M.; Subramaniyan, V.; Kokare, C.; Lum, P.T.; Begum, M.Y. Drug delivery of natural products through nanocarriers for effective breast cancer therapy: A comprehensive review of literature. Int. J. Nanomed. 2021, 16, 7891–7941. [Google Scholar] [CrossRef]
- He, Y.; Tao, Q.; Zhou, F.; Si, Y.; Fu, R.; Xu, B.; Xu, J.; Li, X.; Chen, B. The relationship between dairy products intake and breast cancer incidence: A meta-analysis of observational studies. BMC Cancer 2021, 21, 1109. [Google Scholar] [CrossRef]
- Farvid, M.S.; Chen, W.Y.; Michels, K.B.; Cho, E.; Willett, W.C.; Eliassen, A.H. Fruit and vegetable consumption in adolescence and early adulthood and risk of breast cancer: Population based cohort study. BMJ 2016, 353, i2343. [Google Scholar] [CrossRef] [Green Version]
- Nechuta, S.J.; Caan, B.J.; Chen, W.Y.; Lu, W.; Chen, Z.; Kwan, M.L.; Flatt, S.W.; Zheng, Y.; Zheng, W.; Pierce, J.P. Soy food intake after diagnosis of breast cancer and survival: An in-depth analysis of combined evidence from cohort studies of US and Chinese women. Am. J. Clin. Nutr. 2012, 96, 123–132. [Google Scholar] [CrossRef] [Green Version]
- Lv, Z.-D.; Liu, X.-P.; Zhao, W.-J.; Dong, Q.; Li, F.-N.; Wang, H.-B.; Kong, B. Curcumin induces apoptosis in breast cancer cells and inhibits tumor growth in vitro and in vivo. Int. J. Clin. Exp. Pathol. 2014, 7, 2818–2824. [Google Scholar]
- Bonofiglio, D.; Giordano, C.; De Amicis, F.; Lanzino, M.; Ando, S. Natural Products as Promising Antitumoral Agents in Breast Cancer: Mechanisms of Action and Molecular Targets. Mini Rev. Med. Chem. 2016, 16, 596–604. [Google Scholar] [CrossRef]
- Imran, M.; Aslam Gondal, T.; Atif, M.; Shahbaz, M.; Batool Qaisarani, T.; Hanif Mughal, M.; Salehi, B.; Martorell, M.; Sharifi-Rad, J. Apigenin as an anticancer agent. Phytother. Res. 2020, 34, 1812–1828. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Saeed, F.; Gilani, S.A.; Shariati, M.A.; Imran, A.; Afzaal, M.; Atif, M.; Tufail, T.; Anjum, F.M. Fisetin: An anticancer perspective. Food Sci. Nutr. 2021, 9, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Liu, M.; Wang, X.; Jiang, Q.; Wang, S.; Santhanam, R.K.; Lv, C.; Zhao, Q.; Lu, J. Cimigenoside functions as a novel γ-secretase inhibitor and inhibits the proliferation or metastasis of human breast cancer cells by γ-secretase/Notch axis. Pharmacol. Res. 2021, 169, 105686. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.-J.; Kuo, P.-L.; Hsu, Y.-C.; Huang, Y.-F.; Tsai, E.-M.; Hsu, Y.-L. Arctigenin, a dietary phytoestrogen, induces apoptosis of estrogen receptor-negative breast cancer cells through the ROS/p38 MAPK pathway and epigenetic regulation. Free Radic. Biol. Med. 2014, 67, 159–170. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Wang, X.; Ma, Q.; Zhao, W.; Jia, Y.; Dong, G.; Zhu, Y.; Jia, X.; Tong, Z. Glycyrrhizin inhibits the invasion and metastasis of breast cancer cells via upregulation of expressions of miR-200c and e-cadherin. Trop. J. Pharm. Res. 2020, 19, 1807–1813. [Google Scholar] [CrossRef]
- Chung, H.; Choi, H.S.; Seo, E.-K.; Kang, D.-H.; Oh, E.-S. Baicalin and baicalein inhibit transforming growth factor-β1-mediated epithelial-mesenchymal transition in human breast epithelial cells. Biochem. Biophys. Res. Commun. 2015, 458, 707–713. [Google Scholar] [CrossRef]
- Akiel, M.A.; Alshehri, O.Y.; Aljihani, S.A.; Almuaysib, A.; Bader, A.; Al-Asmari, A.I.; Alamri, H.S.; Alrfaei, B.M.; Halwani, M.A. Viridiflorol induces anti-neoplastic effects on breast, lung, and brain cancer cells through apoptosis. Saudi J. Biol. Sci. 2022, 29, 816–821. [Google Scholar] [CrossRef]
- Cansaran-Duman, D.; Yangin, S.; Çolak, B. The role of vulpinic acid as a natural compound in the regulation of breast cancer-associated miRNAs. Biol. Res. 2021, 54, 37. [Google Scholar] [CrossRef]
- El Gaafary, M.; Saber, F.R.; Mahrous, E.A.; Ashour, R.M.; Okba, M.M.; Jin, L.; Lang, S.J.; Schmiech, M.; Simmet, T.; Syrovets, T. The phloroglucinol calcitrinone A, a novel mitochondria-targeting agent, induces cell death in breast cancer cells. Food Chem. Toxicol. 2022, 162, 112896. [Google Scholar] [CrossRef]
- Ghosh, S.; Pal, A.; Ray, M. Methylglyoxal in combination with 5-Fluorouracil elicits improved chemosensitivity in breast cancer through apoptosis and cell cycle inhibition. Biomed. Pharmacother. 2019, 114, 108855. [Google Scholar] [CrossRef]
- Schuster, C.; Wolpert, N.; Moustaid-Moussa, N.; Gollahon, L.S. Combinatorial Effects of the Natural Products Arctigenin, Chlorogenic Acid, and Cinnamaldehyde Commit Oxidation Assassination on Breast Cancer Cells. Antioxidants 2022, 11, 591. [Google Scholar] [CrossRef] [PubMed]
- Andrijauskaite, K.; Wargovich, M.J. Role of natural products in breast cancer related symptomology: Targeting chronic inflammation. Semin. Cancer Biol. 2020, 380, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-H.; Hwang, K.-A.; Choi, K.-C. Treatment with kaempferol suppresses breast cancer cell growth caused by estrogen and triclosan in cellular and xenograft breast cancer models. J. Nutr. Biochem. 2016, 28, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Y.; Zhang, C.-Z.; Ren, Q.; Chen, Y.; Li, X.; Yang, J.-R.; Li, H.-X.; Tang, W.-T.; Hing-Man, H.; Sun, C. Activation of mitochondrial-associated apoptosis signaling pathway and inhibition of PI3K/Akt/mTOR signaling pathway by voacamine suppress breast cancer progression. Phytomedicine 2022, 99, 154015. [Google Scholar] [CrossRef]
- Son, Y.; Cheong, Y.-K.; Kim, N.-H.; Chung, H.-T.; Kang, D.G.; Pae, H.-O. Mitogen-activated protein kinases and reactive oxygen species: How can ROS activate MAPK pathways? J. Signal Transduct. 2011, 2011, 792639. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, X.; Qin, J.-J.; Voruganti, S.; Nag, S.A.; Wang, M.-H.; Wang, H.; Zhang, R. Natural product ginsenoside 25-OCH3-PPD inhibits breast cancer growth and metastasis through down-regulating MDM2. PLoS ONE 2012, 7, e41586. [Google Scholar] [CrossRef]
- Feng, Y.; Spezia, M.; Huang, S.; Yuan, C.; Zeng, Z.; Zhang, L.; Ji, X.; Liu, W.; Huang, B.; Luo, W. Breast cancer development and progression: Risk factors, cancer stem cells, signaling pathways, genomics, and molecular pathogenesis. Genes Dis. 2018, 5, 77–106. [Google Scholar] [CrossRef]
- Reis-Filho, J.S.; Pusztai, L. Gene expression profiling in breast cancer: Classification, prognostication, and prediction. Lancet 2011, 378, 1812–1823. [Google Scholar] [CrossRef]
- Hudis, C.A.; Gianni, L. Triple-negative breast cancer: An unmet medical need. Oncologist 2011, 16, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Bhardwaj, P.; Brown, K.A. Obese adipose tissue as a driver of breast cancer growth and development: Update and emerging evidence. Front. Oncol. 2021, 11, 638918. [Google Scholar] [CrossRef]
- Evans, D.G.; van Veen, E.M.; Byers, H.J.; Evans, S.J.; Burghel, G.J.; Woodward, E.R.; Harkness, E.F.; Eccles, D.M.; Greville-Haygate, S.L.; Ellingford, J.M. High likelihood of actionable pathogenic variant detection in breast cancer genes in women with very early onset breast cancer. J. Med. Genet. 2022, 59, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Thomas, G.; Ghoussaini, M.; Healey, C.S.; Humphreys, M.K.; Platte, R.; Morrison, J.; Maranian, M.; Pooley, K.A.; Luben, R. Newly discovered breast cancer susceptibility loci on 3p24 and 17q23. 2. Nat. Genet. 2009, 41, 585–590. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Kumar, K.; Purohit, D.; Verma, R.; Pandey, P.; Bhatia, S.; Malik, V.; Mittal, V.; Rahman, M.H.; Albadrani, G.M. Exploration of therapeutic applicability and different signaling mechanism of various phytopharmacological agents for treatment of breast cancer. Biomed. Pharmacother. 2021, 139, 111584. [Google Scholar] [CrossRef] [PubMed]
- Varghese, E.; Samuel, S.M.; Abotaleb, M.; Cheema, S.; Mamtani, R.; Büsselberg, D. The “yin and yang” of natural compounds in anticancer therapy of triple-negative breast cancers. Cancers 2018, 10, 346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marino, N.; German, R.; Rao, X.; Simpson, E.; Liu, S.; Wan, J.; Liu, Y.; Sandusky, G.; Jacobsen, M.; Stoval, M. Upregulation of lipid metabolism genes in the breast prior to cancer diagnosis. NPJ Breast Cancer 2020, 6, 50. [Google Scholar] [CrossRef] [PubMed]
- Colotta, F.; Allavena, P.; Sica, A.; Garlanda, C.; Mantovani, A. Cancer-related inflammation, the seventh hallmark of cancer: Links to genetic instability. Carcinogenesis 2009, 30, 1073–1081. [Google Scholar] [CrossRef] [Green Version]
- Suman, S.; Sharma, P.K.; Rai, G.; Mishra, S.; Arora, D.; Gupta, P.; Shukla, Y. Current perspectives of molecular pathways involved in chronic inflammation-mediated breast cancer. Biochem. Biophys. Res. Commun. 2016, 472, 401–409. [Google Scholar] [CrossRef]
- Huang, A.; Cao, S.; Tang, L. The tumor microenvironment and inflammatory breast cancer. J. Cancer 2017, 8, 1884. [Google Scholar] [CrossRef]
- Puar, Y.R.; Shanmugam, M.K.; Fan, L.; Arfuso, F.; Sethi, G.; Tergaonkar, V. Evidence for the involvement of the master transcription factor NF-κB in cancer initiation and progression. Biomedicines 2018, 6, 82. [Google Scholar] [CrossRef] [Green Version]
- Oshi, M.; Newman, S.; Tokumaru, Y.; Yan, L.; Matsuyama, R.; Endo, I.; Nagahashi, M.; Takabe, K. Intra-tumoral angiogenesis is associated with inflammation, immune reaction and metastatic recurrence in breast cancer. Int. J. Mol. Sci. 2020, 21, 6708. [Google Scholar] [CrossRef]
- Michels, N.; van Aart, C.; Morisse, J.; Mullee, A.; Huybrechts, I. Chronic inflammation towards cancer incidence: A systematic review and meta-analysis of epidemiological studies. Crit. Rev. Oncol. Hematol. 2021, 157, 103177. [Google Scholar] [CrossRef] [PubMed]
- Alzahrani, A.S. PI3K/Akt/mTOR inhibitors in cancer: At the bench and bedside. Semin. Cancer Biol. 2019, 59, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Verret, B.; Cortes, J.; Bachelot, T.; Andre, F.; Arnedos, M. Efficacy of PI3K inhibitors in advanced breast cancer. Ann. Oncol. 2019, 30, x12–x20. [Google Scholar] [CrossRef] [PubMed]
- Dandoti, S. Mechanisms adopted by cancer cells to escape apoptosis–A review. Biocell 2021, 45, 863–884. [Google Scholar] [CrossRef]
- Porta, C.; Paglino, C.; Mosca, A. Targeting PI3K/Akt/mTOR signaling in cancer. Front. Oncol. 2014, 4, 64. [Google Scholar] [CrossRef] [Green Version]
- Dong, C.; Wu, J.; Chen, Y.; Nie, J.; Chen, C. Activation of PI3K/AKT/mTOR pathway causes drug resistance in breast cancer. Front. Pharmacol. 2021, 12, 628690. [Google Scholar] [CrossRef]
- Dornan, G.L.; Burke, J.E. Molecular mechanisms of human disease mediated by oncogenic and primary immunodeficiency mutations in class IA phosphoinositide 3-kinases. Front. Immunol. 2018, 9, 575. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Meng, T.; Zheng, X.; Liu, Y.; Hao, R.; Yan, Y.; Chen, S.; You, H.; Xing, J.; Dong, Y. Transgelin 2 promotes paclitaxel resistance, migration, and invasion of breast cancer by directly interacting with PTEN and activating PI3K/Akt/GSK-3β pathway. Mol. Cancer Ther. 2019, 18, 2457–2468. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Cheng, H.; Roberts, T.M.; Zhao, J.J. Targeting the phosphoinositide 3-kinase pathway in cancer. Nat. Rev. Drug Discov. 2009, 8, 627–644. [Google Scholar] [CrossRef] [Green Version]
- Cidado, J.; Park, B.H. Targeting the PI3K/Akt/mTOR pathway for breast cancer therapy. J. Mammary Gland Biol. Neoplasia 2012, 17, 205–216. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.; Fan, D. Ginsenoside Rk1 induces cell cycle arrest and apoptosis in MDA-MB-231 triple negative breast cancer cells. Toxicology 2019, 418, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Fan, D. Ginsenoside Rg5 induces apoptosis and autophagy via the inhibition of the PI3K/Akt pathway against breast cancer in a mouse model. Food Funct. 2018, 9, 5513–5527. [Google Scholar] [CrossRef] [PubMed]
- Bing, Z.; Shi-ke, Y.; Chun-ling, B.; Ning, L. Research progress of natural products targeting tumor angiogenesis. Nat. Prod. Res. Dev. 2021, 33, 510–520. [Google Scholar]
- Condello, M.; Pellegrini, E.; Multari, G.; Gallo, F.R.; Meschini, S. Voacamine: Alkaloid with its essential dimeric units to reverse tumor multidrug resistance. Toxicol. Vitr. 2020, 65, 104819. [Google Scholar] [CrossRef] [PubMed]
- Grynkiewicz, G.; Demchuk, O.M. New perspectives for fisetin. Front. Chem. 2019, 7, 697. [Google Scholar] [CrossRef]
- Sun, X.; Ma, X.; Li, Q.; Yang, Y.; Xu, X.; Sun, J.; Yu, M.; Cao, K.; Yang, L.; Yang, G. Anti-cancer effects of fisetin on mammary carcinoma cells via regulation of the PI3K/Akt/mTOR pathway: In vitro and in vivo studies. Int. J. Mol. Med. 2018, 42, 811–820. [Google Scholar] [CrossRef] [Green Version]
- Mirza-Aghazadeh-Attari, M.; Ekrami, E.M.; Aghdas, S.A.M.; Mihanfar, A.; Hallaj, S.; Yousefi, B.; Safa, A.; Majidinia, M. Targeting PI3K/Akt/mTOR signaling pathway by polyphenols: Implication for cancer therapy. Life Sci. 2020, 255, 117481. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Herrera-Bravo, J.; Salazar, L.A.; Shaheen, S.; Ayatollahi, S.A.; Kobarfard, F.; Imran, M.; Imran, A.; Custódio, L.; López, M.D. The therapeutic potential of wogonin observed in preclinical studies. Evid. Based Complement. Altern. Med. 2021, 2021, 9935451. [Google Scholar] [CrossRef]
- Gharari, Z.; Bagheri, K.; Khodaeiaminjan, M.; Sharafi, A. Potential therapeutic effects and bioavailability of wogonin, the flavone of Baikal skullcap. J. Nutri. Med. Diet Care 2019, 5, 039. [Google Scholar]
- Zhao, K.; Song, X.; Huang, Y.; Yao, J.; Zhou, M.; Li, Z.; You, Q.; Guo, Q.; Lu, N. Wogonin inhibits LPS-induced tumor angiogenesis via suppressing PI3K/Akt/NF-κB signaling. Eur. J. Pharmacol. 2014, 737, 57–69. [Google Scholar] [CrossRef]
- Ganesan, K.; Xu, B. Telomerase inhibitors from natural products and their anticancer potential. Int. J. Mol. Sci. 2017, 19, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganesan, K.; Xu, B. Molecular targets of vitexin and isovitexin in cancer therapy: A critical review. Ann. N. Y. Acad. Sci. 2017, 1401, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Pons, D.G.; Vilanova-Llompart, J.; Gaya-Bover, A.; Alorda-Clara, M.; Oliver, J.; Roca, P.; Sastre-Serra, J. The phytoestrogen genistein affects inflammatory-related genes expression depending on the ERα/ERβ ratio in breast cancer cells. Int. J. Food Sci. Nutr. 2019, 70, 941–949. [Google Scholar] [CrossRef]
- Seo, H.S.; Choi, H.S.; Choi, H.S.; Choi, Y.K.; Um, J.-Y.; Choi, I.; Shin, Y.C.; Ko, S.-G. Phytoestrogens induce apoptosis via extrinsic pathway, inhibiting nuclear factor-κB signaling in HER2-overexpressing breast cancer cells. Anticancer. Res. 2011, 31, 3301–3313. [Google Scholar]
- Pan, H.; Zhou, W.; He, W.; Liu, X.; Ding, Q.; Ling, L.; Zha, X.; Wang, S. Genistein inhibits MDA-MB-231 triple-negative breast cancer cell growth by inhibiting NF-κB activity via the Notch-1 pathway. Int. J. Mol. Med. 2012, 30, 337–343. [Google Scholar] [CrossRef] [Green Version]
- Ganesan, K.; Du, B.; Chen, J. Effects and mechanisms of dietary bioactive compounds on breast cancer prevention. Pharmacol. Res. 2022, 178, 105974. [Google Scholar] [CrossRef]
- Bauer, D.; Mazzio, E.; Soliman, K.F. Whole transcriptomic analysis of apigenin on TNFα immuno-activated MDA-MB-231 breast cancer cells. Cancer Genom. Proteom. 2019, 16, 421–431. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.-M.; Kim, D.-H.; Park, J.-H.; Surh, Y.-J.; Na, H.-K. Ginsenoside Rg3 inhibits constitutive activation of NF-κB signaling in human breast cancer (MDA-MB-231) cells: ERK and Akt as potential upstream targets. J. Cancer Prev. 2014, 19, 23–30. [Google Scholar] [CrossRef]
- Ozbay, T.; Nahta, R. Delphinidin inhibits HER2 and Erk1/2 signaling and suppresses growth of HER2-overexpressing and triple negative breast cancer cell lines. Breast Cancer Basic Clin. Res. 2011, 5, 143–154. [Google Scholar] [CrossRef]
- Imran, M.; Rauf, A.; Khan, I.A.; Shahbaz, M.; Qaisrani, T.B.; Fatmawati, S.; Abu-Izneid, T.; Imran, A.; Rahman, K.U.; Gondal, T.A. Thymoquinone: A novel strategy to combat cancer: A review. Biomed. Pharmacother. 2018, 106, 390–402. [Google Scholar] [CrossRef]
- Peng, B.; He, R.; Xu, Q.; Yang, Y.; Hu, Q.; Hou, H.; Liu, X.; Li, J. Ginsenoside 20 (S)-protopanaxadiol inhibits triple-negative breast cancer metastasis in vivo by targeting EGFR-mediated MAPK pathway. Pharmacol. Res. 2019, 142, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Xue, L. Kaempferol suppresses proliferation and induces cell cycle arrest, apoptosis, and DNA damage in breast cancer cells. Oncol. Res. 2019, 27, 629. [Google Scholar] [CrossRef] [PubMed]
- Heiden, M.G.V. Targeting cancer metabolism: A therapeutic window opens. Nat. Rev. Drug Discov. 2011, 10, 671–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azevedo, C.; Correia-Branco, A.; Araújo, J.R.; Guimaraes, J.T.; Keating, E.; Martel, F. The chemopreventive effect of the dietary compound kaempferol on the MCF-7 human breast cancer cell line is dependent on inhibition of glucose cellular uptake. Nutr. Cancer 2015, 67, 504–513. [Google Scholar] [CrossRef] [PubMed]
- Hung, H. Inhibition of estrogen receptor alpha expression and function in MCF-7 cells by kaempferol. J. Cell. Physiol. 2004, 198, 197–208. [Google Scholar] [CrossRef]
- Xu, Y.; Shu, B.; Tian, Y.; Wang, G.; Wang, Y.; Wang, J.; Dong, Y. Oleanolic acid induces osteosarcoma cell apoptosis by inhibition of Notch signaling. Mol. Carcinog. 2018, 57, 896–902. [Google Scholar] [CrossRef]
- Kiesel, V.A.; Stan, S.D. Modulation of Notch Signaling Pathway by Bioactive Dietary Agents. Int. J. Mol. Sci. 2022, 23, 3532. [Google Scholar] [CrossRef]
- Li, X.; Zhang, C.-T.; Ma, W.; Xie, X.; Huang, Q. Oridonin: A Review of Its Pharmacology, Pharmacokinetics and Toxicity. Front. Pharmacol. 2021, 12, 1642. [Google Scholar] [CrossRef]
- Xia, S.; Zhang, X.; Li, C.; Guan, H. Oridonin inhibits breast cancer growth and metastasis through blocking the Notch signaling. Saudi Pharm. J. 2017, 25, 638–643. [Google Scholar] [CrossRef]
- Huyen, C.T.T.; Luyen, B.T.T.; Khan, G.J.; Oanh, H.V.; Hung, T.M.; Li, H.-J.; Li, P. Chemical constituents from Cimicifuga dahurica and their anti-proliferative effects on MCF-7 breast cancer cells. Molecules 2018, 23, 1083. [Google Scholar] [CrossRef] [Green Version]
- Einbond, L.S.; Wen-Cai, Y.; He, K.; Wu, H.-A.; Cruz, E.; Roller, M.; Kronenberg, F. Growth inhibitory activity of extracts and compounds from Cimicifuga species on human breast cancer cells. Phytomedicine 2008, 15, 504–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rickels, R.; Shilatifard, A. Enhancer logic and mechanics in development and disease. Trends Cell Biol. 2018, 28, 608–630. [Google Scholar] [CrossRef] [PubMed]
- Vassilev, L.T. MDM2 inhibitors for cancer therapy. Trends Mol. Med. 2007, 13, 23–31. [Google Scholar] [CrossRef]
- Tan, B.X.; Khoo, K.H.; Lim, T.M.; Lane, D.P. High Mdm4 levels suppress p53 activity and enhance its half-life in acute myeloid leukaemia. Oncotarget 2014, 5, 933–943. [Google Scholar] [CrossRef] [Green Version]
- Qin, J.-J.; Sarkar, S.; Voruganti, S.; Agarwal, R.; Wang, W.; Zhang, R. Identification of lineariifolianoid A as a novel dual NFAT1 and MDM2 inhibitor for human cancer therapy. J. Biomed. Res. 2016, 30, 322–333. [Google Scholar]
- Zeng, A.; Liang, X.; Zhu, S.; Liu, C.; Luo, X.; Zhang, Q.; Song, L. Baicalin, a potent inhibitor of NF-κB signaling pathway, enhances chemosensitivity of breast cancer cells to docetaxel and inhibits tumor growth and metastasis both in vitro and in vivo. Front. Pharmacol. 2020, 11, 879. [Google Scholar] [CrossRef]
- Jin, Y.; Huynh, D.T.N.; Nguyen, T.L.L.; Jeon, H.; Heo, K.-S. Therapeutic effects of ginsenosides on breast cancer growth and metastasis. Arch. Pharmacal Res. 2020, 43, 773–787. [Google Scholar] [CrossRef]
- Qin, J.-J.; Jin, H.-Z.; Huang, Y.; Zhang, S.-D.; Shan, L.; Voruganti, S.; Nag, S.; Wang, W.; Zhang, W.-D.; Zhang, R. Selective cytotoxicity, inhibition of cell cycle progression, and induction of apoptosis in human breast cancer cells by sesquiterpenoids from Inula lineariifolia Turcz. Eur. J. Med. Chem. 2013, 68, 473–481. [Google Scholar] [CrossRef] [Green Version]
- Qin, J.-J.; Huang, Y.; Wang, D.; Cheng, X.-R.; Zeng, Q.; Zhang, S.-D.; Hu, Z.-L.; Jin, H.-Z.; Zhang, W.-D. Lineariifolianoids A–D, rare unsymmetrical sesquiterpenoid dimers comprised of xanthane and guaiane framework units from Inula lineariifolia. RSC Adv. 2012, 2, 1307–1309. [Google Scholar] [CrossRef]
- Wang, W.; Zafar, A.; Rajaei, M.; Zhang, R. Two birds with one stone: NFAT1-MDM2 dual inhibitors for cancer therapy. Cells 2020, 9, 1176. [Google Scholar] [CrossRef]
- Lee, C.; Ryu, H.W.; Kim, S.; Kim, M.; Oh, S.-R.; Ahn, K.-S.; Park, J. Verminoside from Pseudolysimachion rotundum var. subintegrum sensitizes cisplatin-resistant cancer cells and suppresses metastatic growth of human breast cancer. Sci. Rep. 2020, 10, 20337. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zou, T.; Wang, S.; Chen, H.; Su, D.; Fu, X.; Zhang, Q.; Kang, X. Genistein-induced differentiation of breast cancer stem/progenitor cells through a paracrine mechanism. Int. J. Oncol. 2016, 48, 1063–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarkowski, M.; Kokocińska, M.; Latocha, M. Genistein in chemoprevention and treatment. Pol. Merkur. Lek. Organ Pol. Tow. Lek. 2013, 34, 54–57. [Google Scholar]
- Wang, K.; Zhang, C.; Bao, J.; Jia, X.; Liang, Y.; Wang, X.; Chen, M.; Su, H.; Li, P.; Wan, J.-B. Synergistic chemopreventive effects of curcumin and berberine on human breast cancer cells through induction of apoptosis and autophagic cell death. Sci. Rep. 2016, 6, 26064. [Google Scholar] [CrossRef] [Green Version]
- Kashyap, A.; Umar, S.M.; Dev, A., Jr.; Mendiratta, M.; Prasad, C.P. In vitro anticancer efficacy of a polyphenolic combination of Quercetin, Curcumin, and Berberine in triple negative breast cancer (TNBC) cells. Phytomed. Plus 2022, 2, 100265. [Google Scholar] [CrossRef]
- Shehatta, N.H.; Okda, T.M.; Omran, G.A.; Abd-Alhaseeb, M.M. Baicalin; a promising chemopreventive agent, enhances the antitumor effect of 5-FU against breast cancer and inhibits tumor growth and angiogenesis in Ehrlich solid tumor. Biomed. Pharmacother. 2022, 146, 112599. [Google Scholar] [CrossRef]
- Rai, G.; Suman, S.; Mishra, S.; Shukla, Y. Evaluation of growth inhibitory response of Resveratrol and Salinomycin combinations against triple negative breast cancer cells. Biomed. Pharmacother. 2017, 89, 1142–1151. [Google Scholar] [CrossRef]
- Chimento, A.; Santarsiero, A.; Iacopetta, D.; Ceramella, J.; De Luca, A.; Infantino, V.; Parisi, O.I.; Avena, P.; Bonomo, M.G.; Saturnino, C. A phenylacetamide resveratrol derivative exerts inhibitory effects on breast cancer cell growth. Int. J. Mol. Sci. 2021, 22, 5255. [Google Scholar] [CrossRef]
- Huang, J.; Li, H.; Ren, G. Epithelial-mesenchymal transition and drug resistance in breast cancer. Int. J. Oncol. 2015, 47, 840–848. [Google Scholar] [CrossRef] [Green Version]
- Linn, S.C.; Pinedo, H.M.; van Ark-Otte, J.; van Der Valk, P.; Hoekman, K.; Honkoop, A.H.; Vermorken, J.B.; Giaccone, G. Expression of drug resistance proteins in breast cancer, in relation to chemotherapy. Int. J. Cancer 1997, 71, 787–795. [Google Scholar] [CrossRef]
- Tang, Y.; Wang, Y.; Kiani, M.F.; Wang, B. Classification, treatment strategy, and associated drug resistance in breast cancer. Clin. Breast Cancer 2016, 16, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Noel, B.; Singh, S.K.; Lillard, J.W., Jr.; Singh, R. Role of natural compounds in preventing and treating breast cancer. Front. Biosci. 2020, 12, 137–160. [Google Scholar]
- Ghosh, S.; Banerjee, S.; Sil, P.C. The beneficial role of curcumin on inflammation, diabetes and neurodegenerative disease: A recent update. Food Chem. Toxicol. 2015, 83, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Chopra, K.; Mukherjee, M.; Pottabathini, R.; Dhull, D.K. Current knowledge and pharmacological profile of berberine: An update. Eur. J. Pharmacol. 2015, 761, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Park, W.; Amin, A.R.; Chen, Z.G.; Shin, D.M. New perspectives of curcumin in cancer prevention. Cancer Prev. Res. 2013, 6, 387–400. [Google Scholar] [CrossRef] [Green Version]
- Kaboli, P.J.; Rahmat, A.; Ismail, P.; Ling, K.-H. Targets and mechanisms of berberine, a natural drug with potential to treat cancer with special focus on breast cancer. Eur. J. Pharmacol. 2014, 740, 584–595. [Google Scholar] [CrossRef]
- Batool, A.; Hazafa, A.; Ahmad, S.; Khan, H.A.; Abideen, H.M.; Zafar, A.; Bilal, M.; Iqbal, H.M. Treatment of lymphomas via regulating the Signal transduction pathways by natural therapeutic approaches: A review. Leuk. Res. 2021, 104, 106554. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Harikumar, K.B. Potential therapeutic effects of curcumin, the anti-inflammatory agent, against neurodegenerative, cardiovascular, pulmonary, metabolic, autoimmune and neoplastic diseases. Int. J. Biochem. Cell Biol. 2009, 41, 40–59. [Google Scholar] [CrossRef] [Green Version]
- Priyadarsini, K.I. The chemistry of curcumin: From extraction to therapeutic agent. Molecules 2014, 19, 20091–20112. [Google Scholar] [CrossRef] [Green Version]
- Czarny, P.; Pawlowska, E.; Bialkowska-Warzecha, J.; Kaarniranta, K.; Blasiak, J. Autophagy in DNA damage response. Int. J. Mol. Sci. 2015, 16, 2641–2662. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Yu, Z.; Li, Y.; Fichna, J.; Storr, M. Effects of berberine in the gastrointestinal tract—A review of actions and therapeutic implications. Am. J. Chin. Med. 2014, 42, 1053–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umar, S.M.; Patra, S.; Kashyap, A.; Dev, A., Jr.; Kumar, L.; Prasad, C.P. Quercetin Impairs HuR-Driven Progression and Migration of Triple Negative Breast Cancer (TNBC) Cells. Nutr. Cancer 2021, 74, 1497–1510. [Google Scholar] [CrossRef] [PubMed]
- Qian, K.; Tang, C.-Y.; Chen, L.-Y.; Zheng, S.; Zhao, Y.; Ma, L.-S.; Xu, L.; Fan, L.-H.; Yu, J.-D.; Tan, H.-S. Berberine reverses breast cancer multidrug resistance based on fluorescence pharmacokinetics in vitro and in vivo. ACS Omega 2021, 6, 10645–10654. [Google Scholar] [CrossRef] [PubMed]
- George, B.P.; Chandran, R.; Abrahamse, H. Role of phytochemicals in cancer chemoprevention: Insights. Antioxidants 2021, 10, 1455. [Google Scholar] [CrossRef]
- Li-Weber, M. New therapeutic aspects of flavones: The anticancer properties of Scutellaria and its main active constituents Wogonin, Baicalein and Baicalin. Cancer Treat. Rev. 2009, 35, 57–68. [Google Scholar] [CrossRef]
- Liu, D.-K.; Dong, H.-F.; Liu, R.-F.; Xiao, X.-L. Baicalin inhibits the TGF-β1/p-Smad3 pathway to suppress epithelial-mesenchymal transition-induced metastasis in breast cancer. Oncotarget 2020, 11, 2863–2872. [Google Scholar] [CrossRef]
- Talukdar, D.; Chaudhuri, B.; Ray, M.; Ray, S. Critical evaluation of toxic versus beneficial effects of methylglyoxal. Biochemistry 2009, 74, 1059–1069. [Google Scholar] [CrossRef]
- Mele, L.; Paino, F.; Papaccio, F.; Regad, T.; Boocock, D.; Stiuso, P.; Lombardi, A.; Liccardo, D.; Aquino, G.; Barbieri, A. A new inhibitor of glucose-6-phosphate dehydrogenase blocks pentose phosphate pathway and suppresses malignant proliferation and metastasis in vivo. Cell Death Dis. 2018, 9, 572. [Google Scholar] [CrossRef] [Green Version]
- El-Far, A.H.; Tantawy, M.A.; Al Jaouni, S.K.; Mousa, S.A. Thymoquinone-chemotherapeutic combinations: New regimen to combat cancer and cancer stem cells. Naunyn-Schmiedeberg Arch. Pharmacol. 2020, 393, 1581–1598. [Google Scholar] [CrossRef]
- Muthusamy, T.; Yadav, L.; Ramalingam, S.; Ramachandran, I. Synergistic effect of 5-fluorouracil combined with naringin in MDA-MB-231 human breast cancer cells. Int. Res. J. Oncol. 2020, 3, 13–27. [Google Scholar]
- Botticelli, A.; Scagnoli, S.; Roberto, M.; Lionetto, L.; Cerbelli, B.; Simmaco, M.; Marchetti, P. 5-Fluorouracil degradation rate as a predictive biomarker of toxicity in breast cancer patients treated with capecitabine. J. Oncol. Pharm. Pract. 2020, 26, 1836–1842. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Ishfaq, M.; Liu, Y.; Wu, Z.; Wang, J.; Li, R.; Qian, F.; Ding, L.; Li, J. Baicalin attenuates endometritis in a rabbit model induced by infection with Escherichia coli and Staphylococcus aureus via NF-κB and JNK signaling pathways. Domest. Anim. Endocrinol. 2021, 74, 106508. [Google Scholar] [CrossRef] [PubMed]
- Hazafa, A.; Iqbal, M.; Javaid, U.; Tareen, M.; Amna, D.; Ramzan, A.; Piracha, S.; Naeem, M. Inhibitory effect of polyphenols (phenolic acids, lignans, and stilbenes) on cancer by regulating signal transduction pathways: A review. Clin. Transl. Oncol. 2021, 24, 432–445. [Google Scholar] [CrossRef] [PubMed]
- Gomez, L.S.; Zancan, P.; Marcondes, M.C.; Ramos-Santos, L.; Meyer-Fernandes, J.R.; Sola-Penna, M.; Da Silva, D. Resveratrol decreases breast cancer cell viability and glucose metabolism by inhibiting 6-phosphofructo-1-kinase. Biochimie 2013, 95, 1336–1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Zhang, H.; Zhu, Y.; Wu, Z.; Cui, C.; Cai, F. Anticancer mechanisms of salinomycin in breast cancer and its clinical applications. Front. Oncol. 2021, 11, 654428. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, M.; Patro, B.S. Salinomycin reduces growth, proliferation and metastasis of cisplatin resistant breast cancer cells via NF-kB deregulation. Toxicol. Vitr. 2019, 60, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Dewangan, J.; Tandon, D.; Srivastava, S.; Verma, A.K.; Yapuri, A.; Rath, S.K. Novel combination of salinomycin and resveratrol synergistically enhances the anti-proliferative and pro-apoptotic effects on human breast cancer cells. Apoptosis 2017, 22, 1246–1259. [Google Scholar] [CrossRef]
- Aung, T.N.; Qu, Z.; Kortschak, R.D.; Adelson, D.L. Understanding the effectiveness of natural compound mixtures in cancer through their molecular mode of action. Int. J. Mol. Sci. 2017, 18, 656. [Google Scholar] [CrossRef]
- Saini, M.; Sangwan, R.; Khan, M.F.; Kumar, A.; Verma, R.; Jain, S. Specioside (SS) & verminoside (VS)(Iridoid glycosides): Isolation, characterization and comparable quantum chemical studies using density functional theory (DFT). Heliyon 2019, 5, e01118. [Google Scholar]
- Singh, I.P.; Sidana, J.; Bharate, S.B.; Foley, W.J. Phloroglucinol compounds of natural origin: Synthetic aspects. Nat. Prod. Rep. 2010, 27, 393–416. [Google Scholar] [CrossRef]
- Larayetan, R.; Ololade, Z.S.; Ogunmola, O.O.; Ladokun, A. Phytochemical constituents, antioxidant, cytotoxicity, antimicrobial, antitrypanosomal, and antimalarial potentials of the crude extracts of Callistemon citrinus. Evid. Based Complement. Altern. Med. 2019, 2019, 5410923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, R.K.; Suh, Y.; Yoo, K.C.; Cui, Y.H.; Hwang, E.; Kim, H.J.; Kang, J.S.; Kim, M.J.; Lee, Y.Y.; Lee, S.J. Phloroglucinol suppresses metastatic ability of breast cancer cells by inhibition of epithelial-mesenchymal cell transition. Cancer Sci. 2015, 106, 94–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidic, D.; Maksimovic, M.; Cavar, S.; Solic, M.E. Comparison of essential oil prof iles of Satureja montana l. and Endemic Satureja visianii Šilic. J. Essent. Oil Bear. Plants 2009, 12, 273–281. [Google Scholar] [CrossRef]
- Esmaeili, A.; Rustaiyan, A.; Masoudi, S.; Nadji, K. Composition of the essential oils of Mentha aquatica L. and Nepeta meyeri Benth. from Iran. J. Essent. Oil Res. 2006, 18, 263–265. [Google Scholar] [CrossRef]
- Medjahed, F.; Merouane, A.; Saadi, A.; Bader, A.; Cioni, P.L.; Flamini, G. Chemical profile and antifungal potential of essential oils from leaves and flowers of Salvia algeriensis (Desf.): A comparative study. Chil. J. Agric. Res. 2016, 76, 195–200. [Google Scholar] [CrossRef] [Green Version]
- Loizzo, M.R.; Menichini, F.; Tundis, R.; Bonesi, M.; Nadjafi, F.; Saab, A.M.; Frega, N.G.; Menichini, F. Comparative chemical composition and antiproliferative activity of aerial parts of Salvia leriifolia Benth. and Salvia acetabulosa L. essential oils against human tumor cell in vitro models. J. Med. Food 2010, 13, 62–69. [Google Scholar] [CrossRef]
- Ghantous, A.; Gali-Muhtasib, H.; Vuorela, H.; Saliba, N.A.; Darwiche, N. What made sesquiterpene lactones reach cancer clinical trials? Drug Discov. Today 2010, 15, 668–678. [Google Scholar] [CrossRef]
- Memariani, T.; Hosseini, T.; Kamali, H.; Mohammadi, A.; Ghorbani, M.; Shakeri, A.; Spandidos, D.A.; Tsatsakis, A.M.; Shahsavand, S. Evaluation of the cytotoxic effects of Cyperus longus extract, fractions and its essential oil on the PC3 and MCF7 cancer cell lines. Oncol. Lett. 2016, 11, 1353–1360. [Google Scholar] [CrossRef] [Green Version]
- Furtado, F.B.; Borges, B.C.; Teixeira, T.L.; Garces, H.G.; de Almeida Junior, L.D.; Alves, F.C.B.; da Silva, C.V.; Junior, A.F. Chemical composition and bioactivity of essential oil from Blepharocalyx salicifolius. Int. J. Mol. Sci. 2018, 19, 33. [Google Scholar] [CrossRef] [Green Version]
- Solárová, Z.; Liskova, A.; Samec, M.; Kubatka, P.; Büsselberg, D.; Solár, P. Anticancer potential of lichens’ secondary metabolites. Biomolecules 2020, 10, 87. [Google Scholar] [CrossRef] [Green Version]
- Sahin, E.; Dabagoglu Psav, S.; Avan, I.; Candan, M.; Sahinturk, V.; Koparal, A.T. Vulpinic acid, a lichen metabolite, emerges as a potential drug candidate in the therapy of oxidative stress-related diseases, such as atherosclerosis. Hum. Exp. Toxicol. 2019, 38, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Kılıç, N.; Derici, M.K.; Buyuk, I.; Aydın, S.; Aras, S.; Duman, D. Evaluation of in vitro anticancer activity of vulpinic acid and its apoptotic potential using gene expression and protein analysis. Indian J. Pharm. Educ. Res. 2018, 52, 46–54. [Google Scholar] [CrossRef] [Green Version]
- Kılıç, N.; Aras, S.; Cansaran-Duman, D. Determination of vulpinic acid effect on apoptosis and mRNA expression levels in breast cancer cell lines. Anti-Cancer Agents Med. Chem. 2018, 18, 2032–2041. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yang, B.; Zhou, K.; Li, H.; Li, D.; Gao, H.; Zhang, T.; Wei, D.; Li, Z.; Diao, Y. Potential therapeutic mechanism of genistein in breast cancer involves inhibition of cell cycle regulation. Mol. Med. Rep. 2015, 11, 1820–1826. [Google Scholar] [CrossRef] [Green Version]
- Nichols, J.A.; Katiyar, S.K. Skin photoprotection by natural polyphenols: Anti-inflammatory, antioxidant and DNA repair mechanisms. Arch. Dermatol. Res. 2010, 302, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Xie, Q.; Bai, Q.; Zou, L.Y.; Zhang, Q.Y.; Zhou, Y.; Chang, H.; Yi, L.; Zhu, J.D.; Mi, M.T. Genistein inhibits DNA methylation and increases expression of tumor suppressor genes in human breast cancer cells. Genes Chromosom. Cancer 2014, 53, 422–431. [Google Scholar] [CrossRef]
- Zhu, W.; Qin, W.; Zhang, K.; Rottinghaus, G.E.; Chen, Y.-C.; Kliethermes, B.; Sauter, E.R. Trans-resveratrol alters mammary promoter hypermethylation in women at increased risk for breast cancer. Nutr. Cancer 2012, 64, 393–400. [Google Scholar] [CrossRef] [Green Version]
- Ávila-Gálvez, M.Á.; Giménez-Bastida, J.A.; Espín, J.C.; González-Sarrías, A. Dietary phenolics against breast cancer. A critical evidence-based review and future perspectives. Int. J. Mol. Sci. 2020, 21, 5718. [Google Scholar] [CrossRef]
- Beatty, E.R.; O’Reilly, J.D.; England, T.G.; McAnlis, G.T.; Young, I.S.; Halliwell, B.; Geissler, C.A.; Sanders, T.A.; Wiseman, H. Effect of dietary quercetin on oxidative DNA damage in healthy human subjects. Br. J. Nutr. 2000, 84, 919–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, M.R.; Islam, F.; Nafady, M.H.; Akter, M.; Mitra, S.; Das, R.; Urmee, H.; Shohag, S.; Akter, A.; Chidambaram, K. Natural small molecules in breast cancer treatment: Understandings from a therapeutic viewpoint. Molecules 2022, 27, 2165. [Google Scholar] [CrossRef]
- Li, J.; Kataoka, K. Chemo-physical strategies to advance the in vivo functionality of targeted nanomedicine: The next generation. J. Am. Chem. Soc. 2020, 143, 538–559. [Google Scholar] [CrossRef] [PubMed]
- Burguin, A.; Diorio, C.; Durocher, F. Breast Cancer Treatments: Updates and New Challenges. J. Pers. Med. 2021, 11, 808. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S.; Dash, R. Natural products for the management and prevention of breast cancer. Evid. Based Complement. Altern. Med. 2018, 2018, 8324696. [Google Scholar] [CrossRef] [PubMed]
- Buhrmann, C.; Shayan, P.; Brockmueller, A.; Shakibaei, M. Resveratrol suppresses cross-talk between colorectal cancer cells and stromal cells in multicellular tumor microenvironment: A bridge between in vitro and in vivo tumor microenvironment study. Molecules 2020, 25, 4292. [Google Scholar] [CrossRef] [PubMed]
- Buhrmann, C.; Popper, B.; Kunnumakkara, A.B.; Aggarwal, B.B.; Shakibaei, M. Evidence that Calebin A, a component of curcuma longa suppresses NF-κB mediated proliferation, invasion and metastasis of human colorectal cancer induced by TNF-β (Lymphotoxin). Nutrients 2019, 11, 2904. [Google Scholar] [CrossRef] [Green Version]
- Shakibaei, M.; Kraehe, P.; Popper, B.; Shayan, P.; Goel, A.; Buhrmann, C. Curcumin potentiates antitumor activity of 5-fluorouracil in a 3D alginate tumor microenvironment of colorectal cancer. BMC Cancer 2015, 15, 250. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Wang, H.Q.; Zhang, H.L.; Kong, D.; Wu, X.Z. Ginsenoside Rh2 reverses P-glycoprotein-mediated multidrug resistance of MCF-7/ADM cells. Tumor 2007, 27, 365–369. [Google Scholar]
- Li, X.; Chu, S.; Lin, M.; Gao, Y.; Liu, Y.; Yang, S.; Zhou, X.; Zhang, Y.; Hu, Y.; Wang, H. Anticancer property of ginsenoside Rh2 from ginseng. Eur. J. Med. Chem. 2020, 203, 112627. [Google Scholar] [CrossRef]
- Wang, P.; Yang, H.L.; Yang, Y.J.; Wang, L.; Lee, S.C. Overcome cancer cell drug resistance using natural products. Evid. Based Complement. Altern. Med. 2015, 2015, 767136. [Google Scholar] [CrossRef] [Green Version]
- Wang, S. The multidrug-resistant mechanism study of reverse effect of Baicalin for hepatocellular carcinoma cell line (Bel 7402/ADM). Chin. J. Mod. Drug Appl. 2012, 6, 10–11. [Google Scholar]
- Tang, X.; Tang, Y. Efficiency and mechanisms of peimine reversing multi-drug resistance of A549/DDP cell line. Shandong Med. J. 2012, 52, 4–6. [Google Scholar]
- Tang, S.-M.; Deng, X.-T.; Zhou, J.; Li, Q.-P.; Ge, X.-X.; Miao, L. Pharmacological basis and new insights of quercetin action in respect to its anti-cancer effects. Biomed. Pharmacother. 2020, 121, 109604. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Liu, X.; Chang, Y.; Wang, R.; Lv, T.; Cui, C.; Liu, M. Investigation of the anti-inflammatory and antioxidant activities of luteolin, kaempferol, apigenin and quercetin. South Afr. J. Bot. 2021, 137, 257–264. [Google Scholar] [CrossRef]
- Wu, Y.; Zeng, Y.; Liang, S. The exploratory research on the berberine reverse the HCT-8/VCR multidrug resistance and its relationship with P-gp functional variance. Guide China Med. 2010, 8, 18–20. [Google Scholar]
- Warowicka, A.; Nawrot, R.; Goździcka-Józefiak, A. Antiviral activity of berberine. Arch. Virol. 2020, 165, 1935–1945. [Google Scholar] [CrossRef]
- Yan, L.; Zizheng, W.; Tengfei, Y. In vitro study on the reversal of multidrug resistance (MDR) in HL60/VCR cell line with ginsenoside-Rb1. J. Radioimmunol. 2005, 18, 362–365. [Google Scholar]
- Gao, H.; Kang, N.; Hu, C.; Zhang, Z.; Xu, Q.; Liu, Y.; Yang, S. Ginsenoside Rb1 exerts anti-inflammatory effects in vitro and in vivo by modulating toll-like receptor 4 dimerization and NF-kB/MAPKs signaling pathways. Phytomedicine 2020, 69, 153197. [Google Scholar] [CrossRef]
- Qin, Y.; Fu, Z.; Lu, W.; Li, L.; Tang, W. Experimental study of multidrug-resistance reversing of curcumin on human colon carcinoma HCT-8/VCR cells. Chin. J. Biochem. Pharm. 2011, 32, 172–173. [Google Scholar]
- Kabir, M.; Rahman, M.; Akter, R.; Behl, T.; Kaushik, D.; Mittal, V.; Pandey, P.; Akhtar, M.F.; Saleem, A.; Albadrani, G.M. Potential role of curcumin and its nanoformulations to treat various types of cancers. Biomolecules 2021, 11, 392. [Google Scholar] [CrossRef]
- Guo, J.; Pan, X.; Feng, C.; Zhou, J. Study on reversal effects of Oridonin on multidrug resistant cell line K562/A02. Shanghai Med. J. 2002, 25, 43–45. [Google Scholar]
- Li, J.; Wu, Y.; Wang, D.; Zou, L.; Fu, C.; Zhang, J.; Leung, G.P.-H. Oridonin synergistically enhances the anti-tumor efficacy of doxorubicin against aggressive breast cancer via pro-apoptotic and anti-angiogenic effects. Pharmacol. Res. 2019, 146, 104313. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, S.; Wang, J.; Jiang, W.; Zhang, Y.; Liu, Y.; Wang, Z. Reversal effect of ginsenoside Rg3 on cisplatin-resistant human lung adenocarcinoma cell line (A549DDP) and its mechanisms. Chin. J. Respir. Crit. Care Med. 2002, 1, 100–103. [Google Scholar]
- Holohan, C.; Van Schaeybroeck, S.; Longley, D.B.; Johnston, P.G. Cancer drug resistance: An evolving paradigm. Nat. Rev. Cancer 2013, 13, 714–726. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.-B.; Li, L.; Fu, J.; Mao, X.-P.; Xu, L.-Z. Reversion of multidrug resistance in a chemoresistant human breast cancer cell line by β-elemene. Pharmacology 2012, 89, 303–312. [Google Scholar] [CrossRef]
- Wang, W.; Lv, M.; Wang, Y.; Zhang, J. Development of novel application of 3,3′-diindolylmethane: Sensitizing multidrug resistance human breast cancer cells to γ-irradiation. Pharm. Biol. 2016, 54, 3164–3168. [Google Scholar] [CrossRef] [Green Version]
- Shindikar, A.; Singh, A.; Nobre, M.; Kirolikar, S. Curcumin and resveratrol as promising natural remedies with nanomedicine approach for the effective treatment of triple negative breast cancer. J. Oncol. 2016, 2016, 750785. [Google Scholar] [CrossRef] [Green Version]
- Shao, C.; Wu, J.; Han, S.; Liu, Y.; Su, Z.; Zhu, H.-L.; Liu, H.-K.; Qian, Y. Biotinylated curcumin as a novel chemosensitizer enhances naphthalimide-induced autophagic cell death in breast cancer cells. Eur. J. Med. Chem. 2022, 228, 114029. [Google Scholar] [CrossRef]
- Al Dhaheri, Y.; Attoub, S.; Ramadan, G.; Arafat, K.; Bajbouj, K.; Karuvantevida, N.; AbuQamar, S.; Eid, A.; Iratni, R. Carnosol induces ROS-mediated beclin1-independent autophagy and apoptosis in triple negative breast cancer. PLoS ONE 2014, 9, e109630. [Google Scholar] [CrossRef]
- Sukalingam, K.; Ganesan, K.; Das, S.; Thent, Z.C. An insight into the harmful effects of soy protein: A review. Clin. Ter. 2015, 166, 131–139. [Google Scholar]
- Ahmad, A.; Li, Y.; Sarkar, F.H. The bounty of nature for changing the cancer landscape. Mol. Nutr. Food Res. 2016, 60, 1251–1263. [Google Scholar] [CrossRef]
- Kim, D.; Park, M.; Haleem, I.; Lee, Y.; Koo, J.; Na, Y.C.; Song, G.; Lee, J. Natural product Ginsenoside 20 (S)-25-methoxyl-dammarane-3β, 12β, 20-triol in cancer treatment: A review of the pharmacological mechanisms and pharmacokinetics. Front. Pharmacol. 2020, 11, 521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Zhang, Y.; Guo, S.; Bai, F.; Wu, T.; Zhao, Y. Improved oral bioavailability of 20 (R)-25-methoxyl-dammarane-3β, 12β, 20-triol using nanoemulsion based on phospholipid complex: Design, characterization, and in vivo pharmacokinetics in rats. Drug Des. Dev. Ther. 2016, 10, 3707–3716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Yang, J.; Yang, Y.; Zhang, J.; Xu, Y.; Lu, X. The natural products and extracts: Anti-triple-negative breast cancer in vitro. Chem. Biodivers. 2021, 18, e2001047. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extracted Compound | Biochemical Structure | Biochemical Nature | Source | Study Type | BC Type | Animal Model | Key Finding | Mechanism of Action | Reference |
|---|---|---|---|---|---|---|---|---|---|
| VOA |  | Alkaloid | V. africana | In vitro | ER-positive, TNBC, and HER2-positive BC | --- | Downregulating the PI3K/Akt/mTOR. VOA showed its usefulness against MCF-7 and 4T1 cells with IC50 values (0.99, 1.48 μM). | VOA significantly inhibits the phosphorylated AKT and mTOR in BC cells and also decreases the expression of CDK2, cyclin A, E. It also induces apoptosis and cell death in MCF-7 and 4T1 cells by arresting the S phase of the cell cycle. | [34] |
| Lin A |  | Sesquiterpenoid | I. lineariifolia | In vitro | TNBC, and HER2-positive BC | --- | Lin A induced apoptosis at a higher concentration of 50% in BC cells (MCF7 and MDA-MB-231 with IC50 (4.5 ± 0.3, 7.8 ± 0.6). | Lin A arrests the cell cycle at the G2/M phase, and inhibits cell invasion and cell proliferation in BC cells. | [100,95] |
| Fisetin |  | Flavonol | Cucumber, apple, strawberry | In vitro and in vivo | ER-positive, TNBC, and HER2-positive BC | BALB/c mice | Fisetin induced apoptosis in MCF-7, 4T1, and MDA-MB-231 at 40 and 80 μM. | Fisetin acts as an inhibitor of PI3K/Akt/mTOR signaling and inhibits the proliferation and dysregulation of this signaling pathway. | [66] |
| WG |  | Flavone | S. baicalensis | In vitro and in vivo | ER-positive, TNBC, and HER2-positive BC | Chicken chorioallantoic membrane (CAM) | WG showed inhibitory effects on MCF-7 and MDA-MB-231 at 20 and 40 μM. | WG acts as an inhibitor of PI3K/Akt/mTOR signaling and shows inhibition in cell proliferation. | [70] |
| AP |  | Flavone | A. cepa, C. sinensis | In vitro | ER-positive, HER2-positive BC | --- | It influenced the NF-κB pathway by suppressing the VEGF through deactivating progesterone receptors in BC cells. | It inhibits cell proliferation and migrations by arresting the cell cycle at the G2/M phase. It also suppresses the cyclin A, B, and CDK1 which controls the G2/M phase. | [77] |
| Oridonin |  | Diterpenoid | R. rubescens | In vivo | --- | BALB/C athymic nude mice | It induced apoptosis and cell death in BC cells. | Notch 1-4 protein expression is lowered by oridonin therapy, which hinders cancer cell migration and invasion. | [89] |
| Genistein |  | Isoflavones | Soy-based foods | In vitro | ER-positive, TNBC | --- | Activation of NF-κB showed potential against MCF-7 at an IC50 value of 20 µM. | It inhibits the phosphorylation of IκBα in MCF-7/T47D/MDA-MB-231 cell lines, thus playing a significant role in the regulation of IκBα to the p50. | [73] |
| GLA |  | Terpenoid | G. glabra | In vitro | ER-positive, TNBC, and HER2-positive BC | --- | It showed anticancer activity against MDA-MB-231/BT549. | It inhibits invasion and cell proliferation, as well as promote the expression of E-cadherin. | [25] |
| ATG |  | Isoflavones | S. heteromalla | In vitro and in vivo | ER-positive, TNBC | BALB/cA-nu | It showed anticancer potentials in MDA-MB-231 cells at 200 μM. | Inhibiting the phosphorylation of MAPK/ERK in MDA-MB-231 cells. | [24] |
| PPD |  | Glycoside | P. notoginseng | In vitro and in vivo | TNBC, and HER2-positive BC | BALB/C nude mice | It showed maximum activity below 20 μM against MDA-MB-231. | PPD targets BC cell lines by suppressing the MAPK pathway through the deactivation of ERK1/2, p38, and JNK. | [81] |
| Kaempferol |  | Flavonols | Onions, lettuce | In vitro | ER-positive, TNBC | --- | The number of cancerous cells decreased from 85.2% to 50.32% in the G1 phase of the cell cycle. Kaempferol significantly inhibited the BC cells (BT474 and MDA-MB-231) by blocking the critical phases of cell cycles. | Inhibitory actions against different breast cell lines can inhibit the expression of genes involved in MAPK/ERK. This shows that binding with estradiol causes degradation of Erα. | [82] |
| Cimigenoside |  | Glycoside | C. dahurica | In vitro and in vivo | ER-positive, TNBC | BALB/C nude Crlj mice | Cimigenoside showed maximum anticancer activity against BC cell lines (MDA-MB-231, MCF-7) with IC50 (12.6 ± 1.47, 15.6 ± 2.47 μM). | Cimigenoside induces apoptosis in BC cells by arresting the G2/M phase of the cell cycle. An in vitro study of cimigenoside also inhibits/attenuates BC cell proliferation and invasion. An in vivo study inhibited the growth of tumor growth in mice models. | [23,90] |
| Ginsenosides |  | Glycosides | P. notoginseng | In vitro and in vivo | ER-positive, TNBC | Nu/nu mice | It showed maximum anticancer activity against BC cell lines (MDA-MB-231). | 25-OCH3-PPD is involved in arresting the G1 phase of the cell cycle and induces apoptosis in BC cells by downregulating MDM2. | [36] |
| BA |  | Flavonoid | Scutellaria baicalensis | In vitro and in vivo | ER-positive, HER2-positive BC | BALB/c mice | It showed suppression of the NF-κB pathway in the development of human breast epithelial cells (MCF10A). | Suppress the NF-κB signaling pathway, as well as IL-1β, Bcl-2, and VEGF. | [26,96] |
| VMS |  | Monoterpenoids | P. rotundum | In vitro and in vivo | ER-positive, TNBC, and HER2-positive BC | PyMT/FP635 mouse | It showed maximum activity against MDA-MB-231 and MCF7 cells with IC50 value (10 µM). | VMS suppresses the growth of epithelial lining and the transition of mesenchymal breast cells. | [101] |
| Calcitrinone A |  | Phloroglucinol | C. citrinus | In vivo and in vitro | ER-positive | Chick chorioallantoic membrane (CAM) | Calcitrinone A induced apoptosis and cell death in MDA-MB-231 cells. | Calcitrinone A interferes with mitochondrial function by blocking succinate coenzyme Q reductase and ultimately inhibits the complex II that increases the production of ROS. | [29] |
| Vulpinic acid |  | Butenolide | Lichens | In vitro | ER-positive, TNBC, and HER2-positive BC | --- | Vulpinic acid induced apoptosis in MCF-7. | Elevate the levels of FOXO-3 and Bax, and suppress the expression of Bcl-2 and procaspase-3/9 to enhance the activity of tumor suppressor miRNAs. | [28] |
| Genistein |  | Isoflavone | Fabaceae family | In vitro | TNBC, and HER2-positive BC | --- | It induced apoptosis and cell death in MCF-7 and MDA-MB-231 cells. It also inhibited cell proliferation and progression in BC. | Arresting the cell cycle at G2/M phase, downregulating CDK-1, and inhibiting the expression of Bcl-2 and the function of DNA polymerase II. | [102,103] |
| CUR + BBR |  | Diarylheptanoid, isoquinoline alkaloid | Curcuma longa, berberine from Rhizoma coptidis | In vitro | ER-positive, TNBC, and HER2-positive BC | --- | It showed effects against BC cell lines (MDA-MB-231 and MDA-MB-468) at p ≤ 0.010. | The EMT process in the case of BC is impaired. | [104,105] |
| BA + 5-FU |  | Flavonoid (BA) | Scutellaria baicalensis | In vivo | --- | Swiss albino mice | It showed inflammation by inhibiting the VEGF, IL-1β, and NF-κB. | Inflammation is inhibited by the VEGF, IL-1β, and NF-κB, which play significant roles in preventing BC. | [106] |
| MG + 5-FU |  | Polyphenolic (MG) | Coffee, wine | In vivo and in vitro | ER-positive, TNBC, and HER2-positive BC | BALB/c mice and Swiss albino mice | It showed anticancer activity against the BC cell line (MCF-7). | Arresting the cell cycle at MG G0/G1 phase induces apoptosis and cell death by increasing Bac-2 and caspase 9. | [30] |
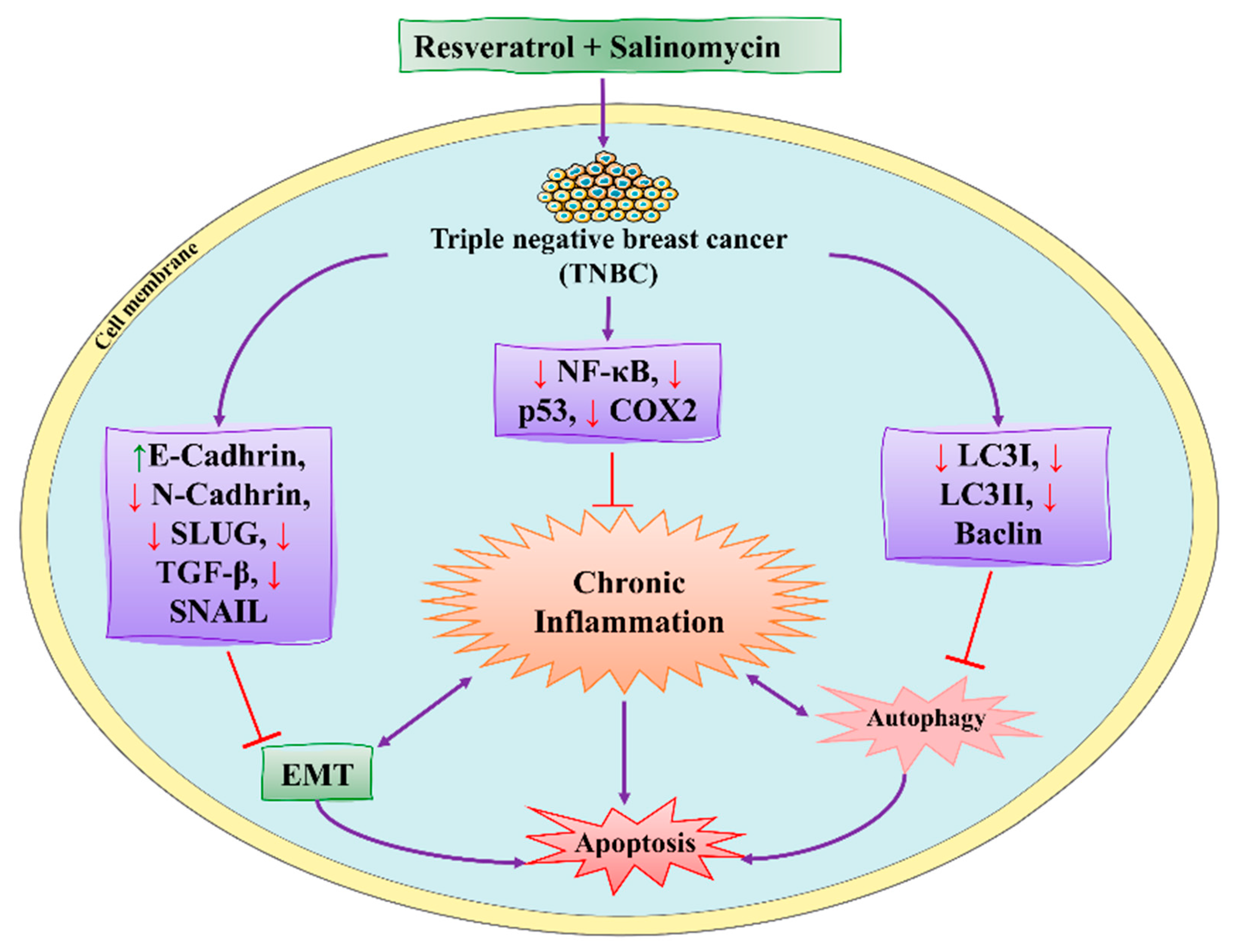
| RSVL + SAL |  | Polyphenol | Red grapes | In vivo and in vitro | ER-positive, TNBC, and HER2-positive BC | Swiss albino mice | It showed anticancer activity against MDA-MB-231. | Arresting the S1 phase of the cell cycle also induces apoptosis in BC cells. Other activities include inhibition and cell proliferation. It acts as an antioxidant by preventing the DNA dame and suppressing the tumor growth. SAL inhibits the epithelial mesenchymal transition, and suppresses p53, COX-2, and Beclin. | [107,108] |
| Natural Compound | Nature | NCT No. | Number of Participants | Disease Type | Dose/Concentration | Duration of Trial | Trial Phase | Study Location | Reference |
|---|---|---|---|---|---|---|---|---|---|
| Curcumin | Polyphenol | NCT01740323 | 30 | BC | 8 g | 8 weeks | Phase II | USA | www.clinicaltrials.gov (accessed on 27 April 2022) |
| Resveratrol | Stilbenoid | --- | 39 | Metastatic BC | 5 or 50 mg | 3 months | Phase I | USA | [157,158] |
| Berberine | Alkaloids | NCT03281096 | 1000 | Invasive BC, colorectal | 300 mg | 4 weeks | Phase II and III | China | www.clinicaltrials.gov (accessed on 27 April 2022) |
| Curcumin (iv) + Paclitaxel | Polyphenol | NCT03072992 | 75 | Metastatic BC | 300 mg | 12 weeks | Phase II | Armenia | www.clinicaltrials.gov (accessed on 27 April 2022) |
| Quercetin | Carotenoids | --- | 42 | Advanced BC | 200 mg | 2 weeks | Randomized crossover clinical trial | UK | [159,160] |
| Resveratrol | Stilbenoid | NCT04266353 | 50 | TNBC | 150 mg | 2–4 weeks | Suspended (due to COVID-19) | California | www.clinicaltrials.gov (accessed on 27 April 2022) |
| Curcumin | Polyphenol | NCT01975363 | 30 | BC (obese women) | 100 mg | 3 months | Pilot trial | USA | www.clinicaltrials.gov (accessed on 27 April 2022) |
| Genistein | Isoflavon | NCT00099008 | 30 | BC | 10 or 20 mg | 84 Days | Completed | US | www.clinicaltrials.gov (accessed on 27 April 2022) |
| Natural Compound | Nature | Dose/Concentration | Target | Cell Mode | Chemo Drug | IDS (x-Fold) | Reference |
|---|---|---|---|---|---|---|---|
| Ginsenosides | Glycosides | 40 µM | MCF | ADM | Doxorubicin | 29.2 | [167,168] |
| Baicalin | Flavonoid | 150 µg/mL | MDR1 and MRP1 | MCF7/ADR | Doxorubicin | 6.5 | [169,170] |
| Quercetin | Carotenoids | 50 µM | MDR1 and MRP1 | MCF7/ADM | Cisplatin | 3.5 | [171,172,173] |
| Berberine | Alkaloids | 20 µM | MDR1 | MDR1 | Vincristine | 3.2 | [174,175] |
| Ginsenoside Rb1 | Glycosides | 80 µM | MDR1 | MCF-7/ADR | Vincristine | 2.5 | [176,177] |
| Apigenin | Flavone | 13 µM | MDR1 | HCT | 5-FU | 4.9 | [169,173] |
| Curcumin | Alkaloids | 25 µM | MDR1 | Various | Various | 4.5 | [178,179] |
| Oridonin | Diterpenoid | 3 µM | MDR1 | MCF7/ADM | Various | 8.5 | [180,181] |
| Ginsenoside Rg3 | Glycosides | 30 µg/mL | MDR1 and MRP1 | MCF7/ADR | Various | 8.5 | [182] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naeem, M.; Iqbal, M.O.; Khan, H.; Ahmed, M.M.; Farooq, M.; Aadil, M.M.; Jamaludin, M.I.; Hazafa, A.; Tsai, W.-C. A Review of Twenty Years of Research on the Regulation of Signaling Pathways by Natural Products in Breast Cancer. Molecules 2022, 27, 3412. https://doi.org/10.3390/molecules27113412
Naeem M, Iqbal MO, Khan H, Ahmed MM, Farooq M, Aadil MM, Jamaludin MI, Hazafa A, Tsai W-C. A Review of Twenty Years of Research on the Regulation of Signaling Pathways by Natural Products in Breast Cancer. Molecules. 2022; 27(11):3412. https://doi.org/10.3390/molecules27113412
Chicago/Turabian StyleNaeem, Muhammad, Muhammad Omer Iqbal, Humaira Khan, Muhammad Masood Ahmed, Muhammad Farooq, Muhammad Moeen Aadil, Mohamad Ikhwan Jamaludin, Abu Hazafa, and Wan-Chi Tsai. 2022. "A Review of Twenty Years of Research on the Regulation of Signaling Pathways by Natural Products in Breast Cancer" Molecules 27, no. 11: 3412. https://doi.org/10.3390/molecules27113412
APA StyleNaeem, M., Iqbal, M. O., Khan, H., Ahmed, M. M., Farooq, M., Aadil, M. M., Jamaludin, M. I., Hazafa, A., & Tsai, W. -C. (2022). A Review of Twenty Years of Research on the Regulation of Signaling Pathways by Natural Products in Breast Cancer. Molecules, 27(11), 3412. https://doi.org/10.3390/molecules27113412